K
家畜衛生学雑誌 家畜衛生学雑誌
Vol.45 No.1-2 2019. JUL.
日 本 家 畜 衛 生 学 会
The Japanese Society of Animal Hygiene
The Japanese Journal of Animal Hygiene
家畜衛生学雑誌
第 45巻第1
-2号
二〇一九年七月
日本家畜衛生学会
乳剤
家畜衛生学雑誌
45
-1
-2
表14( 附 ) 日本家畜衛生学会第90回大会 要旨集
複写される方へ
日本家畜衛生学会は有限責任中間法人 学術著作権協会(学著協)に複写に関する権利委託をしていますので,本誌に掲 載された著作物を複写したい方は,学著協より許諾を受けて複写して下さい.但し,社団法人日本複写権センター(学著協 より複写に関する権利を再委託)と包括複写許諾契約を締結されている企業の社員による社内利用目的の複写はその必要は ありません.(※社外頒布用の複写は許諾が必要です.)
権利委託先: 有限責任中間法人 学術著作権協会
〒107-0052 東京都港区赤坂9-6-41 乃木坂ビル3階
電話:03-3475-5618 FAX:03-3475-5619 E-mail:info@jaacc. jp
注意:複写以外の許諾(著作物の転載・翻訳等)は,学著協では扱っていませんので,直接日本家畜衛生学会へご連絡下 さい.[電話:042-367-5780]
また,アメリカ合衆国において本書を複写したい場合は,次の団体に連絡して下さい.
Copyright Clearance Center, Inc.
222 Rosewood Drive, Danvers, MA 01923 USA
Phone:1-978-750-8400 FAX:1-978-646-8600
家 畜 衛 生 学 雑 誌
日本家畜衛生学会 発行
President : Junsuke SHIRAI(Tokyo Univ. of Agric. and Technol.)
Vice President : Shigeru MIYAZAKI(Res. Inst. for Anim. Sci. in Biochem. and Toxicol.)
Editor-in-Chief : Shigeru MIYAZAKI(Res. Inst. for Anim. Sci. in Biochem. and Toxicol.)
Editorial Board : Masuo SUEYOSHI(Miyazaki Univ.)
Shinji TAKAI (Kitasato Univ.)
Makoto NAGAI(Ishikawa Pref. Univ.)
Sadao NOGAMI(Nihon Univ.)
Hideto FUKUSHI(Gifu Univ.)
"""""""""""""""""""""""""""""""""""""""""""
The Japanese Journal of Animal Hygiene
Published by the Japanese Society of Animal Hygiene
理 事 長 :白井淳資 副理事長 :宮﨑 茂 編集委員長 :宮﨑 茂
編集委員 :末吉益雄・髙井伸二・長井 誠 野上貞雄・福士秀人
誌
45
-1
-2
表23会員の皆様におかれましては,ますますご清栄のこととお慶び申し上げます.ここに,「家畜衛生学雑誌」第45巻 第 1 − 2 号を刊行する運びとなりました.
本号では,総説 1 編の他,「第90回大会講演要旨」を掲載しています.
家畜衛生学雑誌では,原著論文・短報以外にも,総説,数ページ程度のミニレビュー,技術資料等の原稿を受け付 けております.原著論文,総説はもちろんでございますが,ミニレビューや技術資料も含め,会員の皆様の積極的な ご投稿をよろしくお願い致します.ご不明な点はご遠慮なく編集委員会事務局へお問い合わせください.
日本家畜衛生学会理事長 白井淳資 家畜衛生学雑誌編集委員長 宮﨑 茂
(日本家畜衛生学会副理事長)
日本家畜衛生学会・学会費納入のお願い
ご承知のように,学会は会員の皆様からの会費をもって運営されております.学会の運営を円滑に運ぶために,所 定の会費を納入していただきますようお願い致します.
*会費は,正会員5,000円,学生会員2,000円です.
*2015年度までの未納分をお支払いいただく場合,正会員年会費は4,000円です.
日本家畜衛生学会 理事長 白井淳資
00
口座記号 口座番号(右詰めで記入) 金額料金 特殊
取扱
千 百 十 万 千 百 十 円
※
※
※
※
加入者名通信欄ご依頼人 受付局日附印
※
※
(電話番号 - - ) おところ(郵便番号 - )
※ おなまえ
払 込 取 扱 票
口座記号番号加入者名金額料金特殊取扱ご依頼人
※
受 付 局 日 附 印
※
※
※
※
※
円
千 百 十 万 千 百 十 円
おなまえ
郵便振替払込請求書兼受領証
記載事項を訂正した場合は、その箇所に訂正印を押してください。切り取らないで郵便局にお出しください。
各票の※印欄は、ご依頼人において記載してください。
裏面の注意事項をお読みください。
これより下部には何も記入しないでください。
(消費税込み)
様
様
日本家畜衛生学会
0 0 2 4 0
4 3 1 7 1 3 0 0 2 4 0 3 4 3 1 7 1
日本家畜衛生学会
2015 2016 2017 2018 2019 年度
( ) 計 円
ので、口座番号及び金額を記入す る際は、枠内にはっきりと記入し てください。
また、本票を汚したり、折り曲 げたりしないでください。
・この払込請求書を郵便局の派遣 員にお預けになるときは、引換え に預り証を必ずお受け取りください。
この受領証は、郵便振替の払込 みの証拠となるものですから大切
に保存してください。 この払込取扱票の裏面には、何も記載しないでください。
家畜衛生学雑誌
第45巻 第 1 − 2 号 2 0 1 9
目 次
〈総説〉動物由来のパスツレラ科に属する細菌のPCRによる同定・検出及び血清型の同定法ならびにRTX毒素について
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 伊藤博哉 ・・・・・・・・・・・・・・・・・・・・・・・・・ 1〜38
〈第90回大会一般講演要旨〉
散歩中の犬間の接触率および狂犬病に対する犬飼育者の行動
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ ナイジェル クワン・井上 舞・山田章雄・杉浦勝明 ・・・・・・・・・・・・・・・・・・・・・・・・ 42〜43 携帯品で持ち込まれた非加熱肉等のウイルス汚染状況調査
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 小林芳史・柴田明弘・稲垣麻由子・栁澤成江
尾坂優之・遠藤明仁・岩中麻里・大友浩幸 ・・・・・・・・・・・・・・・・・・・・・・・・ 44〜45 養豚場72戸における2015〜2017年の抗菌剤使用量の記述的調査
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 雷 志皓・山根逸郎・山崎久則
内藤 恵・呉 克昌・杉浦勝明 ・・・・・・・・・・・・・・・・・・・・・・・・ 46〜47 クレブシエラ乳房炎の重篤度と分離された原因菌の好中球貪食抵抗性の関連
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ インペラトリス瑠伽・楠本周平・石村麻莉乃
澤松祐人・篠塚康典・河合一洋 ・・・・・・・・・・・・・・・・・・・・・・・・ 48〜49 食性の異なる各種動物からのウェルシュ菌の分離調査
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 清水かな・上塚浩司 ・・・・・・・・・・・・・・・・・・・・・・・・ 50〜51 競走用馬の感染性とみられる皮膚炎からの細菌・真菌の分離同定と薬剤感受性調査( 2 )
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 牛屋重人・上塚浩司 ・・・・・・・・・・・・・・・・・・・・・・・・ 52〜53
〈第90回大会教育講演要旨〉
家畜の下痢症から検出されるウイルスについて
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 長井 誠 ・・・・・・・・・・・・・・・・・・・・・・・・・ 55〜56
会員へのおしらせ ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 57 家畜衛生学雑誌投稿規程 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 59〜60 日本家畜衛生学会会則 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 61〜62
家 畜 衛 生 学 雑 誌
The Japanese Journal of Animal Hygiene Vol. 45 No. 1-2 2 0 1 9
Contents
〈Review〉
Identification, detection and serotyping of Pasteurellaceae species from animals by PCR and their RTX toxins
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ Hiroya Ito ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 1〜38
〈Abstracts of oral presentations on 90th academic meeting〉
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 39〜53
〈Abstracts of the educational lecture on 90th academic meeting〉
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 55〜56
Information for Members ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 57 Instruction for Authors ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 59〜60 The Regulations of The Japanese Society of Animal Hygiene ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 61〜62
Jpn. J. Anim. Hyg.
1 .はじめに
家畜・家禽に病原性を示すfamily Pasteurellaceae(パ スツレラ科)に属する細菌は多いが,表現型のみでは同 定が容易ではない菌種が多数存在する.本稿では,家 畜・家禽に病原性を示す病原性パスツレラ科細菌の(i)
PCR による菌種の同定及び検出,(ii)パスツレラ科細 菌の血清型,(iii)PCRによる血清型の同定,(iv)病原 性因子である RTX 毒素(パスツレラ科細菌のいくつか の菌種が産生する毒素)とPCRによるRTX遺伝子の検 出法について概説する.RTX 毒素はパスツレラ科細菌 のいくつかの菌種が分泌する病原性因子であり,宿主特
異性にも関連する.さらに,RTX毒素遺伝子の検出は,
菌種同定の参考になる菌種も存在するため紹介する.
2 .PCRによる同定及び検出について 2.1.はじめに
パスツレラ科には,2019年 1 月現在17の属,74の菌種 が認められている137).これらの菌種は,家畜・家禽の 扁桃等に存在する常在菌が多いが,病原性を示す菌種も 多数存在する,またパスツレラ科細菌には,生物学的・
生化学性状等の表現型の性状のみでは同定・識別しにく い菌種が多い.そのため既存の菌種とは異なるが,菌種 名の命名までには至らず,未整理のままのtaxon(分類 群)も多い,しかし,近年,遺伝子の性状を利用するこ とによって,未整理であったパスツレラ科に属する taxonが次々と新菌種として提唱されている,
細菌の分類・同定のためのgold standardは,DNA-DNA ハイブリダイゼーション法(DNA-DNA hybridization
動物由来のパスツレラ科に属する細菌のPCRによる 同定・検出及び血清型の同定法ならびにRTX毒素について
伊 藤 博 哉1 )*
Identification, detection and serotyping of Pasteurellaceae species from animals by PCR and their RTX toxins
Hiroya ITO 1)*
(1)National Institute of Animal Health, NARO, 3-1-5 Kannondai, Tsukuba, Ibaraki 305-0856, Japan)
(2019. 3. 31 受付/2019. 6. 19 受理)
Summary
Here, I review the current knowledge on (i) PCR identification and detection of Pasteurellaceae from animals, (ii) serovars of the Pasteurellaceae and (iii) PCR identification of serovar of the Pasteurellaceae as well as (iv) RTX toxins of the Pasteurellaceae and PCR detection of their genes. I hope that information above will be helpful for identification, detection and serotyping of Pasteurellaceae from animals as well as for understanding of the RTX toxins.
Key words: Pasteurellaceae, diagnosis, serovars, RTX toxins, PCR
家畜衛生学雑誌 45, 1 〜38(2019)
1 )
国立研究開発法人農業・食品産業技術総合研究機構 動物衛生研究部門
〒305−0856 茨城県つくば市観音台 3 − 1 − 5
*
連絡著者:伊藤博哉([email protected])
(DDH))である.しかし,DDHは操作が煩雑であり,
さらに基準となる株を全て揃えなければならず,どこで でも実施できる方法ではない.そのため,現在では各菌 種でその塩基配列が保存されている16S rRNA 遺伝子
(16S rrn)の配列を決定して,菌種同定の際の参考にし ている検査室や研究室も多いと思われる. 異なる菌種 と判断して構わないと考えられる16S rrnの塩基配列の 相同性は,当初(かつて)は97%と言われていた(注:
細菌分類学の世界では,厳密に言うと,97%以上の相同 性を示しただけでは,その菌種であると同定はできな い.すなわち,その菌種である可能性が高いため,引き 続き,上述の DDH を実施して,その菌種であるか,ま たは新種であるかを確定する必要があると言われてい る154).しかしながら,病性鑑定の現場では,同定目的 でそこまでする検査室はほとんどに無いと思われる.).
しかしその後,異なる菌種であっても相同性の高い近縁 な菌種が多数存在し,閾値を98.7〜99.0%程度にすべき とされている 154,160).しかし,この数値は菌種によって は例外もあり,たとえば牛の慢性肉芽腫性炎を主徴とす る 牛 ア ク チ ノ バ チ ル ス 症( 木 舌 症 ) の 原 因 菌 Actinobacillus lignieresii の基準株の16S rrnの塩基配列 を用いてBLAST相同性検索すると,豚胸膜肺炎の原因 菌 Actinobacillus pleuropneumoniae の基準株の16S rrn 配列と99%以上の相同性を示す.さらに豚のアクチノバ チルス・スイス感染症の原因菌Actinobacillus suis の基 準株の16S rrn の塩基配列を用いて BLAST 相同性検索 すると,子馬病の原因菌 Actinobacillus equuli の基準株 の16S rrn 配列と約98.7%の相同性を示す.このように 16S rrnの配列のみでは,種を同定することが困難な菌 種も存在するが,16S rrn解析は,両菌種を識別可能な 生物学的性状,生化学性状及び他の遺伝学的性状の検査 を加えて総合的に判断する際の有用な情報となる.
Christensen らの総説では,一部の例外(A. equuli.後 に説明)を除いて,Actinobacillus 属では,分離宿主を 同定のための参考性状としてもよいと記載されている
31).
このように16S rrnの塩基配列は,菌種同定に大いに 役立つ方法であるが,塩基配列の決定は一部の検査機関 しか実施できないまたは実施していないのが実態である と考えられる.本稿では,現在ではおそらくほとんどの 検査室で実施可能と思われる PCR による遺伝学的菌種 同定法や臨床材料からの検出法を紹介する.
2.2.PCRによる同定・検出法の利点と限界 2.2.1.PCRによる同定・検出法の利点
操作が定型的で簡便である.また,用いるプライマー
の種類や,PCR 反応の条件が菌種によって違うだけで あり,使用する機械や酵素は同一で良いので,あらゆる 病原体に対応可能である.さらに操作方法も基本的には 同一である.
2.2.2.PCRによる同定・検出法の問題点と注意点 生化学的性状等の表現型の検査等と併せて,総合的に 診断する必要がある.さらに,陽性対照株を準備してお くことが重要である.また一番重要な点であるが,誤診 につながる非特異反応の増幅が認められることもあり,
その際には見極める能力が必要である.非特異的な増幅 かどうか確認するためには,サザンハイブリダイゼー ションや,増幅された PCR 産物の塩基配列の決定等を 行えば良い.使用している酵素の違い及び PCR 装置の 機種の違いによって,DNA 伸張(複製)の能力や,温 度上昇の度合いに差があるため,文献どおりに PCR を 実施してもうまくいかないことがある.その時は,使用 している酵素や機種に適応した反応時間や温度等の調節 が必要な場合がある.
3 .PCRによる血清型の同定法について 3.1.はじめに
細菌は表層に存在する構造物の抗原性の違いに基づ き,複数の血清型に型別される.細菌種によっては,ワク チンの有効性が血清型特異的であること120,151,157,165,166)
及び血清型によって病原性が異なることがある等の理 由75)や,疫学解析の目的で,臨床材料から分離された 細菌の血清型別は,古くから行われてきている.血清型 別を行うためには,通常,細菌をホルマリン等で不活化 したのち,ウサギ等の動物に接種して作製した高力価 で,かつ特異性の高い免疫血清(抗血清)を使用する必 要がある.しかし,動物由来の重要な細菌の多くについ ては,血清型別用の抗血清は市販されておらず,自家製 の血清型別用抗血清を作製するか,各菌種のリファレン スラボラトリーに依頼して血清型別を実施する必要があ る.
本稿では,重要と考えられる動物由来の主要なパスツ レラ科細菌の血清型及び抗血清を用いずに PCR で血清 型を同定する方法について,文献を紹介して概説を行 う.
3.2.PCRによる血清型の同定法の利点と限界 3.2.1.PCRによる血清型の同定法の利点
抗血清を用いた血清型別を実施するには,実験動物を 用いて抗血清を作製する必要があるが,PCR 法の場合 はその必要がない.また抗血清を用いた血清型別を行う ためには,全ての血清型に対する抗血清を用意する必要
4 .動物由来のパスツレラ科に属する細菌の RTX毒素について
4.1.はじめに
RTX 毒素は,菌体外に分泌される外毒素であり,赤 血球,好中球等の白血球,マクロファージ等の細胞膜に 穴を開ける孔形成毒素(pore forming toxin)である,
ほとんどの RTX 毒素は溶血活性をもっているため,赤 血球の溶解が各疾病の出血性病変の一因と考えられてお り,さらに RTX 毒素は上記の生体防御に関連する細胞 に対して細胞障害性を示すため,宿主の免疫機能から回 避できると考えられ,RTX 毒素は病原細菌の重要な病 原性因子の一つと考えられている.RTX 毒素はタンパ ク質毒素で,その大きさは,100-200 kDaである.RTX 毒素のC末端部分にはグリシンが多く含まれるカルシウ ムイオン結合性のモチーフが10〜40個繰り返し並んでい る特徴的な構造(グリシンが豊富な繰り返し領域)を有 していることからRTX(repeats in toxins)毒素と呼ば れており,様々な菌種が RTX 毒素を産生する(図 1 −
3 )44,46).
本項では,動物由来のパスツレラ科に属する RTX 毒 素について概説するとともに,それらの RTX 毒素の PCR法による検出法について,紹介する.
4.2.RTX毒素の概説
RTX 毒素の構造タンパク質は, 4 つの遺伝子からな るオペロン(rtxCABD)によって,まず細胞質内で産 生・活性化され,そして菌体外に分泌される.それぞれ があるため,全ての各血清型の参考株を入手しなければ
ならないが,PCR 法の場合,PCR プライマーや PCR 酵 素等の試薬等は簡単に購入でき,PCR の装置があれば 型別が可能である.PCR 装置の価格はやや高額である が,現在では多くの検査室が保有していると思われる.
また一般的に抗血清を用いた血清型法では,異なる血清 型間での交差反応が認められる例が多いが,PCR では その問題を克服できる例が多い.
3.2.2.PCRによる血清型の同定法の問題点及び注意点 変異によって,血清型に関係する抗原が発現されなく なった場合には,表現型phenotype(抗血清を用いた検 査による血清型)と遺伝子型 genotype(PCR による血 清型)が一致しなくなることがあり,我々もその例に遭 遇したことがある73).さらにその事例では, 2 種の異な る PCR 法で検査すると,異なる結果が得られた(片方 は 血 清 型15, も う 一 方 は 型 別 不 能 と 同 定 さ れ た A. pleuropneumoniae)73).その原因は,遺伝子の変異 による欠失であり, 2 種類の PCR のうち片方の PCR は 欠失した遺伝子領域に結合するプライマーを設定してい たが74),もう一方の PCR は,欠失していない他の DNA 領域に結合するプライマーを設計していたためであっ た168).結局その株は,元々血清型15であったが,血清 型に関与する遺伝子の一部を欠失したために,抗血清を 用いた血清型別では型別不能となる株であった73).
図 1 .A. pleuropneumoniaeのApxI,ApxII及びApxIII毒素遺伝子オペロンのフィジカルマップ21,42,69,72,130,140). 実線の長方形,機能している遺伝子.破線の長方形,一部を欠損しているために機能していない遺伝子.長方 形の横の長さは,遺伝子の長さに基づいてはいない.
の遺伝子は,rtxC,rtxA,rtxB,rtxDの順番にタンデム に並んでいる.rtxA は,RTX 毒素の構造タンパク質
(RtxA)をコードし,さらに rtxC にコードされた RtxC のアシル化作用によって,RtxAは細胞質内で活性化さ れる.活性化された RtxA は,さらにタイプ I 分泌シス テム(type I secretion system(TISS))(rtxB及びrtxD にコードされた細胞内膜に存在する分泌タンパク質
RtxB及びRtxD)によって,菌体外に分泌される.本稿 では,動物由来のパスツレラ科細菌の RTX 毒素(図 1
〜 3 )について概説する.
パスツレラ科細菌に属する菌種の病原性は,宿種特異 性を示すものが多いが,RTX 毒素の各動物の細胞に対 する毒性が,宿種特異的であることがその一因であると 考えられている46,90,170).例えば,Kuhnert ら90)は豚に 図 2 .A. pleuropneumoniae 以 外 の 細 菌 の Apx 毒 素 遺 伝 子 オ ペ ロ ン の フ ィ ジ カ ル マ ッ プ.var. suis,var.
porcitonsillarum,var. rossii,var. lignieresii 及 び var. Glaessereella は, そ れ ぞ れ,A. suis,A.
porcitonsillarum,A. rossii及びA. lignieresii及びGlaesserella sp.におけるapxのバリアントである.白い長 方形からなるオペロン,Apx毒素が発現しているオペロン. 黒色の長方形からなるオペロン,Apx毒素の発現 が認められないオペロン.A. porcitonsillarumのaspC(縦線の入った長方形)の毒素との関係は不明である.
長方形の横の長さは,遺伝子の長さに基づいてはいない.
図 3 .Actinobacillus属細菌以外(Glaesserella sp.を除く)のRTX毒素遺伝子オペロン.縦線の入った長方形,他の 典型的なRTX毒素よりも遺伝子が長く,5’-末端領域部分は他のRTX毒素遺伝子の塩基配列との相同性がない rtxA.黒色の長方形,他の典型的なRTX毒素遺伝子オペロンには認められない遺伝子.
特異的であるため,毒素遺伝子プロファイルで,おおよ その血清型(群)を推定できる43,45,52,69)(図 1 )( “5.1.4.
A. pleuropneumoniae” の項参照)また,それぞれの菌 種に特有なRTX毒素も存在する(図 1 − 3 )ことから,
RTX 毒素遺伝子の検出を実施することによって,菌種 または亜種の同定の参考にすることができる.
5 .豚から分離されるパスツレラ科に属する 病原細菌
5.1.Actinobacillus pleuropneumoniae
A. pleuropneumoniae は 豚 胸 膜 肺 炎 の 起 因 菌 で あ る66,67,69,141).
5.1.1.A. pleuropneumoniaeのPCRによる同定・検出 法について(表 1 )
Sirois らは,A. pleuropneumoniae の同定・検出用の PCR を本菌で初めて開発している150)(表 1 ).しかし,
A. pleuropneumoniaeと非常に近縁なA. lignieresiiでも 陽性となり,必ずしも種特異的ではない.しかし,
高い宿種特異性を示す A. suis の RTX 毒素(ApxI 及び ApxII 毒素)(図 2 )は,馬のリンパ球よりも豚のリン パ球により高い毒性を示し,逆に馬に高い宿種特異性を 示 す Actinobacillus equuli subspecies haemolyticus の RTX毒素(Aqx毒素)(図 3 )は,豚のリンパ球よりも 馬のリンパ球により高い毒性を示すことを明らかにして いる.
4.3. PCRによる動物由来のパスツレラ科細菌に属する 細菌のRTX毒素遺伝子の検出法について
RTX 毒素タンパク質の発現,産生及び分泌を確認す るには,RTX毒素に特異的な抗体を用いたウエスタンブ ロッティング等のタンパク質解析や,細胞を用いた細胞 障害性試験が必要である.さらに,PCR を用いて RTX 毒素遺伝子の存在を確認する方法が各種細菌で開発され ている.
A. pleuropneumoniaeには 4 種のRTX毒素及びその遺 伝子が存在するが,その組み合わせはほぼ血清型(群)
表 1 .A. pleuropneumoniaeの種同定用または検出用PCR
標的遺伝子等 同時に型別可能な血清型 文献 異なる菌種での増幅a, b
未同定のDNA 150 A. lignieresii陽性b
未同定のDMA(文献150と同じプライマー使用) 51 A. lignieresii陽性b
cpx 5 99 A. lignieresii陽性a
omlA 50 A. lignieresii陽性b
omlA 29 A. lignieresii陽性b
aroA 58 A. equuli陽性b
tbpA/tbpB+RFLP 35 RFLPを併用すれば,なしb
omlA 143 なしb
omlA (5’-nuclease assay) 7 なしb
apxIV 144 なしb
dsbE 30 A. lignieresii及びA. porcitonsillarum陽性b
omlA 2, 5, 6 76 A. lignieresii陽性 (n=50)a
cpxD 1, 2, 8 148 なし(ただし,A. lignieresiiやA. genomospecies 1の検査なし)a
apxIV 3 180 A. genomospecies 1で陽性 (n=2/3)a
apxIV 3, 6, 8 181 A. genomospecies 1で陽性 (n=2/3)a
omlA 1, 7, 12 4 A. genomospecies 1で陽性 (n=4/6)
Metalloprotease遺伝子 149 なしb
apxIV 1-3, 5-8, 10, 12 18 Pig onlya, c
apxIV 61 なしb
apxIV(RT-PCR) 161 なしb
omlA 1, 5, 7, 12, 15 168 Actinobacillusの 3 菌種で非特異バンドa, d.
BOX-SCAR-PCR 138 A. lignieresii陽性b
omlA 1, 2, 5, 7, 15 74
apxIV 16 20 Pig onlya, c
apxIV 1-8, 10, 12, 15 19 Pig onlya, c
apxIV 13, 14, 16-18 19 Pig onlya, c
a, b オリジナルの論文aまたは総説47bから転用.
c Pig only, 豚由来の菌種全て陰性,豚以外の動物から分離された菌種の検査なし.
d ウサギ由来のActiobacillus capsulatus( 1 株)と豚由来のA. indolicus( 1 株)及びA. suis( 1 株)で非特異増幅.
Actinobacillus 属に分類される菌種の種特異性は極めて 高く(後述する A. equuli を除く),分離された宿主は 同定のために参考となる重要な性状と考えられているた め31),現時点では実用的には大きな問題とはならないと 考えられている.以降もA. pleuropneumoniae特異的と 謳った様々な PCR 法(表 1 )が開発されているが,A.
lignieresiiでも陽性となる例が多い.
さらにいくつかの遺伝子が A. pleuropneumoniae の PCR による同定または検出のための標的遺伝子に適し ていると報告されている(表 1 ).それらは,(i) omlA
(outer membrane lipoprotein gene(外膜リポタンパク 質遺伝子))50),(ii) apxIV(A. pleuropneumoniae toxin IV gene(A. pleuropneumoniae毒素IV遺伝子)144),(iii)
dsbE(a thiol:disulfide interchange protein gene(チ オール:ジスルフィド交換タンパク遺伝子)30)及び(iv)
cpxD(capsular polysaccharide export gene(莢膜輸送 タンパク質D遺伝子)99 )等である(表 1 ).
著者は,A. pleuropneumoniae 用の omlA を標的とし たPCRを菌種同定の参考にすることが多い52,74).Gram らは,omlA を標的とした PCR を開発し50),Savoye ら は,さらなる特異性の上昇及び解析時間の短縮のため に,プライマーと PCR の条件に若干の変更を加え,新 たなomlA PCR法を開発した143). そして,さらにPCR の阻害による偽陰性反応を除去するために,internal positive control を ア ッ セ イ 系 に 加 え た. そ れ 故,A.
pleuropneumoniae に特異的で,他の細菌が高度に汚染 されたサンプルであってもA. pleuropneumoniaeを検出 可能としている.また論文には種特異的 PCR であると いう記載は無いが,Osaki らは,omlA PCR-restriction fragment length polymorphisms(PCR-RFLP) に 用 い る PCR も種特異的であることを日本で分離された野外 株で確認している125).詳細は公表されていないが,海 外ではomlAを標的としたPCR診断キットも市販されて いる29).omlA PCRは,他のPCR法と同様に,残念なが らA. lignieresii及びActinobacillus genomospecies 1等,
近縁な菌種でも陽性となる株があると報告されている
(表 1 ).しかし繰り返しになるが,Actinobacillus 属 の 菌 種 は 一 部 の 例 外 を 除 い て 宿 主 特 異 性 が 高 く,
A. lignieresiiは反芻獣,A. genomospecies 1は馬を宿主 とするため,大きな問題はないと考えられている.
apxIV は,全ての A. pleuropneumoniae が保有と言わ れている遺伝子である.この apxIV を標的とした PCR は,A. pleuropneumoniaeの同定及び検出が可能とされ ている144).Schallerの開発したapxIV PCRは,感度を上 げ る た め に,nested PCR を 採 用 し て い る.apxIV が コードするApxIVタンパク質は,A. pleuropneumoniae
感染抗体を特異的に検出するApxIV ELISAの抗原にも 使用されているため,apxIVをA. pleuropneumonie種特 異的 PCR の標的遺伝子にすることは理にかなっている と考えられる.apxIV PCR は,omlA PCR 同様に,A.
pleuropneumoniae と 近 縁 な A. lignieresii 及 び A.
genomospecies 1で も 陽 性 と な る( 表 1 ). し か し,
Angen らは,omlA PCR と apxIV PCR の他菌種との特 異性を比較した結果,omlA PCRの方が高い特異性を示 すことを報告している3 ).
Xiao らは,omlA 及び apxIV の 2 つの遺伝子を標的と した PCR 法を開発している177).本法は,高感度で特異 性が高く,同定だけで無く,材料からの直接検出に効果 的方法としている
dsbE を 標 的 に し た PCR が 開 発 さ れ た が, 近 縁 な A. lignieresii でも陽性となる30).Actinobacillus 属に分 類される細菌は宿主特異性が高く,A. lignieresiiは豚か ら分離されたことはないため,大きな支障にはならない と考えられるが,しかし,豚から分離されるActinobacillus porcitonsillarum でも陽性となることが明らかになって いる41).
cpx PCRは,血清型 1 〜12のうち,血清型 4 では増幅 が認められないので,全ての血清型を同定できるわけで は な い99). ま た A. lignieresii や 豚 か ら 分 離 さ れ る Actinobacillus minor,Actinobacillus indolicus 及び A. suisでも陽性となった報告がある3 ).
aroA(3- ホスホシキミ酸 -1- カルボキシビニルトラン スフェラーゼ)を標的としたaroA PCRも開発されてい るが,本PCRでは豚から分離されるA. equuliやA. suis でも陽性となる41,58).
このようにA. pleuropneumoniaeの種特異的PCRが複 数開発されているため,それぞれの方法を比較・評価し た報告がある3 ).上記の 4 つの遺伝子を標的にしたPCR
(omlA PCR,apxIV PCR,dsbE PCR及びcpx PCR)の 特異性を比較した結果,omlA を標的とした omlA PCR の特異性が最も高かった3 ).反芻獣由来A. lignieresii 45 株,馬由来A. lignieresii(A. genomospecies 1) 6 株,
豚由来のA. indolicus 8株,A. minor 13株,A. porcinus 8株,A. suis 30株を用いてそれぞれの PCR の特異性を 調 べ る と,omlA PCR は 馬 由 来 の A. lignieresii(A.
genomospecies 1) 4 株でPCR増幅が認められた(特異 性96.4%).一方,dsbE,apxIV 及び cpxD を標的とする PCR の場合,反芻獣及び馬由来の A. lignieresii の全て または92%で陽性を示すほか,非病原性と考えられてい るが,豚の上部呼吸器官から分離されるNAD要求性の A. indolicus,A. minor,A. porcinusでは,少数である が PCR 陽性となった.cpxD PCR では,A. suis 全ての
2015年に提唱された血清型16は,ユニークな莢膜(K16 抗原)合成遺伝子構造を保有しているが20)(図 4 (D)),
血清型16のO抗原合成遺伝子に関する情報はない.
抗血清を用いた古典的な血清型別では,スライド凝集
試験108,133)及び共凝集試験110−112)等が分離株の簡易・迅
速な方法として広く用いられている.血清型別には,高 力価で特異性の高い血清型別用抗血清が必要であるが,
市販されていない.そのため通常は ホルマリン不活化 した各血清型参考株の全菌体をウサギに免疫して作製し た自家製の抗血清が用いられている.しかし,この方法 で作製した抗血清中には,血清型特異抗原に対する抗体 だけでなく,共通抗原を含む様々な菌体抗原に対するポ リクローナルな抗体を含有すると考えられる.そのた め,異なる血清型間で交差反応がしばしば認められ,血 清型別不能となることがある107,113).
交差反応が認められた場合には,間接赤血球凝集反応 法(indirect hemagglutination assay(IHA))109)や 免 疫沈降試験111)による確定診断を実施することが推奨さ れている.なお莢膜合成遺伝子の欠失変異によって,ゲ ル内沈降反応(gel immunodiffusion(GID))では型別 できずに型別不能株となる株73)も存在する.
型別用抗血清の入手が難しいこと,凝集反応等の迅速 法での交差反応性や,確定診断の IHA や GID では迅速 性に少し難があること等から,PCR による血清型の同 定法の開発が多数試みられてきている.
5.1.3.A. pleuropneumoniaeのPCRによる血清型の同 定法について(表 2 )
筆者の2016年に刊行された家畜衛生学雑誌の総説69)
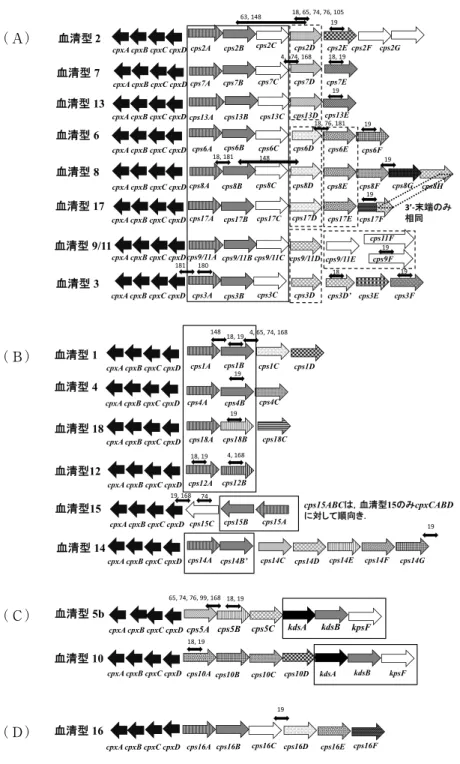
では血清型 1 〜15までの莢膜抗原遺伝子の遺伝子構造の 図を示したが,あらたに 3 つの血清型が増えた.血清型 の同定用 PCR の開発には,血清型特異的遺伝子または 特異的な塩基配列をもったDNA領域の同定が必要であ る.血清型 1 〜18までの莢膜合成遺伝子の塩基配列が 決定されているので,それらの遺伝子構造を図 4 に示し た19−21,69,178).A. pleuropneumoniae の莢膜抗原にはこ れまで 3 種の莢膜タイプがあったが,血清型16はタイプ I〜IIIと共通する遺伝子がなく,全く新しい莢膜タイプ
(タイプIV)であった19,20)(図 4 (D)).一方,血清型17 は,血清型 2 , 3 , 6 , 7 , 8 , 9 ,11及び13が属する タ イ プ I( タ イ コ 酸 グ リ セ ロ ー ル 転 移 酵 素 遺 伝 子 cps17A,グリセロール -3- リン酸シチジル転移酵素遺伝 子 cps17B,機能不明な遺伝子 cps17C 及びタイコ酸合成 酵素遺伝子 cps17D が存在する)19)(図 4 (A)),血清型 18は,血清型 1 ,4 ,12,14及び15が属するタイプ II(莢 膜のリニアバックボーンを構築するリン酸結合に関与す る莢膜多糖リン酸転移酵素 cps18A が存在する)に属し 株(30株),反芻獣及び馬由来の A. lignieresii のほとん
ども(46/51株)陽性を示すほか,豚由来の A. minor
(2/13株)及びA. indolicus(1/8株)でも陽性を示した.
さ ら に Fittipaldi ら は,(i) arbitrary PCR 150),(ii)
omlA PCR 143),(iii) dsbE PCR 30),(iv) aroA PCR 58),
(v) apxIVA nested PCR 144),(vi) cps-cpx multiplex PCR 99),(vii) omlA multiplex PCR 52)及び(viii)海外 で市販されている multiplex PCRキット(Adiavet App PCR test; Adiagène) の 8 種のA. pleuropenumoniae用 PCR について,扁桃のバイオプシーサンプルを用いた 比較試験を行った41).その結果,omlA PCR の感度が 93%と最も高く,次いで83%の Adiavet App PCR と続 くと報告している143).
A. pleuropneumoniaeのPCRによる同定または検出法 について,上記に説明した方法以外のものも含めて,表
1 にまとめたので,参照されたい.
5.1.2.A. pleuropneumoniaeの血清型について
A. pleuropneumoniaeは,莢膜の抗原性の違いに基づ き型別される.2002年までは15の血清型の存在が知られ ていた15).しかし,2015年には血清型16 140),2018年に は血清型17(かつては型別不能株と処理されていた)21)
及び血清型18(かつては血清型K2:O7とミスタイピン グされていた)21,86,121)があらたに提唱され,現在まで に18の(莢膜)血清型の存在が報告されている21).しか し,野外株の血清型別を実施すると,しばしば異なる血 清型間での交差反応が認められる.この理由の大きな原 因はリポ多糖(lipopolysaccharide(LPS))によるもの と考えられている.すなわち,血清型1-9-11,血清型3-6- 8-15,血清型4-7-13の LPS の O 多糖抗原の化学構造及び 抗原構造は同一であるか,あるいは類似しているため,
これらの血清型間ではしばしば交差反応が認められると 考えられている13,39,66,67,69,129).このように,莢膜だけ でなく,LPSのO抗原も血清型別に影響を与えるため,
莢膜抗原(K抗原)及びLPSのO抗原の抗原性に基づい たK:O血清型別が提唱されている.血清型 1 〜15まで のK:O血清型については,既報13,39,66,67,69,129)を参照 されたい. 2018年に新しく提唱された血清型17は,ユ ニークな莢膜(K17抗原)合成遺伝子構造を保有してい るが19)(図 4 (A)),血清型 3 , 6 , 8 ,15と同じO抗原 合成遺伝子構造を保有するため K17:O3と言える.一 方,血清型18(かつてはK2:O7とミスタイピングされ ていた)は,ユニークな莢膜(K18抗原)合成遺伝子構 造を保有しているが19)(図 4 (B)),血清型 7 と同じO抗 原合成遺伝子構造を保有するため,K18:O7と言える.
K2とされた理由は不明であるが,おそらくO7抗原を発 現することがミスタイピングの理由であると思われる.
図 4 .A. pleuropneumoniaeの莢膜合成遺伝子の遺伝子構造.(A),Type I;(B),Type II;(C),Type III;(D),
Type IV19,69,178).両側に矢尻がある小さな矢印は,血清型特異的PCRのプライマーで増幅されるおおよその 位置(遺伝子)をあらわす.その上の数字は,文献番号をあらわす.全ての血清型の莢膜合成遺伝子の上流に は,cpxDCBA(莢膜輸送遺伝子)(cps と逆方向.血清型15のみ cps の下流に順方向),さらにその上流には modF(莢膜合成に無関与)(cpsの順方向)が存在する.一方,Type I,III及びIVの下流には,552-555 bp のオープンリーディングフレーム(orf)(cpsの順方向),さらにその下流にはydeN(cpsの逆方向)が存在す る.Type IIの下流には,順方向にpartial lysA及び114bp orfが存在し,さらにその下流には他のTypeと同 様に,ydeN(cpsの逆方向)が存在する.modF,552-555bp orf,partial lysA,114bp orf及びydeNは,莢膜 の合成には関与しないと考えられている.実線のボックス内の遺伝子,各 Type のコア遺伝子(Type I,
cpsABC;Type II,cpsA;Type III,kdsAB-kpsF;Type IV,不明).破線のボックス内の遺伝子,各血清型 間で有意な相同性を示すcps遺伝子.矢印の長さは,遺伝子の長さに基づいてはいない.
(A)
(B)
(C)
(D)
ための PCR 型別法が開発されてきた,筆者も日本での 分離率が高い血清型 1 , 2 ,5 65)や,その後,豪州で最 も良く分離が認められている血清型15の分離例が,我が 国でも増加してきたことから,血清型 1 , 2 , 5 , 7 , 15の同定用multiplex PCRを開発した74).さらに,型別 可能な血清型の数は増え,血清型 1 〜 3 , 5 〜 8 ,10及 び12( 9 種の血清型)同定用のmultiplex PCRが開発さ れた18).さらにその後,血清型 9 と11とを識別できない が,血清型1-8,9/11,10,12,15及び血清型13,14,
ていた19)(図 4 (B)).
莢膜多糖遺伝子を標的にした PCR 以外にも種々の型 別法がいくつか報告されており43,45,52,57),過去の総説 でもそれらの一部を紹介した69).しかし,各血清型の抗原 性は莢膜多糖によって決定されるため,本稿では莢膜多 糖を合成する遺伝子を標的にした血清型の同定用 PCR に絞り,それらを中心に表 2 にまとめた.流行している 血清型は,国及び地域に特徴的であるため, 1 種類また は数種類のその国・地域に関心の高い血清型を型別する
表 2 . A. pleuropneumoniaeの血清型同定用に開発されたPCR法(菌種同定または検出と血清型同定を同時に行える PCRを含む)の一覧a
血清型の同定が
可能な血清型 血清型同定用PCRの標的遺伝子 菌種同定または
検 出 用 PCR の
標的遺伝子a 文献 他菌種での増幅
5 cps
(cps
5
A+cps5
B) cpx 992, 5, 6 cps
(cps
2
D, cps5
A+cps5
B, cps6
D+cps6
E) omlA 762 cps
(cps
2
B+cps2
D) 631, 2, 8 cps
(cps
1
A+cps1
B, cps2
B+cps2
D) cpx 1483 cps
(cps
3
A) apxIV 1803, 6, 8 cps & cpx
(cpxD+cps
2
A, cps6
D+cps6
E, cps8
A+cps8
B) apxIV 1811, 7, 12 cps
(cps
1
B+IRb between cps1
B/cps1
C, cps7
C+cps7
D, cps12
B) omlA 4A. lignieresii 46株中 1 株:
血清型 1 用プライマーでDNA増幅 A. lignieresii 46株中 5 株:
血清型 7 用プライマーでDNA増幅 1, 2, 5 cps
(cps
1
B+IR between cps1
B/cps1
C, cps2
D, cps5
A+cps5
B) 651-3, 5-8, 10, 12
cps & hypothetical protein
(cps
1
B, hypothetical protein downstream of cps2
D, cps9
D’, cps5
B, cps6
D + cps6
E, cps2
D, cps8
A + cps8
B, cps10
A, cps1
A)apxIV 18
1, 5, 7, 12, 15, cps & cpx
(cps
1
B+IR between cps1
B/cps1
C, cps5
A+cps5
B, cps7
C+cps
7
D, cps12
B, cpxD+cpx15
C) omlA 1682, 1/9/11
cps (cps
2
D)lps
(ORF
17
(rhamnosyl transferase gene))105
1, 2, 5, 7, 15 cps
(cps
1
B+IR between cps1
B/cps1
C,cps
2
D, cps5
A+cps5
B, cps7
C+cps7
D, cps15
C) omlA 7416 cps
(cps
16
C+cps16
D) apxIV 201-8, 9/11, 10, 12, 15
cps
(cps
1
B, cps2
E, cps3
F, cps4
B, cps5
B, cps6
F, cps7
E, cps8
F+cps
8
G, cps9
E+cps9
F, cps10
A, cps12
A, cpxD+cps15
C) apxIV 19 13, 14, 16-18 cps(cps
13
E, cps14
G+ydeN, cps16
C+cps16
D, cps17
F, cps18
B apxIV 19a 菌種同定または検出と血清型同定を同時に行えるmultiplex PCRにおける種同定用PCRプライマーの標的遺伝子.
b IR (intergenic region), 2 つの遺伝子間の非翻訳DNA.
16-18を型別可能な PCR が,開発されている19).なお,
血清型 7 と13の莢膜合成遺伝子の塩基配列は,ほぼ同一 であるというドラフトゲノム塩基配列解析の報告があ り178),莢膜合成遺伝子を標的にした PCR による血清型
7 と13の識別は困難であると考えられていた.しかし,
Bosse19)らは,既報の血清型13の莢膜合成遺伝子の配列 に誤りがあることを明らかにした19).さらに,血清型 7 と13の莢膜合成遺伝子の配列の違いを利用して,両血清 型を識別できると報告している.すなわち,血清型 7 と 13のそれぞれのタイコ酸合成遺伝子 cps13D と cps7D の アミノ酸レベルでの相同性は43%であり,cps13E に 至っては,異菌種のGram陽性菌のhypothetical protein と相同性(<35%)を示すユニークなタンパク質をコー ドしている.そのため,cps13E を標的とした PCR 法が 開発された(図 4 (A))
なおZhouら181)の血清型 3 用及び血清型 6 用の両方の PCR プライマーで増幅が認められる新しい非典型的な 遺伝子型の血清型 6 野外株が日本で見つかり71,152),そ の後欧州18)や,米国49)でも同様に,非典型的な遺伝子 型の血清型 6 が見つかっている. Bosseらはその後,そ れらの非典型的な血清型 6 にも対応できる新しい改良型 のPCR法を開発している18).
A. pleuropneumoniaeのPCRによる血清型の同定法に ついて表 2 にまとめたので,参照されたい.
5.1.4.A. pleuropneumoniaeのRTX毒素とPCRによる RTX毒素遺伝子の検出法について
A. pleuropneumoniae は,赤血球,白血球や,マクロ ファージの細胞膜に穴を開け溶解するApx毒素を分泌す る.その機能によって,A. pleuropneumoniaeは血液寒天 平板上で溶血性を示し,上記の免疫に重要な細胞に障害 を与える.そのため,Apx毒素はA. pleuropneumoniae の重要な病原因子であると考えられている42).Apx 毒 素 に は,ApxI,ApxII,ApxIII 及 び ApxIV 毒 素 が あ
る42,66,67,69,141,145).ApxI毒素は非常に強い溶血性及び
細胞毒性をもち,ApxII毒素は弱い溶血性と中程度の細 胞毒性を示す.ApxIII 毒素には溶血能はないが,細胞 毒性が非常に強い42,66,67,69,145).ApxIV毒素の溶血性及 び 細 胞 毒 性 に つ い て は 検 討 さ れ て い な い141).A.
pleuropneumoniae の ApxI 〜 ApxIII 毒素は,豚胸膜肺 炎対策用ワクチンの主成分の一つとして用いられてい る.一方,ApxIV 毒素はワクチン成分には用いられて いないが,下記のように,A. pleuropneumoniae 感染の 診 断 用 抗 原 に 用 い ら れ て い る66,67,69,141).A.
pleuropneumonia は ApxI 〜 ApxIII の 3 種 の RTX 毒 素 のうち, 1 つあるいは 2 つのRTX毒素を分泌する42)(図 1 ).apxIA,apxIIA 及び apxIIIA は,それぞれ ApxI 毒
素,ApxII 毒素及び ApxIII 毒素の構造遺伝子である.
apxIC,apxIIC 及 び apxIIIC は, そ れ ぞ れ ApxI 毒 素,
ApxII 毒素及び ApxIII 毒素を活性化(アシル化)する タ ン パ ク を コ ー ド す る 遺 伝 子 で あ る.apxIBD 及 び apxIIIBD は,それぞれ ApxI 毒素及び ApxIII 毒素を菌 体外に分泌する機能を持った内膜タンパク質をコードす る遺伝子である.ApxII 毒素遺伝子オペロンは,ApxII 毒素の菌体外への分泌に必要な apxIIBD を欠く.しか し,ApxI毒素遺伝子オペロンのapxIBDの産物(ApxIB 及び ApxID)を利用して,ApxIIA 毒素が分泌される
(図 1 ).分泌する Apx 毒素のパターンは各血清型で概 ね保存されており,Apx 毒素の発現及びその Apx 毒素 遺伝子の保有パターンで,血清型を推定できる42,43,52). しかし,典型的な株とはapxの保有パターンが異なる非 典型的な株の存在も報告されている70,72,141,158).血清型 3 は非溶血性で,病原性がないか弱いと考えられている が,apxIBD を欠く(図 1 )ために,ApxII 毒素は分泌 できず,ApxIII 毒素のみ分泌することがその一因であ ると考えられている42).
ApxIII 毒素の白血球溶解性に豚のβインテグリンの CD18が 関 係 す る こ と が わ か っ て お り, そ の た め に ApxIII 毒素の細胞毒性は豚に宿主特異性を示すことが わかっている170).
A. pleuropneumoniae の ApxIV 毒素は ApxI 〜 ApxIII 毒素とは違い,apxICABD 等の RTX 遺伝子で認められ る 4 つの遺伝子からなる典型的な遺伝子構造(図 1 )を 持たない145).ApxIV 毒素の推定アミノ酸配列は,ヒト の髄膜炎菌 Neisseria meningitidis の FrpA 及び FrpC タ ンパク質のアミノ酸配列と有意な相同性を有する145). apxIVの上流にはORF1が存在する.ORF1はapxIVと同 じ mRNA で転写されることがわかっているが,その機 能はまだ解明されていない.apxIV の下流には,apxIV とは一緒に転写されないlacZが存在する.
ApxIV 毒素は in vivo でしか発現しない145).また,
ApxIV 毒素を産生しない株も何株か報告されている が141),ほとんど全ての株は,ApxIV毒素を分泌する.そ のため,ApxIV毒素は,豚胸膜肺炎の血清診断用ELISA 抗原に用いられ141),海外や日本でも体外診断薬として 市販されている.また apxIV は A. pleuropneumoniae の 同定・検出用nested PCRの標的遺伝子に用いられてい る144).apxIV の 塩 基 配 列 に は variation が あ り,
Rayamajhi らは,彼らが設計したプライマーを用いた PCRで増幅されるDNAのサイズは,血清型によって異 な る と 報 告 し て い る((i) 1600 bp( 血 清 型 4 , 9 , 11),(ii) 2000 bp(血清型 6 ),(iii) 2400 bp(血清型 1 , 3 ,12,13,14),2800 bp( 血 清 型 2 , 5 , 7 , 8 ,
5.2.Actinobacillus porcitonsillarum
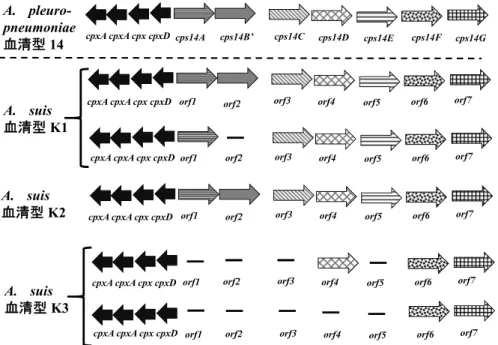
A. porcitonsillarum は, 豚(porci-) の 扁 桃(tonsil)
に生息する菌で,A. pleuropneumoniaeと表現型性状が 似ており,同定の際に注意が必要である48).本菌は非病 原性と考えられているが,日本の食肉衛生検査所で,リ ンパ節炎の豚から分離されたという報告もある123). 5.2.1.A. porcitonsillarumのPCRによる同定・検出法に
ついて(表 3 )
A. porcitonsillarum と近縁な A. pleuropneumoniae と の識別が可能な duplex PCR 開発の報告がある162)(表 3 ), 詳 細 に つ い て は, “5.2.4.A. porcitonsillarum の RTX毒素とPCRによるRTX毒素遺伝子の検出法につい て” を参照されたい.
5.2.2.A. porcitonsillarumの血清型について(表 4 ) A. porcitonsillarum 分 離 株 に お い て,A.
pleuropneumoniae の血清型 1 及び 9 と血清学的交差性 が認められる株が,それぞれ分離されている48).さらに 血清型12抗血清で凝集する株も日本で分離されてい る123),したがって,A. porcitonsillarumには血清型が存 在 す る 可 能 性 が あ る と 推 測 さ れ る が( 表 4 ),
systematic な血清型別を行った研究報告はない.A.
porcitonsillarum は非病原性と考えられており,本菌感 染による重大な被害の恐れは少ない.しかし,本菌に感 染している豚は,北米で開発された LPS を抗原に用い た A. pleuropneumoniae の血清型1-9-11に対する抗体検 出用の ELISA では陽性反応になり,豚胸膜肺炎の正確 10,15)134).しかし,筆者も血清型 6 等の株で試した
経験があるが,増幅されるDNAのサイズが比較的大き いためか,PCR 増幅が認められない株もあった.また 最近の某県での病性鑑定で分離された血清型15の株の場 合も,増幅が認められなかったと聞く(私信).
5.1.5.A. pleuropneumoniaeのPCRによる生物型の型 別法について
A. pleuropneumoniaeには,培地での発育にニコチン アミドアデニンジヌクレオチド(nicotinamide adenine dinucleotide(NAD))を要求する生物型(生物型 1 )と,
NAD を要求しない生物型(生物型 2 )の 2 つの生物型 が存在する.近年,生物型 1 と 2 を識別可能な NAD 合 成に関与する遺伝子nadVを標的としたPCR法が開発さ れている19).この PCR では,PCR 産物が生物型 2 では 増幅されるが,生物型 1 では増幅されない.臨床材料か ら分離されるA. pleuropneuminiaeのほとんどは生物型 1 であり,生物型 2 分離の報告は非常に少ない66,67,69). その理由としては,生物型 2 は生物型 1 よりも病原性が 弱いか75),あるいは NAD 非要求性である生物型 2 の認 知度が低いことから,他の類縁菌と誤認されてきたため と考えられている.A. suisはNAD非要求性であるが,
上記のA. pleuropenumoniae用のnadV PCRは,陰性で ある19).そのため,nadV PCR は A. pleuropneumoniae 生物型 2 と A. pleuropneumoniae 以外の NAD 非要求性 細菌との識別にも利用可能と考えられる.
表 3 .豚から分離されるA. pleuropneumoniae以外の細菌の同定用PCR
菌種 同定用PCRの有無 標的遺伝子 文献a 備考
A. porcitonsillarum なし(備考参照) apsC & apxC 162 左記のプライマーを用いたPCRで,
A. pleuropneumoniaeとの識別可能 他 の 細 菌 同 定 用
PCR で の 非 特 異 増 幅の報告あり
A. pleuropneumoniae dsbE 30 A. pleuropneumoniae同定用のdsbE PCRで陽性
A. suis あり 23S rrn 80
[A]. rossi なし
[P]. aerogenes なし [P.] mairii なし
[H.] parasuis あり 16S rrn 124
16S rrn 9
infB 167
HPS_
219690793
5916S rrn 61(124)
P. multocida あり kmt
1
61(163, 164)kmt
1
+(DNT毒素遺伝子toxA) 101(163, 164)kmt
1
+(DNT毒素遺伝子toxA) 135(163, 164)a 括弧内の数字は,既報の文献のオリジナルな方法をそのまま使用したか,または改変して用いたPCRにおけるオリジナルの文献.