薬理学によって解明された
情報伝達機構
A.生体内情報伝達機構 1.生体内情報伝達機構の概念 2.情報伝達の過程と場 B.薬理学的受容体 の構造と分類 C.細胞内情報伝達系 D. イオンチャネル E. 血管平滑筋・内皮細胞 cf.コミュニケーションの4公理:量・質・関連性・作法 教科書:新薬理学入門(3版)南山堂(2008)参考文献:Katzung, 2014; Goodman & Gilman, 2011; Science, 2001; Hille, 2001; Encyclopedic Ref Mol Pharmacol, 2003
機関リポジトリに「柳澤輝行」のPDFファイルが約70あります。
20180906
(木) 1-3講時 柳澤輝行
柳澤輝行
東北大学・医学部・分子薬理学分野前教授循環器系・神経系薬理学
(1976年卒業)
イオンチャネル、受容体、情報伝達、細胞内Ca2+濃度新薬開発
カルシウム拮抗薬[狭心症・高血圧治療薬] ニコランジル[狭心症治療薬、急性心不全治療薬] ベスナリノン、ミルリノン [急性心不全治療薬] β1アドレナリン受容体刺激薬[心不全治療薬] β2アドレナリン受容体刺激薬[喘息治療薬] β3アドレナリン受容体刺激薬[抗肥満薬、過活動膀胱薬]新薬理学入門(3版) 南山堂 (2008)
カッツング薬理学、イラストレイテッド薬理学
丸善出版休み時間の薬物治療学 講談社
からだの中でも情報が働いている。
• 細胞間情報伝達機構の概念
• 神経系nervous system
• 内分泌 endocrine系
• オータコイド autacoid 系
• 免疫immune系
• 細胞内情報伝達系
内なる声
6細胞間情報伝達機構
intercellular signal transduction mechanism
・免疫系
Exosome Nanotube tunneling
Tunneling nanotubes (TNT) and extracellular vesicle mediated intercellular communication and cargo transfer. Frontiers in Molecular Biosciences 2017(7)
情報伝達の基本課程(図2
−2)
(構造→機能)変化 可逆的なリン酸化反応 転写 trascription 翻訳 translation cAMP cGMP IP3/Ca ProteinsNetwork
Receptor proteins
cell-surface receptors• intrinsic effector function (+)
• Ligand-gated (receptor-operated) ion channels
• receptors with intrinsic enzymatic activity (receptor tyrosine kinases, receptor serine/threonine kinases or receptor guanylyl cyclases)
• intrinsic effector function (-)
• G-protein coupled receptors (GPCRs)
• cytokine receptors → cytosolic protein kinases (Jak/STAT-coupled)
• TNF-receptor superfamily
• IL-1/Toll-like receptors → adaptor proteins (e.g. TRADD, MyD88)
intracellular receptors “nuclear receptors”
Enzymes
Organellae
Cytoplasm
Mechanism of
glucocorticoid
action inside of
cells
Nucleus
heat-shock protein, hsp90 ジンクフィンガー構造 Nuclear pore核内受容体の構造(図2-9)
転写調節因子受容体
Mechanism of activation of the EGF receptor, a
representative receptor tyrosine kinase.
Insulin, PDGF, ANP*, TGFβ, many other
trophic hormones
*ANP: Guanylyl cyclase↑ cGMP↑
Cytokine receptors
STAT dimers then travel to the
nucleus, where they regulate transcription of specific genes.
growth hormones, erythropoietin, interferons, and other regulators of growth & differentiation
細胞膜貫通パターンによるイオンチャネルの型 イオンチャネル内蔵型 M1 M2 P 内向き整流K+ チャネル 電位依存性K+チャネル 4量体→イオンチャネル 4/5量体→イオンチャネル COOH H N2 膜貫通構造 M1 M2 M3 M4 細胞膜 細胞外 細胞内 M1 M2 M3 M4 COOH M1 M2 P(ポア) H N2 2回の遺伝子重複 Ca2+, Na+ チャネル S1 S4 S5 S6 P S2 S3 COOH + + + + S1 S2 S3 S5 S6 P S4 H N2 Inward rectification 内向き整流性(後述) TRP channel
イオンチャネル内蔵型受容体
channel-containing receptor, ligand-gated ion channel, ionotropic receptor; 例 ACh nicotinic receptor
受容体タンパク質は原則として4回細胞膜を貫通する構造(ただし NMDA受容体は3回+ポア構造)をしており、それが4〜5量体で一 つのチャネルを形成する。作動薬は細胞膜の外側で受容体に結合し、 受容体の構造変化を生じる。例えば内蔵するイオンチャネルが開口 して、主にNa+の透過性が亢進すれば、脱分極を生ずる。 この情報伝達の形式は、グルタミン酸受容体(特に NMDA受容体) でも同様である。また、興奮抑制をきたす神経伝達物質であるGABA (γ-aminobutyric acid)のGABAA受容体やグリシンglycine受容体 (ともにCl- チャネルを内蔵、過分極、興奮抑制)でも見られる。作動
薬とは別の部位に結合して、イオンチャネルの活性を変化させる薬 物 (Benzodiazepines) もある(アロステリック効果)。
脱分極depolarization 電位依存性筋細胞膜全体興奮Na+チャネル:開 →収縮
The nicotinic ACh receptor (N
M, N
N)
,
a ligand-gated ion channel.
E/Emax=αX [L]n/(EC50n + [L]n); n=2 (Hill coeffecient) 図 3-18, p77
At binding to and activation of
nicotinic ACh receptor
At binding to and activation of
muscarinic ACh receptor
イオンチャネル内蔵型
Gタンパク質共役型
N
N N
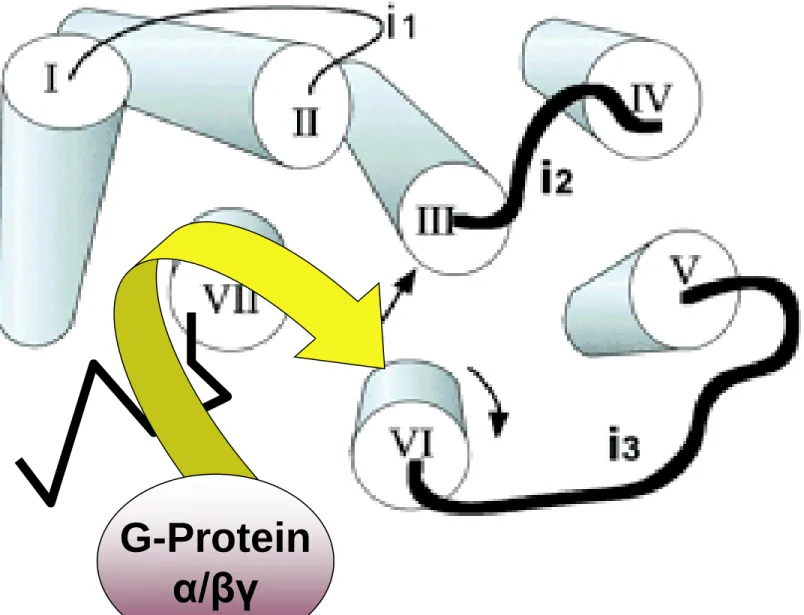
Gタンパク質共役型受容体
G-protein coupled receptor =
GPCR, metabotropic receptor
作動薬の受容体との結合を引き金として、受容体が 膜の中で酵素の活性化を調節するものがある。作動 薬−受容体複合体はGTP結合タンパク質(Gタンパク 質)に結合する。これにより活性化されたGタンパク質 が情報伝達signal transduction を担う種々の酵素や イオンチャネルと結合してそれらの活性の調節をする。Transmembrane topology of a typical serpentine receptor (GPCR)
Illustration of the central core of rhodopsin. The core is viewed from the cytoplasm.
G-Protein α/βγ
Gタンパク質=変換器(transducer)(図2-6)
小分子G-Protein (Rho) MAP kinase (by βγ)
細胞内情報伝達系 Gタンパク質
細菌の毒素を用いてGタンパク質の研究は進展してきた。 コレラ毒素はGsと呼ばれるGタンパク質をADPリボシル 化して、アデニル酸シクラーゼ活性を高め、腸管上皮細 胞中にcAMPを過度に蓄積させる。このcAMPの作用で、 コレラ特有の下痢症状が出現する。 百日咳毒素は、抑制性タンパク質GiをADPリボシル化し て受容体から情報を受け取れないようにしてしまう。Scaffolding b-arrestin Caveolins AKAPs GKAPs SNAREs Components effector substrates protein kinases phosphoprotein phosphatases phosphodiesterases Receptors for biogenic peptides, amines, acetylcholine (M), eicosanoids, Wnt proteins, adhesion protein, odorants, and photons
‘Evolution is molecular tinkering.’ (F. Jacob)
Effectors
Regulated by Gα ↑adenylyl cyclase ↓adenylyl cyclase ↑phospholipase Cβ Regulated by Gβγ KIRcurrents (KGIR) Ca2+ channels Pl-3 kinase G Proteins Classified by α-Subunit αs αi αq αo αtαolf α12/13Ca
2+-phosphoinositide signaling pathway
ER, SR Ca store
Enzyme
Regulation
NO synthases MLCK CaMKⅡ calucineurin PDEs smooth muscle contraction↑[Ca
2+]
i Ca2+ for Contraction striated musclesCalcium Release Channels
IP3R MLCK RyR nNOS eNOS Calcium storage organelle CaM CaM CaM: calmodulin
MLCK: myosin light-chain kinase NO: nitric oxyde
PDE: phosphodiesterase
CaM
Desensitization of the
b
-adrenoceptor.
The response to a b-adrenoceptor agonist (ordinate) versus time (abscissa). The break in the time axis
indicates passage of time in the absence of agonist.
Desensitization & Down-regulation
薬理学
によって解明
された情報伝達機構
D. イオンチャネル ionic channels
•はじめに
<心筋活動電位、イオンチャネル> •イオンチャネルの進化(Dobzhansky)
Nothing in biology makes sense except in light of evolution.
•Ca2 + channels構造と機能 これを介する調節
•K+ channels 構造と機能(多種多彩)
•K+ channel openersの血管拡張機序
教科書:新薬理学入門(3版)南山堂(2008)
参考文献:Katzung, 2011; Goodman & Gilman, 2010; Golan, 2011; Science, 2001; Hille, 2001;
イオン
膜輸送タンパク質の分類
膜
イオンチャネル
輸
チャネル
送
水チャネル
タ
受動輸送
ン
ユニポーター
パ
トランス
シンポーター
ク
ポーター
アンチポーター
質
ポンプ
能動輸送
Na,K ATPase, Na pump ATP ADPIn CNS~120 ~-80mV GABAA
K
+
Cl
-Ca
2+
Na
+
イオン濃度とイオンチャネル
個体を構成する様々なレベル
を理解するために
進化 evolution の観点(生命の歴史)
Nothing in biology makes sense except in
light of evolution. (Dobzhansky)
Hille, 2001
電位依存性Ca
2+チャネルの分子進化系統樹
α1サブユニット相同性 (%) (CLUSTAL W) 100 80 60 40 20 0 高閾値活性型 低閾値活性型 L型(C) L型(D) L型(F) L型(S) P/Q型(A) N型(B) R型(E) T型(G) T型(H) T型(I) C. eleglans C54 電位依存性Na +チャネル DHP感受性 DHP非感受性 非選択性陽イオンチャネル 電位依存性K +チャネル (遺伝子重複 2回) 電位依存性 Ca2+チャネルL型Caチャネルの
分子模型
2
b
outside inside H 2N S-S II III I COOH COOH NH 2 HOOC NH 2 NH 2 COOH
IV1
Varadi (1999) 受容体刺激によるCa チャネル機能の修飾 Ca2+ Caチャネル病L型Ca
2+チャネルはリン酸化を受けて、その電
流量が著明に増加する。
C1 ⇆ C2 ⇆ O交感神経
β受容体
Gs, cAMP
Aキナーゼ
リン酸化
内向き電流 0電流-80 +30 0 5 mN 収縮力 膜電位 (mV) 0.1 mM 0 200 ms 1 mM [Ca2+] i Ca トランジェント 弛緩亢進効果 Ca感受性低下作用 コントロール(点線) 弛緩時 [Ca2+] i ↑ ジギタリス アドレナリン
Ca拮抗薬の冠血管拡張作用に対する5種類の心機能への影響(洞房結節自動能抑制・房室 結節内伝導抑制・心室内伝導抑制・心室自動能抑制・心室筋収縮力抑制作用)の相対効力 比(選択性)を示す。パターンが正六角形に近ければ、冠血管と5種類の心臓機能のいずれ にもそのCa拮抗薬は選択性がないことを示す。逆に六角形の中心に近ければ近いほど、そ のCa拮抗薬は問題にしている心臓機能に対してよりも、冠血管に選択性が高いことを示す。
Taira N: Am J Cardiol, 1989
Ca拮抗薬とK
+チャネル開口薬の比較
verapamil diltiazem nifedipine
L型Ca チャネルの3状態2+ 活性化(開)状態 Ca2+ Ca2+ DHP Ver DHP Vm= -85 mV 0 mV Ca 結合部 位 (ポア) DHP 結合部位 PAA 結合部位 PAA 静止状態 Ca 電流 不活性化状態 2+ 細胞外 細胞膜 細胞内
DHP Ver DHP -85 mV 0 mV PAA 活性化(開)状態 静止状態 ベラパミル 不活性化状態 薬物結合状態(遮断) フェニルアルキルアミン(ベラパミル)による遮断様式 電位依存性解離
DHP系Ca拮抗薬による遮断様式 DHP Ver 薬物結合状態(遮断) DHP DHP DHP -85 mV 0 mV PAA 活性化(開)状態 静止状態 不活性化状態 DHP DHP DHP DHP DHP高親和性 DHP低親和性 DHP
DHP 系Ca拮抗薬
の血管選択性
膜電位 Vh (mV) Ca チャネル利用率血管平滑筋細胞
Test potential (+10 mV) Vh (mV)I
Ca心室筋細胞
膜電位(mV) Ca チャネル利用率DHP 系Ca拮抗薬の血管選択性
(治療用量では心室筋細胞にほとんど効果がない) Rested Open Inactivated RestedI
Ca興奮分泌連関
Ca
2+channel
Presynaptic inhibition
興奮分泌連関 Ca2+ channel
の抑制
神経のN型Ca2+チャネルはGiタンパク質による抑制を受け ると活性化の速度が遅くなることが特徴です。
神経終末
抑制性神経伝達物質
(
α
2, M
2, μ
受容体)
GABA
B 脱抑制I II III IV Extracellular Intracellular Gb +NH 3 b subunit COO -+NH 3 COO -Gb RIM1 Ca2+流入 1 シナプス小胞 α1サブユニットの機能調節タンパク質
Current Opinion in Neurobiology, 1998;8:351-356
歯学部若森教授 RIM1 (Rab3-Interacting Molecule l)
α1subunit
‘Willing’
Closed Inactivated Extracellular Intracellular Gb‘Reluctant’
Closed Closed Closed Closed Closed Open Gb ‘Willing’ ‘Reluctant’State dependence of Gb modulation of presynaptic Ca2+ channels
Prepulse deinhibition of N-type Ca
2+channel
Gβ1 PP:prepulse I(+PP)/I(-PP) 3 2 1 0 τ(-PP)/ τ(+PP) 3 2 1 0 4 電流値 時定数-NE, noradrenaline; Ado, adenosine; 5-HT, serotonin; SS, somatostatin; ATII, angiotensin II; BK, bradykinin.
7.11 Synaptic Sensitization and Depression Hille 3rd p221 Receptors
Channels
GIRK, Kir3.x
一種の
IPSPEzogabine
Ezogabine (Retigabine ): KCNQ/Kv7 opener, anticonvulsant. Flupirtine: the same mechanism , non-opioid analgesic.
Kir3.x regulated by Gi/o βγ
受容体刺激
Gi/o
βγ
Kir3.x 開口
過分極
・徐拍
・伝導抑制
・鎮痛
・鎮静
M1 M2 M1 M2 P 4量体→イオンチャネル1)K
+チャネルは種々の細胞機能に影響する。
2)K
+チャネルの構造と機能を理解すると、生体
現象や病態の理解が深まる。
3) K
+チャネルチャネル開口薬の作用は過分極が
その機序にある。K
+チャネル開口薬は間接的
なCa拮抗薬ではない。
4)平滑筋での弛緩機序を総合して過分極弛緩連
関と呼ぶ。
Potassium (K
+
) channels
The historical view. Potassium channels, key controllers of
resting and action potentials. Armstrong C: Science 1998;280: 56-57.
deactivation
0 100 2 00 3 00 400 0 mV 膜電位 オーバーシュート: +30mV プ ラ ト ー: +20mV -85 mV 静止膜電 位 再 分 極 膜 電流 イオン濃度、平衡電位、電流の方向と膜電位効果 細胞外 細胞内 平衡電位 方向 効果 Na+ 20 nA 145 mM 10 mM 70 mV 内向き 脱分極 外向き電流 4 mM 150 mM -94 mV 外向き 再分極 過分極 K+ a b c 0.5 nA 心筋の膜電位・膜電流・イオンチャネルの概観 向 内 き電流 2 mM 100 nM 129 mV 内向き 脱分極 C a2+ 2 nA 時間(msec)
チャネル K+ 内向き整流 (a) K+ チャネル 一過性外向き (b) K+ 遅延整流 チャネル (c) Na+チャネル 電位依存性 局所麻酔薬、テトロドトキシン(TTX ) Ca2+チャネル 電位依存性 Ca拮抗薬 K+チャネル 電位依存性 III群抗不整脈薬、テトラエチルアンモニウム(TEA ) 遮断薬 ; KA Ch;KATP 心筋の膜電位・膜電流・イオンチャネルの概観 (Kir3.x, Kir6.x)
2 2 2 O O O 神 経 末 梢 頚 動 脈 洞 神 経 Kチャネル I 型 細 胞 Ca電流 毛細血管 受容体 ドパミン*
頚動脈小体での
P
O2の受容
生理学テキスト 図16-26 PO2 正常 PO2 低下 Na電流 *Transmitter:ACh, Glu 舌咽神経,延髄の孤束核Oxygen Sensing: It’s a Gas!
SCIENCE VOL 306 17 DECEMBER 2004 p2050
ヘム分解系の調節・シグナリング
遊離ヘムはラジカル発生(毒性) HO-1 (誘導型): HO-2(構成型) K+チャネル開口 cGMP生成 血管拡張 イオン輸送 ビリベルジン ヘム Fe Fe CO Fe ビリベルジン還元酵素 ビリルビン(抗酸化作用)HO: Heme oxigenase
ヘム、NO、重金属、酸化ストレスで誘導
転写抑制因子Bach1で発現抑制
シグナルのCO
輸送系、
K
+を再循環
させる系として
胃の壁細胞
,HCl分泌
K
+channel, p.181 (COLUMN7)
尿細管細胞
, Na-K-2Cl再吸収
K
+channel, p.165 (図5-6)
他の
K
+channelが関与する系は?
Bartter’s syndrome
図9-13β細胞におけるインスリン分泌機序とその調節因子 グルコース Ca 2+ Gi Aキナーゼ インス リン インス リン Gs AC PLC Gq IP3 Ca2+ cAMP ATP グリベン クラミド + -+ P 脱分極 + KATP -ATP ACh [K ] + 0↑ + + Ca ストア SU 受容体 + K チャネル活性↓+ + -L型Ca チャネル 2+ (M 3受容体) (b2受容体) (2受容体) GLP-1 ソマトスタチン アドレナリン [小腸L細胞] [D細胞] ジアゾキシド +
Closure of the pancreatic β-cell type KATP channel by ATP and sulfonylurea.Increased ATP caused by glucose metabolism closes the KATP channel by binding to the Kir6.2 subunit (A). Sulfonylureas close the channel by binding to the SUR1 subunit (B). The SUR2 subunit has binding sites for benzamide-derivatives in addition to
sulfonylureas (SU). Although there are four binding sites for the ATP molecule and four binding sites for the sulfonylurea molecule in each channel complex, occupation of only one of these sites by ATP or sulfonylurea is sufficient to close the channel.
Encyclopedic Reference of Molecular Pharmacology, p153
4Kir + 4SUR hetromultimer
血糖値
と
インスリン分泌
の連関機構
K
ATPchannel, p.225 (図9-13)
他にも、
K
ATPchannelが関与する系は?
K
ATPchannelは虚血時の心筋保護効果
を持つ。
p.28; p.136 (
preconditioning
)
K
ATPchannel opener,
minoxidil
は発毛
Kir, 内向き整流 inward rectification
膜電位固定法
voltage clamp
0 膜電位 Vm (mV) 膜電流 内向き電流 外向き電流 平衡電位 EK (-96 mV) 脱分極とともに 膜抵抗が増加するKirチャネル内向き整流性
R1 R2Mg
2
+
polyamine+ + +K+
強
D
E
中
D
S
弱
N
G
整流性
R1
R2
2価以上の陽イオン結合
チャネルに栓をする
K
+電流遮断
(1.5 mM) M1 M2静止膜電位
Resting membrane potential
• The membrane potential during diastole • K+
out concentration: [K+]out, Ke
• Hypokalemia低K血症; ectopic pacemaker generation due to the decrease in gK
• カリウムの2つの効果
(EK vs. PK (gK)に対する細胞外(血清)K+濃度Keの影響
を区別せよ。)
K
+電流 vs. K
+e & E
K低K
+e、低P
K;
高K
+e、高P
K 0 膜電位 Vm (mV) Im 膜電流平衡電位
E
K(-96 mV)
K
+i: 150, K
+e: 4 mM
-40 EK (-72 mV) K+e: 10 mME
K(-108 mV)
K
+e: 2.5 mM
E
Kカリウムの2つの効果;
E
Kと
P
K(g
K)
Cf.
Two-site model for the potassium channel• カリウムの
電気化学的勾配(
カリウムの平衡電位
EK)
で予想される影響だけをもとに,血漿カリウム濃度Ke 変化による心筋の活動電位持続時間,ペースメーカ 調律,不整脈に対する影響を考えると,次々と矛盾が 生じてくる. • 心臓では,血漿カリウム濃度が膜のカリウム透過性PK を変化させる効果がしばしば優位になるためである. 細胞外カリウム濃度を増加させるとPKが増加するとい う性質(カリウムチャネルが本来持っている性質)は, 単純な電気化学的勾配EKが小さくなることとは独立し て働くからである.神経細胞とグリア細胞
astrocyte
細動脈arteriole
毛細血管capillary
endfoot
Glia cells behave as messenger transfers like K+ syphon.
神経活動に伴
う脳局所血流
増加
Inward rectifier K+ currents are absent in cerebral artery myocytes isolated from Kir2.1-/- mice.
[K+]
o=140, [K+]i=132 (mM) Zaritsky et al. (Circ Res. 2000;87:160.)
外向き電流増加 →過分極
K+-induced relaxation is absent in the cerebral artery
from Kir2.1-/- animals.
K+-induced relaxation is absent in the cerebral artery
from Kir2.1-/- animals. Zaritsky et al. (Circ Res. 2000;87:160.)
Neuronal activity increases the blood flow in the active area.
Action potential [K+ ]
膜電位 (mV) -55 -90 収縮刺激・ 高血圧 細胞外 細胞内 K + チャネル Ca 2+ チャネル 血管平滑筋 の緊張 血管径 K+チャネルオープナー 過分極 閉 K+平衡電位 -75 -40 静止時 Ca2+ 細胞膜 脱分極 K+ 開 過分極弛緩連関 K+ K+ K + K+ K+ Ca2+ Ca2+Ca2+Ca2+ Ca2+ 細胞膜 閉 開
E. 血管平滑筋・内皮細胞
• 血管、その構造と機能・病態
• 内皮細胞由来弛緩物質
• 細胞内Ca
2+濃度 [Ca
2+]
iと収縮/弛緩
• 狭心症治療薬Nicorandil (NK hybrid*) から
見た過分極弛緩連関
Endothelium Vasa vasorum Sympathetic nerve (vasomotor)
Structure
of the
coronary
artery
Sm. muscle: α receptor(少) β receptor(多)Two types of nitric oxide (NO) synthesis in endothelium (left, eNOS) and
macrophages (right, iNOS). NO causes vasodilation, and NO free radicals are cytotoxic to microbial and mammalian cells. NOS: NO synthase. (Fig. 3-22,
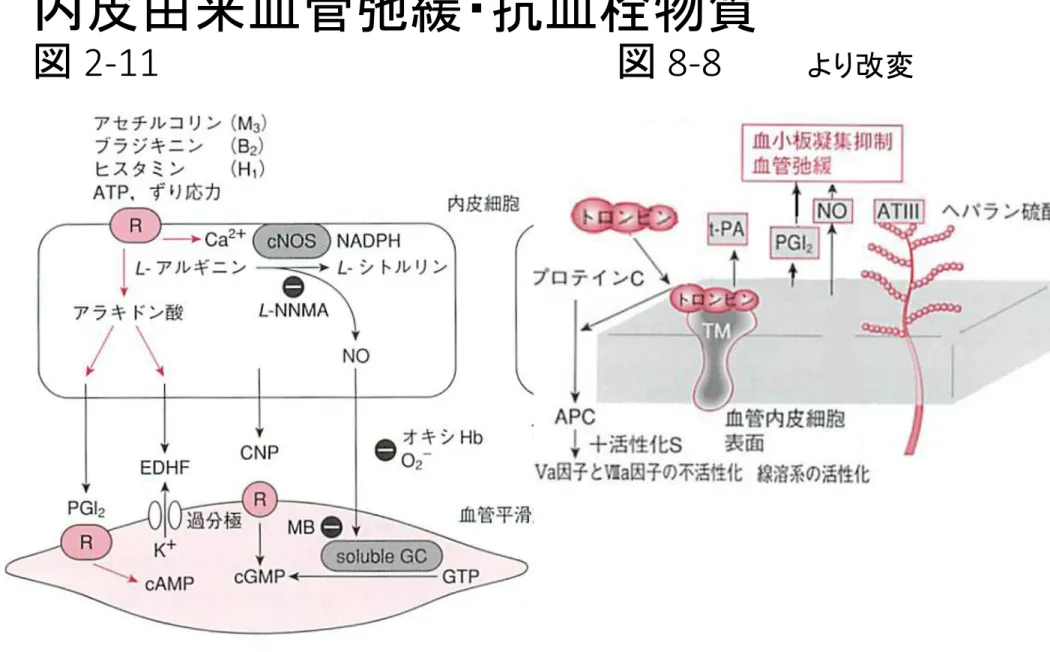
Robbins) 図2-11、 2-16参照
Vascular smooth muscle relaxation & vasodilation
Endothelial stimulation Activation stimulus Macrophage Microbe Endothelium Cytotoxicity胞傷害性 細 NO Ca2+ influx & eNOS activation NO
Virchow’s triad in thrombosis. Endothelial integrity is the single most
important factor. Note that injury to endothelial cells can affect local blood flow and/or coagulability; abnormal blood flow (stasis or turbulence), in turn, can cause
endothelial injury. The factors may act independently or may combine to cause thrombus formation. (Fig.5-12)
収縮因子
血管
拡張因子
筋原性張力
(血流)
内皮由来血管弛緩・抗血栓物質
血管径に応じた3種類のEDRFの役割分担 内皮依存性弛緩反応を生じる3種類のEDRFには血管径に応じた役割分担がある。 Vasodilator PGs は血管径によらずほぼ一定の関与がある。導管血管ではNOの寄与度が 大きく、血管径が細くなるに従ってEDHFの寄与度が大きくなり、抵抗血管ではEDHFによ る弛緩反応が主となる。NOS系は導管血管においては文字通り「NO合成酵素」として働 き、NO産生を介して血管弛緩反応を生じるが、抵抗血管においてはEDHFとしてのH202産 生源となり血管弛緩反応に関与している。Cu,Zn-SOD(copper, zinc-superoxide dismutase;銅・亜 鉛スーパーオキシドジスムターゼ)、sGC(soluble guanylyl cyclase;可溶性グアニル酸シクラーゼ)。
脳血流の自己調節
Autoregulation;
Myogenic control
0
100
200
潅流圧(動静脈圧差
(mmHg)
血流量
(ml/min)
0
500
1000
1500
:過分極、再分極 b 肺細動脈 肺胞内低酸素 低酸素性肺血管収縮 BKCa (電位依存性) Kv and/or BKCa の抑制 抑制 a 全身循環の細動脈 KATP Kv 低酸素、虚血、アデノシン K+チャネル開口薬 血管内圧上昇 脱分極 細胞内Ca2+濃度上昇 筋原性張力(収縮) (電位依存性) 電位依存性Ca2+ チャネルを通して のCa2+流入 血流自己調節 (伸展受容チャネル開口) イオンチャネルによる筋原性張力調節 p52 BKCa (電位依存性) 電位依存性Ca2+ チャネルを通して のCa2+流入 脱分極 細胞内Ca2+濃度上昇 TRPなど
Hypoxic pulmonary vasoconstriction
ventilation-perfusion ratio inequality
A B
Hypoxic (A) Normoxic (B)
air
Pulmonary artery A B Hypoxemia PaO2↓ intrapulmonary shunt↓ Maintain PaO2 (short term) Pulmonary hypertension (long term)alveolus
Pulmonary vein[Ca2+]i
Force of
contraction
(high K contraction)90 mM KCl
(10 min)Verapamil 10-5 M
Ca2+ channel blocker冠動脈
平滑筋
脱分極depolarization Voltage-dependent L-type Ca2+ channel open Ca2+ influx[Ca2+]i
Force of
contraction
+ Verapamil 10
-5 M
Ca2+ channel blocker (10 min)Thromboxan A
2 (agonist contraction)血小板
血栓形成
血管収縮
Ca
in-Fc Relation in Agonist Contraction
TXA
2analogue
-10 0 10 20 30 40 50 60[Ca
2+]
i 0 20 40 60 80 100 120 +Ryanodine (10 min) +Verapamil +0 CaoutKCl
Fc
血管平滑筋アゴニスト収縮機序 p.51
Cav TRPC
Kv
PCI-17- P
冠動脈攣縮
spasm
血管平滑筋の過剰収縮
冠血管狭窄
K
p50
収縮 Ca感受性↑ MLCK Ca Ca2+ から平滑筋収縮まで; Ca感受性をめぐってIP3 DAG PKC Ca2+-CaM MLCK MLCP pCPI-17 pMLCP MLC pMLC RhoK Fasudil Y27632 RhoA GDP RhoA GTP RhoGEF RhoGAP CPI-17 収縮 Staurosporine PLCβ U73122 Telokin pMLCK ↑[Ca2+] i Ca2+ウェーブ
収縮アゴニストの細胞内情報伝達系
参照:『新薬理学入門 第3版』 p.51 Kv channel block ↓ DepolarizationMolecular signal mechanisms of
contraction
of
vascular smooth muscle
HistamineSerotonin TXA2 , ET-1
Depolarization-dependent enhancement
CPI-17 -P
Dissociation of
[Ca2+]i & Force of contraction
KCl (90 mM) contraction
Repolarization vs.
[Ca
2+
]
i
-Fc relation;
Vm
-10mV -34mV -47mV -55mV -42mV -38mVCa desensitization by repolarization
Relationship
between [Ca
2+]
iand force of
contraction
induced by
serotonin ( 10
-6.5M) in the absence
(
○) and presence
of
levcromakalim
( KCO, 10
-5.5M,
●) or nicardipine
(Ca antagonist,
■).
-60 -40 -20 0 20 40 60 80 0 10 20 30 40 50 60 70 80 90 [Ca2+ ]i (%)c
-10 100 F o rc e (% ) 0 0.1 0.3 1.0 2.5 10 0 0.1 0.3 1.0 2.5 10 *** 10-6.5 M 10-5.5 MCa sensitivity ↓
by
KCO
,
hyperpolarization
Canine basilar artery
0-10: [Ca2+]
Control Fura-2 ratio 0.6 0.5 Force (mN) 0 3 1 0 min U46619 Caffeine Cromakalim 10 M-5 0 Ca
Naunyn-Schmied. Arch. Pharmacol. 1992;346:691-700. Biochem. Biophys. Res. Commun. 1992;187:1517-22. Hyperpolarization-relaxation coupling. U46619:thromboxane A2
cromakalim-induced hyperpolarization in porcine coronary artery. (in the absence of [Ca2+]
o)
血管平滑筋の過分極弛緩連関分子機序
過分極 過分極 過分極K
ATP
K
Ca
K
ir
Cav TRPC (Ca2+ ウェーブ) Kv CPI-17 -Pp50
弛緩
Ca感受性↓
Ca拮抗薬
X
Circulation 2011;124:177-1782