新規な N- 結合型
糖タンパク質糖鎖の合成研究
2005 年度
中野 淳
目 次
緒 論
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 1
本 論
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 9
第 1 章 凍結反応場を利用した効率的糖鎖合成法の開発 ・・・・・・・・・・・・・・・ 10 1.1 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 10 1.2 凍結反応の確立 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 12 1.2.1 溶媒の選定 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 12 1.2.2 グリコシルドナーとグリコシルアクセプターの選定 ・・・・・・・・・・・・・・・・・・ 12 1.2.3 凍結反応による反応加速 - 1 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 13 1.2.4 凍結反応による反応加速 - 2 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 14 1.3 凍結反応を用いた構築困難な糖鎖の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 16
第 2 章 寄生虫由来の多分岐型 N-型糖鎖の合成研究 ・・・・・・・・・・・・・・・・・・・・ 18 2.1 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 18
2.2 逆合成解析 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 21 2.3 フラグメント A の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 23 2.3.1 フラグメントA-1 の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 23 2.3.2 フラグメントA-2 の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 27 2.3.3 6 糖骨格の構築 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 28 2.3.4 官能基変換とフラグメント A の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 29 2.4 フラグメント B の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 33 2.4.1 α-フコシル化反応による2糖の合成
〜α1-3フコシド結合の構築〜 ・・・・・・・・・・・・・・・・・・・ 33 2.4.2 PMB 基の変換 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 35 2.4.3 α-フコシル化反応によるフラグメントBの合成
〜α1-6フコシド結合の構築〜 ・・・・・・・・・・・・・・・・・・・ 37
2.5 天然型 9 糖の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 39 2.5.1 9 糖骨格の構築 〜その 1〜 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 39 2.5.2 新規グリコシルアクセプターの設計と合成 〜その 1〜 ・・・・・・・・・・・・・・・ 40 2.5.3 新規グリコシルアクセプターの設計と合成 〜その 2〜 ・・・・・・・・・・・・・・・ 43 2.5.4 9 糖骨格の構築 〜その 2〜 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 45 2.6 非天然型糖鎖の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 48 2.6.1 フラグメント A を用いたフコース欠落型糖鎖の合成 ・・・・・・・・・・・・・・・・・・ 48 2.6.2 キシロース欠落型フラグメントAの合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 51 2.6.3 キシロース欠落型糖鎖の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 54
第 3 章 N-結合型糖アミノ酸の新規合成法の開発 ・・・・・・・・・・・・・・・・・・・・・・・・ 58 3.1 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 58 3.2 ヒドロキサム酸エステルを用いた N-結合型糖アミノ酸の合成 ・・・・・・・・・・・・・ 60 3.2.1 反応系の設定 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 60 3.2.2 ヒドロキサム酸エステルの合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 63 3.2.3 N-グリコシル化反応の検討 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 64 3.3 分子内N-グリコシル化反応を用いたN-結合型糖アミノ酸の合成 ・・・・・・・・・・ 67 3.3.1 反応系の設定 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 67 3.3.2 化合物の設計と反応の拡張性 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 68 3.3.3 反応基質の逆合成解析とその合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 71 3.3.4 分子内 N-グリコシル化反応の検討 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 73
3.4 N-(ベンジルオキシ)カルバミン酸エステルを用いた N-グリコシル化反応 ・・ 78
3.4.1 反応系の設定 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 78 3.4.2 N-(ベンジルオキシ)カルバミン酸エステルの合成 ・・・・・・・・・・・・・・・・・・・・・ 79 3.4.3 N-グリコシル化反応の検討 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 80
第 4 章 総 括 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 81
実験の部
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 87
実験に関する一般事項 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 88 第 1 章 凍結反応場を利用した効率的糖鎖合成法の開発 ・・・・・・・・・・・・・・・・ 89
1.1 凍結反応の確立 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 89
第2 章 寄生虫由来の多分岐型N-型糖鎖の合成研究 ・・・・・・・・・・・・・・・・・・・・・ 91 2.1 フラグメント A の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 91 2.2 フラグメント Bの合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 100 2.3 フラグメント Ba及びBbの合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 104 2.4 天然型 9 糖の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 109 2.5 非天然型糖鎖の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 114 2.5.1 フラグメントAを用いた非天然型糖鎖の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・ 114 2.5.2 フラグメントAa の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 119 2.5.3 フラグメントAaを用いた非天然型糖鎖の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・ 125
第 3 章 N-結合型糖アミノ酸の新規合成法の開発 ・・・・・・・・・・・・・・・・・・・・・・・・ 132 3.1 ヒドロキサム酸エステルを用いた N-結合型糖アミノ酸の合成 ・・・・・・・・・・・・・ 132 3.2 分子内 N-グリコシル化反応を用いた N-結合型糖アミノ酸の合成 ・・・・・・・・・・ 135 3.2.1 糖-アミノ酸複合体の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 135 3.2.2 分子内N-グリコシル化反応 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 140 3.2.3 N-結合型糖アミノ酸の合成 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 142 3.3 N-(ベンジルオキシ)カルバミン酸エステルを用いたN-グリコシル化反応 ・・ 143 3.3.1 N-(ベンジルオキシ)カルバミン酸エステルの合成 ・・・・・・・・・・・・・・・・・・・・・ 143 3.3.2 N-グリコシル化反応 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 144
参考文献
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 147謝 辞
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 153
本論文に用いた略号
Ac Acetyl
All Allyl
Alloc Allyloxycarbonyl Asn Asparagine Asp Aspartic acid
Bn Benzyl
Boc tert-Butoxycarbonyl
Bu Butyl
Bz Benzoyl
Cbz Benzyloxycarbonyl CDI 1,1'-Carbonyldiimidazole Cp Cycropentadienyl
CPME Cycropentyl methyl ether CSA Camphor-10-sulfonic acid DAST (Diethylamino)sulfur tirifluoride DBU 1,8-Diazabicyclo[5.4.0]undec-7-ene DDQ 2,3-Dichloro-5,6-dicyano-1,4-benzoquinone DDQH 2,3-Dichloro-5,6-dicyanohydroquinone DIEA N,N-Diisopropylethylamine
DMAP 4-Dimetylaminopyridine DMF N,N-Dimethylformamide
DMSO Dimethyl sulfoxide
DMTST Dimethyl(methylthio)sulfonium trifluoromethanesulfonate DTBMP 2,6-Di-tert-butyl-4-metylpyridine
Fmoc 9-Fluorenylmethoxycarbonyl Fuc L-Fucose
Gal D-Galactose
GalNAc N-Acetyl-D-galactosamine Glc D-Glucose
GlcNAc N-Acetyl-D-glucosamine M Moles per liter Man D-Mannose
Me Methyl
MS Molecular sieves
NMR Nuclear magnetic resonance NeuAc N-Acetylneuraminic acid (sialic acid) NIS N-Iodosuccinimide
NPhth Phthalimide
Ph Phenyl
PMB p-Methoxybenzyl Pro Proline
pTLC Preparative TLC Py Pyridine
r.t. Room temperature
Ser Serine
TBAB Tetrabutylammonium bromide TBAF Tetrabutylammonium fluoride TBDMS tert-Butyldimethylsilyl
TBDPS tert-Butyldiphenylsilyl
TBTU O-Benzotriazol-1-yl-N,N,N’,N’-tetramethyluronium tetrafluoroborate Tf Trifluoromethanesulfonyl
TFA Trifluoroacetic acid
THF Tetrahydrofuran Thr Threonine
TLC Thin layer chromatography TMS Trimethylsilyl
Tr Trityl (Triphenylmethyl) Troc 2,2,2-Tirchloroethoxylcarbonyl Trp Tryptophan
Ts Tosyl (p-Toluenesulfonyl) Xyl D-Xylose
緒 論
真核生物が産出するタンパク質の多くは、翻訳後修飾として糖鎖が付加された糖タン パク質であり、動物細胞においては合成されたタンパク質の約 50%が糖鎖付加を受ける と推定されている1)。タンパク質は糖鎖の付加により、しばしばその性質が大きく変化 し、例えば立体構造、フォールディング、安定性、酵素やホルモンとしての活性に影響を 与える例が知られている2)。また、糖タンパク質の多くは細胞表層に存在しており、細胞 間、細胞−マトリックス間の認識、接着現象に大きな役割を果たしている3)。
糖タンパク糖鎖は著しい構造多様性を持っているが、その多くはタンパク質に対する 糖鎖付加の様式により N-結合型(アスパラギン結合型)、O-結合型(セリン/スレオニン結合 型)の二つの型に分類される(Figure 0-1)。N-結合型は、アスパラギン側鎖のカルボキサミ ドの窒素原子を介したC-N結合で、GlcNAcを還元末端とする糖鎖が付加し、O-結合型は Serまたは Thrのヒドロキシル基に GalNAc が C-O結合で結合している。この他、Trp 残 基のインドール環の C-2 炭素とマンノースが C-C 結合で結合した C-マンノシルトリプト ファンの存在も近年明らかになり、大きな話題となった4)。このような糖鎖付加の有無や 糖鎖構造は、同一タンパク質の特定部位をとってみても必ずしも均一ではなく、多くの場 合ミクロ不均一性(microheterogeneity)に起因する構造多形(glycoforms)が存在するため
5)、糖鎖や糖タンパク質の構造と機能を一義的に関連づけるためには、それらを均一な分 子として合成することが求められている。
N-結合型糖タンパク質の生合成は、細胞内の小胞体においてドリコール二リン酸に結 合した14糖Glc3Man9GlcNAc2が、糖転移酵素Oligasaccharyl Transferaseの作用を受けて、
新生ポリペプチド上の Asn-X-Thr/Ser 配列(X は Pro 以外の任意のアミノ酸)の Asn 残基へ 転移される事から開始される6)。タンパク質に転移された Glc3Man9GlcNAc2は、小胞体内 で種々のプロセッシングを受けながら分子シャペロンであるカルネキシン(CNX)/カルレ
O AcHN OH HO R1O
HN NH
HN O
O O
OH NH
O
N-Linked Glycoprotein
R' O
AcHN OH R1O
O-Linked Glycoprotein
OH
NH
HN O O
HN R2
O
ティキュリン(CRT)に認識され、タンパク質を正しい高次構造へと導く役割を担う。小胞 体内において正しい高次構造を獲得できた糖タンパク質は、その後ゴルジ体へと輸送され、
α-マンノシダーゼの作用で Man5GlcNAc2-Asn 構造にまでトリミングされる。ゴルジ体に おいて、この Man5GlcNAc2-Asn 構造は様々な糖転移酵素の作用を受け、ハイマンノース 型、ハイブリッド型、複合型の3種の糖鎖構造として完成されるが、これらの糖鎖は全て Man3GlcNAc2という共通の5糖構造を有している(Figure 0-2)。
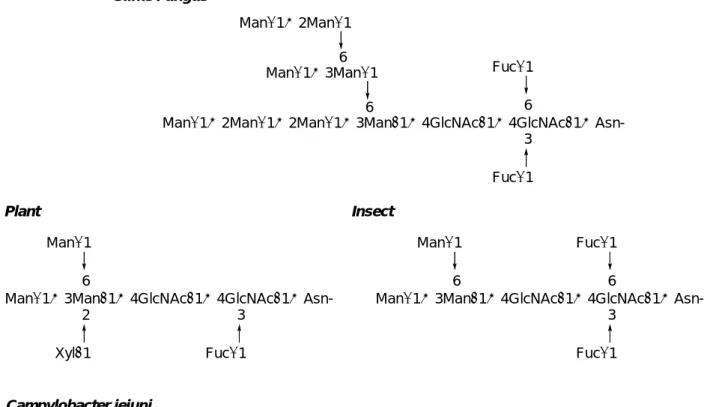
また、これら N-結合型糖鎖の生合成の初期経路は、全ての植物及び動物において共通 であるものの、その後の糖鎖プロセッシングの過程において種による相違が表れ、Figure 0-3に示すような多様性をもっている6b)。
一方で、近年の研究の結果、寄生虫や原核生物を由来とする特異的な構造を有する新
規な N-結合型糖鎖の存在が明らかになり、アレルギーや種々の疾病との関連性から、こ
れらの糖鎖は大きな注目を浴びている。例えばこれまで N-結合型糖鎖を有していないと されていた原核生物である Campylobacter jejuni から、新規な N-結合型糖タンパク質が近 年同定された7)。その構造は、N-アセチルバシロサミン(Bac(NAc)2)という特異な糖残基を
NeuAcα2→3Galβ1→4GlcNAcβ1→2Manα1
NeuAcα2→3Galβ1→4GlcNAcβ1→2Manα1→3Manβ1→4GlcNAcβ1→4GlcNAcβ1→Asn- Fucα1
6 6
Manα1→2Manα1→2Manα1→3Manβ1→4GlcNAcβ1→4GlcNAcβ1→Asn-6 Manα1→2Manα1
Manα1→2Manα1→3Manα16
Galβ1→4GlcNAcβ1→2Manα1→3Manβ1→4GlcNAcβ1→4GlcNAcβ1→Asn-6 Manα1
Manα1→3Manα16 High Mannose Type
Hybrid Type
Complex Type
Figure 0-2. Three Types of N-Glycans
Manα1→2Manα1→2Manα1→3Manβ1→4GlcNAcβ1→4GlcNAcβ1→Asn-6 Manα1→2Manα1
Manα1→3Manα16 Slime Fungus
Manα1→3Manβ1→4GlcNAcβ1→4GlcNAcβ1→Asn- Manα1
6
Xylβ1 2
Fucα1 3
Fucα1 3 Fucα1 6
Plant
Manα1→3Manβ1→4GlcNAcβ1→4GlcNAcβ1→Asn- Manα1
6
Fucα1 3 Insect
Fucα1 6
GalNAcα1→4GalNAcα1→4GalNAcα1→4GalNAcα1→4GalNAcα1→3Bac(NAc)2β1→Asn-
Glcβ1 3 Campylobacter jejuni
Figure 0-3. Evolution of N-Glycans
還元末端に持ち、GalNAc がα1-4 結合で繰り返すといった真核生物とは大きく異なったも のである。このCampylobacter jejuniはCampylobacter感染症の原因菌といわれているが、
これまでの研究成果からこの新規な糖鎖構造が病原としての働きに重要であるとされてい るため8)、本糖鎖の機能には大きな注目が集まっている。
共同研究者の伊藤らは早くから N-結合型糖鎖に着目し、有機合成化学的手法を用いた ハイマンノース型糖鎖の効率的合成法を確立すると共に9)、合成した糖鎖を用いてタンパ ク質の品質管理機構(quality control)に関わる種々の生命現象の解明に取り組んできた10)。 有機合成化学的手法によって糖鎖を合成することにより得られる利点は大きく分けて二つ あり、一つは構造の均一な糖鎖を必要充分量得られる点、もう一つは非天然型糖鎖の合成 が可能である点である。前述の通り、生物試料由来の糖鎖や糖タンパク質は、その構造多 様性が災いとなり、必ずしも均一な構造を有していないばかりか、その供給量も限られて
その構造の均一性は不可欠な要素であるから、化学合成法の持つ利点は極めて大きいとい える。また、糖鎖にまつわる生命現象を理解する上で、非天然型の構造を有する糖鎖の持 つ意義も極めて大きい。非天然型糖鎖は、その名の通り天然の試料から供給することは不 可能であり、化学合成法を通じてのみ合成可能であるが、これにより、天然型糖鎖の立体 異性体や位置異性体のみならず、一部の糖残基が欠落もしくは付加した糖鎖や、他の糖残 基に置換された糖鎖をも合成できる。これらの非天然型糖鎖を用いることにより、機能発 現に関与している糖鎖の立体構造や位置を正確に見積もることができ、より定量的な議論 を行うことが可能となる。また、化学合成法を用いれば、糖鎖のみならず、糖鎖に対して 機能性分子を付加させた分子プローブ11)や人工糖タンパク質12)の合成も可能となり、これ らの化合物は生命現象の解明に大きな効果を発揮することが期待される。
しかしながら、ここに述べたような化学合成法による糖鎖合成の利点を完全に享受す るためには、任意の糖鎖を自在に合成できる方法論の確立が不可欠であることはいうまで もない。糖は周知の通り多くの水酸基を持ち、糖鎖を構成するグリコシド結合もα-結合と
β-結合の2種類が存在しうるため、位置及び立体選択性を制御した糖鎖合成を行わなけれ
ば、数多くの副生成物を得るのみで、均一な糖鎖を合成することは不可能である。糖鎖合 成の歴史をひもとけば、Koenigs-Knorr 反応13)以来、多くの方法論が確立されて現在に至 っており14)、それらを効果的に用いることで、実に多くの糖鎖合成がなされてきた。例え ば、グリコシル化反応の立体制御方法としては、隣接基関与による立体制御といった古典 的なものから、in situ Anomerization 法15)や分子内アグリコン転移反応16)等が開発され、
1,2-trans及び1,2-cis配置17)のグリコシド結合の作り分けが可能となった。また、複雑な糖
鎖を効率よく合成する方法として、Armed and disarmed の概念18)や Orthogonal19)の概念に 基づく糖鎖合成法、固相合成法20)、可溶性ポリマー21)やフルオラスタグ22)を用いた固相-液 相ハイブリッド法等が、開発された。近年では、西村らや Seeberger らにより、糖鎖の自 動合成機の開発も成し遂げられた23)。
とはいえ、今なお合成が困難な糖鎖は少なくないため、従来の方法論の見直しと新規 反応の開発が継続して進められている。今日においても合成が困難な糖鎖の一例として、
本研究において著者が合成を試みた多分岐型糖鎖が挙げられる。多様に分岐した骨格を有 する糖鎖を合成するためには、位置選択性を確実なものとするために保護基の選定が重要
となる。多くの場合Lweis酸条件を必要とするグリコシル化反応の条件下では、用いるこ とができる保護基は限られており、また、保護基はグリコシル化反応の反応性や選択性に 直結する要素であるため、その選定は容易ではない。さらに、糖鎖の分岐が多くなれば多 くなるほど、分岐の中心に存在する糖の持つ水酸基の反応性が著しく低下することが予想 され、効率の良い糖鎖合成が困難になる。また、近年糖鎖合成において威力を発揮してい る酵素法を用いても、糖転移酵素や糖加水分解酵素の基質特異性を考えれば、多分岐型糖 鎖の合成は困難であるといえる。例えば、梶原らは鶏卵から得られる複合型糖鎖に対して、
種々の糖加水分解酵素を作用させて様々な種類の糖鎖や糖ペプチドの合成を成し遂げてい るが24)、前記の理由から複雑に分岐した糖鎖の合成には至っていないのが現状である。
著者はこのような現状を鑑み、より効率的な糖鎖合成法の確立を目指して凍結反応場 を用いたグリコシル化反応の最適化を図ると共に、これまで、その構造の複雑さから合成 がなされたことがなかった寄生虫由来の多分岐型 N-結合型糖鎖の合成研究に取り組んで きた。また、N-結合型糖鎖とペプチド部分の Asn 残基との間で効率よく N-グリコシド結 合を構築する手法の開発にあたり、これらの研究成果を通じて糖タンパク質の化学合成に 向けた方法論の確立を目指した。以下に本研究の概略を示す。
第1章 凍結反応場を利用した効率的糖鎖合成法の開発25)
糖鎖をより効率よく構築するために、様々な手法が開発されているが、本研究では反 応溶媒を凍結させてグリコシル化反応を行うことにより、その反応性が著しく向上するこ とを見いだした。種々反応条件を検討し、最適な反応条件を確立すると共に、本法を構築 困難な糖鎖の合成に応用した。第2章の合成研究においては、凍結反応を鍵反応に採用す ることで、標的糖鎖の骨格構築を達成することができた。
第2章 寄生虫由来の多分岐型N-型糖鎖の合成研究26)
寄生虫、日本住血吸虫及びマンソン住血吸虫の卵に存在する N-型糖鎖の合成を達成し た。この糖鎖は寄生虫の感染とアレルギーの発症との間に存在するとされる因果関係を明
効率的に供給する意義は極めて大きいといえる。また、糖鎖の構造活性相関を明らかにす るために必要となる様々な種類の非天然型糖鎖の合成も達成し、天然から得ることのでき ない糖鎖をも供給できるという化学法の持つ大きなアドバンテージを示すことができた。
第3章 N-結合型糖アミノ酸の新規合成法の開発27)
アスパラギン側鎖のカルボキサミドをヒドロキサム酸エステルへと変換することによ って窒素原子の求核性を向上させ、N-グリコシル化を経由する新規な N-結合型糖アミノ 酸の合成法の開発にあたった。さらに、この合成法を改良すべく、分子内 N-グリコシル 化反応を用いた N-結合型糖アミノ酸の合成法の確立にも取り組んだ。ヒドロキサム酸エ ステルを用いた手法では、N-グリコシル化反応の立体選択性が完全ではなく、O-グリコ シル化が副反応として進行するといった欠点があったため、分子内反応を用いることでこ れらの欠点の克服にあたった。また、N-(ベンジルオキシ)カルバミン酸エステルを用いた N-グリコシル化反応の開発し、効率的なグリコシルアミン誘導体の合成法を確立した。
以下、本研究の成果について順に詳述する。
本 論
第 1 章
凍結反応場を利用した 効率的糖鎖合成法の開発
1.1 はじめに
一般的な化学反応は、均一溶液系においてもっともスムーズに進行するとされている ため、反応温度は常に溶媒の融点よりも高い温度を求められる。凍結した溶媒中では反応 基質や試薬の拡散が極めて制限されるため、反応の進行は極めて遅くなるはずであるが、
凍結させた溶媒中で反応を行うことにより、その進行が大きく加速するとの報告も同時に 知られている。この効果は、化学反応28)、酵素反応29)共に見られ、溶媒系についても水系、
有機溶媒系共に見られるが、これらの特殊な実験事実は”concentration effect”と呼ばれる効 果で説明されている30)。反応温度が溶媒の融点以下であっても共融温度以上であれば、凍 結溶媒中には極少量の液体相が存在し、固体相との間で平衡関係が成立している。このと き、反応基質や試薬は液体相に溶解されることとなるため、その基質濃度は極めて高くな り、分子間反応に代表される二分子系の反応が急激に加速されるというわけである。この 説に従えば、単一分子でのみ進行する分子内反応については反応性の向上は期待できず、
かえって分子間反応という望まない副反応の進行を助けることになりかねない。
糖鎖構築に欠かすことのできないグリコシル化反応は、グリコシルアクセプターとグ リコシルドナーとの間でグリコシド結合を構築させる分子間反応であるため、凍結反応場 を利用すれば、その反応性が大幅に向上することが期待できる。N-結合型糖タンパク質の 化学合成を目指すにあたっては、多くの糖鎖を効率よく合成し、また構築が困難な糖鎖の 合成に挑戦していかなければならない。凍結反応によりグリコシル化反応の加速が認めら れれば、多くの糖鎖を短時間で効率よく合成可能となるばかりか、今まで合成が不可能で
あった糖鎖の合成を実現できる可能性がある。
本章においては、著者が確立した凍結反応場を用いたグリコシル化反応について詳述 するが、第2節においては凍結反応場を用いたグリコシル化反応の結果を均一系溶媒にお けるグリコシル化反応の結果と比較すると共に、その温度依存性について述べる。第3節 では構築困難な糖鎖を凍結反応で合成した結果を述べ、その優位性を示す。
1.2 凍結反応の確立
凍結反応場を利用したグリコシル化反応の確立にあたり、モデルとする反応の溶媒と グリコシルドナー及びグリコシルアクセプターの選定を行った。凍結反応の優位性を示す と共に、グリコシル化反応の立体選択性や温度依存性にまで議論が可能となる反応系の構 築が重要である。
1.2.1 溶媒の選定
多くのグリコシル化反応は、0 ˚C以下の低温条件において進行させることが多いため、
使用される溶媒もジクロロメタン(融点 : −97 ˚C)やトルエン(融点 : −93 ˚C)、ジエチルエー テル(融点 : −116 ˚C)といったように融点が低いものが多い。凍結反応場の利用により、反 応速度が飛躍的に向上することが期待されるとはいえ、一般的な反応速度の温度依存性を 考えれば、用いる溶媒の融点は高い方が望ましい。そこで、グリコシル化反応に汎用され るトルエンと似た物理的性質を有し、融点が 12-13 ˚C と比較的高い p-キシレンを凍結反 応に利用することとした。
1.2.2 グリコシルドナーとグリコシルアクセプターの選定
反応系の確立においては、できるだけ単純な構造を有するグリコシルドナーとグリコ シルアクセプターを用いる点に留意したほか、両者の反応性があまり高くない基質を採用 することとした。これは、あまりに反応性が高くては、均一系溶媒における反応との比較 において、凍結反応の優位性を明瞭に示すことができないためである。また、凍結反応に よる立体選択性に関する知見をも得たいと考えたので、用いるグリコシルドナーの保護基 は隣接基関与が発現されないベンジル基を用いた。以上の条件を満たすグリコシルドナー とグリコシルアクセプターとして、マンノース誘導体 1 および 231)をそれぞれ選定した。
マンノース誘導体 1 はチオメチルグリコシドであるが、チオメチル基はグリコシル化反 応において脱離基として機能する他に、糖鎖合成において効果的な保護基になるという側 面も有している。グリコシルドナーに利便性の高いチオメチルグリコシドを採用すれば、
次に、チオメチルグリコシドであるグリコシルドナーの活性化試薬を選定しなければ ならない。チオメチルグリコシドの活性化を行う試薬としては、NIS/TfOH32)、DMTST33)、
MeOTf34)等がよく知られているが、このうちNIS/TfOH とDMTST を用いるグリコシル化
反応は、多くの場合低温条件下で行うことからもわかるように、その活性化能力は比較的 高い。しかし、MeOTf に関しては、室温や加熱条件下で反応を行うことが多く、前出の 活性化試薬と比較して強力とは言い難い。強力な活性化試薬を必要としない本反応系にお いては、MeOTfを用いることが最適であるといえる。
1.2.3 凍結反応による反応加速 - 1
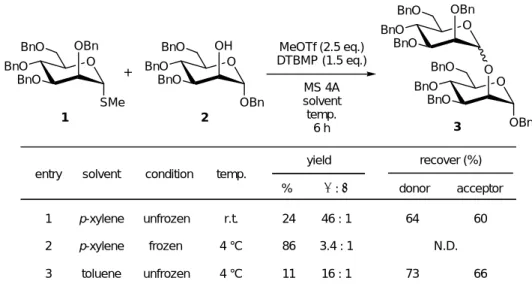
これまでの議論を元に、グリコシルドナー1 とグリコシルアクセプター2 を用いて、p- キシレン溶媒中でグリコシル化反応を試みることとした(Table 1-1)。
一般的なグリコシル化反応と同様、反応系には脱水剤として MS 4A を、プロトンスカ ベンジャーとして DTBMP をそれぞれ加え、活性化試薬として MeOTf を用いて、まずは 室温で反応を行った(entry 1)。6 時間で反応混合物を処理したところ、目的の 2 糖 3 はわ ずか 24%しか得られず、グリコシルドナーとグリコシルアクセプターが共に 60%程度回 収された。これと同じ反応を、今度は4 ˚Cで行った(entry 2)。前述の通り、p-キシレンの
融点は 12-13 ˚C であるから、この条件において反応系は凍結される。反応液は室温条件
下で混合し、最後に活性化試薬である MeOTf を加えた後に反応容器ごと液体窒素に浸し
BnO O BnO
BnO OH
OBn BnO O
BnO
BnO OBn
SMe
BnO O BnO
BnO
OBn BnO O
BnO
BnO OBn
O MeOTf (2.5 eq.)
DTBMP (1.5 eq.) MS 4A solvent temp.
6 h
entry temp.
yield recover (%) donor acceptor
% α : β 1
2
r.t.
4 °C
24 86
46 : 1 3.4 : 1
64 N.D.
60 condition
frozen unfrozen
3 unfrozen 4 °C 11 16 : 1 73 66
solvent
p-xylene p-xylene toluene
Table 1-1. Glycosylation under Frozen Media - 1
+
1 2
3
て反応系を一気に凍結させた。その後反応容器を4 ˚Cの冷蔵庫内で静置し、6時間後に取 り出して溶媒を融解させた後に、反応を終了させて後処理を行った。この反応では収率 86%で目的の2糖3が得られ、室温で行った全く同じ反応と比較して大幅な収率上昇が見 られた。
一方で、立体選択性に目を向けるとこれもまた大きく異なった結果となっている。室 温条件で反応を行った場合はα:βの比が46:1であったが、凍結反応においては3.4:1とα選 択性が大幅に低下している。マンノースは2位水酸基がアキシャル配置であり、アノマー 効果が強く表れるといった立体電子的要因が働くため、α体の生成する傾向が強い。entry 1 ではこれが顕著に表れているわけだが、entry 2 でα選択性が低下した要因は、立体選択 性を発現させる種々の要因が無視されるほどに、反応速度が飛躍的に向上したためと思わ れる。
また、entry 3にはentry 2と同様の反応をトルエン中で行った場合の結果を示す。溶媒
がトルエンであるため、この際反応系は凍結せずに均一溶液のままであるが、6 時間の反 応で得られた 2 糖 3 はわずか11%にとどまってしまった。この結果から、entry 2 で反応 速度が大幅に向上した要因は、(一般的には考えられないことであるが)温度の低下に依存 したものではなく、反応系を凍結させたことが要因であると結論づけることができる。
以上の結果から、凍結反応場を用いることでグリコシル化反応の反応速度が飛躍的に 向上することを示すことができた。
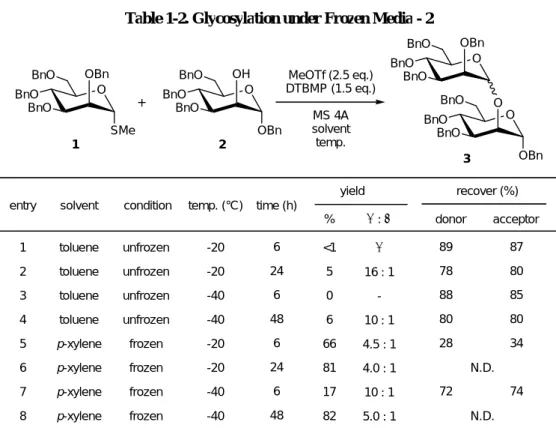
1.2.4 凍結反応による反応加速 - 2
化学反応は一般的に反応温度が高ければ、反応速度が加速される。そのような関係が 凍結反応という特殊な反応系においても成立しているか調べることとした(Table 1-2)。
entry 1〜entry 4にあるように、トルエン溶媒中で行った均一系溶媒の反応においては、反
応温度の低下に伴って反応速度は低下し、また反応時間が長ければ反応収率は向上する。
一方凍結反応に目を向けると、4 ˚C では完結に 6 時間要した反応が、−20 ˚C という条件 では反応完結にまで 24 時間を要し、6 時間では 66%しか生成物を得ることができなかっ
た(entry 5, 6)。さらに、反応温度を−40 ˚Cにまで低下させると、6時間ではわずか17%し
BnO O BnO
BnO OH
OBn BnO O
BnO
BnO OBn
SMe
BnO O BnO
BnO
OBn BnO O
BnO
BnO OBn
O MeOTf (2.5 eq.)
DTBMP (1.5 eq.) MS 4A solvent temp.
entry temp. (°C)
yield recover (%) donor acceptor
% α : β condition
solvent 1
2 3 4 5 6 7 8
unfrozen unfrozen unfrozen unfrozen frozen frozen frozen frozen toluene
toluene toluene toluene p-xylene p-xylene p-xylene p-xylene
-20 -20 -40 -40 -20 -20 -40 -40
<1 5 0 6 66 81 17 82
α 16 : 1
- 10 : 1 4.5 : 1 4.0 : 1 10 : 1 5.0 : 1
89 78 88 80 28
72
87 80 85 80 34
74 N.D.
N.D.
6 24
6 48
6 24
6 48 time (h)
Table 1-2. Glycosylation under Frozen Media - 2
+
1 2
3
以上の結果から明らかなように、凍結反応においても一般的な化学反応と同様に温度 依存性が存在し、反応温度が高ければ高いほど反応速度も加速されると結論づけることが できる。
反応系を凍結させるためには、反応温度を融点以下に設定しなければならないため、
凍結反応において最大の反応速度を得るためには、融点にできるだけ近い、融点以下の反 応温度を設定する必要がある。
1.3 凍結反応を利用した構築困難な糖鎖の合成
N-型糖鎖の一種であるハイマンノース型糖鎖は糖鎖の翻訳後修飾に深く関与しており、
その機構について詳細な検討が行われている9),10)。ハイマンノース型分子プローブはその 機構解明を行う際に用いられる分子であり、用途に応じて様々な種類のものが合成されて いる11)。著者は共同研究者らと共に、新たな分子プローブとして 2糖 4 や 3糖 5、9 糖 6 を設計した(Figure 1-1)。
これらの糖鎖の合成を試みたところ、2糖4や3糖5の合成においては、マンノシルド ナーの有するTBDMS基の立体障害が影響してか、グリコシル化反応が円滑に進行せず、
また、9糖 6の合成にあたっては、その複雑な糖鎖構造が影響してか、マンノトリオース 構造を有するマンノシルドナーを効率よく導入することが困難であった。s
前節において確立した凍結反応は、グリコシル化反応の反応速度が大幅に向上すると いう利点を有している。この利点は、反応時間の短縮に貢献するばかりか、通常の反応条 件で進行しづらいグリコシル化反応をも進行させるという可能性を秘めていると考えられ たので、構築が困難であったこれら三種類の分子プローブを合成する際に凍結反応を試み、
その反応性が向上することを期待した。その結果、いずれの反応においても反応速度の向
O OAc
BnO O BnO
BnO
O
BnO O BnO
BnO
O HO
BnO O BnO
BnO
O
OTBDMS
O O
OBn PhthN
BnO O
NPhth OAll OBn O
BnO BnO O
BnO BnO BnO O
BnO BnO
O BnO O
BnO BnO
O OTBDMS
O O
O O OBz O
OR BnO O
BnO
BnO OTBDMS
O O OBz O
OR BnO O
BnO BnO
O
O BnO O
BnO
BnO OTBDMS R = (CH2)8CO2Me
4
5 6
: Structure derived from glycosyl donor
上が認められ、従来法では構築困難であったこれらの糖鎖を効率よく合成することができ た。
これまで見てきたように、凍結反応場においてグリコシル化反応を行うと、その反応 性の飛躍的な向上が見られため、本法は、構造が複雑な糖鎖を構築する上で有効な方法論 になりえるといえる。特に本法は、特別な実験器具や複雑な操作を要求されることなく、
単に反応溶液を凍結させ、冷蔵庫内に静置するとういごく単純な操作で実現されている点 が大きな利点であるといえる。また、ある種のグリコシル化反応では、その立体選択性が 均一溶媒系の反応と比較して変化するという特徴も有している。
第 2 章
寄生虫由来の 多分岐型 N- 型糖鎖の合成研究
2.1 はじめに
住血吸虫(Schistosome)は寄生性の蠕虫(helminth)であり寄生虫の一種であるといえるが、
開発途上国を中心に二億人以上が感染の被害に遭っている35)。その一方で、衛生環境の整 った先進国ではその感染件数は減少傾向にあり、例えばその名に「日本」を持つ日本住血 吸虫(Schistosoma japonicum)は、確かに過去に日本国内に生息し猛威をふるっていたが、
1978 年以降の新患は認められていない。しかしながら、世界的規模で見れば寄生虫の脅 威が払拭されたとはいえず、寄生虫感染や寄生虫疾患に関する研究が今なお盛んになされ ている。
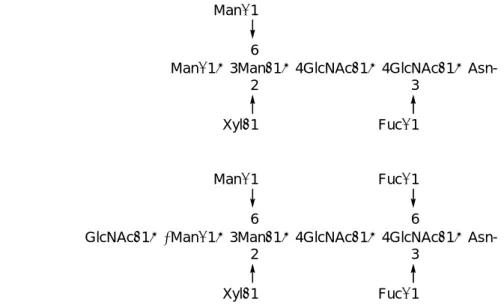
最近の研究から、これらの寄生虫は哺乳動物とは異なる特徴的な N-結合型糖鎖を発現 していることが明らかになってきた。興味深いことに、それらの糖鎖は植物由来の糖鎖と
同様に D-キシロースや L-フコースを含有しているが、それらの糖残基は、β-マンノース
(Xylβ1→2Man)と還元末端側の N-アセチルグルコサミン(Fucα1→3GlcNAc)にそれぞれ結合 している36)。また、これらの糖鎖の部分構造は、人に対して抗原性を有していることが示 されており37)、植物性アレルゲンのIgE抗体との結合に関与している(Figure 2-1)38)。
一方で、マンソン住血吸虫(Schistosoma mansoni)の感染により TH2 型の免疫応答が、寄 生虫由来の糖鎖構造を認識すること39)により誘導されること40)が報告されている。TH2 が 優位になることは、アレルギーを発症しやすくなる要因となるにも関わらず、寄生虫に感 染するとアレルギーに感染しづらくなると言う説は、比較的古くから提唱されている。こ の矛盾した関係を明らかにするために、両者の因果関係については多くの興味が持たれ、
NeuAcα2→3Galβ1→4GlcNAcβ1→2Manα1
NeuAcα2→3Galβ1→4GlcNAcβ1→2Manα1→3Manβ1→4GlcNAcβ1→4GlcNAcβ1→Asn- Fucα1
6 6
Manα1→3Manβ1→4GlcNAcβ1→4GlcNAcβ1→Asn- Manα1
6
Xylβ1 2
Fucα1 3
GlcNAcβ1→2Manα1→3Manβ1→4GlcNAcβ1→4GlcNAcβ1→Asn- Manα1
6
Xylβ1 2
Fucα1 3 Fucα1 6 (A)
(B)
(C)
Figure 2-1. Typical Structures of Animal (A), Plant (B), and Helminth (C) Derived Complex-type Glycans
その解明に向けた研究が行われてきた41),42)。当初、この矛盾は、”IgE blocking hypothesis”
と呼ばれる仮説で説明されてきた。これは、寄生虫の感染によって産生されるポリクロナ ールなIgE抗体が、体内に存在する肥満細胞のIgEレセプターを塞ぎ、アレルゲンの進入 により産生される特異的なIgE抗体とIgEレセプターの結合を阻害するとすることにより、
アレルギーの発症が抑制されるとする説である。しかしながら、近年ではこの学説を否定 する研究結果が示され、その妥当性に疑問を持つ声も多い。最近では、アレルギー感染が 衛生環境に依存しているという衛生仮説(hygiene hypothesis)という学説が主流になりつつ ある。これは、衛生環境が整い、抗生物質等の投与が盛んである都市部では、細菌やウイ ルス等の感染が激減することで TH1 型の免疫応答が弱くなり、その結果 TH2 が優位とな ってアレルギーに感染しやすくなってしまうとういものである。この説にしたがえば、衛 生環境の整っていない環境下においては、TH1 型の免疫応答が充分に刺激されているため に、TH2 とのバランスが保たれ、アレルギー感染が抑制されていると説明することはでき る。しかしながらこの説では、寄生虫感染とアレルギー感染の間に存在する因果関係を説 明することはできない。さらに近年、この衛生仮説を元に新たな学説が示されたが、この
説においては、インターロイキン-10 (IL-10)といった抗炎症性サイトカインの増加が、ア レルギーの感染を抑制していると主張している。この IL-10 は、長期にわたる寄生虫感染 により制御性 T 細胞から産生されるサイトカインであるから、この説においては両者の 因果関係も成立している。
前述の通りある種の寄生虫由来の糖鎖は、TH2 型の免疫応答を誘導することが知られて いるため、これまで示されてきた学説を分子化学的に解明していくためには、これらの糖 鎖は必要不可欠な存在であるといえるが、各々の事象における糖鎖や糖タンパク質の役割 を正確に理解することは極めて困難である。その原因として、生物試料を由来とするそれ らの化合物の供給量が絶対的に少なく、またその構造が必ずしも均一ではないことが挙げ られる事は前にも述べたとおりである。そこで著者は、寄生虫由来の N-型糖鎖を均一な 分子として有機合成化学的手法により合成し、供給することを目的にその合成研究を開始 することとした。標的とする糖鎖は、寄生虫、日本住血吸虫及びマンソン住血吸虫の卵に 存在するN-型糖鎖である(Figure 2-1-c)43)。また、本糖鎖の構造と機能の関連性を明らかに するために、天然型の9糖の他、糖鎖の機能発現に関与しているとされるキシロースやフ コースを含まないような非天然型の糖鎖も合成の標的糖鎖にすることとした。一方で、標 的とする糖鎖は N-アセチルグルコサミン、マンノース、フコースさらにはキシロースと 多くの糖残基から構成されているばかりか、多くの分岐点を持つ非常に複雑な糖鎖骨格を 有している。このため、生物学的視点のみならず、有機合成化学的視点から見ても、本糖 鎖は非常に興味深い標的化合物であるといえよう。
本章では標的糖鎖の合成研究に関する結果について詳述するが、まず第 2 節において は標的とする天然型の糖鎖のみならず、非天然型の糖鎖をも効率よく合成できるよう考慮 した標的糖鎖の逆合成解析を示す。第3節および第4節では逆合成解析により分割した各 フラグメントの合成について述べ、第5節ではそれらのフラグメントを元に達成された標 的糖鎖の合成法を示す。また第6節においては、天然型糖鎖の合成結果を踏まえて確立し た、キシロースやフコースを含まない非天然型糖鎖の合成法を示す。
2.2 逆合成解析
前節で述べたとおり、標的糖鎖の構造と機能の関連を明らかにするためには、天然型 の糖鎖のみならず非天然型の糖鎖を合成しなければならないので、本合成研究においはそ れらの糖鎖をも効率よく合成できるような合成経路の確立が必須になるといえる。そこで 著者は、天然型の9糖7を6糖からなるフラグメントA 8と3糖からなるフラグメントB 9 の二つのフラグメントに分割し、両者を合成した後にグリコシル化を行って標的糖鎖へ と導く集約的合成法を採用することとした。フラグメントA 8にはキシロースが、フラグ メントB 9にはフコースがそれぞれ含まれているので、各フラグメントを合成する際にこ れらの糖残基を含まないような糖鎖を別途合成すれば、それらの組み合わせにより様々な 種類の糖鎖を効率よく合成することができると考えた(Figure 2-2)。
O O O
OH NHAc HO
O OH
OH HO
O
HO O HO
NHAc
O OH
OH O OH HO
O HO
O HO
OH
O HO
HO
O O
NHAc OH O
OHOH
OH O
O HO OH HO
O
O O
NPhth OBn O
OAcOAc
OAc O
O AcO OAc BnO
O HO O O
OAc PhthN AcO
O OAc
OAc AcO
O
AcO O AcO
NPhth
O OAc
OAc O OAc AcO
O AcO
O AcO
OAc
O AcO
AcO
O CCl3 NH
O BnO
OAc BnO
BnO
Cl
BnO O BnO
NPhth
O OBn
OBn O OBn BnO
O HO
O O
O OBn O
OBn PhthN BnO
O OAc
OAc AcO
O SMe
Fragment B Fragment A
Fragment A-1 Fragment A-2
Figure 2-2. Retrosynthesis of Target Sugar Chain 7
8 9
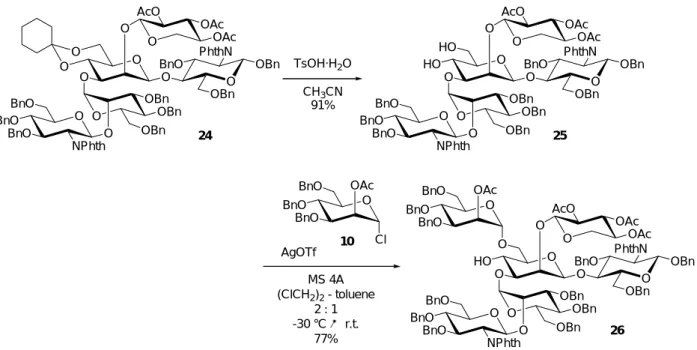
10 11 12
さらに、各フラグメントの逆合成解析を進めると、フラグメントA 8は3糖からなるフ ラグメントA-1 11と2糖からなるフラグメントA-2 12、およびマンノース誘導体1044)の 3つのブロックに分割可能である。フラグメントA-1 11の合成に関しては、構築困難なβ- マンノシド結合を立体選択的に構築することが重要となる他、多様に分岐した糖鎖構造を 構築していくために、効率的な保護、脱保護が求められる。
フラグメントB 9に関しては、3糖の中心となるグルコサミン誘導体に対して段階的に フコースを導入する方法で合成が可能となるが、この際フコースをα選択的に導入するこ とが、合成の鍵段階として挙げられる。
2.3 フラグメント A の合成
フラグメントA 8の合成においては、その合成ブロックとなるフラグメントA-1 11、A- 2 12を位置及び立体選択的な糖鎖伸長を経て、効率よく合成することが肝要となる。特に、
フラグメント A-1 11 は構築が困難なβ-マンノシド結合を有するほか、多様に分岐した構 造を有するフラグメントA 8の中心骨格であるだけに、位置選択的な糖鎖伸長が可能とな るよう保護基を選定し、選択的な脱保護反応を行わなければならない。以下、フラグメン トA 8の合成の詳細について述べる。
2.3.1 フラグメントA-1の合成
フラグメント A-1 11 の合成にあたっては、β-マンノシド結合の立体選択的合成を第一 に行わなければならない。β-マンノシド結合は、1,2-cis 配置のグリコシド結合であり、
1,2-trans 配置のグリコシド結合のように、隣接基関与を用いて立体選択的に構築すること
は困難である。この問題を解決するために、これまでに様々な方法論が提唱されてきた。
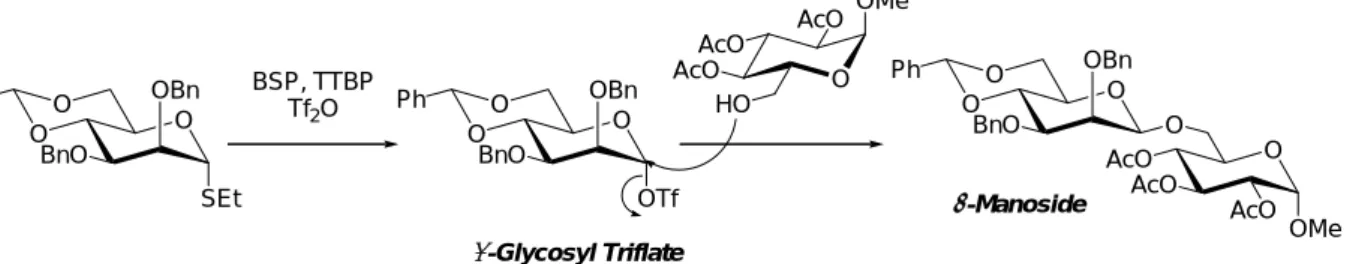
例えば近年 Crich らは、低温条件下で安定に存在するα-マンノシルトリフレートを系内で 形成させた後、グリコシルアクセプターとのグリコシル化反応を行うことで SN2 型の置 換反応が進行して、β-マンノシド結合が得られると報告している(Figure 2-3)45)。
また、グリコシルアクセプターをマンノシルドナーの 1 位のβ面に一時的に固定させ、
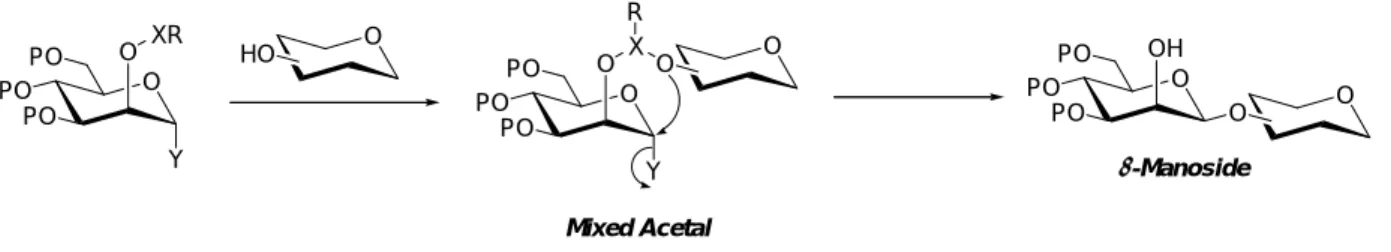
続く活性化により分子内でグリコシルアクセプターを転移させて望みのβ-マンノシドを得 るという、分子内アグリコン転移反応と呼ばれる反応も著名である(Figure 2-4)16)。この反 応において鍵段階となる、両者を一時的に連結させてグリコシルアクセプターをβ面に固 定する方法としては、マンノシルドナーの2位を介した混合アセタールを合成する手法が よく知られている。Barresi と Hindsgaul はイソプロピリデンアセタール46)を、Kim と
BSP, TTBP Tf2O
β-Manoside α-Glycosyl Triflate
O O
BnO O O
SEt Ph OBn
O BnO
O O
OTf
Ph OBn AcO O
AcO AcO
OMe
HO O
BnO O O Ph OBn
O AcO
AcO
AcO OMe
Figure 2-3. β-Mannosylation Developed by Crichet. al.
O PO
Y O XR PO
PO
O PO
Y O X PO
PO O
PO OH PO
PO
β-Manoside R
HO O
O O O O
Mixed Acetal
Figure 2-4. Intramolecular Aglycon Delivery (IAD)
Stork はシリルアセタール47)をそれぞれ用いた例を過去に報告している。また近年では、
Fairbanks らがビニルエーテルやアリルエーテルを介した混合アセタールの合成法を確立
している48)。
著者は、共同研究者の伊藤らにより開発された p-メトキシベンジリデンアセタールを 経由する、分子内アグリコン転移反応49)を用いて立体選択的にβ-マンノシド結合を構築し た。本法では、既知のマンノシルドナー1349c)とグルコサミン誘導体1450)をDDQで処理し て混合アセタール 15 を合成し、グリコシルアクセプターをマンノシルドナーのβ面に固 定させている。しかる後に、マンノシルドナーを MeOTf で活性化し、グリコシルアクセ プターを分子内で転移させてβ面からのみの選択的なグリコシル化反応を実現し、2 糖 16 の立体選択的合成を達成した(Scheme 2-1)。
このように、本反応を用いることにより、効率よくβ-マンノシド結合を構築することが 可能であったが、本法をフラグメント A-1 11 の合成に採用することで得られる利点はこ れのみではない。反応前にマンノシルドナー13の2位水酸基を保護していたPMB基は、
反応後の後処理で脱保護されるため、得られた2糖16においては、マンノースの2 位水
O TBDPSO
O O
SMe OPMB
O TBDPSO
O OH O
O OBn O
OBn PhthN BnO HO O
BnO
NPhth OBn
OBn
O TBDPSO
O O O
O OBn OBn PhthN BnO OMe
O
SMe +
MeOTf DTBMP
DDQ MS 4A CH2Cl2
MS 4A (ClCH2)2
45 °C 63%
14
15
16 13
MS 4A CH2Cl2 -40 ℃ promoter O
TBDPSO O OH O
O OBn O
OBn PhthN
BnO O
TBDPSO O O
O OBn O
OBn PhthN BnO
O OAc
OAc AcO
O
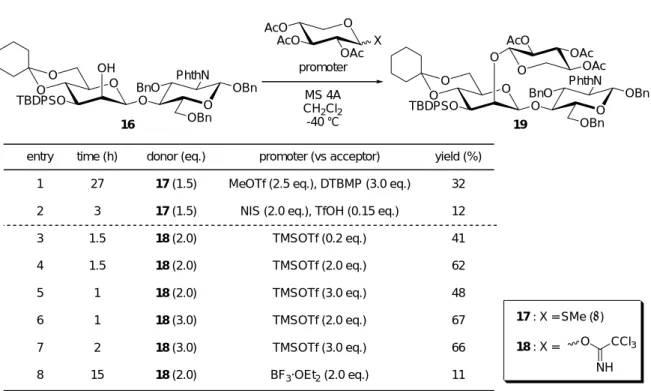
entry 1 2 3 4 5 6 7 8
time (h) 27
3 1.5 1.5 1 1 2 15
promoter (vs acceptor) MeOTf (2.5 eq.), DTBMP (3.0 eq.)
NIS (2.0 eq.), TfOH (0.15 eq.) TMSOTf (0.2 eq.) TMSOTf (2.0 eq.) TMSOTf (3.0 eq.) TMSOTf (2.0 eq.) TMSOTf (3.0 eq.) BF3·OEt2 (2.0 eq.)
yield (%) 32 12 41 62 48 67 66 11 AcO O
AcO