去勢抵抗性前立腺癌における OCT1 の機能解析
日本大学大学院医学研究科博士課程 外科系泌尿器科学専攻
山本 慎一郎
修了年 2019 年
指導教員 髙橋 悟
【目次】
【概要】
... 1【略語一覧】
... 3【緒言】
... 61
前立腺癌の疫学、治療
... 62
アンドロゲン受容体
(androgen receptor; AR)の作用機構 ... 73
前立腺癌の進行、去勢抵抗性獲得のメカニズム
... 94 Octamer transcription factor 1 (OCT1)についての報告 ... 11
【目的】
... 14【方法】
... 151
細胞と細胞培地、材料
... 152 Small interfering RNA
(siRNA)
... 163 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H- tetrazolium (MTS) assay ... 16
4 Cell migration assay ... 17
5 Quantitative reverse transcription-polymerase chain reaction (qRT-PCR) ... 17
6 Western blot analysis ... 19
7
マイクロアレイ解析
... 208
統計学的解析
... 20【結果】
... 211 OCT1発現抑制のcastration resistant prostate cancer (CRPC)細胞への影響 .... 21
2 CRPC
進展に関わる
OCT1の直接的な標的遺伝子の同定
... 223 OCT1
標的遺伝子発現抑制の
CRPCモデル
22Rv1細胞への影響
... 244 AR
発現抑制の
CRPC細胞への影響
... 24【考察】
... 261 CRPC
モデル細胞における
OCT1の役割
... 262 CRPC
の進展に関わると想定される
OCT1標的遺伝子の抽出
... 263 OCT1
標的遺伝子の前立腺癌における働き
... 284 OCT1
標的遺伝子による細胞周期の制御、腫瘍の増殖
... 295
今後の研究の展望
... 34【まとめ】
... 36【謝辞】
... 37【図・図説】
... 38【引用文献】
... 65【研究業績】
... 751
【概要】
アンドロゲンおよびアンドロゲン受容体(androgen receptor: AR)を介した アンドロゲンシグナル経路が前立腺癌の増殖・進行に重要な役割を果たしてい る。進行性前立腺癌に対するホルモン療法は有効な治療法だが、次第にホルモ ン療法が効かなくなる去勢抵抗性前立腺癌 (castration resistant prostate
cancer: CRPC)となってしまうことが問題である。Octamer transcription factor 1 (OCT1)は生体内に広く存在するが、前立腺においてはAR
と協調して 転写因子として働く。これまでの前立腺癌における
OCT1の機能解析では、
AR陽性前立腺癌細胞である
LNCaP細胞において最も高発現の
OCT1標的遺伝子 が
acyl-coenzyme A synthetase 3 (ACSL3)であることや、ACSL3が前立腺癌細 胞内でのテストステロン合成を介して前立腺癌の進展に働くことが報告されて
いるが、
CRPCにおける
OCT1の働きは明らかではなかった。本研究では、
OCT1が
CRPCモデル細胞
22Rv1細胞で
AR陽性前立腺癌細胞
LNCaP細胞に比較し て高発現であることを明らかとし、
kinesin family member 15 (KIF15)、 NUF2, NDC80 kinetochore complex component (NUF2)、non-SMC condensin I
complex subunit G (NCAPG)、disks large associated protein 5 (DLGAP5)、anillin actin binding protein (ANLN)の5
遺伝子が
22Rv1細胞における
ARお
2
よび
OCT1の標的遺伝子であることを同定した。
siRNAを用いて
NUF2の発現 を抑制することで
22Rv1細胞の増殖が抑制され、
KIF15、NUF2、DLGAP5の 発現を抑制することで
22Rv1細胞の遊走能が抑制された。加えて
LNCaP細胞 において
ARおよび
OCT1の標的遺伝子として見出されている
ACSL3は
22Rv1細胞においても
ARおよび
OCT1の標的遺伝子であり、
ACSL3を抑制すること
で
22Rv1細胞の細胞増殖能が抑制されることを確認した。本研究により、
CRPCにおける
ARおよび
OCT1の標的遺伝子として新たに見出した
5つの遺伝子は、
ACSL3
とともに
CRPCの進行に関わることが想定され、さらなる研究により前
立腺癌の治療標的や予後予測因子となり得る可能性が示唆された。
3
【略語一覧】
ACSL3: acyl-coenzyme A synthetase 3 AF: activation function
ANLN: anillin actin binding protein AR: androgen receptor
ARE: androgen response element
CAP-D2: chromosome-associated protein D2 CAP-H: chromosome-associated protein H CCP: cell cycle progression
CRPC: castration resistant prostate cancer DBD: DNA-binding domain
DHEAS: dehydroepiandrosterone DHT: dihydrotestosterone
DLGAP5: disks large associated protein 5 E2F: E2F transcription factor
ER: estrogen receptor
ERG: ERG, ETS transcription factor
ETS1: ETS proto-oncogene 1 transcription factor
4
FBS: fetal bovine serum FOXA1: forkhead box A1 FOXP1: forkhead box P1
GAPDH: glyceraldehyde-3-phosphate dehydrogenase GATA2: GATA binding protein 2
GR: glucocorticoid receptor
KIF15: kinesin family member 15 LBD: ligand binding domain
MAB: maximum androgen brockade
MYC: v-myc avian myelocytomatosis viral oncogene homolog NCAPG: non-SMC condensin I complex subunit G
NKX3-1: NK3 homeobox 1 NTD: N-terminal domain
NUF2: NUF2, NDC80 kinetochore complex component OCT1: octamer transcription factor 1
PARP1: poly (ADP-ribose) polymerase 1 PBS: phosphatase buffered saline
PR: progesterone receptor
5
qRT-PCR: quantitative reverse transcription-polymerase chain reaction RUNX1: runt related transcription factor 1
RXR: retinoic receptor
siRNA: small interfering RNA
SMC2: structural maintenance of chromosomes protein 2 SMC4: structural maintenance of chromosomes protein 4 SPC24: spindle pole body component24
SPC25: spindle pole body component 25 SRC 1: steroid receptor coactivator-1
STAT: signal transducer and activatior of transcription TACC2: transforming acidic coiled-coil-containing protein 2 TR: thyroid hormone receptor
UBE2C: ubiquitin-conjugating enzyme E2 C
6
【緒言】
1
前立腺癌の疫学、治療
本邦において、前立腺癌患者数は近年急激な増加傾向にある。本邦における
2013
年の男性癌総数
498,720人のうち前立腺癌は
74,861人、
15.0%で胃癌、肺癌に次いで
3位であった
(1)。
2017年の前立腺癌罹患数予測は
86,100人と予測さ れており
(1)、年間推定罹患数は
2020~2024年には
105,800人に到達し、1 位に なることが予測されている
(2)。2016 年の男性癌死亡総数
219,785人のうち前立 腺癌は
11,803人、5.4%であり、部位別では
6位であった
(1)。本邦の前立腺癌死 亡者数も増加傾向にあり、
2025年~2029 年には約
15,000人に達すると予想さ れている
(2)。
1940
年代に
Hugginsらにより去勢術によって前立腺癌の進行が抑制される
ことが報告されて以来
(3)、進行性前立腺癌治療の第一選択はホルモン療法である。
ホルモン療法としてはアンドロゲン除去療法や抗アンドロゲン薬を使用するこ
とにより前立腺癌の進行が抑制され
(4)、前立腺癌はアンドロゲンにより増殖・進
行することが明らかとなっている。精巣および副腎由来のアンドロゲンを十分
に抑制するために、アンドロゲン除去療法と抗アンドロゲン薬を併用する
Maximum androgen brockade (MAB)療法が本邦の前立腺癌に対するホルモン7
療法の主流となっている
(5)。
進行性前立腺癌に対するホルモン療法は有効な治療法であるが、治療が進む
とホルモン療法が効かなくなる去勢抵抗性前立腺癌
(castration resistant prostate cancer; CRPC)となる(6)。
CRPCに対してはタキサン系の抗癌剤である ドセタキセルやカバジタキセル
(7)、CYP17 阻害剤であるアビラテロン
(8)、新規 抗アンドロゲン薬であるエンザルタミドなどを用いて治療が行われているが
(9)、
次第にそれらの薬物に対しても治療抵抗性を獲得してしまう。そのため、
CRPCに対する新たな治療薬の開発が模索されている状況である。
2
アンドロゲン受容体 (androgen receptor; AR)の作用機構
男性ホルモンであるアンドロゲンは
ARを介して機能する。
ARは核内転写因
子として機能する
110 kDaの蛋白である
(10)。
AR遺伝子は
X染色体上の
q11-12に位置し、8 個のエクソンより構成されている
(11)。AR は
N-terminal domain (NTD)、DNA-binding domain (DBD)、hinge domain、ligand binding domain (LBD)より構成される(12)。
LBDは
C末端に位置し、アンドロゲンと結合するこ
とでアンドロゲンシグナル経路を開始させる。DBD は
ARの核内移行および標
的遺伝子近傍にある
androgen response element (ARE)と呼ばれるゲノム上の配列への結合能を有している。NTD に含まれる
activation function (AF)1や
8
LBD
にある
AF2は
ARによる転写を促進している
(11)。
アンドロゲンの中でも精巣由来のテストステロンは前立腺細胞に取り込まれ
ると
5α還元酵素によりARへの結合力が高い
dihydrotestosterone (DHT)に変換される
(13)。
ARは
DHTなどのリガンドにより活性化されると、核内に移動し
DNA上の
AREに結合する。
ARが
AREに結合すると
steroid receptor coactivator-1 (SRC 1)等の共役因子と相互作用して標的遺伝子の転写が促進される
(14)(図
1)。
AR
による下流遺伝子の発現制御には転写因子も重要な役割を果たしている。
遺伝子はヌクレオソームに存在している。ヌクレオソームは
4つのコアヒスト ンに
DNAが巻き付いた構造をしており
ARの
AREへの結合を妨げているが、
転写因子によりヌクレオソームの構造が変化することで
ARの結合を可能にし ている
(15)。ARE には
non canonical AREと
canonical AREがある。90 ある機 能的
AR結合領域のうち
ARのホモダイマーと結合することで標的遺伝子の転写 が可能となる典型的な
canonical AREはわずか
10%と報告されている(16)。 一方、
ARE
の多くを占める
non canonical AREでは、GATA binding protein 2
(GATA2)やforkhead box A1 (FOXA1)等の転写因子と相互作用することによって標的遺伝子の転写を制御している
(16)。GATA および
FOXAファミリーは
pioneer factor
と呼ばれ、クロマチンが凝集している状態にあっても直接
DNA9
に結合することが可能である
(17)。また、
pioneer factorはクロマチンの凝集を開 く こ と で 他 の 因 子 が
DNAと 結 合 す る こ と を 可 能 に す る
(18,19)。
Octamer transcription factor 1 (OCT1)はpioneer factorの下流で機能する転写因子であ る。前立腺癌細胞においては
GATA2が
ARに結合することで、
OCT1がゲノム 上の
AR結合部位近傍に結合することが可能になり、標的遺伝子の発現を制御 している
(20)。
OCT1
の 他 に
ARと 協 調 す る 転 写 因 子 に は 、
ETS proto-oncogene 1 transcription factor (ETS1), ERG, ETS transcription factor (ERG), forkhead box P1 (FOXP1), NK3 homeobox 1 (NKX3-1), runt related transcription factor 1 (RUNX1)などがある(12)。ETS1 は
ARの活性化以外に、様々な癌の進 展に働くことが報告されている
(21-26)。前立腺癌では、
ETS1は
ARとの相互作用 のほか、NKX3-1 の転写を促進する
(27,28)。NKX3-1 は前立腺癌で頻繁に欠損を
認める癌抑制遺伝子である
(29)。
NKX3-1は
ARの標的遺伝子であると同時に
ARと相互作用する転写因子でもある
(23,25,26)。
3
前立腺癌の進行、去勢抵抗性獲得のメカニズム
進行性前立腺癌に対するホルモン療法は有効な治療法であるが、治療が進む
とホルモン療法が効かなくなる
CRPCとなってしまう
(6)。近年の研究では、
10
CRPC
細胞内ではアンドロゲン濃度が上昇しており、CRPC の増殖・進行にお いてもアンドロゲンシグナル経路が重要な役割を果たしていることが明らかに
なっている
(30)(図2)。去勢抵抗性の原因には、アンドロゲンシグナル経路が関与する機序としては
AR
の発現増加
(31)、AR の点突然変異
(32)や
splice variant(33)によるリガンド特異 性の変化、腫瘍内でのステロイド合成
(34,35)あるいは
AR転写協調因子群の発現 パターンの変化がみられることが報告されている
(36)。
CRPC
細 胞 で は 、
E2F transcription factor (E2F)(37,38)、
v-myc avian myelocytomatosis viral oncogene homolog (MYC)(39,40)や
signal transducer and activatior of transcription (STAT)(41)などの前立腺癌の進行に働く転写協 調因子の発現がホルモン感受性のある前立腺癌細胞に比較して多いことが報告
されており、AR 転写協調因子としての役割を
CRPC細胞で担っていると推察 されている
(29)。
AR
を介さない去勢抵抗性獲得の機序には
glucocorticoid receptor (GR)の発現増加
(42)や前立腺癌細胞の神経内分泌様細胞への変化
(43)などが明らかとなって いる。
このような機序により、前立腺癌の進行や去勢抵抗性を獲得する過程におい
てアンドロゲンシグナル経路の活性化が維持されている。そのため去勢抵抗性
11
前立腺癌においても
ARを介したアンドロゲンシグナル経路は腫瘍の増殖・進 展に重要な働きをしている。
4 Octamer transcription factor 1 (OCT1)についての報告
OCT1
遺伝子は染色体上の
1q24.2に位置し、
18個のエクソンより構成されて いる
(44)。OCT1 は核内受容体の共役因子として働き、AR、GR、progesterone
receptor(PR)
、retinoic receptor (RXR) 、thyroid hormone receptor (TR) 、
estrogen receptor(ER)などを制御する(45)。前立腺では、OCT1 は
ARの転写 因子として
ARの結合部位近傍に結合することで、
ARの転写活性を制御してい る。臨床検体を用いた免疫染色により、
AR陽性の前立腺癌組織における
OCT1の高発現が、前立腺癌特異的生存率低下と相関することが報告されている
(46)。
OCT1
は前立腺癌以外に、胃癌
(47)や結腸癌
(48)、子宮頸癌
(49)でも発現が上昇し、
胃癌や結腸癌では
OCT1の高発現が予後不良と相関することが報告されており、
AR
転写協調因子としての作用以外の働きが示唆されている。
OCT1
は細胞のストレス反応に対する抵抗性を亢進し
(50,51)、OCT1 の欠損し た繊維芽細胞では放射線や抗がん剤に対する感受性が亢進するとの報告がある
(51)
。OCT1 により制御されるストレス反応抵抗性を引き起こす遺伝子には
poly (ADP-ribose) polymerase 1(PARP1)が含まれている(52)。PARP1 は放射線な
12
どの細胞ストレス反応により損傷した
DNAの修復および転写を行うが、進行し た前立腺癌では
ARに結合し
ARによる転写を促進することで前立腺癌の増殖を 促進すると報告されている
(53)。
OCT1
の標的遺伝子としては
AR陽性前立腺癌細胞
LNCaP細胞では、
acyl-coenzyme A synthetase 3 (ACSL3)が最も高発現の遺伝子として報告され
ている
(54)。
ACSL3は正常前立腺組織に比較し、前立腺癌で発現が増加し
(55)、前 立腺癌組織における
ACSL3の高発現が癌特異的生存率低下に相関する
(54)。
ACSL3は前立腺癌細胞内で副腎由来の
dehydroepiandrosterone(DHEAS)か らテストステロンの合成を促進し、前立腺癌の増殖、進行に関わることが報告
されている
(56)(図3)。これまでに、
AR陽性前立腺癌細胞
LNCaP細胞での
OCT1の働きは解析され てきたが、OCT1 の
CRPCにおける働きについては十分明らかではない。前述 のとおり、ホルモン感受性前立腺癌と
CRPCでは転写因子の発現や標的遺伝子 が変化している可能性が考えられる
(36)。そのため、私は
CRPCモデル細胞であ
る
22Rv1細胞における
OCT1の機能や標的遺伝子を解析することが前立腺癌の
去勢抵抗性獲得の機序の解明や新規治療薬の開発につながると考えた。
LNCaP
細胞が未治療の前立腺癌患者のリンパ節転移巣から樹立したのに対
し、22Rv1 細胞はヌードマウスに移植した前立腺癌細胞
CWR22細胞を去勢下
13
に増殖させて細胞株化した由来を持つ
(57,58)。そのため、
22Rv1細胞では
LNCaP細胞同様
AR陽性であるが、アンドロゲン除去処理に抵抗性を有しており、実
臨床の
CRPC症例に近い特性を有している
(59)。また、22Rv1 細胞は
ARの発
現増加や
AR-V7などの
ARの
splice valiantの発現を認めることにより、ホル
モン除去処理に対する抵抗性を有していると報告されている
(57)。
14
【目的】
本研究では
OCT1による
CRPCの進行に関わる分子作用機序の解明を目的と する。
OCT1
による前立腺癌進行に関わる分子作用機序の解析は、これまで
AR陽性
前立腺癌細胞
LNCaP細胞において行われてきた。本研究では
CRPCのモデル
細胞として
22Rv1細胞を用いて
OCT1の機能を解析した。
15
【方法】
1
細胞と細胞培地、材料
ヒト前立腺癌細胞株として、CRPC モデル細胞
22Rv1細胞と
AR陽性前立腺 癌細胞
LNCaP細胞、AR 陰性前立腺癌細胞
DU145細胞を
American Type Culture Collection (Rockville, MD, USA)から購入した。細胞培養は、全て37℃、5% CO2
条 件 下 で 、
10% ウ シ 胎 児 血 清 (
fetal bovine serum; FBS, JPH Bioscience)を添加したRPMI 1640培地(Sigma, St Louis, MO, USA)にて行 った。アンドロゲン刺激前には、ホルモン除去として
72時間
charcoal処理済
みの
2.5%FBSを混合したフェノールレッドフリーの
RPMI1640培地にて培養
を行った。
2 Small interfering RNA
(siRNA)
コ ン ト ロ ー ル
siRNA(
siControl、 標 的 配 列 :
5’-GUACCGCACGUCAUUCGUAUC-3’)(60)、および下記の
siRNAを使用した。
siRNA
は
Sigma Genosys (Tokyo, Japan)より購入した。トランスフェクションには
Lipofectamine RNAiMAX(Life Technologies, Carlsbad, CA, USA)を用いた。
OCT1に対する
siRNA (siOCT1 #A、#B)は10 nMで使用し、それ以外の
16
siRNA
では
5 nMで使用した。
siOCT1 #A,
標的配列:5’-GUGAAGGCUAGGUGAGAAGC-3’
siOCT1 #B,
標的配列:5’-GUGCUAGAUAGGUUUAUAAGU-3’
siAR #1, (Thermo Fisher, S1539) siAR #2, (Thermo Fisher, S1538) siKIF15, (Thermo Fisher, S32546) siNUF2, (Thermo Fisher, S37981) siNCAPG, (Thermo Fisher, S34514) siDLGAP5, (Thermo Fisher, S18914)
siACSL3,
標的配列: 5’-GGAACAAUUUCCGAAGUGUGG-3’
3 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)- 2H- tetrazolium (MTS) assay
96
ウエルディッシュの各ウエルに
22Rv1細胞(500 細胞/ウエル)を入れ、
24
時間後に
siRNAをトランスフェクションした。37℃で培養後に
10μl ずつ
Cell Titer 96 Aqueous One Solution Cell Proliferation Assay kit(Promega, Madison, WI, USA)を付加し、microplate reader(Mithras LB 940; Berthold Technologies, Bad Wildbad, Germany)のabsorbance 490 nm
にて測定した。
各条件は
5ウエルずつ行った。
17
4 Cell migration assay
フィブロネクチン処理をした
Cell Culture Insert with an 8.0-μm pore size PET filter(Falcon, NY, USA)を用いて、各ウエルに5×104個の細胞を入れ、
48
時間培養後にメタノール固定及びギムザ染色を行い、
poreを通過した細胞数 を蛍光顕微鏡(Nikon eclipse TE 2000-U; Nikon, Tokyo, Japan)×100 視野に てランダム
5視野で撮影、検討した。
5 Quantitative reverse transcription-polymerase chain reaction (qRT-PCR)
細胞よりトータル
RNAを
ISOGEN(Nippon Gene, Tokyo, Japan)を用いて 溶解、抽出した。
1本鎖
cDNAは
PrimeScript RT reagent kit(Takara, Kyoto,
Japan)を用いてmRNAより逆転写合成した。
qRT-PCRは
ABI Step One (Life Technologies, Carlsbad, CA, USA)を用いて行い、試薬はKAPA SYBR green mix (KAPA Biosystems, Woburn, MA, USA)を使用して、遺伝子発現量を定量化した。
glyceraldehyde-3-phosphate dehydrogenase (GAPDH)の発現量に対する比として、各遺伝子の発現量をΔΔCt 法により計算した。
ΔΔCt 法は
qRT-PCRによる相対定量法である。目的遺伝子と内在性コント
ロール遺伝子の
Ct値(Threshold Cycle)を求め、目的遺伝子と内在性コント
ロール遺伝子のΔCt 値(Ct 値の差)をもとに、未知サンプル間のΔΔCt 値(Δ
18
Ct
値の差)からサンプル間の目的遺伝子の発現量の比較定量を行う
(61)。 使用したプライマー配列は以下の通りである。
GAPDH
Forward: 5’- GGTGGTCTCCTCTGACTTCAACA -3’
Reverse: 5’- GTGGTCGTTGAGGGCAATG -3’
KIF15
Forward: 5’- CCTTGGAGGTAATGCCAAAA -3’
Reverse: 5’- GCTCACATTTCCTTGGGTGT -3’
NUF2
Forward: 5’- TCCAAATCCAAAGCCTGAAG -3’
Reverse: 5’- GTCATTCACCCGGCAGATAG -3’
NCAPG
Forward: 5’- TGGGAAGTATGCCAGAAATG -3’
Reverse: 5’- TAACCACTGGGCATTCATCA -3’
DLGAP5
Forward: 5’- GCCAAGGGCAATGAAAACTA -3’
Reverse: 5’-TCTTTGGCCTTTGACCTTGT -3’
ANLN
19
Forward: 5’- GGGCTGAACTCAAGATTGGA -3’
Reverse: 5’- GAGGCATTTGAAAGCAGAGG -3’
ACSL3
Forward: 5’- TGCTTTCCGAAGCTGCTATT -3 Reverse: 5’- AAGGCATCTGTCACCAGACC -3
6 Western blot analysis
細胞からの蛋白抽出液の回収は
whole cell lysateを
lysis buffer (50 mM Tris-HCl, [ph8.0], 150 mM NaCl, 1.0% NP40, protease inhibitor cocktail)にて溶解して回収した。蛋白濃度の測定は
PierceTM BCA Protein Assay kit (Life Technologies, Carlsbad, CA, USA)を用いて実施した。それぞれの蛋白30μg を
8% SDS-polyacrylamide gelで 電 気 泳 動 し 、
Immobilon-P Transfer Membrane (Millipore, Billerica, MA, USA)に転写させた。メンブレンは1次抗体と
24時間反応後、2 次抗体と
1時間反応させ、Pierce
TM ECL Plus Western Blotting Substrate (Life Technologies, Carlsbad, CA, USA)を用いて発色させた。
今回使用した抗体は以下の通りである。
Rabbit polyclonal Anti-OCT1 antibody (abcam, ab15112)、rabbit polyclonal
20
anti-AR-antibody (Santa Cruz, H-280)
、
mouse monoclonal anti-β
-actin monoclonal antibody (Sigma, St Louis, MO, USA)を用いた。7
マイクロアレイ解析
22Rv1
細胞より抽出した
RNAを用いて、各
RNAの
RNA integrity number (RIN)が 9.0以上であることを確認した後に、Clariom S Arrays HT, human
(Affymetrix, Santa Clara, CA, USA)によりマイクロアレイを行った。データ解析には
Affymetrix Microarray Suite softwareを用いた。
8
統計学的解析
実験結果は
Student’s t検定を用いて評価した。
p < 0.05を統計学的有意差あ
りとした。
21
【結果】
1 OCT1
発現抑制の
CRPC細胞への影響
CRPC
モデル細胞
22Rv1細胞、AR 陽性前立腺癌細胞
LNCaP細胞、および
AR陰性前立腺癌細胞
DU145細胞より蛋白抽出液を回収し、western blot
analysis
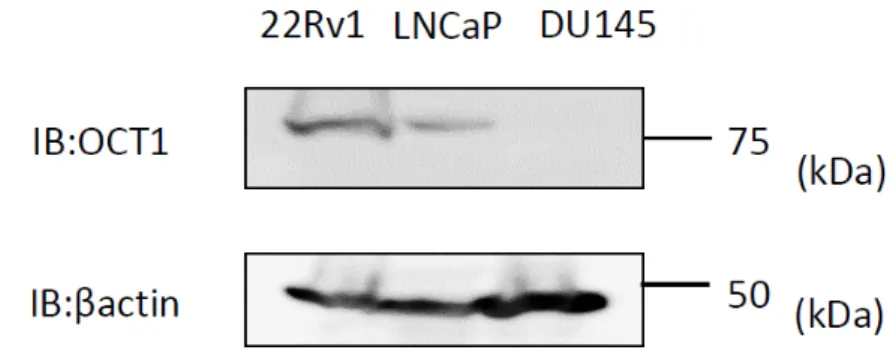
を行いそれぞれの前立腺癌細胞株での
OCT1の発現量を比較した。
22Rv1
細胞で
LNCaP細胞に比較し
OCT1は高発現で、
DU145細胞では
OCT1の発現は検出限度以下であった(図
4)。OCT1
の
CRPC細胞内での機能を解析するため、
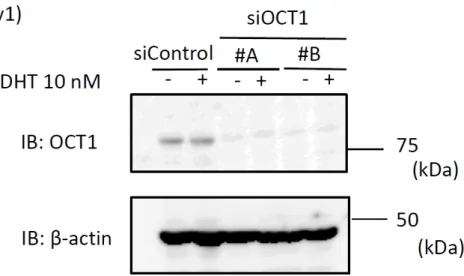
siRNAを用いて
OCT1の発 現を特異的に抑制する実験を行った。まず、
siOCT1 #A、#Bおよび
siControl 10nM
を
22Rv1細胞にトランスフェクションし、
48時間後に蛋白抽出液を回収し
western blot analysis
を行い、OCT1 の発現抑制効果について検討した。いず れの
siOCT1をトランスフェクションした場合も、
siControlと比較し
OCT1の 蛋白レベルでの発現量は顕著に抑制されていた(図
5)。次に、このsiOCT1 #A、#B
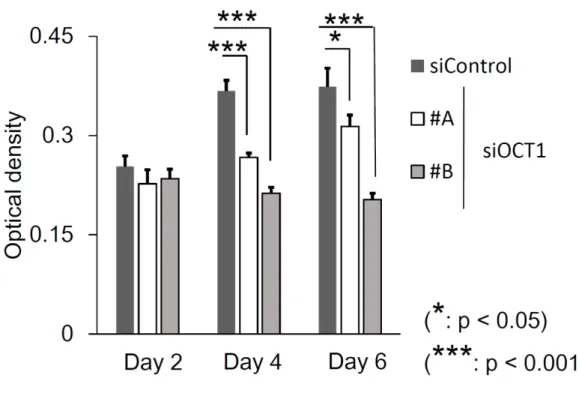
を用いて細胞増殖能を評価する
MTS assayと細胞遊走能を評価する
migration assayを
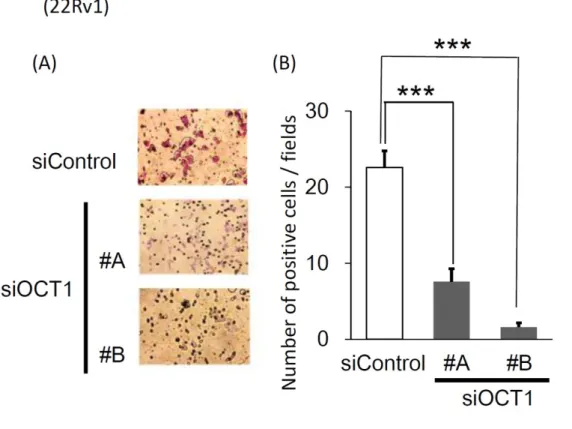
22Rv1細胞に対して実施したところ、OCT1 の発現を抑制
することで細胞増殖能および細胞遊走能が有意に抑制された(図
6、図7)。22Rv1細胞を用いて、
siOCT1 #A、#Bをトランスフェクションすることで、
siControl22
に比較した
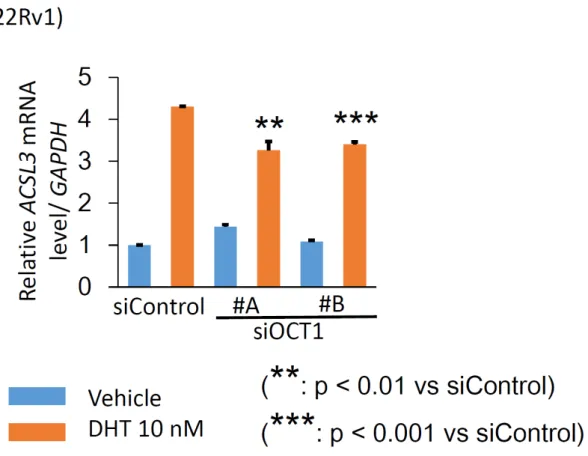
ACSL3の
mRNAレベルでの有意な発現低下を認めた(図
8)。2 CRPC
進展に関わる
OCT1の直接的な標的遺伝子の同定
CRPC
細胞における
OCT1の標的遺伝子を同定するために、siOCT1 #A、
siControl
をそれぞれ
10 nMトランスフェクションし、DHT もしくはエタノー
ル処理を行った
22Rv1細胞より回収した
RNAを用いてマイクロアレイ解析を 行った。当研究室で過去に行った
22Rv1細胞における
OCT1 Chromatin immunoprecipitation-sequencing (ChIP-seq)データ(投稿準備中)を利用し OCT1結合部位が転写開始点より
3 kilobase (kb)以内にある遺伝子に関して、OCT1
の発現抑制による影響を解析した。まず、siOCT1 #A により
siControlと比較し
1/2以下に発現減尐していた遺伝子群を
OCT1標的遺伝子として抽出
した(図
9)。さらにOCT1標的遺伝子のうち、①22Rv1 細胞で高発現(マイクア
ロアレイシグナル>9.0, Log2 変換値) 、②エタノール処理下では
22Rv1細胞で
LNCaP
細胞に比較し発現が上昇し、③DHT 刺激によりエタノールに比較し
2.0倍以上の発現上昇を認めた
33遺伝子を抽出した。②については、以前
LNCaP細胞を用いて発現を検証したマイクアロアレイデータを用いて解析した
(54)。さ
らに、遺伝子発現データベース
Oncomine(www.oncomine.org)内のGrasso et alのデータセットを用いて
(62)、CRPC での発現上昇が予想される
14遺伝子を
23
CRPC
における
OCT1標的遺伝子の候補として選択した(図
10)。次に 14遺伝 子について
OCT1による発現制御を
qRT-PCRにより検討した。siOCT1 #A、
#B
もしくは
siControlを
22Rv1細胞にトランスフェクションし、48 時間後に
DHTもしくはエタノールを加え、
24時間後に抽出した
RNAより逆転写合成し た
cDNAを用いて
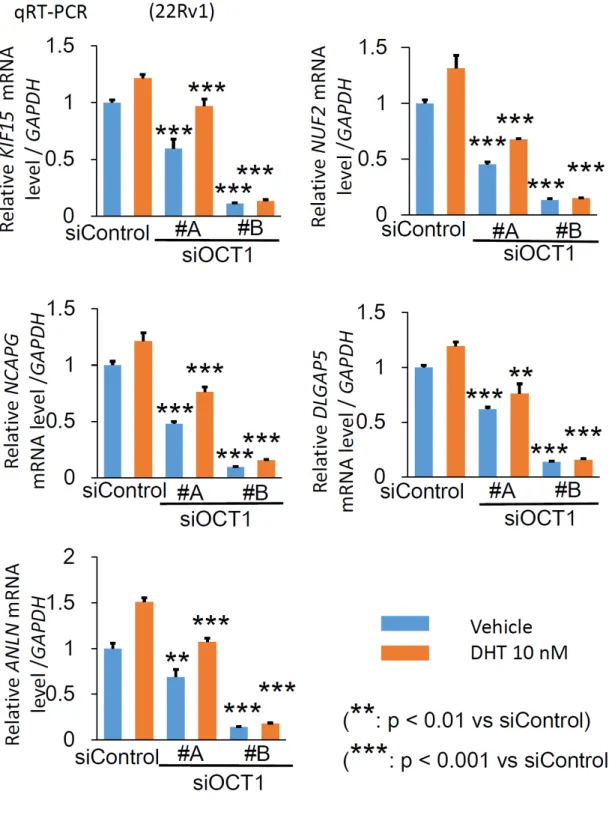
qRT-PCRを行った。その結果、kinesin family member 15
(KIF15)、NUF2, NDC80 kinetochore complex component (NUF2)、non-SMC condensin I complex subunit G (NCAPG)、disks large associated protein 5 (DLGAP5)、anillin actin binding protein (ANLN)の5遺伝子では、
siOCT1 #A、#B
により
siControlと比較し
mRNAレベルでの発現量が有意に減尐すること
を確認し、CRPC 進展に関わると想定される
OCT1標的遺伝子として同定した
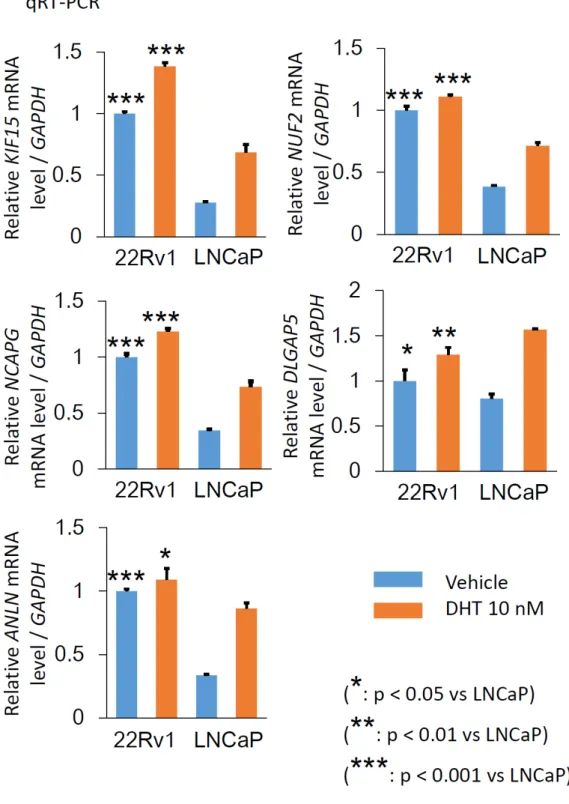
(図11)。同定した5
遺伝子について、22Rv1 細胞で
LNCaP細胞に比較し高発
現であることを確認するため、それぞれの細胞を
48時間培養後に
DHTもしく はエタノールを加え、
24時間後に
RNAを回収し、
qRT-PCRを行った。マイク ロアレイデータから予測されるとおりに
5遺伝子ともにエタノール刺激下では
22Rv1細胞で
LNCaP細胞に比較し高発現であった(図
12)。また、LNCaP細胞 においてもアンドロゲンによる反応性を示した。CRPC 進展に関わると想定さ れる直接的な
OCT1標的遺伝子
5つのうち、前立腺癌での機能解析の報告がな
い
KIF15、NUF2、NCAPG、DLGAP5について前立腺癌細胞での機能解析を
24
行うこととした。
3 OCT1
標的遺伝子発現抑制の
CRPCモデル
22Rv1細胞への影響
KIF15、NUF2、NCAPG、DLGAP5
の前立腺癌細胞内での機能を解析する
ため、 それぞれの発現を特異的に抑制する
siRNAを用いて実験を行った。 まず、
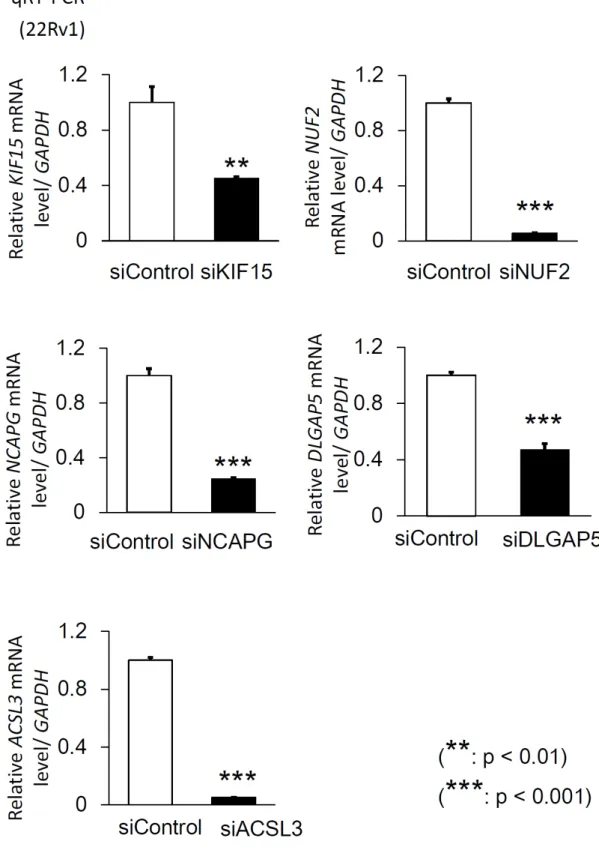
siControl、siKIF15、siNUF2、siNCAPG、siDLGAP5
を
22Rv1細胞にトラン スフェクションし、
qRT-PCRを行い、
siRNAの発現抑制効果について検討した。
いずれの
siRNAをトランスフェクションした場合も
siControlと比較しそれぞ
れの遺伝子の
mRNAレベルでの発現量が有意に抑制されていた(図
13)。次に、これらの
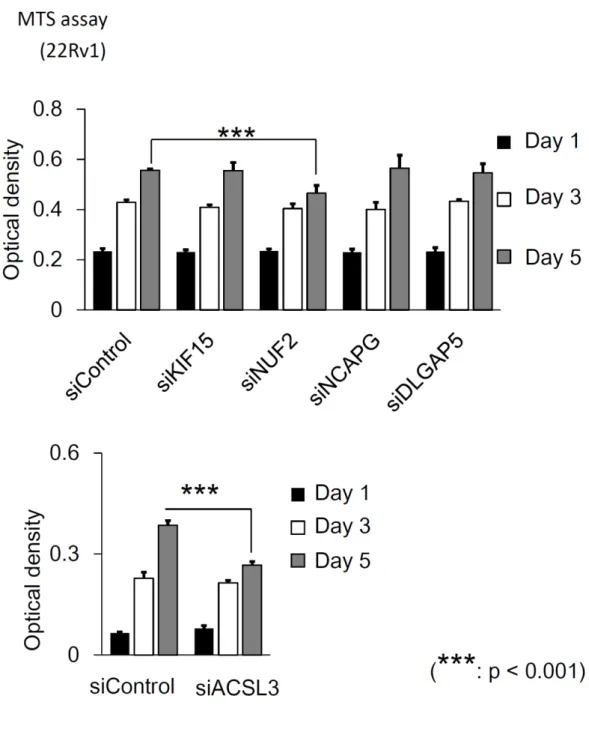
siRNAを用いて、細胞増殖能を評価する
MTS assayと細胞遊走能を
評価する
migration assayを
22Rv1細胞に対して実施した。 細胞増殖能は
NUF2を抑制することで有意に抑制され、細胞遊走能は
KIF15、NUF2、DLGAP5を 抑制することで有意に抑制された(図
14、図15)。細胞増殖能については、22Rv1細胞を用いて
ACSL3を抑制した場合についても検討した。qRT-PCR を行い、
siControl
と比較し
siACSL3により
mRNAレベルでの
ACSL3の発現が抑制さ れ(図
13)、MTS assay では細胞増殖能の有意な抑制を認めた(図
14)。
4 AR
発現抑制の
OCT1標的遺伝子群への影響
25
次に
22Rv1細胞における
ARによる
OCT1標的遺伝子群の発現制御を解析す
るため、
ARを特異的に抑制する
siRNAを用いて実験を行った。まず、
siControlおよび
siAR #1、#2を
22Rv1細胞にトランスフェクションし、western blot
analysisを行い
ARの発現抑制効果につき検討した。siAR #1、#2 ともに
ARの蛋白レベルでの発現を著明に抑制した(図
16)。次にこれらの
siRNAを用いて、
細胞増殖能を評価する
MTS assayを
22Rv1細胞に対して実施した。siAR #1、
#2
ともに
siControlに比較し細胞増殖能を有意に抑制した(図
17)。
そこで、AR を抑制した場合の
CRPC細胞での今回同定した
OCT1標的
5遺 伝子の発現量を検討するため
qRT-PCRを行った。siControl および
siAR #1、#2
を
22Rv1細胞にトランスフェクションした細胞より抽出した
RNAを用いて
qRT-PCR
を行った。siAR #1、#2 により
DHT刺激下での
5遺伝子の
mRNAレベルでの有意な発現量の低下を認めた(図
18)。
26
【考察】
1 CRPC
モデル細胞における
OCT1の役割
本研究ではまず、
AR陽性の
CRPCモデル細胞として
22Rv1細胞を用い、
AR陽性の
LNCaP細胞に比較し
OCT1が高発現であることを明らかにし、OCT1
の高発現が去勢抵抗性獲得に働く可能性を想定した。次に、22Rv1 細胞におい
て
OCT1の発現を抑制することで、細胞増殖能および細胞遊走能が抑制される ことを示し、CRPC においても
OCT1が腫瘍の増殖、進展を促進する可能性が 示唆された。これらのことから、本研究では
22Rv1細胞を用いて、OCT1 の
CRPCにおける機能を解析することとした。
2 CRPC
の進展に関わると想定される
OCT1標的遺伝子の抽出
22Rv1
細胞での
OCT1の機能を解析するため、22Rv1 細胞において
siRNAを用いて
OCT1の発現を抑制し、マイクロアレイ解析を行った。今回は
OCT1結合遺伝子のうち
22Rv1細胞で高発現、
22Rv1細胞で
LNCaP細胞に比較し高
発現、アンドロゲン反応性ありを満たす遺伝子を
CRPC進展に関わる
OCT1標
的遺伝子候補とし、KIF15、NUF2、NCAPG、DLGAP5、ANLN の
5遺伝子
が同定された。
27
22Rv1
細胞、
LNCaP細胞はいずれも
AR陽性前立腺癌細胞株であり
ARに依 存して増殖する
(57)。本研究においても
ARの発現抑制により細胞増殖が抑制さ れることを確認した。しかしながら、
LNCaP細胞はアンドロゲン除去処理によ り増殖を抑えることができるのに対して
(58)、
CRPCモデル細胞である
22Rv1細 胞ではこの処理は十分に有効ではない
(59)。これは
22Rv1細胞においてアンドロ ゲンへの反応性の亢進やアンドロゲン非依存的な
ARの
variantの発現が原因と して考えられる。22Rv1 細胞で
LNCaP細胞に比較して
OCT1が高発現してい ることから、OCT1 が去勢抵抗性に働く機序の解明を目的として
22Rv1細胞に おいて本研究を行った。そのため、CRPC モデル細胞である
22Rv1細胞で高発 現の遺伝子のうち、ホルモン除去処理が効果的である
LNCaP細胞に比較して高 発現の遺伝子を
CRPC進展における重要な
OCT1標的遺伝子の候補になり得る ことを想定した。さらに、
OCT1は前立腺においてはゲノム上の
AR結合部位近 傍に結合し
ARと相互作用することで標的遺伝子の転写を活性化しているので、
アンドロゲン応答性を認め
ARによっても制御されていると考えられる遺伝子
を
CRPC進展に重要な候補遺伝子として選択した。本実験では
22Rv1細胞を用
いて
ARを特異的に抑制することで、今回見出した
OCT1標的
5遺伝子の発現
が抑制され、これらの
OCT1標的遺伝子は
ARによっても制御を受けているこ
とが示された。
28
3 OCT1
標的遺伝子の前立腺癌における働き
新たに見出した5つの
OCT1標的遺伝子のうち、
NUF2では
22Rv1細胞にお いて特異的に発現を抑制することで細胞増殖能が抑制された。ANLN について
は
22Rv1細胞を用いて
siRNAにより発現を特異的に抑制することで細胞増殖
能が抑制されることが報告されている
(63)。以上より、NUF2、ANLN が
OCT1標的遺伝子として
CRPCの増殖を促進している可能性が示唆された。また、
22Rv1
細胞を用いて
KIF15、NUF2、DLGAP5を特異的に抑制させることで細 胞遊走能の減弱を認めたことから、OCT1 がこれらの標的遺伝子の発現を促進
することで細胞遊走能を亢進させて
CRPCの進展に寄与していると考えると興 味深い。
前立腺癌の臨床検体を用いた解析では、NUF2 と
NCAPGについては前立腺 癌転移部で前立腺癌部に比較した
mRNAの発現上昇を認めたことや、それぞれ の遺伝子の発現上昇が癌特異的生存率の増悪と相関し前立腺癌患者における独
立した予後予測因子であることが報告されている
(64,65)。
DLGAP5は臨床検体を 用いた免疫染色により高分化の前立腺癌に比較し低分化の前立腺癌で発現量が
増加することが報告されている
(66)。
ANLNは臨床検体を用いたマイクロアレ
イ解析により、CRPC でホルモン感受性のある前立腺癌に比較し発現が上昇す
る遺伝子として報告がある
(63)。これらの報告は今回同定した
5遺伝子がそれぞ
29
れ前立腺癌の転移や病理学的進行、去勢抵抗性獲得の促進に働くことにより前 立腺癌の進行に働くことと同じ方向のものであった。
ACSL3
は、
LNCaP細胞における
OCT1の標的遺伝子として既に見出され
(54)、 前立腺癌細胞内での
testosterone合成を介して前立腺癌の増殖・進行に働くこ とが報告されている
(56)。ACSL3 は、22Rv1 細胞においても
OCT1の発現を抑 制することで発現量が有意に低下し、OCT1 の標的遺伝子として機能すること
を確認した。また、
22Rv1細胞を用いて、
ACSL3の発現を抑制することで細胞 増殖能が抑制され、
CRPCにおいても
ACSL3が腫瘍の増殖に働くことが考えら れた。
今回
NUF2のみが
siRNAによる発現抑制で
22Rv1細胞の増殖抑制が認めら
れた。これは
NUF2の
siRNAが最も効率よく発現を抑えていることと関連して いるかもしれない。今後
siRNAとその導入や培養の条件、あるいは標的配列の 変更や、過剰発現による検討などにより他の遺伝子についても増殖促進効果を 確認できる可能性が考えられた。
4 OCT1
標的遺伝子による細胞周期の制御、腫瘍の増殖
今回
OCT1標的遺伝子として同定した
5遺伝子はいずれも細胞周期・細胞分
裂に関わることや腫瘍の増殖・促進に関わることが報告されている(図
19)。
30
KIF15
はキネシンスーパーファミリーに属する細胞分裂に働くモーター蛋白
である
(67)。KIF15 は細胞分裂においては微小管上を動くことで染色体の分裂に
働く
(68)。前立腺癌に関する報告はないが、膵癌で
MEK-ERK経路を介して腫瘍 の増殖に働くことや、膵癌や乳癌では
KIF15の高発現が予後増悪と相関するこ とが報告されている
(69,70)。
NUF2
は
NDC80、spindle pole body component 24 (SPC24)、spindle pole body component 25 (SPC25)とNDC80複合体を形成し動原体蛋白として機能 する
(71)。前立腺癌で発現が上昇する
cancer testis antigenの一つとして報告が
ある
(64)。Cancer testis antigen は正常では精巣から分泌され、癌により異所性
に分泌されるという性質を利用して予後予測因子や治療標的として研究されて いる
(72)。CRPC に対するペプチドワクチンを用いた癌免疫療法の標的として臨 床試験が進められている
(73)。
NCAPG
は
structural maintenance of chromosomes protein 2 (SMC2)、structural maintenance of chromosomes protein 4 (SMC4)
、
chromosome-associated protein D2 (CAP-D2)、
chromosome-associated protein H (CAP-H)とともに5量体を形成し、condensin Ⅰ complex として機 能する
(74)。
Condensin complexには
condensinⅠ complex および
condensinⅡ complex があるが、いずれも細胞分裂においては染色体の凝集に働く
(75,76)。
31
DLGAP5
はチューブリンを重合しシート状にすることで微小管の形成を促進
することや
(77)、微小管と動原体の結合を行う動原体蛋白として機能する
(78)。細 胞周期に関わる遺伝子の発現量が前立腺癌の予後予測因子となり得ることにつ いては、cell cycle progression (CCP)スコアを用いた報告がある
(79)。CCP スコ
アは
31個の細胞周期に関わる遺伝子の発現量の平均値をもとに算出する値で、
CCP
が高値であることが前立腺癌の再発率と相関することが報告されている。
今回同定した遺伝子のうち
DLGAP5は
CCPスコアで利用されていた。
ANLN
はアクチンやミオシンの結合タンパクである
anillinを転写している
(80)
。細胞分裂の終期に細胞を二つに分ける収縮環は
anillinがアクチンやミオシ ンと相互作用して形成される
(81)。
細胞周期・細胞分裂を制御する遺伝子の発現量が増加することで細胞の増殖
能の亢進に働き得ると考え
MTS assayを行い評価したが、
22Rv1細胞で腫瘍の 増殖に働いているのは
NUF2のみであった。ANLN についてはこれまでに
22Rv1
細胞を用いて
siRNAにより発現を抑制することで増殖能が抑制されるこ
とが報告されていた
(63)。また、KIF15、
NUF2、DLGAP5は
22Rv1細胞の遊走
能の亢進に働いていたが、これらの遺伝子が細胞の遊走に働く機序の解明につ
いてはこれからの課題である。これらの遺伝子は細胞分裂においてはモーター
タンパクや動原体蛋白として微小管を制御することで働くことが報告されてお
32
り
(67,71,77,78)、細胞の遊走に関しても微小管の運動を介して作用している可能性
も考えられる。今回の検討では
NCAPGが
CRPCモデル細胞の遊走能ならびに 増殖能へ与える影響は明らかとならなかった。一方で
NCAPGの発現増加が前 立腺癌の予後増悪に働く報告があることから
(65)、抗アポトーシス作用や薬剤抵
抗性の獲得、微小環境との相互作用など、別の機序により
CRPCの進行に関与 する可能性が考えられた。
一方で、興味深いことに、AR と細胞分裂については
transforming acidic coiled-coil-containing protein 2 (TACC2)やubiquitin-conjugating enzyme E2C (UBE2C)などのAR
下流遺伝子が報告されており、CRPC における
ARの標
的遺伝子群として細胞分裂に関する遺伝子が重要であると考えられている
(82,83)。
以上より、
OCT1は
ARと協調作用をすることで、細胞分裂に関わる遺伝子の 転写を促進している可能性が示された。本研究は、CRPC において
OCT1およ び
ARがこれらの遺伝子発現を制御することを初めて見出しており、OCT1 の
CRPCにおける新たな知見を得たものといえる。
臨床的には、細胞周期や細胞分裂を標的とする前立腺癌の治療法としては、
ドセタキセルやカバジタキセルによる化学療法が
CRPCに対して有効であるこ
とが知られている。これらはタキサン系の抗癌剤で微小管を阻害し、有糸核分
裂や輸送などの機能を障害することでアポトーシスを誘導している
(84)。今回同
33
定した
5遺伝子のうち、NUF2 と
ANLNは
CRPCでの細胞増殖を促進してお り、前立腺癌細胞、特に
CRPC細胞内における細胞周期及び細胞分裂における 機能の解析から
CRPCの新たな治療法開発や治療標的となり得ることが考えら れた。前立腺癌における報告はないが、繊維芽細胞を用いた報告では
OCT1の 発現により抗癌剤のドキソルビシンに対する感受性が低下することが報告され ている
(52)。この報告では、phosphatase buffered saline (PBS)を加えた場合に 対するドキソルビシンを加えた場合の細胞数を用いてドキソルビシンによる細 胞増殖の抑制を評価しており、OCT1 が欠損した細胞ではコントロールの細胞
に比較し増殖能が抑制されることから
OCT1がドキソルビシンに対する感受性 低下に働くとしている。前立腺癌においても
OCT1が化学療法に対する抵抗性 に働く場合には、OCT1 の発現が増加した前立腺癌に対しては化学療法よりも
OCT1
あるいは今回同定した遺伝子を標的とする治療が有効になる可能性が考 えられた。
限局性前立腺癌に対しては根治的な治療として前立腺全摘除術や放射線治療
が適応になり、低リスクの限局性前立腺癌に対しては過剰治療を避けるために
監視療法も適応となっている
(85)。一方で、進行性前立腺癌に対してはホルモン
療法が有効な治療法ではあるが、多くの症例で
2年から
3年でホルモン療法に
対する抵抗性を獲得し
CRPCとなってしまう
(86)。
CRPCに対しては、本邦では
34
ドセタキセルやカバジタキセルといったタキサン系の抗癌剤を用いた化学療法 や新規抗アンドロゲン薬エンザルタミド、CYP17 阻害薬アビラテロンを用いて 加療を行っているが
(85)、次第にそれらに対しても抵抗性を獲得してしまうこと が問題となっている。本邦の前立腺癌診療ガイドラインにおいて、これらの薬
物の
CRPCに対する使用はいずれも推奨グレードの高い治療とされているが、
薬物の選択に関する明確な基準はない
(85)。最近、個々の症例に対し最適な治療
を分析・選択する
precision medicineが世界的に注目されており
(87)、前立腺癌 治療においてもその導入が望まれる。今後、OCT1 や今回同定した遺伝子が高 発現の症例に対しては、それらの遺伝子を標的とする新規治療薬を選択する
precision medicineの可能性が期待される。
5
今後の研究の展望
今回、
CRPC進展に関わることが想定される
OCT1標的遺伝子として
KIF15、NUF2、NCAPG、DLGAP5、ANLN
の
5遺伝子を同定した。NUF2 と
ANLNは、
CRPC細胞における細胞増殖作用を有すること、
KIF15、NUF2、DLGAP5が細胞遊走能を促進することが示唆された。
NUF2
と
NCAPGについては発現の増加が予後増悪と相関し、前立腺癌の独
立した予後因子であることが報告されている
(64,65)。他の
3遺伝子についても前
35
立腺癌における細胞増殖や転移の促進に働いており、発現の増加が前立腺癌の 予後増悪に働くことが予想される。今までに前立腺癌に関する報告 のない
KIF15
と
DLGAP5については、臨床検体を用いた免疫染色によりそれぞれの遺
伝子の発現量と予後について解析が必要と考えた。
今回、AR および
OCT1が
CRPC細胞内で細胞周期に関わる
5遺伝子の発現
を制御することが明らかとなったが、CRPC 細胞内での役割や細胞分裂、細胞
周期の解析を行うことで新たな治療法の開発や治療標的となり得ることが考え
られた。今後の課題として、細胞分裂における機能解析実験などによる前立腺
癌におけるこれらの遺伝子の役割のさらなる解明が期待される。
36
【まとめ】
本研究は、AR と協調して働く転写因子である
OCT1の
CRPCにおける役割 を解析した(図
20)。OCT1が
CRPCモデル細胞
22Rv1細胞で
AR陽性前立腺癌
細胞
LNCaP細胞に比較して高発現であることを明らかにし、KIF15、NUF2、
NCAPG、DLGAP5、ANLN
の
5遺伝子が
CRPCモデル細胞
22Rv1細胞にお ける
ARおよび
OCT1の標的遺伝子であることを同定した。NUF2、ANLN は
22Rv1細胞の増殖を促進方向に働き、
KIF15、NUF2、DLGAP5は
22Rv1細胞 の遊走能に促進的に関与することが示唆された。既に
LNCaP細胞を用いて
ARおよび
OCT1の標的遺伝子として見出されている
ACSL3は
22Rv1細胞におい ても
ARおよび
OCT1の標的遺伝子であることが明らかとなり、
ACSL3を抑制 することで
22Rv1細胞の細胞増殖能が抑制されることを示した。本研究により、
CRPC
における
ARおよび
OCT1の標的遺伝子として新たに見出した
5つの遺
伝子は、
ACSL3とともに細胞増殖能や遊走能、転移能等を介して
CRPCの進行
に働く可能性が示唆され、さらなる研究により前立腺癌の治療標的や予後予測
因子となり得ることが期待された。
37
【謝辞】
本研究の実施ならびに本論文の作成に当たり、様々なご指導、ご鞭撻を賜り ました日本大学医学部泌尿器科学系泌尿器科学分野主任教授 髙橋 悟博士に 謹んで感謝申し上げます。また、連携大学院生として受け入れ本研究を行うに あたり、親切丁寧なご指導を賜りました東京都健康長寿医療センター研究所研 究部長、日本大学医学部泌尿器科客員教授兼務 井上 聡博士、東京都健康長 寿医療センター研究所 高山賢一博士に心から感謝いたします。実験全般、研 究内容につきご指導いただいた日本大学医学部泌尿器科学系泌尿器科学分野助 教 大日方大亮博士、日本大学医学部内科学系総合内科総合診療医学分野 (現
日本大学歯学部解剖学第
I講座)藤原恭子博士に心から御礼申し上げます。本研
究に当たり、様々なご指導をいただいた日本大学医学部泌尿器科学系泌尿器科
学分野の皆様、東京大学医学部附属病院泌尿器科の皆様、東京都健康長寿医療
センター研究所老化制御システム加齢医学研究の皆様に深く感謝申し上げます。
38
【図・図説】
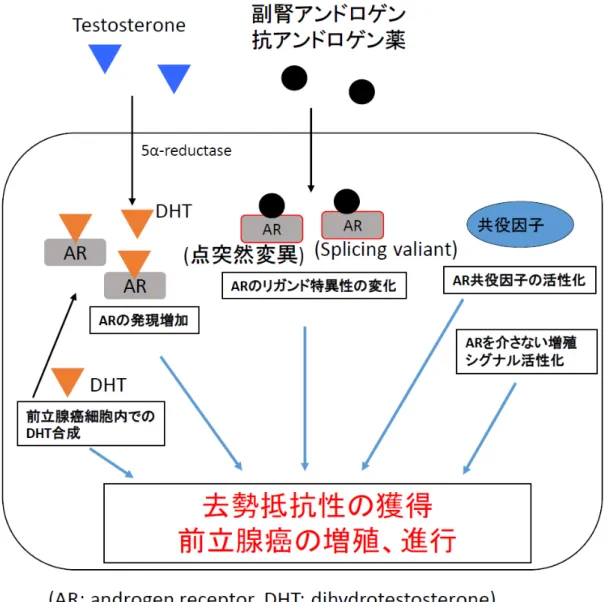
図1 前立腺癌細胞内でのアンドロゲンシグナル経路
テストステロンは前立腺細胞内で
5α還元酵素によりDHTに変換される。
ARは
DHTと結合すると核内に移動し
DNA上の
AREに結合し、共役因子と相互
作用して標的遺伝子の転写を活性化する。その結果、前立腺癌細胞の増殖促進
作用や抗アポトーシス作用に働き、前立腺癌の発生や進行を促進する。
39
図
2前立腺癌の去勢抵抗性獲得のメカニズム
去勢抵抗性の原因には、AR を介する機序としては
ARの発現増加、AR の点
突然変異や
splicing variantによるリガンド特異性の変化、腫瘍内でのステロイ
ド合成、AR 共役因子の活性化などがあげられる。
40
図
3これまでに解析されてきた前立腺癌細胞内での
OCT1の働き
前立腺癌細胞内において、
OCT1はゲノム上の
AR結合部位近傍に結合し、標
的遺伝子
ACSL3の転写を誘導する。また、前立腺癌細胞内で高発現した
ACSL3は前立腺癌細胞内でのテストステロン合成を促進し、前立腺癌の増殖・進行を
促進することが報告されている。
41
図
4前立腺癌細胞での
OCT1発現量の検討
22Rv1
細胞、
LNCaP細胞、
DU145細胞より蛋白抽出液を回収、
western blotanalysis
を実施し、それぞれの細胞での
OCT1の発現量につき検討した。ロー
ディングコントロールとしてβ-actin を用いた。
42
図
5 siRNAを用いた
OCT1の発現抑制効果の検討
22Rv1
細胞にコントロール
siRNA (siControl)および、OCT1に対する
siRNA (siOCT1 #A、#B)をそれぞれ10 nMトランスフェクションし、
48時間 後にエタノールもしくは
DHTを
10 nM投与した。
24時間後に細胞より蛋白抽 出液を回収、
western blot analysisを実施し
OCT1の発現抑制効果を検討した。
ローディングコントロールとしてβ-actin を用いた。
43
図
6 OCT1発現抑制による細胞増殖能への影響
22Rv1
細胞を
96ウエルディッシュの各ウエルに
500細胞ずつ入れ、24 時間
培養後に
siControlおよび
siOCT1 #A、#Bをそれぞれ
10 nMトランスフェク
ションした。
48、96、144時間培養後に
MTS assayを行い、
Microplate readerで吸光度を測定した。結果は平均値±標準偏差 (SD) 、n=5 にて提示した。有
意差検定には
Student’s t検定を用いた( *
: p < 0.05,***
: p < 0.001)。44
図
7 22Rv1細胞に対する
OCT1発現抑制による細胞遊走能への影響
22Rv1
細胞に
siControlおよび、siOCT1 #A、#B をそれぞれ
10 nMトラン
スフェクションし、48 時間培養後に
migration assayを行い細胞遊走能への影
響を検討した。ギムザ染色後に、pore を通過したフィルター下面の細胞数を顕
微鏡
100倍ランダム
5視野にて計測した。結果は平均値±標準偏差 (SD)に
て提示した。有意差検定には
Student’s t検定を用いた( ***
: p < 0.001)。45
図
8 22Rv1細胞を用いた
siOCT1による
ACSL3の発現量の検討
22Rv1
細胞に
siControlおよび、siOCT1 #A、#B を
10 nMトランスフェク
ションし、48 時間後にトータル
RNAを回収、qRT-PCR を行い、ACSL3 の発
現量を検討した。ローディングコントロールとして
GAPDHを用いた。結果は
平均値±標準偏差 (SD) 、n=3 にて提示した。有意差検定には、Student’s t 検
定を用いた( **
: p < 0.01 vs siControl,***
: p < 0.001 vs siControl)。46
図
9 22Rv1細胞における
OCT1標的遺伝子候補の同定
22Rv1
細胞を用いて、
siOCT1 #Aもしくは
siControlを
10 nMトランスフェ クションし、48 時間後に
DHTもしくはエタノール
10 nMを
24時間処理した 細胞より回収したトータル
RNAを用いてマイクロアレイ解析を行った。
siOCT1 #A
により
siControlと比較して
1/2以下に発現が減尐していた遺伝子
群を同定した。
47
図
10 CRPC進展に関わる
OCT1の直接的な標的遺伝子の抽出
22Rv1
細胞で
siOCT1 #Aにより
siControlと比較して発現減尐を認め、
22Rv1細胞で
LNCaP細胞に比較し高発現の
OCT1標的遺伝子
35遺伝子を同定し、ア
ンドロゲンに反応を認めた
33遺伝子を抽出した。遺伝子発現データベース
Oncomine内の
Grasso et alのデータを利用し
(62)、CRPC での発現上昇が予想
される
14遺伝子を選択した。
48
図
11 22Rv1細胞を用いて
siOCT1により発現減尐を認める
5遺伝子を
qRT-PCRにより確認
49
22Rv1
細胞に
siControlおよび、siOCT1 #A、#B をそれぞれ
10 nMトラン スフェクションし、
48時間後にエタノールもしくは
DHTを
10 nM投与し、
24時間後に
RNAを回収した。回収した
RNAを用いて、OCT1 標的遺伝子として 同定した
14遺伝子について
qRT-PCRを行った。ローディングコントロールと
して
GAPDHを用いた。結果は平均値±標準偏差 (SD)、n=3 にて提示した。
有意差検定には
Student’s t検定を用いた( **
: p < 0.01 vs siControl,***
: p <0.001 vs siControl)。
siOCT1 #A、#B
処理により
siControlと比較し有意に発現が減尐する
5遺伝
子を確認した。
50
図
12 22Rv1細胞と
LNCaP細胞での
OCT1標的遺伝子の発現量の比較
22Rv1
細胞および
LNCaP細胞を
48時間後培養後にエタノールもしくは
DHT
を
10 nM投与し、
24時間後に
RNAを回収した。 回収した
RNAを用いて、
51
5
遺伝子につき
qRT-PCRを行った。ローディングコントロールとして
GAPDHを用いた。結果は平均値±標準偏差 (SD) 、n=3 にて提示した。有意差検定に は
Student’s t検定を用いた( *
: p < 0.05 vs LNCaP,**
: p < 0.01 vs LNCaP,***
: p < 0.001 vs LNCaP)。52
図
13 22Rv1細胞に対する
siRNAを用いた
OCT1標的遺伝子の発現抑制効果
の検討
53
22Rv1
細胞に
siControlおよび、
OCT1標的遺伝子に対する
siRNA (siKIF15、siNUF2、siNCAPG、siDLGAP5、siACSL3)をそれぞれ5 nM
トランスフェク
ションし、
48時間後にトータル
RNAを回収、
qRT-PCRを行い、OCT1 標的遺
伝子の発現抑制効果を検討した。ローディングコントロールとして
GAPDHを
用いた。結果は平均値±標準偏差 (SD) 、n=3 にて提示した。有意差検定には
Student’s t検定を用いた( **
: p < 0.01,***
: p < 0.001)。54
図
14 OCT1標的遺伝子発現抑制の
22Rv1細胞に対する細胞増殖能への影響
22Rv1
細胞を
96ウエルディッシュの各ウエルに
500細胞ずつ入れ、24 時間
培養後に
siControlおよび、
siKIF15、siNUF2、siNCAPG、siDLGAP5、siACSL3をそれぞれ
5 nMトランスフェクションした。24、72、120 時間培養後に
MTS assayを行い、microplate reader で吸光度を測定した。結果は平均値±標準
55
偏差 (SD) 、
n=5にて提示した。有意差検定には
Student’s t検定を用いた( ***
:p < 0.001)。
56
図
15 OCT1標的遺伝子発現抑制の
22Rv1細胞に対する細胞遊走能への影響
57
22Rv1