視床と大脳基底核の灰白質体積に対するFK506結合
タンパク質51(FKBP5)遺伝子の一塩基多型と母親の
受容性の交互作用効果
著者
松平 泉
学位授与機関
Tohoku University
学位授与番号
11301甲第19137号
URL
http://hdl.handle.net/10097/00129232
博⼠論⽂
視床と⼤脳基底核の灰⽩質体積に対する
FK506結合タンパク質51(FKBP5)遺伝⼦の⼀塩基多型と
⺟親の受容性の交互作⽤効果
東北⼤学⼤学院医学系研究科医科学専攻 脳科学研究部⾨ 機能画像医学研究分野 松平泉⽬次
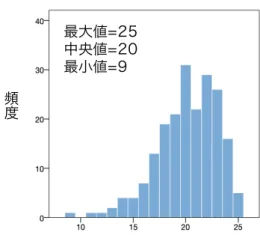
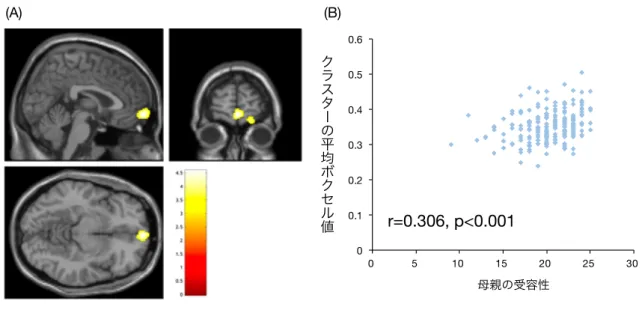
1. 要約 ... 4 2. 研究背景 ... 6 2.1. はじめに ... 6 2.2. 精神障害リスクの神経基盤 ... 7 2.2.1. ⼤脳⽪質−辺縁系回路 ... 8 2.2.2. ⼤脳⽪質−⼤脳基底核−視床−⼤脳⽪質回路 ... 9 2.3. 脳機能・脳構造に対する遺伝要因と環境要因の影響 ... 11 2.3.1. 遺伝要因 ... 11 2.3.1.1. 画像遺伝学 ... 11 2.3.1.2. 遺伝⼦多型による脳機能・脳構造の個体差 ... 12 2.3.2. 環境要因 ... 15 2.3.2.1. 精神障害リスクと養育の関連 ... 15 2.3.2.2. 虐待・ネグレクトによる脳機能・脳構造の個体差 ... 16 2.3.2.3. 規範的な養育による脳機能・脳構造の個体差 ... 18 2.4. 脳機能・脳構造に対する遺伝・環境交互作⽤の影響 ... 20 2.4.1. 画像遺伝学と遺伝・環境交互作⽤の統合 ... 20 2.4.2. FK506 結合タンパク質 51 の遺伝⼦多型と虐待・ネグレクトの交互作⽤による脳機能・ 脳構造の個体差 ... 22 2.4.2.1. FKBP51 の概要 ... 22 2.4.2.2. FKBP5 rs1360780 ... 24 2.4.2.3. FKBP5 rs1360780 と虐待・ネグレクトの交互作⽤ ... 25 2.5. 問題の所在 ... 26 3. 研究⽬的 ... 28 4. 研究⽅法 ... 29 4.1. 研究対象 ... 29 4.2. ポジティブな養育の評価 ... 31 4.3. 遺伝⼦解析 ... 34 4.4. 脳画像撮像 ... 36 4.5. 脳画像前処理 ... 37 4.6. 統計解析 ... 39 5. 研究結果 ... 425.1. 遺伝⼦解析結果 ... 42 5.2. 記述統計 ... 42 5.3. 脳画像解析結果 ... 43 5.3.1. rs1360780 の遺伝⼦型の効果 ... 43 5.3.2. ⺟親の受容性の効果 ... 43 5.3.3. rs1360780 と⺟親の受容性の交互作⽤効果 ... 43 6. 考察 ... 45 6.1. 本研究の主な知⾒ ... 45 6.2. 視床と⼤脳基底核の灰⽩質体積に交互作⽤効果が⾒られたことに関する考察 ... 45 6.3. 先⾏研究の結果との相違に関する考察 ... 47 6.4. C/C 群と T アレルキャリア群における⺟親の受容性と灰⽩質体積の相関の⽅向性の相違に 関する考察 ... 48 6.5. 有意区間における T アレルキャリア群と C/C 群の灰⽩質体積の差に関する考察 ... 51 6.6. rs1360780 の遺伝⼦型による左中帯状⽪質の灰⽩質体積の差に関する考察 ... 52 6.7. ⺟親の受容性と右前頭極の灰⽩質体積の相関に関する考察 ... 53 6.8. 本研究の限界点 ... 54 7. 結論 ... 57 8. ⽂献 ... 58 9. 図 ... 78 図 1. FKBP51 の発現と GR のコルチゾール親和性に対する rs1360780 の影響 ... 78 図 2. ⺟親の受容性の得点ヒストグラム ... 79 図 3. rs1360780 による中帯状⽪質の灰⽩質体積の差 ... 80 図 4. ⺟親の受容性と右前頭極の灰⽩質体積の相関 ... 81 図 5. 左視床, ⼤脳基底核の灰⽩質体積に対する rs1360780 と⺟親の受容性の交互作⽤効果 . 82 10. 表 ... 83 表 1. 記述統計 ... 83 表 2. 灰⽩質体積に対する rs1360780 と⺟親の受容性の交互作⽤効果が有意な脳領域の Montreal Neuroimaging Institute(MNI)座標 ... 84
1. 要約
脳の機能や構造に対する遺伝要因と環境要因の交互作⽤効果を検討することは、 精神障害の発症しやすさ(精神障害リスク)の個体差の理解に有⽤であると⾔われ ている。FK506結合タンパク質51は視床下部‒下垂体‒副腎系のストレス応答機能の 制御に関与するタンパク質であり、その遺伝⼦の⼀塩基多型rs1360780(C/T)のマ イナーアレルTは精神障害リスクを⾼めると考えられている。先⾏研究では、 rs1360780と虐待やネグレクトの被害経験の交互作⽤が脳の機能や構造に関与するこ とが報告されてきた。しかし、虐待やネグレクトは極端に不適切な養育であるた め、それらの知⾒が虐待を受けずに育った群の精神障害リスクの個体差をも説明で きるとは限らない。そこで本研究は、202名の健常⼩児を対象とし、灰⽩質体積に対 するrs1360780とポジティブな養育(⺟親の受容性)の交互作⽤効果を検討した。重 回帰分析の結果、左視床、⼤脳基底核の灰⽩質体積に有意な交互作⽤効果が確認さ れた。下位検定により、C/C群では⺟親の受容性の得点が⾼いほど当該領域の灰⽩ 質体積が⼩さいのに対し、Tアレルキャリア群では⺟親の受容性の得点が⾼いほど灰 ⽩質体積が⼤きいことが明らかとなった。また、⺟親の受容性の得点が全体の70パ ーセンタイル以下の場合において、Tアレルキャリア群はC/C群よりも当該領域の灰 ⽩質体積が有意に⼩さかった。視床と⼤脳基底核は⼤脳⽪質−⼤脳基底核−視床− ⼤脳⽪質回路の中核として精神障害リスクに関わる領域であると⾔われている。本 研究の結果は、精神障害リスクや脳の機能・構造に対する遺伝要因と環境要因の交互作⽤の研究において、虐待やネグレクトのような過度な逆境以外の環境要因に着 ⽬することの重要性を⽰唆していると⾔える。
2. 研究背景
2.1. はじめに ⼈間の精神現象の異常は精神障害であると⾒なされる1)。何を異常あるいは正常と ⾒なすかという根本的な問題も議論が尽きないが、異常と思われる症状も多様を極め ており、診断基準をもってしても明確な診断は困難である1)。2013年から2015年にか けて⾏われた⼤規模な疫学調査によると、⽇本における精神障害の⽣涯有病率は 22.7%であり、国⺠の約5⼈に⼀⼈が⽣涯のどこかで何らかの精神障害に苦しんでい ることになる2)。精神障害は患者と周囲の⼈々を⼼理的苦痛に苛み、個⼈の⼈⽣の質 とも⾔うべきwell-beingに直接の影響を与え得る。有効な治療と同時に、発症を未然 に防ぐことが重要であるが、効果的な予防⽅法の確⽴には⾄っていない3)。 精神障害の予防が困難な理由の⼀つとして、精神障害の発症しやすさ(以下、精神 障害リスク)に個体差があることが挙げられる。精神障害リスクの個体差の存在は、 古くは学習性無⼒感の実験によって裏付けられている。特別な装置を⽤いて⽝に電気 ショックを与えたこの実験では、頭部の横のパネルを叩けば⾃ら電気ショックを⽌め られる逃避可能群と、どれほど⼯夫をしても⽌められない逃避不可能群が⽤意された 4)。その後別の部屋で床から電気ショックが流れてきたとき、逃避可能群はすぐに逃 げ出したが、逃避不可能群は逃げ出そうとせずにその場で電気ショックを受け続けた 4)。逃避不可能群の⽝たちは対処不可能な課題に直⾯したことで、⾃分の⾏動が望ん だ結果に結びつかないことを学習して無気⼒(無⼒感)に陥ったと考えられ、この現 象は学習性無⼒感と呼ばれた。学習性無⼒感が⼈間にも⽣じるかを検証する実験も⾏われた5)。流れてくる⼤⾳響を⾃ら⽌められる群と⽌められない群を⽤意すると、⽌ められない群はやはり無気⼒状態に陥った。しかし、⼤⾳響を⾃分で⽌められない群 のうち、3⼈に⼀⼈は無気⼒にならず、無気⼒になった者たちの中にもすぐに回復す る者と別の課題まで無気⼒を引き摺る者がいることが明らかとなった。学習性無⼒感 はうつ病の危険因⼦であると⾔われている6)。したがってこれらの実験結果から、精 神障害、少なくともうつ病の発症リスクには個体差があると考えることができる。 精神障害リスクの個体差が⽣じる理由、すなわち個体の精神障害リスクが形成され る⽣物学的なメカニズムは未だ明らかにされていない。しかしながら、ストレスへの 応答と適応を担う⼤脳⽪質−辺縁系回路と、⾏動の適応的調整を担う⼤脳⽪質−⼤脳 基底核−視床−⼤脳⽪質回路の機能と構造に対する遺伝要因、環境要因、そして両者 の交互作⽤の影響が、精神障害リスクの基盤であるという仮説が提唱されている7)。 以下では、この仮説を⽀える先⾏知⾒をまとめ、当該分野の現在の問題点を明らかに し、本研究の⽬的に繋げていく。 2.2. 精神障害リスクの神経基盤 機能や構造と精神障害リスクの関連が報告されている脳領域の多くは、⼤脳⽪質− 辺縁系回路と⼤脳⽪質−⼤脳基底核−視床−⼤脳⽪質回路に含まれる。ストレスを知 覚したとき、⽣体内ではホメオスタシスの回復のために⾃律神経系、神経内分泌系、 免疫系などのあらゆるシステムが活動する。これらのストレス応答は⼤脳⽪質−辺縁 系回路の働きによって開始される7)。また、ストレスフルな場⾯では、その状況から 得られる情報や過去の経験の記憶などを統合することにより、適当な⾏動を選択した
り、適応的に⾏動を修正することが必要となる。これらの⾏動調整は、⼤脳⽪質−⼤ 脳基底核−視床−⼤脳⽪質回路の働きによって実現される8)。精神障害とこれらの回 路に含まれる領域の機能・構造の関連について個々の研究の知⾒は必ずしも⼀致して いない。しかしながら、それぞれの回路を構成する領域の機能・構造に個体差があり、 その個体差がストレスと精神障害の関係を媒介し得ることは確かであると考えられ ている7)。 2.2.1. ⼤脳⽪質−辺縁系回路 ⼤脳⽪質−辺縁系回路では、扁桃体をハブとして視床、感覚野、脳幹、視床下部、 島⽪質、海⾺体、前頭前⽪質が結合している。この回路は、環境の中の脅威を認知し て適切な⽣理的・⾏動的応答を⾏うための重要な神経回路であると⾔われている7)。 視床と感覚野から扁桃体基底部に外界の情報が⼊⼒され9)、その情報はさらに扁桃体 中⼼核とレンズ核下拡⼤扁桃体へと送られる10)。扁桃体中⼼核とレンズ核下拡⼤扁桃 体から脳幹への投射は交感神経を覚醒させ、視床下部への投射は神経内分泌系の応答 を引き起こす11)。扁桃体とレンズ核下拡⼤扁桃体は島⽪質や海⾺へも投射する。島⽪ 質への投射は内受容感覚を鋭敏にして⼼拍等の変化を知覚させ12)、海⾺への投射は扁 桃体の活動を引き起こした⽂脈の記憶や想起を促進する13)。⽪質から扁桃体への投射 もある。最も代表的なものは前頭前⽪質からの投射であり、感情制御時に扁桃体の活 動を抑制している14)。 ⼤脳⽪質−辺縁系回路の機能がストレス応答の制御であることから、この回路に含 まれる領域の機能・構造と精神障害の関連を⽰すシステマティック・レビューやメタ
解析論⽂が発表されている。最も多く報告されているのは、⼤うつ病性障害(major
depressive disorder; MDD)・不安障害・⼼的外傷後ストレス障害(post-traumatic stress
disorder; PTSD)などストレスとの関連が特に深い精神障害における、脅威刺激に対 する扁桃体の過活動である15‒17)。ここで⾔う脅威刺激とは主に恐怖表情の画像を指し ている。精神障害患者では恐怖表情画像を⾒た際の扁桃体の活動が中⽴表情画像を⾒ た際よりも有意に⼤きく、脅威に対する扁桃体の過敏性が、臨床的な診断内容によら ず各精神障害間で共通の神経基盤の⼀つではないかと考えられている7,15‒17)。⼤脳⽪ 質−辺縁系回路の構造については、扁桃体活動のように障害間で共通した知⾒は得ら れていないものの、MDD患者では海⾺の体積が⼩さく18)、社交不安障害患者では視 床の体積が⼩さいなど19)、各障害の構造的特徴は明らかにされている。また、PTSD 患者では海⾺・島⽪質・内側前頭前⽪質の体積が⼩さく、脳全体の体積がMDD患者 よりも⼩さいことが報告されている20)。なお、神経症傾向や特性不安など、精神障害 の危険因⼦となる性格特性もまた、⼤脳⽪質−辺縁系回路の機能・構造に差をもたら すことが報告されている。メタ解析の知⾒によると、神経症傾向が強いほど、不快な 情動刺激に対する視床、海⾺、海⾺傍回などの賦活がニュートラルな情動刺激に対す るそれよりも有意に⼤きい21)。また、神経症傾向や特性不安を統合した概念である negative emotionalityが⾼いほど、扁桃体と海⾺傍回の灰⽩質体積が⼤きく眼窩前頭 ⽪質の灰⽩質体積が⼩さいことも⽰されている22)。 2.2.2. ⼤脳⽪質−⼤脳基底核−視床−⼤脳⽪質回路 ⼤脳基底核は、線条体(被殻と尾状核)、淡蒼球、⿊質、視床下核によって構成され
る。線条体は⼤脳基底核の主要な⼊⼒部である。線条体の構造の⼤半を占めるγ−ア ミノ酪酸作動性の中型有棘細胞が、⼤脳⽪質や視床、扁桃体、海⾺などから⼊⼒を受 けると、その情報は淡蒼球、視床下核、⿊質網様部を介して視床へ送られ、さらに視 床から⽪質へ送られる23)。⼤脳⽪質−⼤脳基底核−視床−⼤脳⽪質回路は、それぞれ が起始する領域によって、⾻格運動回路、眼球運動回路、前頭前野回路、辺縁回路の 四種類に分けられる24)。⾻格運動回路は、⼀次運動野・補⾜運動野・運動前野、被殻、 淡蒼球内節と⿊質網様部、視床外腹側核という投射経路を辿る。眼球運動回路は、前 頭眼野・補⾜眼野、尾状核、淡蒼球内節と⿊質網様部、視床前腹側核と視床背内側核 という投射経路を辿る。これらの運動回路は、⾏動の選択、運動の準備、運動の実⾏、 記憶に基づく運動など、運動⾏動の広範な要素に関与すると考えられている。前頭前 野回路は、背外側前頭前⽪質から起始して尾状核頭部、淡蒼球内節と⿊質網様部、視 床前腹側核と視床背内側核を通る回路と、外側眼窩前頭⽪質から起始して尾状核腹内 側部、淡蒼球内節と⿊質網様部、視床前腹側核と視床背内側核を通る回路に分けられ る。前者は複雑な問題に対処するための⾏動反応の調整、後者は共感や社会⾏動に関 与すると⾔われている。辺縁回路は、前帯状⽪質・内側眼窩前頭⽪質・海⾺・扁桃体 から起始して腹側線条体と尾状核、淡蒼球内節と⿊質網様部、視床前腹側核と視床背 内側核、という投射経路を辿り、動機づけ⾏動に関与すると⾔われている。いずれの 回路も、それぞれの視床核から起始領域への投射によって閉じられるループ状の構造 を成している。 多くの先⾏研究により、MDDにおける⼤脳⽪質−⼤脳基底核−視床−⼤脳⽪質回
路の機能異常が報告されてきた。MDD患者は健常者に⽐べて視床と被殻の機能的結 合性が弱く、その傾向は発症年齢が若いほど顕著であるという報告25)があることから、 MDDにおける同回路の機能異常は、⼊⼒部ではなく⼤脳基底核と視床核の連携の異 常によるものであると考えられる。また、MDD患者における⼤脳⽪質−⼤脳基底核 −視床−⼤脳⽪質回路の構造異常を⽰したメタ解析の報告26)によると、MDD患者は 前頭前⽪質、前帯状⽪質、被殻、尾状核の灰⽩質体積が健常者よりも⼩さい。なお、 発症年齢が若年であった群よりも、50歳以上の時点で発症した群の⽅が視床の灰⽩質 体積が⼩さいことも明らかにされている26)。 2.3. 脳機能・脳構造に対する遺伝要因と環境要因の影響 脳は遺伝要因と環境要因の影響を受けて発達する。本項ではまず、精神障害リスク の研究において脳画像と遺伝⼦情報を融合することの利点と、脳機能・脳構造に対す る遺伝要因の効果を⽰した知⾒の例を挙げる。環境要因の「環境」の定義は曖昧であ るが、脳画像研究においては、乳児期以降の養育が脳発達における重要な環境要因の ⼀つとして広く研究されている。したがって本項の後半では養育と精神障害リスクお よび脳機能・脳構造の関連についての知⾒をまとめる。 2.3.1. 遺伝要因 2.3.1.1. 画像遺伝学 脳に対する遺伝要因の効果を検討する研究の多くが、候補遺伝⼦の多型の遺伝⼦型 による脳機能・脳構造の差を調べるものであり、この研究⼿法は画像遺伝学と呼ばれ
ている27‒29)。ここで⾔う候補遺伝⼦とは、神経系に作⽤するタンパク質を⽣成するこ とが神経⽣物学的に明⽩な遺伝⼦であり、その遺伝⼦の転写に影響する⼀塩基多型 (single-nucleotide polymorphism; SNP)などの遺伝⼦構造が遺伝要因として選択さ れる30)。 画像遺伝学は以下の三つの特徴により、精神障害リスクが形成されるメカニズムの 理解に貢献するという利点がある27)。1点⽬は、中間表現型である脳画像を遺伝⼦多 型と結びつけることで、遺伝⼦が⾏動に影響する過程のメカニズムを推定できる点で ある。例えばセロトニントランスポーター遺伝⼦のプロモーター領域における繰り返 し配列の多型(5-HTTLPR)のShortアレルを持つ群では脅威刺激に対する扁桃体の 賦活が⼤きいが31)、これは精神障害患者で脅威刺激に対する扁桃体の過活動が⾒られ ることに共通する。この共通点は5-HTTLPRと精神障害リスクの関係を説明し得る。 2点⽬は、選択された遺伝⼦多型の機能性が明らかになっていれば、脳の個体差の分 ⼦メカニズムを推定できる点である。5-HTTLPRのShortアレルであれば、セロトニ ントランスポーターの転写が少なくシナプスからセロトニンを除去する機能が低下 していること32)が脳の機能や構造に寄与する可能性が考えられる。3点⽬は、画像遺 伝学がモデル動物を⽤いた研究と相性が良い点である。画像遺伝学で明らかになった ことをトランスジェニックモデルマウスやオプトジェネティクスに落とし込むこと で、遺伝⼦多型と脳、⾏動を結びつける分⼦・細胞レベルのメカニズムを検討するこ とができると考えられている27)。 2.3.1.2. 遺伝⼦多型による脳機能・脳構造の個体差
画像遺伝学の⼿法によって脳の機能や構造への影響が最も多く検討されているの は、視床下部‒下垂体‒副腎系(hypothalamic-pituitary-adrenal axis; HPA系)に関わる
タンパク質をコードする遺伝⼦のSNPである7,29)。HPA系はストレスへの応答と適応 を担う中⼼的なシステムである。視床下部室傍核は扁桃体中⼼核、レンズ核下拡⼤扁 桃体、脳幹、海⾺、前頭前⽪質、そして他の視床核から求⼼性の神経⽀配を受けてい る33)。これらの投射によって視床下部室傍核が感覚や情動などの多様な情報を受け取 ることで、HPA系の活動34)が開始される。視床下部室傍核が活動すると副腎⽪質刺激 ホルモン放出ホルモン(corticotropin-releasing hormone; CRH)が分泌され、CRHは 下垂体からの副腎⽪質刺激ホルモン(adrenocorticotropic hormone; ACTH)の放出を 促す。ACTHが副腎を刺激することによりコルチコステロイド(ヒトの場合はコルチ ゾ ー ル ) が 産 ⽣ さ れ る 。 コ ル チ ゾ ー ル が 海 ⾺ で 鉱 質 コ ル チ コ イ ド 受 容 体 ( mineralocorticoid receptor; MR ) や 糖 質 コ ル チ コ イ ド 受 容 体 ( glucocorticoid
receptor; GR)に結合すると35)、これらのリガンド・受容体複合体がCRHとACTHの 分泌を抑制し、結果的にコルチゾールの分泌が抑えられる。コルチゾールが受容体へ の結合を介して更なるコルチゾール分泌を抑制することは、ネガティブフィードバッ ク抑制と呼ばれる34)。ネガティブフィードバック抑制により、⽣体はストレスに適応 しホメオスタシスを回復することができる34)。精神障害患者では以上のストレス応答 とネガティブフィードバック抑制の機能不全があると⾔われている35,36)。 HPA系に関与する遺伝⼦のSNPと⼤脳⽪質−辺縁系回路や⼤脳⽪質−⼤脳基底核 −視床−⼤脳⽪質回路の機能や構造の関連を⽰す知⾒は多数報告されているが、代表
的なSNPとしてCRH受容体遺伝⼦(CRHR1)のrs110402、MR遺伝⼦(NR3C2)の rs5522(rs5522単独、もしくはrs5522とrs2070951のハプロタイプ)、GRの機能を制御 するFK506結合タンパク質51遺伝⼦(FKBP5)のrs1360780が挙げられる29)。CRHR1 rs1104042のマイナーアレルAの保因者は不快な情動を誘起する刺激への腹外側前頭 前野の賦活が⼩さく、感情制御機能が弱い可能性が⽰唆されている37)。NR3C2 rs5522のマイナーアレルGの保因者は脅威刺激に対する扁桃体の賦活が⼤きく、脅威 に対する過敏性が強いと考えられている38)。また、rs5522とrs2070951のハプロタイプ と記憶課題遂⾏中の脳活動の関連を検討した実験では、MRの発現量が多い遺伝⼦型 (rs5522のAアレルとrs2070951のCアレル)を持つ群において、他の群よりも被殻と 尾状核の賦活が⼤きいことが確認されている39)。MRの発現に関わる遺伝⼦多型が、 陳述記憶から⼿続き記憶への切り替えの柔軟性に関与する可能性が⽰唆される。 FKBP5 rs1360780のマイナーアレルTの保因者は背側前帯状⽪質の灰⽩質体積が⼩さ く、PTSD患者においても同領域の体積減少が認められていることから、同SNPと背 側前帯状⽪質の構造の関係がPTSD発症リスクの基盤である可能性が推察されてい る40)。 画像遺伝学は遺伝⼦型を予測変数、脳の機能・構造を媒介変数、⾏動上の精神障害 リスクを⽬的変数とした媒介モデルを仮定している27)。しかし、上記に挙げた先⾏研 究を含め、実際にこの通りの媒介関係を統計的に有意に⽰した研究は少なく41)、「遺伝 ⼦多型がある脳領域の機能・構造に影響し、その脳領域の機能・構造は精神障害リス クに関連する。したがって遺伝⼦多型と精神障害リスクの関係はその脳領域の機能・
構造によって媒介されていると考えられる」という三段論法30)の域を脱していない。 しかしながら、遺伝⼦・脳・⾏動の間に想定される関係性を介して分⼦から神経系ま での多様なレベルで精神障害の病因を考察できるという点で、画像遺伝学は有効な研 究⼿法であると⾔える。 2.3.2. 環境要因 2.3.2.1. 精神障害リスクと養育の関連 精神障害リスクに関与する重要な環境要因の⼀つとして養育が挙げられる。発達⼼ 理学では、⼈間のあらゆる社会情緒的スキルの発達の⼟台は親⼦間の愛着形成にある と考えられている42)。愛着は親と⼦の情緒的絆を意味する⼼理学的概念である。親と の間に安定した愛着関係を築いた⼦どもは、いざという時に親は⾃分を助けてくれる という信念に基づいて⾏動することができる43‒45)。愛着関係を拠り所とした⾃律的な ⾏動の積み重ねにより、レジリエンスなどの社会情緒的スキルが育まれていくと考え られている42)。レジリエンスはストレスフルな状況に適応し良好な⼼⾝状態を回復す る⼼理的機能であり、精神障害の予防に必要な社会情緒的スキルであると⾔われてい る42,46)。不安定な愛着関係がうつ病の発症を予測することもメタ解析論⽂によって報 告され47)、発達⼼理学の愛着理論が裏打ちされたと⾔える。 愛着関係をはじめ、養育にまつわる諸因⼦が⼤脳⽪質−辺縁系回路や⼤脳⽪質−⼤ 脳基底核−視床−⼤脳⽪質回路の発達に影響することを⽰唆する知⾒が多数報告さ れている。当該分野において養育はmaltreatmentとmaltreatment以外に⼤別される。
つまり虐待やネグレクトに該当しない養育は、normative parenting(規範的な養育)
またはparenting in the normal range(正常な範囲内の養育)として括られる48)。何を
もって規範的や正常と⾒なすかは曖昧で、明確なコンセンサスは得られていないが、 本稿では便宜的に「規範的な養育」という表現を⽤いる。規範的な養育の中でも、温 かく受容的な態度はpositive parenting(ポジティブな養育)、拒絶や⼲渉はnegative parenting(ネガティブな養育)に分けられる。しかし両者は完全に分離できるもので はなく、双⽅のバランスによって成り⽴った「虐待・ネグレクトではない養育」が規 範的な養育であると考えられる49)。 2.3.2.2. 虐待・ネグレクトによる脳機能・脳構造の個体差 虐待とネグレクトはその⾏為内容に応じて下位分類される50)。虐待は⾝体的虐待、 性的虐待、⼼理的虐待に分けられる。⼼理的虐待には、恐怖⼼や罪悪感を煽ることで ⼦どもをコントロールすることや、⾔葉によって侮辱すること(⾔語的虐待)、両親 のドメスティック・バイオレンスを⽬撃させることが含まれている。ネグレクトは⼼ 理的ネグレクトと⾝体的ネグレクトに分けられる。⼼理的ネグレクトには⼦どもから の情緒的な要求に応じないことや⼦どもの成熟レベルを超えた⾏動を期待すること などが含まれ、⾝体的ネグレクトは⾷・⾐服・健康・安全など、⼦どもの基本的な欲 求に応じないことを意味する。⼀つ以上の虐待またはネグレクトの経験がうつ病の⼈ ⼝寄与危険度(population attributable risk; PAR)の54%、⾃殺企図のPARの67%を説 明する(すなわち、⼀般⼈⼝から虐待・ネグレクトが無くなればうつ病の発症が54%、
「虐待やネグレクトは慢性的なストレッサーとして⼦どもの脳発達過程に変化を 引き起こし、精神症状が現れる基盤を作る」という仮説のもと、虐待・ネグレクトと 脳の機能・構造の関係を調べる研究が展開されてきた52)。多くの研究の間で⼀貫して 得られている知⾒は、虐待・ネグレクトの被害者において前帯状⽪質・背外側前頭前 ⽪質・眼窩前頭⽪質・海⾺の灰⽩質体積が⼩さいことと、脅威刺激に対する扁桃体の 賦活が⼤きいことである53)。報酬に基づく学習や動機づけといった機能に対する虐待 の影響も報告されている。虐待を受けていた群は、報酬刺激に対する左淡蒼球の活動 が対照群よりも有意に低い54)。また、重度のネグレクトを受けた⼩児において⾒られ る反応性アタッチメント障害の患者では、価値の程度(実験課題の中で与えられる⾦ 銭の額)に依らず、報酬刺激に対する被殻、尾状核、視床の賦活が対照群よりも低い と報告されている55)。以上に挙げた先⾏研究の対象者は、必ずしも精神障害を発症し ていない。しかし、脅威に対する扁桃体の機能亢進や報酬に対する線条体の機能低下 はMDDをはじめとする精神障害で広く報告されていることから15-17, 25, 26)、上述の領 域の機能や構造の異常が、虐待被害経験による精神障害リスクの指標である可能性が ⽰唆される。 虐待の種類によって、その影響を受ける脳領域が異なることも明らかにされている。 精神障害を発症していない⻘年らを対象とした研究によると、⾝体的虐待を受けた群 は背外側前頭前⽪質・眼窩前頭⽪質・島⽪質・腹側線条体の灰⽩質体積が、⾝体的ネ グレクトを受けた群は⼩脳の灰⽩質体積が、⼼理的ネグレクトを受けた群は背外側前 頭前⽪質・眼窩前頭⽪質・線条体・扁桃体・海⾺・⼩脳の灰⽩質体積が、対照群より
も⼩さいことが報告されている56)。また、受けた虐待の中核となる刺激を処理する感 覚野の発達が各種の虐待の影響を受けるという、興味深い知⾒も⽰されている50)。⼼ 理的虐待の⼀部である⾔語的虐待を受けた場合は、成⼈期の聴覚野の灰⽩質体積が⼤ きく57)、聴覚野と前頭葉を結ぶ⽩質線維である⼸状束の拡散異⽅性(fractional anisotropy; FA)が低い58)。⼀⽅、両親のドメスティック・バイオレンスを⽬撃して育 った場合は、成⼈期の⼆次視覚野の灰⽩質体積が⼩さく59)、後頭葉と視覚‒辺縁系経路 を繋ぐ⽩質線維である上縦束のFAが低い60)。また、性的虐待を受けた場合は、成⼈期 の⼀次視覚野と視覚連合野の灰⽩質体積が⼩さく61)、体性感覚野のうち⽣殖器部の感 覚を司る領域の⽪質厚が薄いことが報告されている62)。これらの知⾒から、虐待やネ グレクトによる脳構造の変化はストレスフルな状況への適応によるもので、その状況 の知覚を妨げることで⽣体を保護した結果なのではないかという、新たな仮説が提⽰ されている51)。 2.3.2.3. 規範的な養育による脳機能・脳構造の個体差 虐待やネグレクトの被害者を対象とした⽂献を含まずに⾏われたシステマティッ ク・レビューとメタ解析により、親の温かみの薄さ、拒絶、過⼲渉、両親間の不和が うつ病と不安障害の予測因⼦となることが報告されている63)。したがって、規範的な 養育のバリエーションも精神障害リスクに関与すると⾔える。規範的な養育と脳の機 能や構造の関連を検討した研究は虐待・ネグレクトのそれに⽐べて圧倒的に少なく、 研究間で⼀貫した知⾒は得られていない。しかし、ポジティブな養育が扁桃体や⼤脳 基底核の機能に関与することが、いくつかの先⾏研究によって報告されている。具体
的には、⺟親の養育態度が⽀援的であるほど、⻘年期において脅威刺激に対する扁桃 体の賦活が⼩さい64)。また、⺟親の温かみを感じている⼦どもほど、報酬を失ったと きの尾状核の賦活が⼤きい65)。さらに、親⼦間の愛着形成の基礎となる⺟親の対⼈親 和性が⾼いほど、その⼦どもは報酬刺激に対する腹側線条体と尾状核の賦活が⼤きい 66)。前述の通り、精神障害患者や虐待・ネグレクト被害者は脅威刺激に対する扁桃体 の活動や報酬に対する⼤脳基底核の活動に異常を⽰すという知⾒があることから、こ れらの報告はポジティブな養育が精神障害の防御因⼦として働く可能性を⽰唆して いると⾔える。 脳の構造と規範的な養育の関連についても、発達段階ごとに興味深い知⾒が得られ ている49)。まず、乳児期の⺟⼦間の愛着形成が扁桃体の発達に⻑期的に影響する可能 性が⽰されている。1歳半時点での⺟親への愛着が不安定であった群は、安定してい た群よりも22歳時点での扁桃体の体積が有意に⼤きかった67)。また、1歳半時点での ⺟親への愛着が無秩序的(養育者に対し接近⾏動も回避⾏動も⽰す)であった群にお いて、⼦どもとの交流における⺟親の⾏動が不安定(⼦どもからの要求に適切に応じ られない・⼦どもに対し過度に⼲渉的になるなど)であったほど、29歳時点での扁桃 体の体積が⼤きいことも報告されている68)。学童期・⻘年期における⺟親の養育と⼦ どもの扁桃体の発達の関係についても、いくつか関連する知⾒が得られている。12歳 時点での⺟親の愛情や温かみが強いほど、12歳から16歳にかけての扁桃体の体積の増 加傾向が緩やかであることが⽰されている69)。また、扁桃体の活動を制御すると⾔わ れている腹内側前頭前⽪質の灰⽩質体積が⺟親の温かみと有意な相関関係を⽰すこ
とも報告されている70)。著者らの研究グループもまた、扁桃体と機能的結合性を持つ 後部島⽪質71)の灰⽩質体積が、⺟親が⼦どもを褒めようという意識を強く持っている ほど⼤きいことを明らかにしている72)。海⾺の体積と虐待・ネグレクトの負相関は成 ⼈期のみに⽣じる頑健な知⾒であると⾔われているが、規範的な養育と海⾺体積につ いては学童期・⻘年期から有意な相関関係が報告されている。しかし、4歳時点での ⺟親の養護性(⼦どもへの愛情や温かみを総称する概念)が⾼いほど14歳時点での海 ⾺体積が⼩さいという報告73)がある⼀⽅で、3歳から6歳にかけての⺟親の養育態度が ⽀援的であったほど10歳から12歳にかけての海⾺体積の増加傾向が⼤きいという報 告74)もあり、知⾒は⼀貫していない。このような知⾒の不⼀致の原因の⼀つとして、 養育が養護性や⽀援性といったポジティブな側⾯からしか評価されていないことが 指摘されている75)。22歳の⼤学⽣らに16歳以前の両親の養護性と過保護性を回顧的に 評価させた研究によると、過保護性が低い場合においてのみ、養護性が⾼いほど海⾺ 体積が⼤きいという有意な関係が確認された75)。 上記に挙げたいずれの先⾏研究も、規範的な養育による脳の機能・構造の個体差を ⾏動上の精神障害リスクに結びつけるには⾄っていない。しかしながら、虐待・ネグ レクトに該当しない範囲の養育の多様性と⼤脳⽪質−辺縁系回路の関係へのアプロ ーチは、過度な逆境を経験せずに育った群の精神障害リスクの個体差の理解に貢献す る可能性がある。 2.4. 脳機能・脳構造に対する遺伝・環境交互作⽤の影響 2.4.1. 画像遺伝学と遺伝・環境交互作⽤の統合
画像遺伝学で扱われる候補遺伝⼦のSNPの多くは浸透率が低く、精神障害の発症と
は必ずしも結びつかないと⾔われている76)
。また、ゲノムワイド関連解析(genome-wide association study; GWAS)によって同定される精神障害の感受性遺伝⼦とも⼀
致しない27)。⼀⽅、GWASによる感受性遺伝⼦の遺伝寄与率は双⽣児研究で算出され
た遺伝率よりも⼩さいことを意味する「失われた遺伝率」の問題も指摘されている76)。
さらに近年、脳構造への関与がGWASで同定された遺伝⼦多型が精神障害の感受性遺
伝⼦と⼀致しないことも明らかにされた77,78)。遺伝⼦と脳と精神障害リスクの関係を
取り巻くこれらの問題点への対処の⼀つとして、脳に対する遺伝要因と環境要因の交 互作⽤効果(遺伝環境交互作⽤; gene‒environment interaction; G×E)を調べる研究
(imaging G×E)が広がっている7,27)。 G×Eは、⾏動に対する環境要因の効果が遺伝要因によって調整される、または遺 伝要因の効果が環境要因によって調整されることを意味する79)。精神障害リスクに対 するG×Eの効果を検証した先駆的な研究は、ストレスフルなライフイベント(失業・ 離婚・親しい⼈との死別など)が多いほどうつ症状の程度が⼤きいという相関関係が、 セロトニントランスポーター遺伝⼦多型5-HTTLPRのShortアレルを持つ群でより強 く⾒られることを明らかにした80)。このような遺伝要因・環境要因・精神障害リスク の関係の調整モデルと、画像遺伝学が仮定する遺伝要因・脳・精神障害リスクの関係 の媒介モデルを統合することで、精神障害リスクの個体差についてより深い⽰唆が得 られると考えられた27,81)。そこで、遺伝要因から脳、脳から精神障害リスク、そして 遺伝要因から脳を介して精神障害リスクに伸びるそれぞれのパスが環境要因によっ
て調整されるという調整媒介モデルが提案された27)。imaging G×Eは画像遺伝学と同 様に、個体の精神障害リスクが形成されるメカニズムを分⼦から神経系までの多様な レベルで推定することができる点で有意義な研究⼿法であると⾔われている7,27,30)。た だし現在までに発表されている多くの研究では、脳の機能や構造と⾏動上の精神障害 リスクが統計的に結びついておらず30,41)、脳に対する遺伝⼦多型と環境要因の交互作 ⽤を検討する段階に留まっている。 2.4.2. FK506結合タンパク質51の遺伝⼦多型と虐待・ネグレクトの交互作⽤による脳 機能・脳構造の個体差 ⼤脳⽪質−辺縁系回路や⼤脳⽪質−⼤脳基底核−視床−⼤脳⽪質回路の機能・構造 に対するG×Eの効果を調べる研究において最も多く検討されているのが、HPA系機 能に関与するタンパク質の遺伝⼦多型と虐待やネグレクトの被害経験の交互作⽤で ある7)。特に、FK506結合タンパク質51(FK506-binding protein 51; FKBP51)をコード するFKBP5遺伝⼦のSNP rs1360780と虐待・ネグレクトの交互作⽤が⼤脳⽪質−辺縁 系回路や⼤脳⽪質−⼤脳基底核−視床−⼤脳⽪質回路に含まれる領域の機能や構造 に関与することは、頑健性が⾼いと⾔われている7)。 2.4.2.1. FKBP51の概要 FKBPは免疫抑制剤FK506に結合するタンパク質である82)。FK506は藤沢薬品⼯業 が筑波⼭に⽣育する放線菌から単離・同定した物質であり(FKはFujisawa Kaihatsuの 略、506は研究番号)、細胞障害性T細胞の産⽣抑制などの機能を持つことから、臓器 移植後の拒絶反応や⾃⼰免疫疾患の治療に利⽤されてきた83)。これらの機能はT細胞
の表層から核に⾄るシグナル伝達経路を阻害することで発揮されると⾔われている 84)。この作⽤機構が解明される過程で、FK506⾃体には薬理作⽤が無く、FKBPとの結 合によって活性が発現されることが明らかとなった。FKBPはペプチジルプロリルシ ストランスイソメラーゼドメインを持っており、プロリンのイミド結合のシスとトラ ンスを変換し、タンパク質の主鎖の向きを変えるタンパク質構造変換酵素である85-87)。 FKBPは⽣存には必須でないと⾒られているが88)、原核⽣物から多細胞⽣物まで幅広 く進化的に保存されていることから、⽣物機能にとって基本的な因⼦であると考えら れている89)。 FK506に結合して免疫抑制作⽤を発動するFKBP12が同定されて以降、FKBPファ ミリーの多様な構成が明らかにされるとともに、FK506が作⽤しない場合のFKBP本 来の機能とその標的タンパク質が解明されてきた90)。FKBPファミリーの⼀つである FKBP51は、FKBP5(6p21.31)にコードされる51キロダルトンのタンパク質である 82)。体内に広く分布するが、神経系ではGRとほぼ同様の領域に発現すると⾒られてい る91)。マウスの脳では、平常状態の場合、⼤脳新⽪質の第2層と第5層、中隔、脳梁 灰⽩層、梨状⽪質、海⾺、被殻、尾状核、腹側線条体、扁桃体、視床、視床下部、上 丘、腹側被蓋核、橋でのメッセンジャーRNA(mRNA)の発現が確認されている91)。 また、ストレス暴露後は海⾺のCA1領域と⻭状回、扁桃体、視床下部でmRNA量が増 加することが報告されている91)。 FKBP51はC末端のテトラトリコペプチド反復ドメインを介して熱ショックタンパ
Hsp90はGRのシャペロンであり、GRの構造を安定化させコルチゾールへの結合を可 能にするが、FKBP51がHsp90を介してGRに結合すると、GRのコルチゾール親和性 が低下する93)。GRとコルチゾールの結合はHPA系機能のネガティブフィードバック 抑制を⾏う重要なトリガーであるため、FKBP51によるGRのコルチゾール親和性低下 はネガティブフィードバック抑制の不全とコルチゾール量の上昇を引き起こす94)。他 ⽅で、GRとコルチゾールの結合により、FKBP51の転写が促進されることも明らかに されている95)。GRがコルチゾールと結合すると、FKBP51は複合体から分離し、代わ ってFKBP52が複合体に結合する82)。FKBP52はFKBP51と類似した構造を持つタンパ ク質であるが、GR−コルチゾール複合体を微⼩管モータータンパク質であるダイニ ンに渡し、核内移⾏を促進する働きを担う82)。核内に移⾏したGR−コルチゾール複合 体がFKBP5の第2イントロン内のGR応答配列に結合すると、FKBP51の転写が活性 化される96)。FKBP51の転写と翻訳が増加すると、GRの感受性が低下する。この過程 は細胞内のウルトラショートネガティブフィードバックと呼ばれており、末梢神経系 でその存在が確認されているが、中枢神経系での作⽤は未だ明らかにされていない94)。 しかし、仮にウルトラショートネガティブフィードバックが中枢神経系では⽣じなか ったとしても、FKBP51とGRの結合によるコルチゾール量の上昇による脳への影響は 精神障害リスクを充分に⾼め得ると考えられている94)。 2.4.2.2. FKBP5 rs1360780 rs1360780は、FKBP5の第2イントロンのGR応答配列から500塩基対分離れた箇所 に位置するSNPであり、マイナーアレル(T)頻度は30%程度であると⾔われている
94)。rs1360780がFKBP51の発現に関与する(図1)と考えられており、Tアレルはヒ トのリンパ球におけるFKBP51の発現量がCアレルよりも有意に多いことが確認され ている97)。⽇本⼈を対象とした研究でも、50歳以下の層においてTアレルキャリアは 末梢⾎単球中のFKBP51の発現量が多いと報告されている98)。TアレルではFKBP5の 第2イントロンの塩基配列内にTATAボックスが⽣じるために、TATAボックス結合タ ンパク質が結合しやすくなり、それによってGR応答配列と転写開始点の距離が近接 することで、FKBP51の産⽣が促進されると⾔われている99)。さらに、末梢⾎単球の FKBP5のmRNA量が多いほどコルチゾール濃度が⾼いことが確認されていることか ら97)、rs1360780のTアレルではFKBP51が過剰に発現しており、HPA系のネガティブ フィードバック不全とコルチゾール濃度の上昇が⽣じやすいと考えられている94)。 2.4.2.3. FKBP5 rs1360780 と虐待・ネグレクトの交互作⽤ 多くの先⾏研究により、FKBP5 rs1360780と⼩児期のトラウマティックな経験の 交互作⽤が精神障害リスクに影響する可能性が⽰されている。虐待やネグレクトを受 けたTアレルキャリアは、PTSD100)やMDD101)を発症しやすく、⾃殺企図102)に⾄る率 が⾼いと報告されている。不適切な養育という環境下では、Tアレルに特異的なコル チゾール反応が亢進してDNAの3次元構造が変化することで、FKBP5の第7イントロ ンにおけるGREの脱メチル化が⽣じて転写開始点と近接し、FKBP51の産⽣がさらに 促進される99)。このエピジェネティックな変化によるHPA系機能の不全が、虐待やネ グレクトを受けたTアレルキャリアにおける精神障害リスクの⾼まりを説明するメカ ニズムであると考えられている7,99)。
脳の機能や構造に対するrs1360780と虐待・ネグレクトの有意な交互作⽤効果もま た、先⾏研究によって報告されている。特に、虐待やネグレクトを受けていたTアレ ルキャリアにおいて脅威刺激に対する扁桃体の賦活が⼤きいことは複数の研究グル ープ間で確認されている103,104)。また、Tアレルキャリアでのみ、虐待経験の程度が⼤ きいほど島⽪質と弁蓋部の⽩質のFAが低く、平均拡散率が⾼いことも⽰されている 105)。より⼤規模なサンプル数による研究では、虐待を受けていたTアレルのホモ接合 型(T/T型)において、海⾺、海⾺傍回、前帯状⽪質、扁桃体、島⽪質、被殻、尾状 核などの灰⽩質体積が虐待を受けていたC/Tヘテロ接合型(C/T型)・Cアレルホモ接 合型(C/C型)よりも有意に⼩さいことが明らかにされている106)。なお、海⾺の体積 についてはrs1360780の主効果も多く検討されてきたが、遺伝⼦型の間で有意な差は 得られていない。FKBP5の第7イントロンにおけるDNAメチル化レベルが低いほど海 ⾺の体積が⼩さいという報告があることから、海⾺の構造への影響は虐待やネグレク トという逆境が経験された場合のみに限られる可能性が⽰唆されている99)。以上より、 ⼤脳⽪質−辺縁系回路や⼤脳⽪質−⼤脳基底核−視床−⼤脳⽪質回路に含まれる領 域に対するFKBP5 rs1360780と虐待・ネグレクトの交互作⽤効果は、imaging G×E の分野において特に頑健性が⾼いと認知されている7)。 2.5. 問題の所在 以上より、HPA系機能に関与するタンパク質の遺伝⼦多型、虐待やネグレクト、そ してそれらの交互作⽤が脳機能・脳構造に影響することを解明した数々の先⾏研究は、 精神障害リスクの個体差の基盤の理解に貢献してきたと⾔える。しかし、imaging G
×Eを⽤いた研究領域の問題点の⼀つとして、虐待とネグレクトのみに環境要因を絞 ってきたことが挙げられる7)。元来、脳の発達と養育の関係を調べる研究は虐待・ネ グレクトの効果の検討から始まった。しかし、虐待やネグレクトは極端に不適切で特 異的な養育形態であり、虐待・ネグレクト被害者を対象とした研究から得られた知⾒ がそれらを経験していない群にも拡⼤できるとは⾔えないという指摘があった48)。以 降、規範的な養育と脳発達の関連にも関⼼が注がれ始めたのは2.3.2.3で述べた通りで ある。この指摘の観点はimaging G×Eを⽤いた研究にも適⽤されるべきであると考 えられる。虐待・ネグレクトの被害経験を環境要因とした研究のみでは、それらを経 験していない群における精神障害リスクの個体差の基盤を理解することができない。 例えば、⾃殺者の死後脳を解析した先⾏研究によると、虐待を受けていた⾃殺者群は 対照群である事故死群よりもGRプロモーターのメチル化レベルが⾼くGRのメッセ ンジャーRNA量が少なかったが、虐待を受けていなかった⾃殺者群と事故死群の間に この差は検出されなかった107)。この結果から推察されるように、虐待を受けていた群 と受けていなかった群では、精神障害リスクの個体差を形成するメカニズムが異なる 可能性がある。しかし著者の知る限り、虐待を受けていなかった群における精神障害 リスクや脳機能・脳構造への遺伝⼦多型と養育の交互作⽤効果を検討した研究は、こ れまでに⾏われていない。
3. 研究⽬的
以上の研究背景を受け、本研究の⽬的は、脳構造に対するHPA系関連遺伝⼦多型と 規範的な養育の交互作⽤効果の検討により、虐待を受けずに育った群における精神障 害リスクの個体差の神経基盤について⽰唆を得ることとした。 ここで着⽬する遺伝⼦多型として、FKBP5 rs13607807,99)を選択した。このSNPが タンパク質とHPA系に与える作⽤が明確であること94)、このSNPと虐待やネグレクト の交互作⽤効果について脳画像研究の知⾒が蓄積されている103-106)と共にエピジェネ ティックな機構も解明されている99)ことが、rs1360780を選択した理由である。また、 近年の先⾏研究により、Tアレルキャリアはネガティブなライフイベントが多いほど 神経症傾向が⾼いが、⼀⽅でポジティブなライフイベントが多いほど神経症傾向が低 くなることが明らかにされている108)。したがって、rs1360780は虐待やネグレクトの ような極端な要因のみならず、環境要因のより広範なバリエーションとの交互作⽤に よって脳発達および精神障害リスクに影響する可能性が考えられる。そこで本研究で は、虐待やネグレクトとの隔たりを仮定しやすいポジティブな養育の指標を規範的な 養育の因⼦として選択した。 脳に対するrs1360780と虐待・ネグレクトの交互作⽤効果を⽰した先⾏研究の知⾒ 103-106)から、Tアレルキャリアにおいて、⼤脳⽪質−辺縁系回路、⼤脳⽪質−⼤脳基底 核−視床−⼤脳⽪質回路に含まれる領域の構造とポジティブな養育の指標の間に、有 意な相関関係が⾒られると予想した。4. 研究⽅法
4.1. 研究対象 本研究の対象者は5.7歳から18.4歳の健常な⽇本⼈202名(男児101名・⼥児101名) であった。本研究は、東北⼤学加齢医学研究所認知機能発達寄付研究部⾨が2008年か らおよそ4年間に渡って実施した、⼩児脳発達研究プロジェクト109)によって得られた データセットを利⽤して⾏った。同プロジェクトはヘルシンキ宣⾔に基づいて⾏われ、 東北⼤学⼤学院医学系研究科倫理委員会の承認を得た上で実施された(承認課題名 「MRIを⽤いた⼩児の脳形態、脳機能、認知機能に関する研究」、承認番号:2008-129, 2008-371)。 同プロジェクトは脳および認知機能の定型的な発達過程の解明を⽬的としたため、 発達障害や各種疾患の既往歴のない健常な⼩児を研究参加者として募集した。まず、 責任者の同意を得られた宮城県内の幼稚園・⼩学校・中学校・⾼等学校に、研究の概 要を記した募集広告を配布した。この広告を通じて研究に関⼼を持った保護者1,423 名に研究の詳細を記した資料を電⼦メールにて送付したところ、776組の保護者と⼩ 児から研究参加の意思が得られた。悪性腫瘍・5分以上の意識喪失を伴う頭部外傷・ 発達障害・癲癇・精神障害・閉所恐怖症の既往歴がないこと、⾊覚異常がないこと、 その他何らかの疾患の治療のための通院や薬剤服⽤を継続していないことを適⽤基 準とし、これらの基準を満たしていることを電話や電⼦メールによって保護者に確認 した。適⽤基準を満たした⼩児とその保護者に対して検査内容やデータの取り扱いにントおよびインフォームド・アセントが得られた。疲労や閉所恐怖症により8名が脳 画像撮像を中⽌したため、最終的に、290名(5.6歳から18.4歳, 男児145名, ⼥児145 名)が脳画像撮像と認知⼼理検査に参加した。また、保護者を対象とした養育に関す る質問紙検査も⾏われた。なお、すべての対象者が右利きであることをエジンバラ利 き⼿テスト110)によって確認した。 同プロジェクトは縦断調査であり、第⼀期調査の参加者290名のデータ収集が完了 したおよそ3年後に第⼆期調査が実施された。第⼆期調査では、脳画像撮像と認知⼼ 理検査に加え、遺伝⼦解析を⽬的とした唾液採取が⾏われた。本研究は脳画像、養育、 遺伝⼦のデータを必要としたため、第⼀期調査、第⼆期調査の両⽅に参加した234名 のデータのみを利⽤した。親⼦関係の評価と脳画像撮像の時期を統⼀するため、第⼀ 期調査で撮像した脳画像を解析に⽤いた。234名のうち、データ⽋損のあった32名(脳 画像データ⽋損者11名(詳細は4.4.を参照)、親⼦関係データ⽋損者15名(詳細は4.2. を参照)、遺伝⼦データ⽋損者6名(詳細は4.3.を参照))を解析から除外した結果、最 終的に前述の202名が本研究の対象となった。 脳に対するFKBP5 rs1360780と被虐待経験の交互作⽤効果を検討した先⾏研究 103-106)の研究対象者は、多くがヨーロッパ系の成⼈である。しかし成⼈を対象とした場合、 養育は回顧的に評価せざるを得ないため、⼩児期や⻘年期に実際に経験していた養育 とは乖離が⽣じる可能性がある。また、年齢が上がるにつれ、脳の発達には遺伝要因 と養育以外の様々な要因(⽣活習慣やストレスなど)が影響するため、本研究が⽬的 とするFKBP5 rs1360780とポジティブな養育の交互作⽤効果が検出しにくくなる可
能性が考えられる。以上の理由から、本研究は⼩児および⻘年を対象とした。本研究 の対象者が⽇本⼈であることは、保護者への問診によって確認された。著者の知る限 り、⽇本⼈を対象として脳構造に対するFKBP5 rs1360780と虐待・ネグレクトの交互 作⽤効果を検討した研究はこれまでに存在しない。しかし研究背景にて前述の通り、 FKBP5 rs1360780と虐待・ネグレクトの関係のエピジェネティックな機構は解明さ れており99)、この機構は対象者の遺伝的な系統に依らず共通のものであると推察され る。したがって、本研究の対象者が⽇本⼈であることは解析結果およびその解釈に重 ⼤な影響を与えるものではないと考えられる。 4.2. ポジティブな養育の評価 ポジティブな養育を評価する指標として、FDT親⼦関係診断検査(親⽤)111)の下位 尺度である「基本的受容」に含まれる質問項⽬への対象者の⺟親の回答を使⽤した。 FDT親⼦関係診断検査(親⽤)は、親が⼦供を親とは違う⼀⼈の独⾃性を持つ⼈間と して⾒ているか、⼦どもの個性を好んでいるか、といった親⼦関係の⼼理的側⾯を親 の視点から捉えることを主眼とした質問紙検査である。東ら(2002)によって開発され、 594名の⺟親と524名の⽗親を対象とした調査により、因⼦的妥当性および信頼性が 確認されている。下位尺度は上述の基本的受容の他、無関⼼・養育不安・夫婦間不⼀ 致・厳しいしつけ・達成要求・不介⼊の6つで構成される。回答⽅法は、質問項⽬の内 容に対し「1. まったくあてはまらない、2. あまりあてはまらない、3. どちらともい えない、4. だいたいあてはまる、5. よくあてはまる」のいずれか⼀つを選択する形 式である。
FDT親⼦関係診断検査(親⽤)の原版は全40項⽬から成るが、⼩児脳発達研究プロ ジェクトでは2008年当時の担当者の判断により、全20項⽬に短縮して使⽤された。基 本的受容尺度に含まれる項⽬は原版の10項⽬から5項⽬へと短縮された。同プロジェ クトではこの他にも多くの質問紙検査への回答を保護者に依頼していたため、⼀つの 質問紙への回答にかける保護者の負担軽減が短縮の理由であった。原版の10項⽬の内 容は、「⼦どものよいところを、ほめるようにしている」「この⼦は、よい⼦だと思う」 「この⼦は、私の気持ちをよく分かってくれる」「この⼦のことを、誇りに思う」「こ の⼦と話していると楽しい」「⼦どもの性格が、あまり好きではない(逆転項⽬)」「こ の⼦の⽋点ばかりが⽬につく(逆転項⽬)」「この⼦が、もう少し違う⼦であったらよ いのにと思う(逆転項⽬)」「⾃分と⼦どもは、相性があまり良くない(逆転項⽬)」 「⼦どもが何を考えているのか、ほとんど分からない(逆転項⽬)」であった。短縮に 際して選択された項⽬は、「⼦どものよいところを、ほめるようにしている」「この⼦ のことを、誇りに思う」「この⼦と話していると楽しい」「この⼦の⽋点ばかりが⽬に つく(逆転項⽬)」「⼦どもが何を考えているのか、ほとんど分からない(逆転項⽬)」 であった。 原版の意図通りに親⼦関係を正しく評価できない可能性があるという点で、この短 縮は本研究の重⼤な問題点である。しかしながら、本研究の対象者の⺟親202名の回 答データから算出した基本的受容尺度5項⽬のクロンバックのα係数は0.72であった。 これは原版10項⽬のクロンバックのα係数0.82に若⼲劣るものの、おおよそ同程度の 信頼性が担保できていると考えられる。また、この5項⽬の内容的妥当性を確認する
ための指標として、同プロジェクトで収集された親⼦関係の良好性に関する質問項⽬ の回答を利⽤した。親⼦関係の良好性に関する質問は、第⼀期調査当時⼩学4年⽣以 下の参加者の場合は保護者が回答し、⼩学5年⽣以上の参加者の場合は本⼈が回答し た。保護者⽤の項⽬は「あなたは、お⼦様との親⼦関係について以下のどれにあては まると思いますか」であり、本⼈⽤の項⽬は「あなたは、親との関係について以下の どれにあてはまると思いますか」であった。いずれの場合も、「1. うまくいっている、 2. まあうまくいっている、3. あまりうまくいっていない、4. うまくいっていない、 5. 分からない」から⼀つ回答を選択する形式であった。「5. 分からない」を選択した 参加者を除外し、⼩学4年⽣以下91名、⼩学5年⽣以上106名を対象として、親⼦関係 の良好性と基本的受容尺度5項⽬の回答の合計値の2変量相関解析を⾏ったところ、 いずれの群においても有意な相関関係が確認された(⼩学4年⽣以下:r = −0.49, p < 0.001; ⼩学5年⽣以上:r = −3.12, p = 0.001)。親⼦関係の良好性は回答の数値が 低いほど関係が良好であることを表すため、いずれの群も親⼦関係が良好であるほど ⺟親が⼦どもを受容していることが明らかとなった。⼩学5年⽣以上の相関係数r = −3.12は弱い相関を⽰しているが、基本的受容尺度の回答者が⺟親であるのに対して 親⼦関係の良好性の回答者が⼩児であったことと、親⼦関係の良好性についての項⽬ の評価対象が⺟親との関係に限定されていなかったことが、相関の強さを低めた要因 であると考えられる。以上の限界点を考慮する必要があるものの、本研究で使⽤した 基本的受容尺度5項⽬は、⺟親と⼦どもの間のポジティブな関係を評価する項⽬とし て妥当であると考えられる。したがって、本研究では基本的受容尺度5項⽬の回答の
合計値を「⺟親の受容性」の得点と⾒なし、規範的な養育の指標として使⽤した。な お、逆転項⽬は回答された値を6から引いた差分を得点とし、5項⽬の合計値が⼤き いほど⺟親の受容性が⾼いことを意味するように処理した。
4.3. 遺伝⼦解析
各対象者のFKBP5 rs1360780の遺伝⼦型(C/C, C/T, T/Tのいずれか)を特定する
ために、ポリメラーゼ連鎖反応(polymerase chain reaction; PCR)によって増幅する DNAをリアルタイムでモニタリングする⽅法である、リアルタイムPCRを⽤いた SNPタイピングを⾏った。 PCR法は細胞分裂時のDNA複製の仕組みを利⽤して開発された技術であり、DNA の特定の領域を数時間で100万倍に増幅させることができる。DNA複製の過程112)では、 まずDNAヘリカーゼによってDNAの⼆重らせん構造が解体され、2本の鋳型DNAと なる。DNAポリメラーゼと呼ばれるDNA合成酵素が、鋳型DNAの塩基配列と正しい 塩基対を形成できるデオキシリボヌクレオチド3‒リン酸を合成中のDNA鎖の3ʼ末端 に付加していくことにより、互いに相補的な⼆本鎖DNAが完成する。PCR法113)では、 以上のDNA複製の過程が熱変性によって再現される。標的とする遺伝⼦を含んだ⼆ 本鎖DNAと、その両端部分のそれぞれに相補的な⼆種類のDNA断⽚(プライマー)、 そしてDNAポリメラーゼを含んだ反応溶液の温度を、専⽤の機械で周期的に変動さ せる。この処理により、⼆本鎖DNAの解離、解離した⼀本鎖DNAとプライマー分⼦ の相補的な会合、プライマー断⽚からのDNA鎖の伸⻑、という三つの反応が繰り返さ れる。
リアルタイムPCRを⽤いたSNPタイピングにおいて最も簡便で広く使われている ⼿法が、TaqMan®アッセイによる⽅法である114)。この⽅法では、⽬的の遺伝⼦を含ん だ DNA と 、 増 幅 し た い DNA 領 域 に 応 じ て 合 成 さ れ た プ ラ イ マ ー に 加 え 、 TaqMan®MGBプローブという蛍光標識プローブ(VIC®プローブおよびFAMTMプロー ブ)が⽤いられる。VIC®プローブは5ʼ末端がVIC⾊素で標識され、⼀⽅のアレルに特 異的にハイブリダイズするように設計されている。また、FAMTMプローブは5ʼ末端が FAM⾊素で標識され、もう⼀⽅のアレルに特異的にハイブリダイズする設計となって いる。PCRによってDNAが合成されると、例えば鋳型DNAにC(シトシン)が存在す る場合、相補的な配列を持つVIC®プローブがハイブリダイズし、VIC⾊素の蛍光が発 ⽣する。FAMTMプローブはSNPサイトの配列が異なるためこの鋳型DNAにハイブリ ダイズできないが、T(チミン)が存在する鋳型DNAにハイブリダイズし、FAM⾊素 の蛍光を発する。理論上、Cアレルのホモ接合の場合はVIC蛍光が2単位、Cアレルと Tアレルのヘテロ接合の場合はVIC蛍光とFAM蛍光が1単位ずつ、Tアレルのホモ接合 の場合はFAM蛍光が2単位発⽣するため、各サンプルのVICとFAMの蛍光強度を専⽤ のソフトウェア上でグラフにプロットすることにより、各参加者の遺伝⼦型が特定さ れる113)。 ⼩児脳発達研究プロジェクトでは、第⼆期調査に参加した234名の唾液が採取され、 東北メディカル・メガバンク機構災害精神医学分野(富⽥研究室)の技術補佐員がこ れらの唾液からDNAの抽出を⾏った。唾液採取およびDNA抽出には、DNA採取キッ トであるOragene(DNA Genotek, Inc., Ottawa, ON, Canada)が⽤いられた。SNPタ
イピングは災害精神医学分野の実験室にて著者が⾏った。20ngのゲノムDNAを含む
抽出液2.5-μL、FKBP5 rs1360780のタイピング⽤に設計されたプライマーである
TaqMan®アッセイ(ID: C_8852038_10; Applied Biosystems, Foster City, CA, USA) 0.25-μL、蛍光プローブを含んだ反応液であるTaqMan®マスターミックス(Applied Biosystems)5-μL、ヌクレアーゼフリー⽔2.25-μLを混合し、CFX ConnectTMリアル タイムPCR検出システムによってPCRにかけた。PCRによるDNAの増幅が適切に進 ⾏したかを確認するためのコントロールとして、ヌクレアーゼフリー⽔10-μLを⽤い た。TaqMan®Genotyper Softwareを⽤いて蛍光強度をプロットし、参加者の遺伝⼦型 を特定した。但し、試料の質の問題により6名の遺伝⼦型は特定できなかったため、 これらの参加者は統計解析の対象から除外した。SNPタイピングを同じ⼿順で⼆度実 施することによってタイピング結果の妥当性を確認した。SNPタイピングの結果、本 研究の対象者202名のうち、130名がC/C型、66名がC/T型、6名がT/T型であると特 定された(SNPタイピングの結果の詳細は5.1で後述)。T/T型が少数であることから、 本研究ではC/C型をC/C群、C/T型とT/T型をまとめてTアレルキャリア群とした。 4.4. 脳画像撮像 東北⼤学加齢医学研究所に設置されている磁場強度3Tの磁気共鳴画像(magnetic resonance imaging; MRI)装置(Phillips Medical Systems, Best, The Netherlands)を ⽤いて参加者の脳画像が撮像された。Magnetization-prepared-rapid gradient echo法 によって撮像された3次元T1強調画像を本研究の解析に使⽤した。3次元T1強調画像 の撮像に使⽤されたパラメーターは、撮像画⾯マトリクス240 × 240、繰り返し時間
6.5 ms、エコー時間3 ms、反転時間711 ms、撮像野24 cm、スライス数162枚、スライ ス厚1.0-mmであり、撮像時間は483秒であった。撮像中の参加者の体動が激しかった 場合は撮り直しが⾏われた。画像の質は東北⼤学加齢医学研究所機能画像医学研究分 野の放射線科医が確認し、アーチファクトが⾒られた11名の画像は解析対象から除外 された。 4.5. 脳画像前処理 個々⼈の脳形態はそれぞれ異なるため、撮像されたままの3次元T1強調画像は統計 解析に適さず、灰⽩質や⽩質の体積に対する独⽴変数の効果を正しく検出することが できない。そのため、統計解析に先⽴つ画像処理(前処理)として、灰⽩質や⽩質と いった脳組織の分離抽出処理(セグメンテーション)と、標準空間への合わせこみに よる脳形態の変形処理(解剖学的標準化)が必要となる。本研究では、Voxel based morphometry(VBM)による前処理を⾏った115)。VBMは、特定の脳領域ではなく全 脳を統計解析の対象とすることで、灰⽩質・⽩質の密度や体積をボクセルごとに探索 的かつ客観的に評価する⼿法である。VBMのセグメンテーションでは、MR信号の閾 値を設定して脳組織を抽出する⽅法と、既存の組織存在確率マップを⽤いて脳組織ら しさの重み付けをする⽅法を組み合わせることによって、灰⽩質や⽩質の組織画像が ⽣成される116)。続く解剖学的標準化では、セグメンテーションによって得られた組織 画像がテンプレートと呼ばれる基準画像に合わせ込まれる。この処理により、個々⼈ の脳の形態差が最⼩限化されるため、ボクセル密度によって表される脳体積の⼤⼩を 個⼈間で⽐較できるようになる。