家畜衛生学雑誌42-2表14.ai
K
ISSN 1347−6602 昭和62年6月9日学術刊行物認可
家畜衛生学雑誌 家畜衛生学雑誌
Vol.42 No.2 2016. JUL.
日 本 家 畜 衛 生 学 会
The Japanese Society of Animal Hygiene
The Japanese Journal of Animal Hygiene
家畜衛生学雑誌 第
42巻第
2号
二〇一六年七月
日本家畜衛生学会
日本家畜衛生学会第84回大会 要旨集
複写される方へ
日本家畜衛生学会は有限責任中間法人 学術著作権協会(学著協)に複写に関する権利委託をしていますので,本誌に掲 載された著作物を複写したい方は,学著協より許諾を受けて複写して下さい.但し,社団法人日本複写権センター(学著協 より複写に関する権利を再委託)と包括複写許諾契約を締結されている企業の社員による社内利用目的の複写はその必要は ありません.(※社外頒布用の複写は許諾が必要です.)
権利委託先: 有限責任中間法人 学術著作権協会
〒107−0052 東京都港区赤坂9−6−41 乃木坂ビル3階
電話:03−3475−5618 FAX:03−3475−5619 E-mail:info@jaacc. jp
注意:複写以外の許諾(著作物の転載・翻訳等)は,学著協では扱っていませんので,直接日本家畜衛生学会へご連絡下 さい.[電話:042−367−5780]
また,アメリカ合衆国において本書を複写したい場合は,次の団体に連絡して下さい.
Copyright Clearance Center, Inc.
222 Rosewood Drive, Danvers, MA 01923 USA
Phone:1−978−750−8400 FAX:1−978−646−8600
家 畜 衛 生 学 雑 誌
日本家畜衛生学会 発行
President : Junsuke SHIRAI( )
Vice President : Shigeru MIYAZAKI ( )
Editor-in-Chief : Shigeru MIYAZAKI( )
Editorial Board : Norihide KAKIICHI( )
Shinji TAKAI( )
Hajime NAGAHATA( )
Sadao NOGAMI( )
Tadao WATANABE ( )
"""""""""""""""""""""""""""""""""""""""""""
The Japanese Journal of Animal Hygiene
Published by the Japanese Society of Animal Hygiene
理 事 長 :白井淳資 副理事長 :宮﨑 茂 編集委員長 :宮﨑 茂
編集委員 :柿市徳英・高井伸二・永幡 肇
野上貞雄・渡邊忠男
会員の皆様におかれましては,ますますご清栄のこととお慶び申し上げます.ここに,「家畜衛生学雑誌」第42巻 第 2 号を刊行する運びとなりました.
本号では,総説 1 編と第84回大会の講演要旨を掲載しています.
第84回大会では,教育講演 1 題,一般演題 8 題のご発表のほか,平成27年度家畜衛生学雑誌論文賞受賞講演もござ いますので,ご参加の皆様の積極的なご討論をお願いいたします.また,「家畜衛生フォーラム2016」の開催を予定 している12月 2 日の午前中には第85回大会を開催いたしますので,こちらでの一般講演発表についても積極的なエン トリーをお願いいたします.
なお,本号には総説 1 編も掲載しております.家畜衛生学雑誌では,原著論文・短報以外にも,総説,数ページ程 度のミニレビュー,技術資料等の原稿を受け付けておりますので,会員の皆様の積極的なご投稿をよろしくお願い致 します.ご不明な点は遠慮なく編集委員会事務局へお問い合わせください.
日本家畜衛生学会理事長 白井淳資 家畜衛生学雑誌編集委員長 宮﨑 茂
(日本家畜衛生学会副理事長)
日本家畜衛生学会・学会費納入のお願い
ご承知のように,学会は会員の皆様からの会費をもって運営されています.学会の運営を円滑に運ぶために,所定 の会費を納入していただきますようお願いします.
*会費は,正会員5,000円,学生会員2,000円です.
*平成27年度までの未納分をお支払いいただく場合,正会員年会費は4,000円です.
日本家畜衛生学会 理事長 白井淳資
00
口座記号 口座番号(右詰めで記入) 金額料金 特殊
取扱
千 百 十 万 千 百 十 円
※
※
※
※
加入者名通信欄ご依頼人 受付局日附印
※
※
(電話番号 − − ) おところ(郵便番号 − )
※ おなまえ
払 込 取 扱 票
口座記号番号加入者名金額料金特殊取扱ご依頼人
※
受 付 局 日 附 印
※
※
※
※
※
円
千 百 十 万 千 百 十 円
おなまえ
郵便振替払込請求書兼受領証
記載事項を訂正した場合は︑その箇所に訂正印を押してください︒切り取らないで郵便局にお出しください︒
各票の※印欄は︑ご依頼人において記載してください︒
裏面の注意事項をお読みください。
これより下部には何も記入しないでください。
(消費税込み)
様
様
日本家畜衛生学会
0 0 2 4 0
4 3 1 7 1 3 0 0 2 4 0 3 4 3 1 7 1
日本家畜衛生学会
平成 24 25 26 27 28 年度
( ) 計 円
ので、口座番号及び金額を記入す る際は、枠内にはっきりと記入し てください。
また、本票を汚したり、折り曲 げたりしないでください。
・この払込請求書を郵便局の派遣 員にお預けになるときは、引換え に預り証を必ずお受け取りください。
この受領証は、郵便振替の払込 みの証拠となるものですから大切
に保存してください。 この払込取扱票の裏面には、何も記載しないでください。
家畜衛生学雑誌
第42巻 第 2 号 2 0 1 6
目 次
〈総 説〉
豚胸膜肺炎の起因菌Actinobacillus pleuropneumoniaeの生物型,血清型及び遺伝子型について
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・伊藤博哉 ・・・・・・・・・・・・・・・・・・・・・・・・・ 29〜60
〈第84回大会一般講演要旨〉
焼成カルシウムと次亜塩素酸ナトリウムの殺菌作用
・・・・・・・・・・・・・・・・・・・・・・・・・・豊福千遙・山田匡之・古村みゆき・鈴木真結子・Hakim Hakimullah
Sangsriratanakul Natthanan・Alam Md. Shahin・Shoham Dany・竹原一明 ・・・・・・・・・・・・・・・・・・・・・・・・・ 64〜65 Study of virus and bacteria inactivation using various sources of fresh charcoal ash in Thailand
・・・・・・・・・・・・・・・・・・・・・・・・・Kittima Lorcharoenrungroj・Sakchai Ruenphet・Kazuaki Takehara ・・・・・・・・・・・・・・・・・・・・・・・・・ 66〜67 Quantitative risk assessment of the introduction of rabies into Japan through
the importation of dogs and cats worldwide
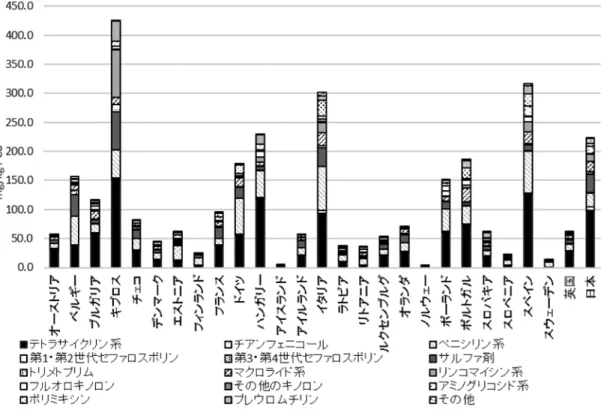
・・・・・・・・・・・・Katsuaki Sugiura・Nigel C.L. Kwan・Y. Hosoi・Akio Yamada・Emma Snary ・・・・・・・・・・・・・・・・・・・・・・・・・ 68〜69 日本の家畜の抗菌剤使用量の欧州との比較
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・松田真理・Nigel Kwan・杉浦勝明 ・・・・・・・・・・・・・・・・・・・・・・・・・ 70〜71 犬の膀胱炎および抗生剤使用に関する調査
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・井上 舞・杉浦勝明 ・・・・・・・・・・・・・・・・・・・・・・・・・ 72〜73 MALDI Biotyperによる動物由来細菌株の遺伝子型・血清型鑑別
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・村田 亮・井上正亮・岡 良彦・菊池直哉 ・・・・・・・・・・・・・・・・・・・・・・・・・ 74〜75 輸入初生ひなから分離された鶏大腸菌症由来E.coliの性状調査
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・下平久美子・小菅千恵子・長谷川未央・楠本正博 ・・・・・・・・・・・・・・・・・・・・・・・・・ 76〜77 馬の敗血症の原因菌Actinobacillus genomospecies 2の日本における初分離及び
16S rDNAとhousekeeping geneの遺伝子解析による同定
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・伊藤博哉・村上美雪・下西儀政・帆保誠二・丹羽秀和 ・・・・・・・・・・・・・・・・・・・・・・・・・ 78〜79
〈平成27年度家畜衛生学雑誌論文賞授賞講演要旨〉
「下痢を呈した豚群におけるデルタコロナウイルス遺伝子の検出と宮城県内浸潤調査」の 平成27年度家畜衛生学雑誌論文賞受賞にあたって
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・小寺 文・曽地雄一郎・西 清志・千葉直幸
植木 洋・矢島りさ・竹田百合子・結城瑞希 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 81
〈第84回大会教育講演要旨〉
ジビエに関する食肉衛生 ─現場からの提言─
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・小谷浩治 ・・・・・・・・・・・・・・・・・・・・・・・・・ 83〜85 会員へのおしらせ ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 87 家畜衛生学雑誌投稿規程 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 88〜89 日本家畜衛生学会会則 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 90〜91
家 畜 衛 生 学 雑 誌
The Japanese Journal of Animal Hygiene Vol. 42 No. 2 2 0 1 6
Contents
〈Review〉
Actinobacillus pleuropneumoniae biotypes, serotypes and genotypes
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・Hiroya Ito ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 29〜60
〈Abstracts of oral presentations on 84th academic meeting〉 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 61〜79
〈Abstract of JJAH award lecture〉 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 81
〈Abstract of the educational lecture on 84th academic meeting〉・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 83〜85
Information for Members ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 87 Instruction for Authors ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 88〜89 The Regulations of The Japanese Society of Animal Hygiene ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 90〜91
Jpn. J. Anim. Hyg.
1 .豚胸膜肺炎及び豚胸膜肺炎菌について 豚胸膜肺炎は世界各国で多発し,養豚産業に多大な経 済的損失を与える豚の重要な呼吸器系疾病である45).本 症の主要な臨床症状は,急性の場合は食欲不振,沈鬱,
発熱,発咳,呼吸促拍,呼吸困難であるが,剖検後,初 めて本病と診断される場合も多い45).飼料効率及び一日 平均増体量の低下等を示す慢性症例も多く,これらでは 食肉衛生検査時に肺と壁側胸膜の癒着,肺における膿瘍 及び結節等の病変が観察されることが多い45).
豚 胸 膜 肺 炎 の 起 因 菌 で あ る Actinobacillus pleuropneumoniae(App)は,莢膜を保有するグラム陰 性の小桿菌であるが,しばしば多形性を示す.ウレアー
ゼ(Actinobacillus 属以外の菌種との鑑別時に重要な性 状),マンニットの分解(Actinobacillus suisとの鑑別時 に重要な性状),CAMPテスト,血液寒天上での溶血性 は,いずれも陽性である68,73).
2 .生物型について 2.1.はじめに
Appは,発育の際のニコチンアミド・アデニン・ジヌ クレオチド(Nicotinamide adenine dinucleotide)(NAD)
要 求 性 の 有 無 に 基 づ き, 2 つ の 生 物 型 に 型 別 さ れ
る68,73,166).NAD 要 求 性 の 生 物 型 1 は, か つ て は
Haemophilus pleuropneumoniae(その前はHaemophilus parahaemolyticus),一方,NAD非要求性の生物型 2 は,
かつては Pasteurella haemolytica-like organism に分類 され別菌種とされていた.しかし,DNA 相同性試験及 び生化学的性状試験の成績から,共に Actinobacillus 属 に移行され,同一菌種の App に再分類された166).この
豚胸膜肺炎の起因菌Actinobacillus pleuropneumoniaeの生物型,
血清型及び遺伝子型について
伊 藤 博 哉*
Actinobacillus pleuropneumoniae biotypes, serotypes and genotypes
Hiroya ITO *
(*National Institute of Animal Health, NARO, 3-1-5 Kannondai, Tsukuba, Ibaraki 305-0856, Japan)
(2016. 4. 22 受付/2016. 6. 6 受理)
Summary
Actinobacillus pleuropneumoniae, a Gram-negative bacterium belonging to the Pasteurellaceae family, is a causative agent of swine pleuropneumonia, a highly contagious repiratory disease that causes important economic losses to the swine industry worldwide. To date, 2 biotypes and 16 serotypes are recognized in A.
pleuropneumoniae. Serotyping is of major interest for A. pleuropneumoniae due to different virulence potential among serotypes and serotype-specific protection with bacterins. Serotype prediction methods based on genotyping have been developed lately and can be used as diagnostic tools by veterinary diagnostic laboratories.
Here, I review current knowledge on A. pleuropneumoniae biotypes, serotypes and genotypes.
Key words: Swine pleuropneumonia, Actinobacillus pleuropneumoniae, biotypes, serotypes, genotypes
家畜衛生学雑誌 42,29〜60(2016)
* 国立研究開発法人農業・食品産業技術総合研究機構 動物 衛生研究部門
〒305-0856 茨城県つくば市観音台3-1-5 連絡著者:伊藤博哉([email protected])
ために,現在でもAppのことを「Haemophilus ヘモフィ ルス」の最初の 2 文字である「ヘモ」と呼ぶ人も多い.
2.2.生物型 1 と 2 の分離頻度について
App生物型 1 による豚胸膜肺炎は頻発し,アウトブレ イク例も多いが,App生物型 2 による豚胸膜肺炎の発生 は少なく,散発例が多い.App生物型 2 の分離例は世界 的にも少ないが72−74)ドイツ35),ハンガリー35),ベルギー94), オランダ94),デンマーク151)及びスペイン120−122)等の欧 州や米国37)等での分離報告がある.日本では,筆者の 知る限り,生物型 2 の分離報告は 2 例のみである81,135). 2.3.生物型 1 及び 2 の病原性について
実験感染の成績に基づき,生物型 2 は生物型 1 よりも 病原性が弱いと考えられてきた31,85).しかし,胸膜肺炎 症例から生物型 2 が純培養的に分離された報告例がいく
つかあり37,120−122),米国で分離された生物型 2 を用いた
実験感染で,胸膜肺炎を再現させた報告もある37).さら に 7 年にわたる調査により,スペインでは App 生物型 2 が豚胸膜肺炎に強く関与している可能性が示唆されて いる.すなわち,典型的な急性胸膜肺炎を呈した豚の肺 から App 生物型 2 がほとんどは純培養状あるいは優勢 菌として分離され,それらの割合は,8.9〜39.4%(平均 25.3%)を占め比較的高率であった122).
2.4. パスツレラ科細菌のNAD合成経路とApp生物型 1 のNAD要求性の遺伝学的背景について
パスツレラ科細菌のNADは,以下の経路で合成され ると考えられている124).(ⅰ)ニコチンアミドと 5 -リン 酸リボシルピロリン酸から,ニコチンアミド・リン酸転 移酵素(NAmPRTase)によって,ニコチンアミド・モ ノヌクレオチド(NMN)が合成される.(ⅱ)次いでニ コチンアミド・アデニル転移酵素によって,NMNから NADが合成される.
なぜApp生物型 1 はNAD要求性であるかを明らかに するための実験が行われている.すなわち,NAD 非要 求 性 の Haemophilus ducreyi の NAmPRTase(495ア ミ ノ酸(aa))をコードする nadV 遺伝子を App 生物型 1
(血清型 1 )に形質転換し,発育にNADが不要になるか 検討された124).その結果,形質転換体は発育にNADを 添加する必要がなくなり,nadV 遺伝子の欠損あるいは 機能不全がApp生物型 1 のNAD要求性の理由であると 考えられた124).その後,生物型 1 (血清型 3 )は495 aa のNAmPRTaseをコードするnadV遺伝子ではなく,ア ミノ酸長が1/2以下と短く,酵素として機能しなくなっ たと思われる偽遺伝子 nadV’(203aa のタンパクをコー
ド)を保有することが明らかにされ,この事が生物型 1 のNAD要求性の生化学的及び遺伝学的背景である可能 性が示唆された204).しかしさらにその後,生物型 2 (血 清型13)も生物型 1 (血清型 3 )と同様にnadV’を保有 することが明らかになったため203),上記の仮説に矛盾 が生じた.したがって,生物型 1 の NAD 要求性の生化 学的及び遺伝学的背景の解明のために,今後のさらなる 解析が必要と思われる.
3 .血清型について 3.1.はじめに
App には,莢膜の抗原性の違いに基づきこれまでに 15の血清型が存在することが知られてきた13,149).しか し,昨年末になって最も新しい血清型16が提唱された175). 血清型 1 及び 5 は,リポ多糖(LPS)のO抗原及び莢膜 の抗原性の違いに基づき,さらに 2 つの亜型(それぞれ 1a,1b及び5a,5b)に型別される88,89,148).しかし,臨床 細菌学的観点からみると亜型の鑑別の重要性は低い44). 生物型 1 では,血清型14を除くすべての血清型の分離が 報告されている13,149,162,167,175).一方,生物型 2 では,
血清型 2 , 4 , 7 , 9 ,11,13,14のみが分離されてい
る9 ,35,120−122,151).また両生物型ともに,型別不能の
株も存在するが,複数の血清型特異抗血清に反応する ために型別不能な場合と,既知の血清型特異抗血清の いずれとも反応しないために型別不能な場合とがあ
る16,86,100,122,136,137,196).血清型別不能株となった遺伝
学的背景(動く遺伝子 ISApl1の莢膜多糖合成遺伝子へ の挿入による変異)が明らかとなっている型別不能株も ある78).
3.2.血清型別の検査法について
スライド凝集試験129,170)及び共凝集試験131,132)等が,
分離株の簡易・迅速な血清型別法として広く用いられて いる.血清型別には,高力価で特異性の高い血清型別用 抗血清が必要であるが,市販されていない.そのため 通常はホルマリン不活化した各血清型参考株の全菌体を ウサギに免疫して作製した自家製の抗血清が用いられて いる.しかし,この方法で作製した抗血清中には,血清 型特異抗原に対する抗体だけでなく,共通抗原を含む 様々な菌体抗原に対するポリクローナルな抗体を含有 すると考えられる.そのため,異なる血清型間で交差 反応がしばしば認められ,血清型別不能となることがあ る45,51,58,68,72−74,127,128,132,134).
交差反応が認められた場合には,間接赤血球凝集試験130)
やゲル内沈降反応132,133,186)による確定診断を実施する ことが推奨されている.なおスライド凝集試験では血清
性の差に強く関与すると考えられている38,45,46,68,72−74). 各 Apx 毒素の性状,各血清型の株が分泌する Apx 毒素 の種類と各血清型の株の病原性について表 1 及び 2 にま とめた.
3.3.1.2.各血清型のAppにおけるApx毒素の分布について 細胞毒性の強い ApxI 及び細胞毒性が中程度の ApxII の 2 種類の Apx 毒素を分泌する血清型 1 , 5 , 9 及び 11が最も病原性が強いと考えられており38,45)(表 1 〜 2 ),様々な国々で問題となっている(表 3 〜 6 ).さら に,ApxII 及び細胞毒性の強い ApxIII の 2 種類の Apx 毒素を分泌する血清型 2 , 4 , 8 及び15による豚胸膜肺 炎が,それぞれ日本・欧州全土,スペイン,イギリス及び オーストラリアで問題となっている.(表 3 〜 6 )45−47), したがって,ApxI及びApxⅢ毒素のいずれかに加え,
Apx Ⅱ毒素を分泌する血清型の臨床学的重要性が高い と考えられる.
ApxI毒素のみ分泌する血清型10(表 2 )は,ApxIを 型 2 と型別可能な株が,ゲル内沈降反応では型別できず
に型別不能株と同定される株や73,138),凝集試験及びゲ ル内沈降反応のいずれの検査法でも型別できない株も存 在する78).
3.3.血清型別実施の意義について
3.3.1.1.血清型間の病原性の強さに差が認められるため に,血清型別は重要である
Appは,血清型に関わらず病原性を有するが,血清型 によって病原性の強さが異なると言われている45,68,72−74). 病原性因子は複数あると考えられるが45),Apx 毒素
(ApxI 〜 ApxIV 毒素)は,App の最も重要な病原性因 子の一つである38).ApxIV毒素は全ての血清型がin vivo で分泌するが,in vitroでは分泌しない毒素である177). 一方,Appは,ApxI〜ApxIII毒素のうち, 1 種類または 2 種類のApx毒素をin vitroで分泌する38).しかし,血清 型毎にその種類が異なり,その違いが各血清型間の病原
表 1 .App毒素の性状について38,45,68,72−74,94)
Apx毒素 細胞毒性a 溶血性b 病原性への
関与 ワクチンへの
利用 診断への
利用 備考
ApxI 強 強 + + △
(種特異性−) Actinobacillus suisも分泌
ApxII 中程度 弱 + + △
(種特異性−) A. suisも分泌
ApxIII 強 無 + + −
ApxIV ? 有 + − + in vivoでのみ発現.
感染抗体とワクチン抗体を識別 可能な診断用抗原に利用.
a 多形核白血球及びマクロファージの膜に穴を開けて溶解することによって,細胞毒性を示す.
b 赤血球の膜に穴を開けて溶解することによって,溶血性を示す.
表 2 .各血清型の株が分泌するApp毒素の種類と各血清型の株の病原性について38,45−47,68,72−74)
毒素
血 清 型 1,5,9,11 2,4,6,8,15 7,12,13 2 45−47)
(北米分離株) 10,14 3 16 175) 非典型的13
(K13:O10 162)) 非典型的12
(K12:O3 77))
ApxI + − − − + − (+)a (+)a (−)a
ApxII + + + + − − (+)a (−)a (−)a
ApxIII − + − − − + (−)a (−)a (+)a
ApxIV + + + + + + ? ? ?
病原性 強 中程度
7 →中程度 12,13→中程 度または弱?
(血清型 7 以外は臨床例
少ない)
弱?
(臨床例 少ない)
弱?
(病豚から はほとんど 分離されて いない46))
弱 強?a 中程度?
(臨床例162)か ら分離)
中程度?
(臨床例77)か ら分離)
a 毒素タンパク分泌の確認はされていないが,Apx毒素の遺伝子プロファイル39,56,171)から推測.
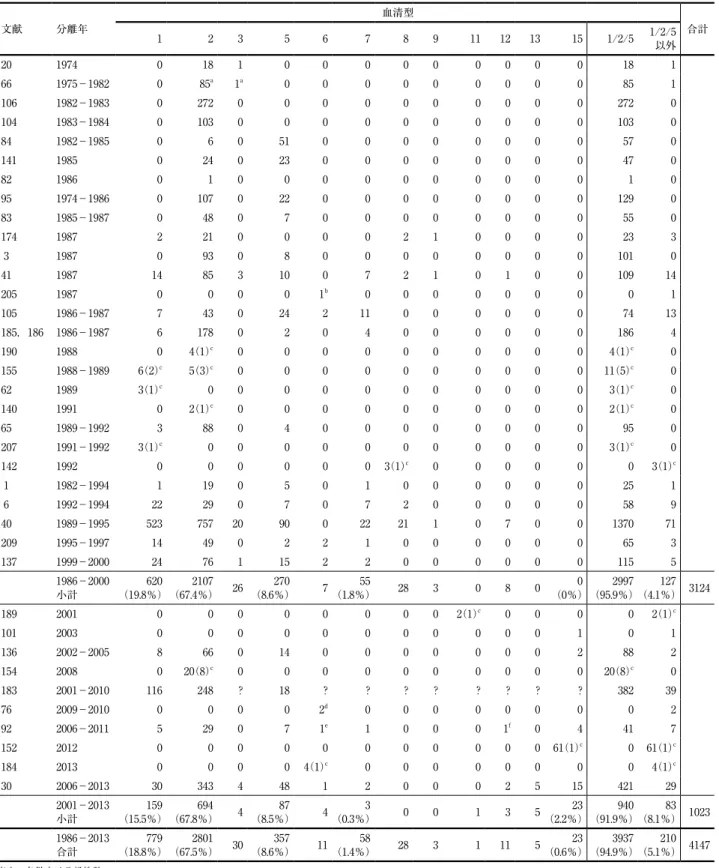
表 3 .1974〜2013年に日本で分離されたAppの各血清型(株数)
文献 分離年
血清型
1 2 3 5 6 7 8 9 11 12 13 15 1/2/5 1/2/5 合計
以外
20 1974 0 18 1 0 0 0 0 0 0 0 0 0 18 1
66 1975−1982 0 85a 1a 0 0 0 0 0 0 0 0 0 85 1
106 1982−1983 0 272 0 0 0 0 0 0 0 0 0 0 272 0
104 1983−1984 0 103 0 0 0 0 0 0 0 0 0 0 103 0
84 1982−1985 0 6 0 51 0 0 0 0 0 0 0 0 57 0
141 1985 0 24 0 23 0 0 0 0 0 0 0 0 47 0
82 1986 0 1 0 0 0 0 0 0 0 0 0 0 1 0
95 1974−1986 0 107 0 22 0 0 0 0 0 0 0 0 129 0
83 1985−1987 0 48 0 7 0 0 0 0 0 0 0 0 55 0
174 1987 2 21 0 0 0 0 2 1 0 0 0 0 23 3
3 1987 0 93 0 8 0 0 0 0 0 0 0 0 101 0
41 1987 14 85 3 10 0 7 2 1 0 1 0 0 109 14
205 1987 0 0 0 0 1b 0 0 0 0 0 0 0 0 1
105 1986−1987 7 43 0 24 2 11 0 0 0 0 0 0 74 13
185,186 1986−1987 6 178 0 2 0 4 0 0 0 0 0 0 186 4
190 1988 0 4(1)c 0 0 0 0 0 0 0 0 0 0 4(1)c 0
155 1988−1989 6(2)c 5(3)c 0 0 0 0 0 0 0 0 0 0 11(5)c 0
62 1989 3(1)c 0 0 0 0 0 0 0 0 0 0 0 3(1)c 0
140 1991 0 2(1)c 0 0 0 0 0 0 0 0 0 0 2(1)c 0
65 1989−1992 3 88 0 4 0 0 0 0 0 0 0 0 95 0
207 1991−1992 3(1)c 0 0 0 0 0 0 0 0 0 0 0 3(1)c 0
142 1992 0 0 0 0 0 0 3(1)c 0 0 0 0 0 0 3(1)c
1 1982−1994 1 19 0 5 0 1 0 0 0 0 0 0 25 1
6 1992−1994 22 29 0 7 0 7 2 0 0 0 0 0 58 9
40 1989−1995 523 757 20 90 0 22 21 1 0 7 0 0 1370 71
209 1995−1997 14 49 0 2 2 1 0 0 0 0 0 0 65 3
137 1999−2000 24 76 1 15 2 2 0 0 0 0 0 0 115 5
1986−2000
小計 620
(19.8%) 2107
(67.4%) 26 270
(8.6%) 7 55
(1.8%) 28 3 0 8 0 0
(0%) 2997
(95.9%) 127
(4.1%) 3124
189 2001 0 0 0 0 0 0 0 0 2(1)c 0 0 0 0 2(1)c
101 2003 0 0 0 0 0 0 0 0 0 0 0 1 0 1
136 2002−2005 8 66 0 14 0 0 0 0 0 0 0 2 88 2
154 2008 0 20(8)c 0 0 0 0 0 0 0 0 0 0 20(8)c 0
183 2001−2010 116 248 ? 18 ? ? ? ? ? ? ? ? 382 39
76 2009−2010 0 0 0 0 2d 0 0 0 0 0 0 0 0 2
92 2006−2011 5 29 0 7 1e 1 0 0 0 1f 0 4 41 7
152 2012 0 0 0 0 0 0 0 0 0 0 0 61(1)c 0 61(1)c
184 2013 0 0 0 0 4(1)c 0 0 0 0 0 0 0 0 4(1)c
30 2006−2013 30 343 4 48 1 2 0 0 0 2 5 15 421 29
2001−2013
小計 159
(15.5%) 694
(67.8%) 4 87
(8.5%) 4 3
(0.3%) 0 0 1 3 5 23
(2.2%) 940
(91.9%) 83
(8.1%) 1023 1986−2013
合計 779
(18.8%) 2801
(67.5%) 30 357
(8.6%) 11 58
(1.4%) 28 3 1 11 5 23
(0.6%) 3937
(94.9%) 210
(5.1%) 4147 表中の各数字は分離株数
a 文献20の菌株数を示してカウントしているので,その株数(それぞれ20及び 1 )を差し引いた数を示した.
b 輸入検疫中に死亡した豚から分離
c 括弧内の数字は農場数.同一農場から分離された株は, 1 株としてカウントした.
d 遺伝学的に非典型的な血清型 6 e 文献76の株と同一株 f 血清型12の非典型的株K12:O3
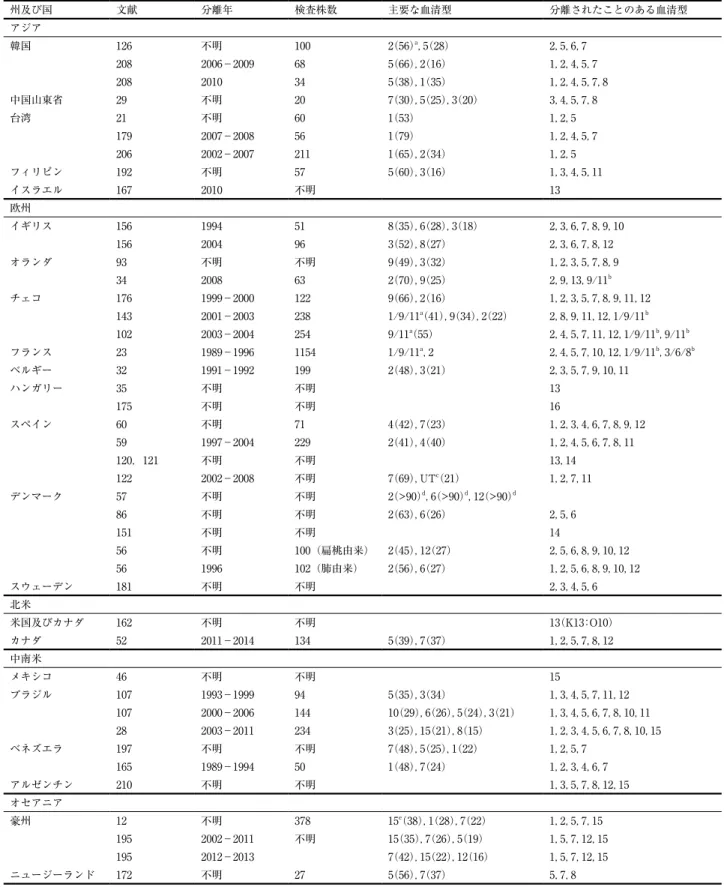
表 4 .日本を除く世界各国で分離されるAppの血清型の割合(%)(型別用抗血清で菌株を血清型別)
州及び国 文献 分離年 検査株数 主要な血清型 分離されたことのある血清型
アジア
韓国 126 不明 100 2(56)a,5(28) 2,5,6,7
208 2006−2009 68 5(66),2(16) 1,2,4,5,7
208 2010 34 5(38),1(35) 1,2,4,5,7,8
中国山東省 29 不明 20 7(30),5(25),3(20) 3,4,5,7,8
台湾 21 不明 60 1(53) 1,2,5
179 2007−2008 56 1(79) 1,2,4,5,7
206 2002−2007 211 1(65),2(34) 1,2,5
フィリピン 192 不明 57 5(60),3(16) 1,3,4,5,11
イスラエル 167 2010 不明 13
欧州
イギリス 156 1994 51 8(35),6(28),3(18) 2,3,6,7,8,9,10
156 2004 96 3(52),8(27) 2,3,6,7,8,12
オランダ 93 不明 不明 9(49),3(32) 1,2,3,5,7,8,9
34 2008 63 2(70),9(25) 2,9,13,9/11b
チェコ 176 1999−2000 122 9(66),2(16) 1,2,3,5,7,8,9,11,12
143 2001−2003 238 1/9/11(41),9(34),2(22)a 2,8,9,11,12,1/9/11b 102 2003−2004 254 9/11(55)a 2,4,5,7,11,12,1/9/11b,9/11b
フランス 23 1989−1996 1154 1/9/11a,2 2,4,5,7,10,12,1/9/11b,3/6/8b
ベルギー 32 1991−1992 199 2(48),3(21) 2,3,5,7,9,10,11
ハンガリー 35 不明 不明 13
175 不明 不明 16
スペイン 60 不明 71 4(42),7(23) 1,2,3,4,6,7,8,9,12
59 1997−2004 229 2(41),4(40) 1,2,4,5,6,7,8,11
120,121 不明 不明 13,14
122 2002−2008 不明 7(69),UT(21)c 1,2,7,11
デンマーク 57 不明 不明 2(>90)d,6(>90)d,12(>90)d
86 不明 不明 2(63),6(26) 2,5,6
151 不明 不明 14
56 不明 100(扁桃由来) 2(45),12(27) 2,5,6,8,9,10,12
56 1996 102(肺由来) 2(56),6(27) 1,2,5,6,8,9,10,12
スウェーデン 181 不明 不明 2,3,4,5,6
北米
米国及びカナダ 162 不明 不明 13(K13:O10)
カナダ 52 2011−2014 134 5(39),7(37) 1,2,5,7,8,12
中南米
メキシコ 46 不明 不明 15
ブラジル 107 1993−1999 94 5(35),3(34) 1,3,4,5,7,11,12
107 2000−2006 144 10(29),6(26),5(24),3(21) 1,3,4,5,6,7,8,10,11 28 2003−2011 234 3(25),15(21),8(15) 1,2,3,4,5,6,7,8,10,15
ベネズエラ 197 不明 不明 7(48),5(25),1(22) 1,2,5,7
165 1989−1994 50 1(48),7(24) 1,2,3,4,6,7
アルゼンチン 210 不明 不明 1,3,5,7,8,12,15
オセアニア
豪州 12 不明 378 15(38),1(28),7(22)e 1,2,5,7,15
195 2002−2011 不明 15(35),7(26),5(19) 1,5,7,12,15
195 2012−2013 7(42),15(22),12(16) 1,5,7,12,15
ニュージーランド 172 不明 27 5(56),7(37) 5,7,8
a 括弧内の数値は分離率.
b 複数の血清型の抗血清で凝集.
c 型別不能.
d 血清型 2 , 6 ,12あわせて90%以上.
e 原著では当時血清型12となっているが,後に実際には血清型15と判明.
表 5 .日本を除く世界各国で分離されるAppの血清型の割合(%)(PCR法で血清型を予測)
州及び国 文献 分離年 株数 主要な血清型 分離されたこと
のある血清型 型別法
アジア
韓国 91 2008−2010 32 5(50),1(22),2(16) 1,2,5,12 PCR(遺伝子不明)
96 2011−2012 48 2(42),5(33) 1,2,4,5,7,12 cps/apx/omlA PCR
111 2012−2013 54 5(37),1(28),UT a(24) 1,2,5,7,12 cps PCR
タイ 191 不明 1 15 apxIV REA b
欧州
イギリス 156 2004 66
(同一検体)
3(52),8(30) 2,3,6,7,8,12 従来の血清型別法
(凝集試験)
8(82) 2,3,6,7,8,12 cps PCR
156 2009 378 8(78) cps PCR
北米
カナダ 51 2007−2013 96 6,8,3/6 c,15? cps PCR( 血 清 型 3 ,
6 及び 8 抗血清で交差 する株のみを検査.)
南米
チリ 144,145 2007−2009 60 7(36),6(17) 4,6,7 apx PCR
コロンビア 173 不明 不明 1,5,7 apxIV PCR
a 型別不能株.
b restriction analysis(制限酵素切断解析)
c 血清型 3 及び 6 に特異的なサイズのPCR産物が 2 本増幅されたため,非典型的な血清型 676). 下線はPCR法ではなく,抗血清を用いた従来の血清型別法(スライド凝集試験)での成績.
表 6 .世界各国におけるAppの各血清型に対する抗体陽性率(%)
州及び国 文献 抗体検査の実施年 頭数 農場数 主要な血清型 抗体陽性を示した血清型 備 考
アジア
日本 106 1982−1983 119 18 2(44)a 2 血清型2のみ検査
104 1983−1984 200 48 2(52) 2 血清型2のみ検査
141 1985 64 6 5(36),2(38) 2,5 血清型2,5のみ検査
83 1983−1986 629 220 2(62),5(10) 2,5 血清型2,5のみ検査
207 1992 473 1 5(91)b,2(83)b,1(47),6(44),7(17) 1,2,5,6,7 血清型1,2,5−7のみ検査 188 2012−2013 31 2(74),5(61),1/9/11(19)c 1/9/11,2,5 血清型1/9/11,2,5のみ検査
188 2012−2013 144 2(62),5(25) 1/9/11,2,5 血清型1/9/11,2,5のみ検査
韓国 96 2011−2012 452 2(19) 1/9/11,2,3/6/8d,4/7e,5
タイ 7 不明 549 1/9/11(29),3/6/8(26),5a(26) 1/9/11,3/6/8,5a,5b,12 欧州
ベルギー 118 不明 150 3(90),2(81),9(39) 農場内平均感染率
血清型2,3,9のみ検査
119 不明 50 2(57),3(54),9(41) 農場内平均感染率
血清型2,3,9のみ検査 オランダ 34 2008 500 2(35),12(28),4/7(21),3/6/8(17) 1/9/11,2,3/6/8,4/7,5a/5b,10,12
スウェーデン 199 2000 500 4/7(59),2(49),3(39) 農場内平均感染率
フィンランド 112 不明 692 3(51),2(26)
61 2007−2008 206 4/7(35),1/9/11(15) 1/9/11,2,3/6/8,4/7,5,12 野生及び家畜化した猪 北米
カナダ 115 2003 50 4/7(26),12(17),3/6/8(15) 1/9/11,2,3/6/8,4/7,5,12 中南米
メキシコ 19 不明 556 2(89),3(86),1(50)
a 括弧内の数値は抗体陽性率.
b ワクチン抗体?
c 血清型LPSのO抗原を用いたELISAで検査したので,血清型1/9/11間の識別はできない.
d 血清型LPSのO抗原を用いたELISAで検査したので,血清型3/6/8間の識別はできない.
e 血清型LPSのO抗原を用いたELISAで検査したので,血清型4/7間の識別はできない.