平成 2 9 年度修士論文
コイ赤血球膜中の糖タンパク質 グリコホリン の糖鎖機能の解析
三重大学大学院地域イノベーション学研究科 博士前期過程地域イノベーション学専攻
氏 名 小 高 耀 平
第
1章 緒 論
1第
2章 実験方法
122‑1
実験材料および試薬
2‑2実験方法
2‑2‑1
コイ赤血球の調製
2‑2‑2コイ赤血球膜の調製
2‑2‑3コイグリコホリンの調製
2‑2‑4タンパク質の定量
2‑2‑5タンパク質の染色
2‑2‑5‑1 CBB染色の方法
2‑2‑5‑2 PAS染色の方法
2‑2‑5‑3銀染色の方法
2‑2‑6静菌活性の測定
2‑2‑7
コイグリコホリンからの糖鎖の分離
第
3章結果と考察
223‑1
コイ赤血球膜の調製
3‑2
陰イオン交換クロマトグラフィー
3‑3
各
Fractionの銀染色によるタンパク質の検出
3‑4各フラクションの静菌活性
3‑5
~-脱離による糖鎖分離
第
4章 総 括
41第
5章 地域イノベーションに関しての総括
42参考文献
43F u n c t i o n a l A n a l y s i s o f O l i g o s a c c h a r i d e s from Carp G l y c o p h o r i n
Yohei Odaka
孔
1arch20181. Introduction
Glycophorin is a membrane glycoprotein in red blood cells and contains a large amount of sialic acid in their oligosaccharides.
I n
our laboratory, the glycophorin企
omblood of ca中 (
cyprim cαrpio) is revealed that their sialo‑oligosaccharides have a bacteriostatic activity [1]. However, preparation of oligosaccharides by using the extract method (LIS‑phenol method) is not adequate because this yield of glycophorin is significantly low (ca. 1 %). To improve the yield of glycophorin and its oligosaccharides, we adapted the anion‑exchange column chromatography [2]. However, ~-eliminationfor the preparation of oligosaccharides
企
om glycophorin fraction is not reacted sufficiently. This reason is supposed to be the existence of NaCl and excess detergent in the glycophorin企
action. To reduce NaCl and detergent, the企
actionwas applied to the desalting column (Econo‑pac lODG, Bio‑rad co. ltd), followed by ~--elimination. We aim to improve the method of oligosaccharides preparation by removing excess NaCl and detergent followed by~-elimin剖ion.
2. Method
We prep訂cred blood cell membranes
企
omcarp blood. Then, red blood cell membranes was solubilized by LIS. After that, desalting was carried out using Econo Pac‑10 DG column, and then anion exchange chromato‑graphy (DEAE Toyopearl 650M;TOSO Co.,ltd.) was carried out. Elution was performed by gradient concentration method with Na Cl. Dialysis and desalting column were used to remove excess NaCl and sodium cholate. After that, ~-elimination
was carried out. 3. Conclusions
We predicted that the existence of NaCl and detergent in glycophorin
企
actioninhibited the reaction of ~-elimin剖ion. Desalting by using an Econo‑PaclO DG column was performed. As a result of~-elimination, no resulting protein aggreg
剖
e wasprep訂ation was successfully by using the anion‑exchange column chromatography [2]. Therefore, obtained glycophorin fraction is suitable to prepare oligosaccharides efficiently. It is suggested th
剖
denaturationof the protein moiety by phenol or other reagents is required. We will study to perform the adequate method by using a weak acidic or alkaline protein denaturing agent other than phenol for oligosaccharides preparation企omglycophorin. AcknowledgementIm grate
白
lto Aoki ,T and other members in our laboratory.References
[1] Aoki ,T., Chimura, K., Nakao, N. and Mizuno, Y.
Isolation and characterization of glycophorin 企omcarp red blood cell membranes. Membranesラ
4,491
・
508(2014).[2] Fuziwara, R. The studies on the glycoproteins in carp red cell membranes. Graduation thesis, Faculty of Bio問sources,Mie University (2009).
第1章 緒論
[背景]近年、医学、理学、農学など様々な分野で遺伝子研究が盛んに行われてい る。遺伝子研究の次の段階としてポストゲノムが注目され、その中でも糖鎖の持つ生理 機能が関心を集めている。生体内で作られるタンパク質の多くは糖鎖で修飾されること によってその機能を発現している。よってタンパク質に結合した糖鎖の機能を解明する ことは大変重要であると認識されている。本研究室では中でも赤血球膜中の糖タンパク 質に結合している糖鎖に注目した。
ヒトの赤血球膜中のヒトグリコホリンは赤血球膜中の糖タンパク質の中でシアル酸を 大量に含む膜貫通タンパク質であり、一方でシアル酸をほとんど含まない膜タンパク質
は
Band 3と呼ばれている。グリコホリンの親水性のアミノ末端ペプチドは細胞膜の外
側に存在している。また親水性のカルボキシル末端は内側の細胞質に露出しており,そ れに連なる親水性のヘリックスが細胞膜の基本構造である脂質二重膜を貫通している
(図
1-1)。グリコホリンはヒト赤血球膜を
SDSで可溶化した後,PAS 染色をすること により、初めて
Fairbanksによって発見された[1]。当初は検出されたヒトグリコホリン を
PAS1, PAS2, PAS3,と呼んでいた。その後Steckら[2]はヒトグリコホリンを 4 種 類,Anstee ら[3]はヒトグリコホリンを
7種類検出した。Dahr ら[4]は血液型活性から
MNおよび
Ss Glycoproteinと命名した。現在はそれぞれヒトグリコホリン
A,B,Cおよ び
Dという別の名称が定着している。ヒトグリコホリンの生理機能として、インフルエ ンザウィルスのレセプターがグリコホリンであることを
Segrest[5]らが、またMN式血 液型の抗原活性があることを
Jackson[6]らが、それぞれ報告している。ヒトグリコホリン
A はMN型、ヒトグリコホリン
Bは
Ss型を持つとされ、それぞれ糖鎖のシアル酸を 取り除くと血液型活性が失われる。gu しかし、上記の機能はグリコホリンの見掛け上の 活性にすぎず,本来の機能は全く不明という報告もある[7]。グリコホリンを持たない赤 血球を有するものも発見され、そのようなグリコホリン欠損赤血球では、
Band 3がグリ コホリンの代わりに糖鎖を担っている。そのため赤血球の外側に一定量以上の糖鎖が存 在することが生命維持に不可欠であると考えられている[8]。
ヒトにおけるグリコホリンを含む赤血球膜中の糖タンパク質の研究は行われているが、
有核であり調製困難な鳥類や魚類の赤血球膜の研究報告はあまり多くはない。当研究室
は水産物の品質について研究を行っており、魚類血液中の筋肉の腐敗に関連する因子の
探索を行い、比較的容易に採取出来るという理由からコイ血液を対象として赤血球膜中 に含まれる糖鎖に注目し、平成
2年より研究を重ねてきた。その結果、コイ赤血球膜の 構成糖タンパク質であるコイグリコホリンに種々の細菌に対する静菌作用がある事を見 いだした[9]。 また、静菌作用の活性部位はコイグリコホリンの糖鎖に存在する事が判 明した(図
1-2)[10]。以降の研究の成果より、コイグリコホリンは血球膜から脱離する際、タンパク質部の疎水性により相互に凝集して球状の多量体として血中を浮遊し、こ の表面を覆っている糖鎖が血中に侵入した細菌と反応、吸着することによって、静菌作 用を生じることが明らかとなった。コイグリコホリンの静菌活性はペーパーディスク法 による感受性確認試験によって確認されている[10]。抗菌活性は
B. subtilis, A.hydrophilla, M. luteusV. anguillarum, E-coli (IFO3301)で確認されている
。A.
hydrophilla
は通性嫌気性のグラム陰性細菌である。エロモナスソブリアと共に淡水域の
常在菌であり、その周辺の魚介類にも存在し、食中毒起因菌として知られている。
V.anguillarum
はビブリオ属の通性嫌気性のグラム陰性桿菌であり、水中に多く存在する。
ヒトに対して病原性は持たないが、ブリ属などの多くの魚種に感染する。サケ、マス類 が発症するビブリオ病の起因菌としても知られ、遊泳が不活発になり、体表の黒色化や 眼球突出、出血斑などの症状がでる、養殖業において問題視されている病原菌である。
養殖業では水産用医薬品としてオキソリン酸やスルファモノメトキシンなどの投与によ る対策がされている。以上のことから、コイグリコホリンはグラム陰性及び陽性双方に 対して広い抗菌スペクトルを有していることが判明している。また、当研究室で、コス ティアやキロドネラといった細菌の寄生が原因で発症する魚類の白雲病に感染したコイ と健康的なコイを比較した結果、白雲病個体は白い斑点が体表面にでき、食欲不振、運 動製の低下など外面的な症状に加え、健康個体と比べその血液にも差異が認められた。
健康個体と比べ白雲病個体は赤血球膜中のグリコホリンを著しく欠いた状態であったと 報告されている[11]。
赤血球膜の構成タンパク質の中で静菌作用を持つものでは,現状はコイグリコホリンで
のみ報告されている[10]。コイが赤血球膜に静菌作用といった生体防御機構を獲得した
要因として、コイの自然界での生育環境が関連していると推測できる。コイは
BOD(生物化学的酸素要求量)の値が高く、汚れた水域でも適応でき、他の魚種と比較して寿命が
長いことが知られている。よって、コイと類似した環境に生息する魚種や、遺伝子的に
近縁の魚種、または魚類以外の生物にもコイと同様の生体防御機構が存在する可能性も 考えられる。
調製方法に関しては魚類の赤血球はヒトと異なり有核であるため、赤血球膜調製では 溶血操作と共に核を除去する操作が必要である。ニジマスの赤血球膜調製法については 既に
Micheland
Rudloff [12]が報告しているが、コイの赤血球はニジマスに比べて直径が約
20 %程小さく、溶血時における核の除去がさらに困難であった。本研究室の深井らはニジマス赤血球膜の調製法に改良を加えてコイ赤血球膜を調製した[13]。コイ赤血球 膜からコイグリコホリンを調製する手段としては、ヒトグリコホリン抽出法を用いる事 とし、まず始めに Michel and Cleve 法[14]を試みたが、コイグリコホリンは抽出でき なかった。そこで Blumenfeld
[15]らによるピリジンによる抽出法を検討したが、ピリジンの臭気が強い事と当時ドラフトが不備だったことから断念し、マルケッシによる
LISによる赤血球膜可溶化の後フェノールを用いて抽出する、LIS-フェノール法[16]を 試みた。図
1-4に概要を示す。しかし魚類の赤血球は有核であり、フェノールは糖タン パク質と共に核酸をも抽出するため、ヒトグリコホリン調製法の操作に加えて核酸除去 の操作を加える事により、混在する核酸を除去した。その結果、コイ赤血球膜からコイ グリコホリンを抽出する事が可能となった。
[目的]以上の検討によって,今までの研究では魚類のグリコホリン調製法はヒトグ リコホリンの抽出法である
LIS-フェノール法に核酸除去の操作を加えた方法を用いて調製してきた。しかしながら、本方法によるコイグリコホリンの回収率は、弱酸である フェノールを用いるために、糖鎖に含有しているシアル酸が遊離し易く、回収率が約1%
と非効率であり、生理機能を検討する十分な量が調製できるとは言い難かった。そこで、
本研究室ではコイグリコホリンの回収率をあげるべく、陰イオン交換クロマトグラフィ
ーを用いたコイグリコホリン調製法を平成
20年に開発を試みた[17]。その結果
LIS-フェノール法と比較し短時間かつ高い回収率でグリコホリンを調製することができた。し
かし、調製の整合性はシアル酸の回収率でしか評価がなされていない。また、コイグリ
コホリンの糖鎖部分のみを再現性良く調製する方法は確立していなかった。コイグリコ
ホリンの糖鎖に関わらず、自然界に存在するタンパク質の半数以上は糖鎖に修飾され生
理機能を発揮している糖タンパク質である。安定的なコイグリコホリンの調製法を確立
し、コイグリコホリンからの糖鎖分離方法が確立した場合、コイグリコホリンのみでな
く他の魚種においても、より発展的な糖鎖研究が可能になると期待される。よって、コ イグリコホリンから静菌作用を有した糖鎖を分離し、更なる生理機能を解明するにあた り糖鎖の分離方法を確立することは非常に重要な課題である。
[本論文の構成] 本論文では章に詳細な実験方法を記す。コイ血液から調製した赤
血球膜を懸濁させ、
LIS(3,3-ジヨードサリチル酸リチウム)を用いて可溶化し、陰イオ
ン交換カラム(DEAE Toyopearl 650M; TOSO Co.
ltd.)に負荷後、NaClを用いたグ
ラジエント溶出法を行ってグリコホリンを分画した。その後、-脱離反応を行って糖鎖
の分離を試みた。糖鎖の精製度の確認は、各フラクションを用いた
SDS-ポリアクリルアミドゲルスラブ電気泳動後の銀染色によるタンパク質染色の結果より確認を行い、糖
鎖の遊離の確認は、ペーパーディスク法による大腸菌に対する静菌活性の観察によって
判別した。3章に詳細な結果と考察を記す。陰イオン交換クロマトグラフィーで
0.2 M NaCl付近で溶出されたグリコホリンには静菌活性は認めらなかった。しかし、ペーパ
ーディスク法を改良した方法で不溶性のコイ赤血球膜の静菌活性を評価した結果、大腸
菌と直接接触した部分で生育阻害が認められた。また、赤血球膜を可溶化する際に用い
た
LISが大腸菌の生育に影響を与え、静菌作用を持つ可能性が示唆された。加えて
SDS-電気泳動によるタンパク質の染色結果から推測すると、-脱離反応によるグリコホリン
からの糖鎖の分離は可能であることが判明した。4章に本論文の総括、5章に地域イノ
ベーションに関する総括を記す。
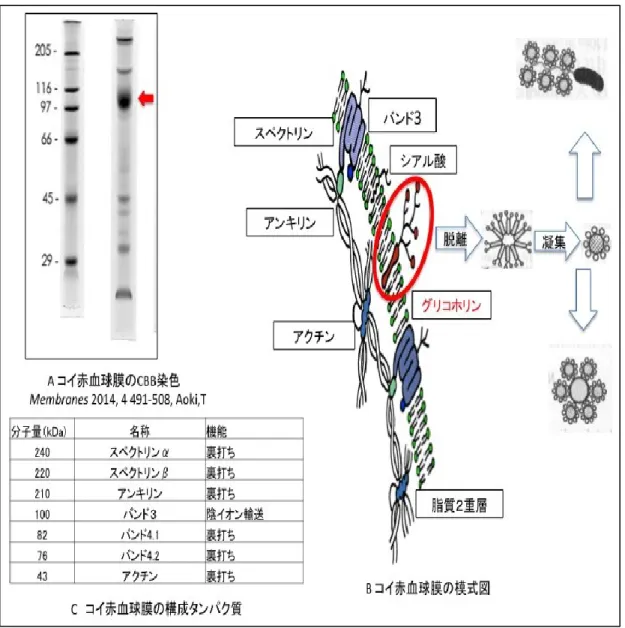
図
1-1.コイ赤血球膜の模式図
A はコイ赤血球膜の CBB 染色画像であり、赤血球膜を構成するタンパク質 を検出している[10]。コイ赤血球膜中には様々なタンパク質が存在するが、
グリコホリンは赤血球膜中の糖タンパク質の中でシアル酸を大量に含む分子
量約 100 kDa の膜貫通タンパク質である。一方でシアル酸をほとんど含まな
い膜タンパク質はバンド 3 と呼ばれている。バンド 3 の生理機能として呼吸
におけるガス交換や pH 調製が知られている。当研究室ではコイの赤血球膜を
調製し、膜構成タンパク質とグリコホリンを検出した。その結果、主要タン
パク質はコイとヒトはほぼ同様であったが、グリコホリンに関しては差異が
認められた[18]。B はコイ赤血球膜の模式図を示している。コイグリコホリ
ンの親水性のアミノ末端ペプチドは細胞膜の外側に露出している。また親水
性のカルボキシル末端は内側の細胞質に露出しており,それに連なる親水性
の ヘリックスが細胞膜の基本構造の脂質二重膜を貫通している。C はグリ
コホリン以外のそれぞれの分子量と細胞内における機能を示している[19]。
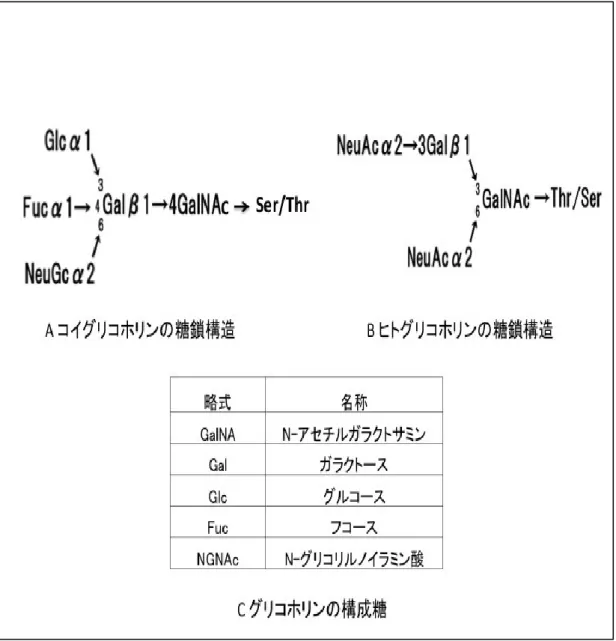
図
1-2.コイグリコホリンの糖鎖構造
A はコイグリコホリンの糖鎖構造、B はヒトグリコホリンの糖鎖構造を示 している。 C はグリコホリンの糖鎖を構成する糖の略式と名称を示している。
コイグリコホリンの糖鎖は GalNAc と Gal とが結合し、更に Gal 側に Glc と Fuc が結合し、NGNA が GalNAc か Gal に結合しているオリゴ糖である。
GalNAc とは、 N−アセチルガラクトサミンを意味しており、ムチン型糖鎖(O
結合型糖鎖(O-linked sugar chain))として、タンパク質のセリン(Ser)又 はスレオニン(Thr)に結合している。 Gal とはガラクトースを、 Glc とはグル コースを、Fuc とはフコースを意味する。コイグリコホリンには、弱い血液 型活性とともに、コイグリコホリンに結合している糖鎖由来の抗菌活性があ ることが確認されている。5種類の糖類の結合順序、結合位置については、
GalNAc と Gal とが結合し、更に Gal 側に Glc と Fuc が結合している。コイ
グリコホリン由来の抗菌性多糖類は、このうちの Gal→GalNAc→Ser/Thr
によって、タンパク質と結合していることが明らかとなった。その後、 GalNAc
と Gal とが結合し、Gal 側に Glc と Fuc と NGNA とが結合するという構造
を取ることが明らかになった。更に、NGNA は Glc、Fuc と同様に Gal 残基
に結合している。コイグリコホリンの糖鎖とヒトグリコホリンの糖鎖の違い
は、糖鎖を構成する糖の種類の他に、結合するシアル酸がコイは NGNA (N-
グリコリルノイラミン酸)なのに対してヒトは NANA (N-アセチルノイラミ
ン酸)という点である[20]。
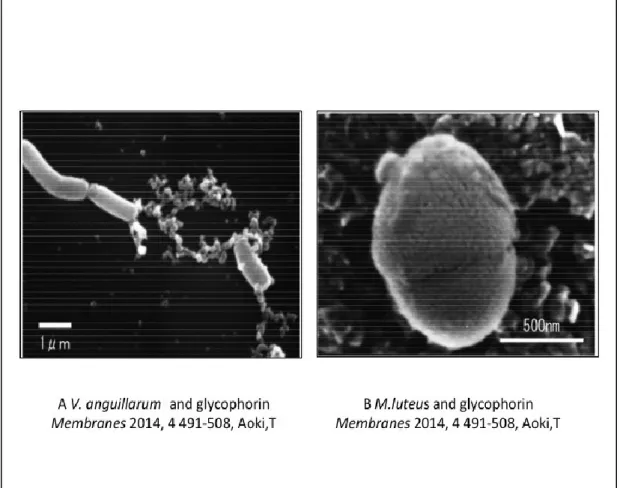
図
1-3.コイグリコホリンと細菌の
SEM画像
A および B は,各々コイグリコホリンと V. anguillarum, M. luteus の SEM
画像である。細菌を含めた全ての生物の生体内には糖鎖と特異的に認識し結
合する性質を持つレクチンが含まれている。レクチンと関連する生理活性と
して,ヒトグリコホリンも血液型活性を持つことの他に、種々のレクチンへ
の結合や、インフルエンザウイルス、センダイウイルスおよびマラリアの受
容体としての生理機能を持つことが知られている。コイグリコホリンが細菌
に対して静菌作用を生じる機構として、グリコホリンの持つ糖鎖が細菌のレ
クチンに結合し、鞭毛、細胞面に吸着し運動阻害をおこすことが原因である
と推測される[10]。
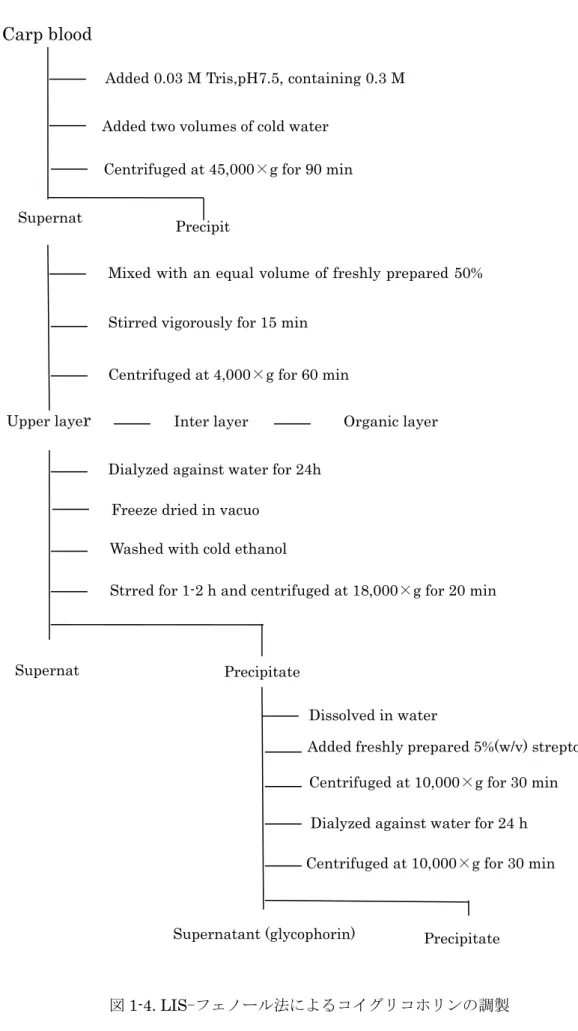
Carp blood
図
1-4. LIS-フェノール法によるコイグリコホリンの調製Added 0.03 M Tris,pH7.5, containing 0.3 M LIS
Added two volumes of cold water
Centrifuged at 45,000×g for 90 min Centrifuged at 45,000×g for 90 min Supernat
ant
Mixed with an equal volume of freshly prepared 50%
phenol
Stirred vigorously for 15 min
Centrifuged at 4,000×g for 60 min
Upper laye
r
Dialyzed against water for 24h
Freeze dried in vacuo
Washed with cold ethanol
Strred for 1-2 h and centrifuged at 18,000×g for 20 min Precipit
ate
Dissolved in water
Added freshly prepared 5%(w/v) streptomycin Centrifuged at 10,000×g for 30 min
Dialyzed against water for 24 h
Centrifuged at 10,000×g for 30 min
Supernatant (glycophorin) Precipitate
Supernat Precipitate
Inter layer Organic layer
第2章 実験
2-1
実験材料及び試薬
供試魚として体重約
3,000 gのコイ(
Cyprinus carpio)を用いた。Ficoll-Paque PLUS は(Pharmacia 製)、クロマトグラフィー用カラムは
DEAE Tyopearl 650M(東ソー株式会社製) 、脱塩用カラムの
Econo-Pac10DGカラム(Bio-Rad 製)、電気泳動用ゲルは
SuoerSepTMAce,10-20%,13well(和光純薬工業社製) を用いた。
CBB染色液は
CBB Stain one Super(ナカライテスク株式会社製)を用いた。銀染色液は Pierce Silver Stain Kit(Thermo Fisher 製)を用いた。
PAS染色液はコールド・シッフ試薬(武藤化学株式会 社製)を用いた。他の試薬は特筆しない限り全て和光純薬工業社製の生化学用または特 級を用いた。各試薬の組成は章末の図表に示す。
2-2 実験方法
2-2-1 コイ赤血球の調製
体長約
30 cmのコイから採血を行った。MS-222(m-aminobezonic acid ethylester
methanesulfonate) 1.9 gを水道水
15 Lに溶かし、薬浴麻酔した後、口腔内の背部動脈 からヘパリン溶液(0.55 %)で湿らせた
20 mlシリンジを用いて血液を採取した[25]。本実 験では以降の実験での使用量、飼育しているコイの数をふまえ, 約
20 mlの血液を採血 した。 得られた血液を直ちに魚類リンゲル液(魚類リンゲル液 A:魚類リンゲル液 B:MilliQ 水=1:1:8)(表
2-1,2,)で等倍に希釈した後、Ficoll-Paque PLUS に重層し、1500×g
で 30 分間、室温で遠心分離を行った。上層と中層に存在する、血漿、白血球
を駒込ピペットで除去し、下層の赤血球のみを採取した。得られた赤血球を魚類リンゲ
ル液に再度懸濁した後、
2500×g で 5 分間遠心分離を行って赤血球を洗浄した。この操作を 3 回繰り返し、コイ赤血球を約
10 ml得た[21]。
2-2-2 コイ赤血球膜の調製
図
2-1に概要を示す。 以下の操作は特記しない限り,全て
4℃で行った。2-2-1 で得ら れた赤血球約
10 mlを
80 mlの氷冷水溶液(表
2-3)に攪拌しながら加えて赤血球の溶血を行った。氷冷水溶液には
Phenylmethylsulfonyl fluoryde (PMSF)を添加した。得られた溶血液を氷冷下で
7分間放置した後、30,000×g で 20 分間遠心分離し、上清を除去 後、 沈殿に
80 mlの
PMSF含有氷冷水溶液を添加して懸濁した。 懸濁液を再度 40,000×g で 20 分間遠心分離した。得られた沈殿に、
2 倍に希釈したBuffer A (表2-4)を約50 ml加え後、
Potter型テフロンホモジナイザーでホモジナイズ(10 往復)した[22]。得られた
懸濁液を 30,000×g で
20 分間 遠心分離した後、沈殿に約30 mlの
Buffer Aを加え再 度ホモジナイズ(20 往復)した。懸濁液 15 ml 毎に
20 mlのショ糖溶液(表
2-6)に重層し、2,500×g, 15 分間遠心分離した。次に、上層および中層を採取し、30,000×g,20 分 間の遠心分離を行った。得られた沈殿から核を除去し、4 ml の
Buffer B(表2-5)に懸濁したものを濃度
1.2 mg/mlの赤血球膜調製液とした。赤血球膜調製液は-80℃ のフリーザ ーで凍結保存した。
2-2-3 コイグリコホリンの調製
図
2-2に概要を示す。凍結保存していたコイ赤血球膜調製液を流水で解凍した後、分取 した
2 ml(タンパク質濃度
1.2 mg/ml)に等量の赤血球膜可溶化試薬(表2-10)を添加してコイ赤血球膜を可溶化した。用いた界面活性剤である
LISはリン酸塩等とキレート を形成して白色沈殿を生じるため,カラムへアプライする前の可溶化試料中の過剰な
LIS除去が不可欠となった。Econo-Pac カラムは従来脱塩カラムであるが,LIS の分子 量が低いことを利用して界面活性剤の除去に応用した 。 可溶化した赤血球膜 (タンパク 質濃度
0.65 mg/ml)を100,000×gで
30分間遠心分離し、上清を
Econo-Pac10 DGカラ ムに負荷して、過剰な
LISの除去を行った。Econo-Pac10DG カラムからの溶出は
Econo-Pac 10 DGカラム平衡化洗浄
Buffer(表
2-7)を1.0 mlずつ負荷して自然落下で 溶出を行い、溶出液は
1.0 mlずつ分画した。得られた画分(FractionNo.4-No.8)毎に、
2.0 M NaCl
を
90 l、10 %コール酸ナトリウム溶液を40 l添加して、終濃度が
130 mM NaClおよび
0.4% コール酸ナトリウムになるように調整した。前段階で得られた画分
のうち、4.5 ml を、あらかじめ
DEAEカラム平衡化洗浄
Buffer(表2-8)で平衡化させた
DEAE Toyopearl 650Mカラムに負荷した。クロマトグラフィーの条件はカラム直径
約
2.0 cm高さ約
3.0 cm、フラクションコレクターは、流速目盛り(30 ml/hr)、分画量(約1.0 ml/tube)で行った。始めに、DEAE
カラム平衡化洗浄
Bufferでカラムの洗浄を行
って不純タンパク質を除去した後、DEAE カラム平衡化洗浄
Bufferと溶出
Buffer(表 2-9)を用いて、直線的濃度勾配溶出法によるコイグリコホリンの溶出を行った。2-2-4
タンパク質の定量
タンパク質の定量は
Proteinassay試薬 (Bio-Rad 製) を用いて行った。標準物質は
BSAを用いた。2-2-2 で調製したコイ赤血球膜
0.1 mlごとに
Proteinassay試薬を蒸留 水で
5倍希釈した溶液を加え発色させた。発色を確認した後、直ちに吸光度
595 nmの 吸光値を測定した。標準物質
BSAの標準曲線から、コイ赤血球膜のタンパク質濃度を 算出した[23]。
2-2-5
タンパク質の染色
SDS-ポリアクリルアミドゲル電気泳動は Laemmli
の方法[24]により行った。泳動槽
用緩衝液はトリス-グリシン溶液(25 mM Tris, 192 mM Glycine, 3.5 mM SDS 含有)
を使用した。電気泳動用試料に等量の試料混合液(50 mM Tris‐HCl 緩衝液, 8% SDS,
40%グリセロール, 320 mM DTT および
0.4 mg/ml BPB含有, pH 6.8)を加え可溶化 し、
100℃で1分間の加熱処理を行った。泳動条件は、ゲル厚 1 mm の
SuperSeoTMAce, 10-20%,13well (和光純薬工業社製)、電流および電圧は30 mA・300 Vとした。タンパ ク質の染色にはクマジーブリリアントブルー R-250 (CBB)および銀染色法、糖タンパ ク質の染色は
PAS染色法で行った。
2-2-5-1 CBB
染色
CBB
染色は電気泳動を行ったポリアクリルアミドゲルを
CBB Stain one Super(ナカライテスク株式会社製)で約
90分間振盪させ、染色を行った。
2-2-5-2 PAS
染色
PAS
染色は電気泳動を行った。ポリアクリルアミドゲルを
12.5 % TCAで
30分間振 盪し、その後純水で振盪した。次に、ゲルを
3.0%酢酸、1.0% 過ヨウ素酸液で
50分間 振盪し、純水で一晩振盪した。次にコールド・シッフ試薬(武藤化学株式会社)で約
2時間振盪して染色を行った。その後、亜硫酸水で
10分間
3回振盪した。最後に純水で 洗浄した。
2-2-5-3
銀染色
銀染色は
Pierce Silver Stain Kitを使用した。電気泳動を行ったポリアクリルアミド ゲルを純水で5分間洗浄した。その後固定液(純水:エタノール:酢酸6:3:1)で ゲルの固定を行い、
10%エタノールで洗浄した。純粋25 mlに
Sensitizer 50 lを加え、
1分間震盪し、純粋で洗浄後、Silver stain 0.5 ml に増感剤の
Enhancer 25 mlを加え
30分間反応させた。その後、5%酢酸で反応を停止させ、純水で洗浄した。
2-2-6
静菌活性の測定
各フラクションでの静菌活性を確認することで、陰イオン交換クロマトグラフィー得 られた各フラクションからコイグリコホリンを検出した。静菌活性はペーパーディスク 法により測定した[19]。作業は全てクリーンベンチ内で行った。供試菌は本研究で培養 した
E-coliを用いた。培地は
HI+BactAgsr (Difco)を用いた。E-coliが含まれる液体培
地
0.2 mlを
45℃の湯浴により溶解させた同培地とアルゴンチューブ内で混和した後、シャーレに均等に散布した。ポジティブコントロールにはアンピシリン(50 l/ml) 、ネ ガティブコントロールにはコイ赤血球膜可溶化溶液、DEAE カラム平衡化洗浄
Buffer(表
2-9)を用いた。静菌活性の測定を行った試料には
Buffer B(表 2-5)に懸濁したコイ赤血球膜(1.2mg/ml)
、可溶化コイ赤血球膜、陰イオン交換クロマトグラフィーで得られた
Fra.No.17、Fra.No.33、Fra.No.47
を用いた。ペーパーディスクに各試料
100 lを添加した。試料 を添加したペーパーディスクは、常温で2分間乾燥させた後、ポジティブコントロール、
ネガティブコントロールとともに乾燥面を下向きにしてシャーレ内に静置した。その後、
30℃で 24 時間インキュベートし、生育阻止円を観察した。
また、ペーパーディスクを用いずに試料を直接、添加し生育阻止円を観察した。試料は
Buffer B(表2-5)に懸濁したコイ赤血球膜(1.2 mg/ml)
、ポジティブコントロールに はアンピシリン、ネガティブコントロールには
Buffer B(表2-5)を用いた。各試料20l
をシャーレ中央に滴下し、20 分間静置し、乾燥を確認した後
30℃で24時間インキ ュベートし観察を行った。
2-2-7 コイグリコホリンからの糖鎖の分離
始めに、過剰な塩を除去するため、得られたコイグリコホリン画分を
Econo-Pac 10DGカラムを用いてコイグリコホリン画分に含まれる、過剰な
NaCl及びコール酸ナトリウ ムを除去した。また、一方はコイグリコホリン画分をセルロースチューブ(Size18,Wako)
に入れた後、脱イオン水
10 Lに対して
24時間の透析を行い、コール酸ナトリウムと
NaClを除去した後、透析内液を
100,000 × gで遠心分離を行った。得られた沈殿を
0.3M LISを加えて溶解した。その後、同条件で透析を行い、過剰な
LISを除去した。過剰な 塩が除去されたコイグリコホリン画分から糖鎖を分離するために脱離を行った[20]。
コイグリコホリン画分
0.4 mlに
5 M NaBH4 (0.5 M NaOH含有) を
0.1 mlを加え
N2ガス存在下の暗室で
48時間のインキュベートを行った[26]。
コイ血液
図
2-1.コイ赤血球膜の調製
Ficoll-Paqueに重層し遠心分離
1500 × g for 30 min赤血球
魚類リンゲル液で洗浄後、氷冷水(表
2-3)で溶血PMSF
を加え、遠心分離
40,000 × g for 20 min min白血球と血漿
上清 上層と沈殿
BufferA(表2-4)に懸濁しホモジナイズ(10
ストローク )
遠心分離
70,000 × g for 20 min上清 沈殿
BufferA(表2-4)に懸濁しホモジナイズ(20
ストローク)
40%ショ糖層(表2-6)に重層し、遠心分離2,500 × g for 15 min
ショ糖層
Overlay and Interphase遠心分離
30,000 × g for 20 min上清 沈殿
BufferB(表2-5)に懸濁しコイ赤血球膜調製液とした
コイ赤血球膜
赤血球膜可溶化溶液で可溶化した。
超遠心分離
30 min at 4℃100,000×g
上清 沈殿
脱塩カラム(Econo-Pac 10DG)での脱塩処理
DEAE Toyopearl 650M columnに付加した。
DEAE
カラム平衡化洗浄
Bufferで平衡化した。
直濃度勾配法でコイグリコホリンを溶出した。
図
2-2.コイ赤血球膜からのグリコホリンの調製
表 2-1「リンゲル液 A」500 ml, pH 7.9 の組成
試薬 mol (最終濃度) 重量
CaCl
2・2H
2O 50 mM 3.67 g
MgSO
4・7H
2O 10 mM 1.23 g
KCl 40 mM 1.49 g
Hepes 100 mM 11.91 g
Glucose 50 mM 4.50 g
超純水
400 mlに上記の試薬を上から順に加え溶解した後、1N NaOH で
pH 7.9を調整
し、500 ml にメスアップした。
表 2-2「リンゲル液 B」500 ml の組成
試薬 mol (最終濃度) 重量
NaCl 1.45 M 42.37 g
超純水
400 mlに
NaClを加え溶解した後、500 ml にメスアップした。
表 2-3 氷冷水」1.2 L, pH 7.7 の組成
試薬 mol (最終濃度) 重容量
CaCl
2・2H
2O 0.88 g
Tris 5 mM 0,73 g
PMSF(stock) 1.0 ml
超純水
1.0 Lに上記の試薬を上から順に加え溶解した後、
conc,HClで
pHを調整し、
1.2 Lにメスアップした。
表 2-4「Buffer A」2.0 L, pH 7.5 の組成
試薬 mol (最終濃度) 重容量
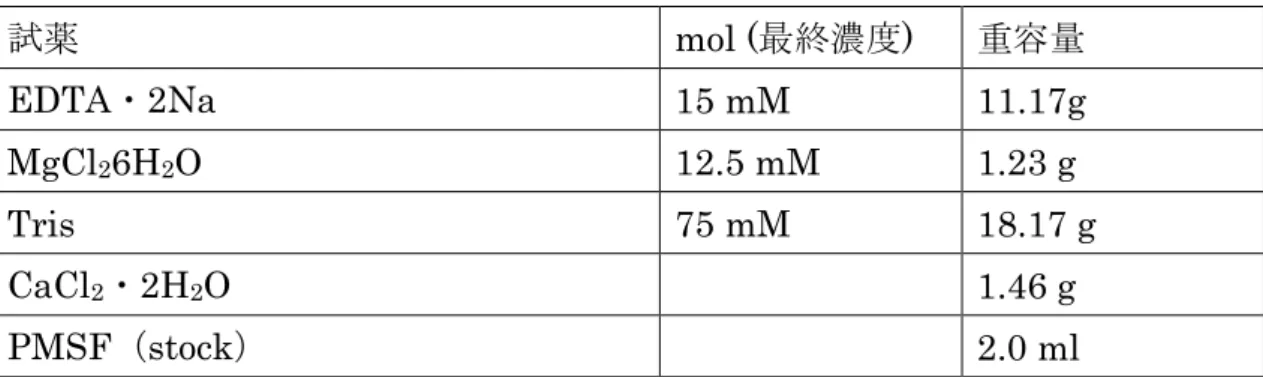
EDTA・2Na 15 mM 11.17g
MgCl
26H
2O 12.5 mM 1.23 g
Tris 75 mM 18.17 g
CaCl
2・2H
2O 1.46 g
PMSF(stock) 2.0 ml
超純水
1.5 Lに上記の試薬を上から順に加え溶解した後、
conc,HClで
pHを調整し、
2.0 Lにメスアップした。
表 2-5「Buffer B」500 ml, pH7.5 の組成
試薬 mol (最終濃度) 重量
Tris 20 mM 0.61g
EDTA・2Na 2 mM 0.372g
超純水
400 mlに上記の試薬を上から順に加え溶解した後、conc,HCl で
pHを調整し、
500 ml
にメスアップした。
表 2-6 「ショ糖溶液」500 ml, pH7.5 の組成
試薬 mol (最終濃度) 重量
Tris 20 mM 0.61g
MgCl
2・6H
2O 1.02 g
ショ糖(サッカロース) 225 g
超純水
400 mlに上記の試薬を上から順に加え溶解した後、conc. HCl で
pHを調整し、
500 ml
にメスアップした。
表 2-7「Econo-Pac 10 DG カラム平衡化洗浄 Buffer」30 ml, pH 7.5 の組成
試薬 mol (最終濃度) 容量
0.2 M Tris-HCl(stock) 50 mM 7.5 ml
0.2 M Tris-HCl (stock) 7.5 ml
を超純水
22.5 mlで希釈した。
表 2-8「DEAE カラム平衡化洗浄 Buffer」100 ml, pH 7.5 の組成
試薬 mol (最終濃度) 重容量
(Stock)1M リン酸カリウム 50 mM 5.0 ml
NaCl 130 mM 0.759 g
Sodium cholate(コール酸ナトリウム) 0.40 % 0.4 g
1
M KPi(stock)5.0 mlを超純水
85 mlに加え、NaCl、Sodium cholate を加えた後、100
mlにメスアップした。
表 2-9「溶出 Buffer」30 ml, pH 7.5 の組成
試薬 mol (最終濃度) 重容量
(Stock) 1M リン酸カリウム 50 mM 1.5 ml
NaCl 0.5 M 0.877 g
Sodium cholate(コール酸ナトリウム ) 0.70 % 0.21 g
1M KPi(stock)1.5 ml
を超純水
20 mlに加え、
NaCl、Sodium cholateを加えた後、
30 mlにメスアップした。
表 2-10「赤血球膜可溶化溶液」2.0 ml, pH 7.5 の組成
試薬 mol(最終濃度) 重容量
(stock) 0.2 M Tris-HCl 40 mM 0.8 ml
LIS 0.3 M 0.475 g
超純水 1.2 ml
試料(コイ赤血球膜) 2.0 ml
第3章 結果と考察
3-1 コイ赤血球膜の調製
本実験で調製したコイ赤血球膜に目的のグリコホリンが含まれ、陰イオン交換クロマト グラフィーで使用可能かを確認することを目的とし、実験を行った。図
3-1の
Aで示し たように、
CBB染色では、本実験で調製した
A-1(コイ赤血球膜)と
A-2(
Membranes 2014, 4 491-508, Aoki,T)による泳動パターンが一致した。また、
PAS染色においても、本実験で調製した
B-1(コイ赤血球膜)と
B-2(
Membranes 2014, 4 491-508, Aoki,T)による泳動パターンが一致した。本実験で調製したコイ赤血球膜では
CBB染色
PAS染色共に分子量約
100 kDaのバンドが検出された。加えて、
グリコホリン以外の赤血球膜構成タンパク質である、アクチン(43 kDa)やアンキリン
(210 kDa)などが検出された。コイグリコホリンは前述したように赤血球膜に存在す る分子量約
100 kDaの糖タンパク質であることから、本実験ではコイ赤血球約
20 mlからグリコホリンが含まれるコイ赤血球膜調製液(タンパク質濃度
1.2 mg/ml)が約7.0 ml調製できた。
また、図
3-2で示したように、界面活性剤(LIS)による可溶化を行ったコイ赤血球 膜においても図
3-1の
A-1, B-1と同様に分子量約
100 kDaのバンドが検出された。こ の結果より、界面活性剤(LIS)による可溶化を行ったコイ赤血球膜においても目的の グリコホリンが含まれると判断し、陰イオン交換クロマトグラフィー供する試料とした。
3-2 陰イオン交換クロマトグラフィー
図
3-3と図
3-4と比較すると、点線で示した吸光度
280 nmの吸光値のパターンが一
致していることがわかった。また、3つ目のピークの吸光値の最大値も
0.40付近と近似
していることがわかった。しかし、2つ目のピークの吸光値では図
3-3では吸光値の最
大が
0.75であるのに対して、図
3-2では最大が
0.20と低い結果になり大きな差が見ら
れた。この結果より、本実験での陰イオン交換クロマトグラフィーによる赤血球膜構成
タンパク質の分画は再現できたと判断した。しかし、目的のグリコホリンの溶出は再考
する必要がある。この原因として使用している吸着単体(DEAE 650M)の劣化などが考え
られる。また、図
3-4ではシアル酸の定量による観点でのコイグリコホリンの分画を評
価しているが、各
Fractionと各ピークに溶出されたタンパク質の同定はなされていない。
そこで、本実験では陰イオン交換クロマトグラフィーで得られた各
Fractionで電気泳動 と銀染色を行うことで各ピークに溶出されたタンパク質の検討を行った。
3-3 各Fraction
の銀染色によるタンパク質の検出
始め、陰イオン交換クロマトグラフィーで得られた各
Fractionからのタンパク質の検 出は
CBB染色と
PAS染色により行ったが、各
Fractionに含まれるタンパク質が陰イオ ン交換クロマトグラフィーの過程で希釈されてしまい、検出可能な濃度を下回る結果と なった。そこで、より感度の高い銀染色法での確認を行った。図
3-5はその結果を示し ている。Lane 2 ,3(Fraction.No.17,18)は、カラム非吸着タンパク質であり、Lane 全体 にバンドが検出され、低分子量の
20 kDa、10 kDaにもバンドが強く検出された。
Lane 7, 8, 9, 10, 11(Fraction.No.45-No.49)では、分子量約90 kDaのバンドの他に
37-57 kDaの付近にバンドが検出され、赤血球膜構成タンパク質のアクチンが多く溶出されたと考 えられる。 塩濃度
0.20 mM付近で溶出された
Lane 4, 5, 6(Fraction.No. 31, 33, 35)では、分子量約
100 kDaの付近にバンドが強く検出され、他の分子量でのバンドは弱く検出さ
れた。コイグリコホリンの分子量が約
100 kDaであることから、夾雑タンパク質が除去 さ れ 、 目 的 と す る コ イ グ リ コ ホ リ ン を 分 画 で き た と 推 測 さ れ る 。 よ っ て 、
Fraction.No.31-No.35をグリコホリン画分とみなした。
Fraction.No.45-No.49で検出さ
れた
90 kDa-100 kDa付近のバンドは塩濃度
0.20 mMで溶出されなかったコイグリコホ
リン、もしくは近い分子量の赤血球膜構成タンパク質のバンド3である可能性が考えら れる。
3-4 各Fraction
の静菌活性
図
3-6は各
Fractionの静菌活性示している。
A-1コイ赤血球膜懸濁液(タンパク質
1.2mg/ml)
、B-1 可溶化後のコイ赤血球膜(タンパク質
0.3 mg/ml)では静菌活性の結果となる生育阻止円を観察することができなかった。また、ネガティブコントロールで使用し た赤血球膜可溶化溶液(表 2-10)で生育阻止円が観察された。陰イオン交換クロマトグ ラフィーで得られた各
Fractionにおいても生育阻止円を観察することができなかった。
しかし、以前の研究成果ではコイグリコホリンの静菌活性は確認されている[8] 。その
際はコイグリコホリン
15 μg/diskの条件で本実験と同様の試験を行っている。しかし、
図
3-7で示したように、ペーパーディスクを使用せず直接シャーレ内にコイ赤血球膜を
20 μl
添加したところ、生育阻止円が観察され、
E.colの生育阻害が認められた。図
3-6で静菌活性が確認されなかった原因は、試料中に含まれるコイグリコホリンの濃度が
15 μgより低いことに加え、本実験で使用したコイグリコホリンは陰イオン交換クロマトグ ラフィーにより調製したコイグリコホリンであるため、LIS-フェノール法[13]により 調製されたコイグリコホリンとは異なり、タンパク質部の変性を受けていないことが考 えられる。陰イオン交換クロマトグラフィーではコイグリコホリンの疎水部が保持され た状態で分画されるため、凝集が生じやすい。また、コイ赤血球膜が可溶化した状態で あってもコイグリコホリンの糖鎖部がタンパク質部に取り込まれているために、静菌活 性を有した糖鎖部がタンパク質から分離しない。その結果、
E.coliに糖鎖が作用しない ために抗菌性を示す生育阻止円が観察されなかったと判断した。以上のことから、コイ グリコホリンの静菌活性を確認するためには試料に含まれるコイグリコホリンの濃度を
15 μg /disk より高くし、コイグリコホリンの糖鎖部分のみを用いる必要があると考えられる。
3-5 -脱離による糖鎖分離
当初は、グリコホリン調製で使用した、過剰な塩と界面活性剤(LIS)が-脱離によ
る反応を阻害していると推定していた。そこで、Econo-Pac10DG カラムおよび透析に
より過剰な塩と界面活性剤(LIS)の除去をした後、-脱離反応を行ったが、目視によ
るグリコホリンからのタンパク質部分の沈殿は確認できなかった。このことから、コー
ル酸ナトリウム及び
LISを用いて可溶化した試料を用いた場合のいずれもがタンパク質
からの糖鎖の分離反応が不十分である事が判明した。しかし、-脱離反応を行った後の
試料で銀染色を行った結果、100 kDa 付近にバンドが検出されず、50 kDa-10 kDa の範
囲に広くバンドが検出された(図
3-5)。この結果から、不十分ではあるが、-脱離反応
がコイグリコホリンに対し作用している可能性が示唆された。よって、陰イオン交換ク
ロマトグラフィーで
0.2 M NaCl付近で溶出された、グリコホリンは-脱離による糖鎖
の分離用の試料として充分使えることが明らかになった。
図
3-1.コイ赤血球膜の
CBB染色および
PAS染色
A;CBB染色,B; PAS 染色
A はコイ赤血球膜の CBB 染色の結果であり、 A-1 は本実験で調製した赤血
球膜(20 μl/lane)、A-2(Membranes 2014, 4 491-508, Aoki,T )である。B
はコイ赤血球膜の PAS 染色の結果であり、B-1 は本実験で調製した赤血球膜
(20 μl/lane)、B-2(Membranes 2014, 4 491-508, Aoki,T )である。分子量
マーカーは CBB 染色では、PAS 染色ではを使用した。図中の矢印はコイグ
リコホリンの分子量である 100 kDa 付近に検出されたバンドを示している。
図3-2. 可溶化およびカラム負荷試料の
CBB染色および
PAS染色
A-1 は界面活性剤(LIS)による可溶化を行ったコイ赤血球膜、A-2 は
Econo-Pac カラムにより、過剰な界面活性剤(LIS)の除去を行ったコイ赤血球
膜の CBB 染色の結果である。B-1 は界面活性剤(LIS)による可溶化を行っ
たコイ赤血球膜、B-2 は Econo-Pac カラムにより、過剰な界面活性剤(LIS)
の除去を行ったコイ赤血球膜の PAS 染色の結果である。図中の矢印はイグリ
コホリンの分子量である 100 kDa 付近に検出されたバンドを示している。
図
3−3.陰イオン交換クロマトグラフィーのクロマトグラム
グラフは吸光度 280 nm で吸光値を測定したクロマトグラムである。
Fra.No.6 から試料の負荷を始めた。 Fraction.No.16-No24 に最初のピークが
見られた。このピークは、DEAE カラムに吸着しなかったタンパク質だと考
えられる。Fraction.No.25 から NaCl による直濃度勾配法による溶出を行っ
た。濃度勾配を開始した直後、NaCl 濃度 0.2 M 付近で2つ目のピークが見
られた。その後、NaCl 濃度 0.5 M まで溶出を続けると NaCl 濃度 0.25 M 付
近で 3 つ目のピークが見られた。
図
3-4.陰イオン交換クロマトグラフィーのクロマトグラム[12]
0.000 0.200 0.400 0.600 0.800 1.000
2 8 14 20 26 32 38 44 50 56 62 68 74 80 Tube No. (1.0 ml/tube)
Absorbance at 280 nm, 630 nm
0.500 0.900 1.300 1.700 2.100
Absorbance at 490 nm
280 nm 630 nm 490 nm
図 3-4 は、本実験と同様の手順で調整したコイ赤血球膜を負荷した陰イオン 交換クロマトグラフィーのクロマトグラムである[17]。グラフ左の縦軸は吸 光度 280 nm および 630 nm の吸光値、右の縦軸は吸光度 490 nm の吸光値 である。吸光度 280 nm はタンパク質の吸光値を示している[27]。中性糖の測 定は、フェノール硫酸法[28]を用いて呈色させ、反応後、 490 nm の吸光度を 測定している。標準物質は D-グルコースを用いている。シアル酸の定量は、
Jourdian らの過ヨウ素酸-レソルシノール法[29]を用いて呈色を行い、反応
後、 630 nm の吸光度を測定している。標準物質として NANA (N-アセチル
ノイラミン酸)を用いている。グリコホリンに含まれるシアル酸は赤血球膜
中に存在する全シアル酸量の約7割を占めるため、Fraction.No.32-No.50 を
グリコホリン画分としている。
図
3-5.各
Fractionの銀染色の結果
図 3-5 は、陰イオン交換クロマトグラフィーで得られた各 Fracion の銀染 色の結果である。Lane 1 ,2 ,3, 12, 13 は SDS 用サンプル Buffer で各画分と も 20 倍希釈した試料を泳動した。それ以外の画分は全て、SDS 用サンプル Buffer で 4 倍希釈した試料を泳動した。 Lane 1 には分子量マーカー (MWM)
を泳動した。Lane 2 , 3 は陰イオン交換クロマトグラフィーで DEAE に吸着 しなかったパススルーの Fraction である。 Lane 4, 5 ,6 では分子量約 100kDa 付近に強くバンドが検出された。Lane 7,8,9,10,11 では、分子量約 90kDa の バンドが検出された他、37-57kDa の付近にバンドが検出された。Lane 12 は本実験で調整した、コイ赤血球膜を界面活性剤(LIS)により可溶化した試 料である。 Lane 13 は陰イオン交換カラムクロマトグラフィーで負荷 した試料
である。
図
3-6.各フラクションの静菌活性の確認
図 3-6 はペーパーディスク法での静菌活性の確認試験の結果を示している。
A-1 はコイ赤血球膜懸濁液(タンパク質 1.2 mg/ml) 、図 3-6- B-1 は可溶化コ
イ赤血球膜溶液(タンパク質 0.3mg/ml)、 A-2、 B-2 はポジティブコントロール
(アンピシリン 50 l/ml)、 A-3 および B-3 はネガティブコントロール(赤血球
膜可溶化用溶液)である(100 l/ディスク)。図 3-6 の C-1、 D-1 および E-1
は陰イオン交換クロマトグラフィーで得られたピークの各々Fra.No.17,
Fra.No.33 および Fra.No.47 である。 C-2、 D-2 および E-2 はポジティブコン
トロール(アンピシリン 50 l/ml)、A-3、B-3 はネガティブコントロール(赤
血球膜可溶化用溶液)である(100 l/ディスク) 。
図
3-7.各フラクションの静菌活性の確認
図 3-7 は図 3-6 とは異なり、各試料をペーパーディスクを使用せずに培地 に直接添加した静菌活性の確認の結果を示している。 A はポジティブコント ロール(アンピシリン 50 l/ml)、B はコイ赤血球膜懸濁液(タンパク質 1.2 mg/ml) 、C はネガティブコントロール(20 mM Tris-HCl)である。各試料
は 20l シャーレ中央に添加している。 B’と C’は肉眼での結果となる生育阻
止円が確認できたため、それぞれを拡大した写真である。B’の黒枠は、試料
の添加場所と確認された生育阻止円の部分である。
図
3-8. -脱離反応の結果A は-脱離反応の結果の結果を示す、銀染色を行った電気泳動写真である、
Lane1 は分子量マーカー(MWM) 、 Lane2, 3 は-脱離反応を行ったコイグリ
コホリン試料、Lane4 は本実験の陰イオン交換クロマトグラフィーで得られ
たコイグリコホリン(Fraction.No.33)である。下の表はそれぞれの Lane
に対応している。B はコイグリコホリンの構造式であり、-脱離反応の作用
を示している。
第
4章 総括
本論文ではコイ血液から赤血球膜を調製し、陰イオン交換クロマトグラフィー[13]を 用いて調製されたコイグリコホリンの静菌活性を確認し、脱離による糖鎖の分離が可 能であるかの検証を目的として実験を行った。検証の結果、陰イオン交換クロマトグラ フィーを用いて調製されたコイグリコホリンには充分な静菌活性がないことが確認され た。また、コイグリコホリンからの糖鎖の分離反応は不十分ではあるが、作用している ことが判明し、本実験で調製されたコイグリコホリンは脱離用の試料として使用可能 であることが明らかになった。従来使用されていた
LIS-フェノール法[16]により調製されたグリコホリンでは、-脱離による遊離糖鎖の調製は充分可能であった[10]。よ って、グリコホリンから効率よく糖鎖を分離するためには、フェノールによるグリコホ リンのタンパク質部分の変性操作が必要不可欠であることが推測できる。今後はフェノ ール等の弱酸性のタンパク質変性剤及びアルカリ性のタンパク質変性剤を使用し、今回 の実験手順にタンパク質変性の操作を加えた糖鎖の分離方法を検討したい。また、コイ グリコホリンの調製においても本実験の結果は不安定であり、大量の試料を用いていな いことから、コイグリコホリンを効率よく調製できたとは言い難い。今後、試料および カラムのスケールを拡大して実験を行い、大量調製における問題点などを検討する必要 がある。加えて、-脱離反応以外での糖鎖の分離方法を検討する必要がある。例として 脱糖鎖酵素による切断が考えられるが、コイグリコホリンの糖鎖は
O-型糖鎖であり、コアがシアル酸で修飾されているため、O-グリカナーゼが阻害されると予想されため
G
al-GalNAc-O-Ser/Thrの状態にまでにする必要があると考えている。
第5章 地域イノベーションに関しての総括
当研究室が実験試料としているコイは昔から日本や中国では、滋養効果を持つ食物と して重宝されてきた。コイの煮汁にはビタミン
B2, B6および B12 が豊富に含まれてお り、血液循環や肝機能の改善といった効果があるとされている。日本では薬膳料理とし て親しまれている。
当研究室では上記で示したコイグリコホリンの種々の細菌に対する抗菌活性を利用し、
魚類養殖や養殖環境の改善に関して何らかの応用ができるのではないかと考えている。
日本の養殖業は、急速な発展をしてきたが、ビブリオ病などの様々な疾病と戦いなが らなされてきた。原因不明の魚病の発生や海外からの新しい疾病の侵入は養殖業の発展 を妨げる大きな阻害要因と認識されている。かつては海面養殖業において、連鎖球菌症 や類結節症など細菌感染症がまん延し、治療のための抗菌性薬剤が多く使用された。し かし、耐性菌の出現や使用薬剤の魚体への残留に対する消費者の懸念などから国単位で の改善が行われ、ワクチンの開発により、薬剤の使用量は減少した。しかし、水産用ワ クチンは種類の不足、抗菌剤が幅広い細菌に対して有効であることに対し、ワクチンは 原則1つの疾病にしか有効ではないなどの問題がある。魚病対策はサルファ剤系、キノ ロン系などの抗菌剤による化学療法とワクチン治療の両方依存しているのが現状である。
水産用抗菌剤は各都道府県の指導のもと適正使用が推進されてきたが、完全な履行は なされていない。耐性菌の出現などの問題から、平成30年から水産用抗菌剤の購入と 使用の際、魚類防疫員や獣医師などの専門家の指導を必要とする取り組みが新たに導入 されることになり、水産用抗菌剤に対する意識は高まりつつある。
上記の養殖業の現状を改善するため、本研究室ではグリコホリンを用いた製品の開発 を考えている。例として、グリコホリンを結合したゲル単体を充填したカラムを作り、
カラム内を通過した水の水質を浄化する水質浄化装置などを考えている。しかし原料と
なるコイの血液の入手手段が確立していないことや、コイグリコホリンの調製が安定的
ではなく、調製量のスケールが小さいことなど解決すべき課題が存在する。
参考文献
1. Fairbanks,G.,Steck,Theodorel.&Wallach,D.F.H.,Biochemistry,10,2606-2 617(1971).
2. Steck,T.L.,Cell Biology, 62,1-19,(1974).
3. Anstee,D.J.,Parsons, S.F.,Ridwell, K.,Tanner, M.J.A., Merry, A.H., Thomson, E.E., Judson, P.A., Johnson, P., Bates, S.and Fraser, I.D., Biocheminal Journal, 218, 615-619(1984).
4. Dahr, W., Uhlenbruck, G., Janssen, E. and Schmalisch, R., Blut, 32, 171-184(1976).
5. Segrest, J.P., Kahane, J., Jackson, R.L. and Marchesi, V.TC Arch.Biochem. Biophys., 155, 167-183,(1973).
6. Jackson, R.L., Segrest, J.P., Kahane, J. & Marchesi, V.T., Biochemistry .,12, 3131-3138(1973).
7.
富田基郎, 赤血球膜糖蛋白質の構造と機能, 生物物理, 17, 4(1997).
8. M.J.A. Tanner and D. J. Anstee., Biocheminal Journal., 153, 271(1976)
9.
中尾暢宏, 魚類赤血球膜中の糖タンパク質について, 修士論文,三重大学,
三重,(1997).
10. Aoki, T., Chimura, K., Nakano, N. and Mizuno., Y., Membranes, 4, 491-508(2014).
11. Aoki, T., Inoue, T., Journal of Fish Diseases., 573-576(2011).
12. Michel, F. & Rudloff, V. Eur. J. Biochem.,181, 181-187(1989).
13.
深井正夫, 魚類赤血球膜中の糖タンパク質について, 修士論文,三重大学,
三重(1994).
14. Hamaguchi., H. and Cleve , H. 271 -279(1972).
15. Olga, O, B. and Bina,Zvilichovsky.., Methods in Enzymology., 245 -252(1972).
16. Marchesi, V.T., Method in Enzymology, 252-254(1972).
17.
藤原里恵, コイ赤血球膜の糖タンパク質に関する研究, 卒業論文, 三重大学, 三重(2008).
18. Aoki, T., Fukai, M. and Ueno, R., Fisheries Sci, 62, 498-499(1996).
19.
照屋忠敬, ディスク法による薬剤感受性試験に及ぼす測定用培地の種類, 沖縄県水産試験場事業報告書, 1982, 201-223(1983).
20. Aoki, T., Chimura, K., Suguira, H. and Mizuno., Y., Membranes, 4, 764-777(2014).
21. Aoki, T., Fukai, M., Ueno, R., Fisheries Sci, 62, 498-499(1996).
22. Michel, F., Rudloff, V., Biochem, 181, 181-187(1989).
23. Bradford, M.M., Biochem, 72, 248(1976).
24. Laemmli, U.K., Nature, 227, 680-685(1970).
25.
岡崎太郎. 「続生化学実験講座, 8, 血液(上) 」 (日本生化学会編)東京化学 同人,東京. 213-227(1987).
26. Carlson, D.M., J.Boil.Chem, 243, 616-626(1968).
27. Lowry , O.H. , Rosebrough , N.J. , Farr , A.L. and Randall , R.J., J.Biol.
Chem. 93, 26(1951).
28. Dubois, M., Gilles, A.K., Hamilton, K.J., Rebers, A.P. and Smith, F., Analytical Chemistry, 28, 350-356(1956).
29. Jourdain, G.W., Dean, L., and Roseman, S., J.Boil.Chem, 246-430(1971).