TUMSAT-OACIS Repository - Tokyo University of Marine Science and Technology (東京海洋大学)
低温ならびに中温性ヒスタミン生成菌の挙動と迅速
同定法に関する研究
著者
通堂 裕子
学位名
博士(海洋科学)
学位授与機関
東京海洋大学
学位授与年度
2013
学位授与番号
12614博甲第319号
URL
http://id.nii.ac.jp/1342/00000987/
博士学位論文
低温ならびに中温性ヒスタミン生成菌の挙動と
迅速同定法に関する研究
平成 25 年度
(2014 年 3 月)
東京海洋大学大学院
海洋科学技術研究科
応用生命科学専攻
通堂 裕子
目次
序論 ・・・・・・2 第 1 章 鮮魚における低温性および中温性ヒスタミン生成菌の 分布 ・・・・・・8 1.1 緒言 ・・・・・・8 1.2 実験方法 ・・・・・・9 1.3 結果 ・・・・・・11 1.4 考察 ・・・・・・12 図表 ・・・・・・16 第 2 章 低温性ヒスタミン生成菌 Photobacterium phosphoreum および P. iliopiscarium の低温保存下における魚肉中での挙動 ・・・・・・22 2.1 緒言 ・・・・・・22 2.2 実験方法 ・・・・・・22 2.3 結果 ・・・・・・24 2.4 考察 ・・・・・・25 図表 ・・・・・・27 第 3 章 高感度融解曲線解析(HRMA)を用いた ヒスタミン生成菌の迅速同定法の検討 ・・・・・・31 3.1 緒言 ・・・・・・31 3.2 実験方法 ・・・・・・32 3.3 結果 ・・・・・・35 3.4 考察 ・・・・・・35 図表 ・・・・・・38 総括 ・・・・・・48 参考文献 ・・・・・・50序論
ヒスタミン(Hm)食中毒、またはアレルギー様食中毒と呼ばれる食中毒は、宮 木らによって Hm が原因となって発生することが報告され(Miyaki, 1954)、主に発 疹・蕁麻疹・顔面の紅潮・吐き気および頭痛など、アレルギーに類似した症状を示 す食中毒である(Taylor, 1985)。その症状は主に摂食後、10 分から 1 時間程度で発 症し、症状の程度は患者の感受性によって異なるとされている。症状は通常 24 時 間程度で治まるが、完治に数日要する場合もある(Hungerford, 2010)。Hm は、あ る種の微生物が有するヒスチジン脱炭酸酵素(histidine decarboxylase; HDC)によっ て、食品中のアミノ酸であるヒスチジンが脱炭酸されることで生成される(Fig. 1) (Kimata and Kawai, 1953; Lerke et al., 1978)。Hm 食中毒は、主に水産食品を原因と して発生し、中でも一般的な赤身魚、特にサバ科に属する赤身魚は遊離ヒスチジン 含量が 700~1,800 mg/100 g と非常に高いため(Auerswald et al., 2006)、原因食品と なりやすい(Lehane and Olley, 2000)。また、赤身魚以外でも、シイラやサバヒーと いった一部の白身魚において発生事例が報告されている(Chen et al., 2011; Tsai etal., 2005)。
Hm 食中毒は、世界中で発生が報告されており、日本においては 1950 年代初頭 には主な食中毒として多数報告され、1970-1980 年の 10 年間には 4122 件の報告が みられた(Lehane and Olley, 2000)。日本においては、干物や調理品における発生が 多く、アメリカなど諸外国においては缶詰などにおける発生が多い(戸田ら, 2009)。 低温貯蔵技術が普及するとともに衛生管理が徹底されるようになったことで、Hm 食中毒の発生件数は減少し、日本においては、現在ではその発生数は年数件から十 数件程度となっている(Fig. 2)(山中英明ら, 2012)。しかし、Hm 食中毒が学校給 食など大量調理施設で発生した際には、100 名以上の患者を出すような大きな被害 を起こす場合があり(戸田ら, 2009)、2009 年には学校給食のマグロごまフライを 原因として 279 名の、2013 年には保育園給食のイワシつみれ汁を原因として 109 名 の 患 者 を 出 し た 事 例 が あ る ( 厚 生 労 働 省 , 食 中 毒 事 件 一 覧 速 報 〔http://www.mhlw.go.jp/topics/syokuchu/04.html〕)。そのため、Hm 食中毒は現在に おいても食品衛生上重要な問題であると考えられている。Hm は加熱や凍結に対し て非常に安定であることから、一度食品中に蓄積した場合、加工・調理工程におけ る除去は困難である。そのため、Hm 食中毒を予防する上では Hm 蓄積の抑制が重 要である。 HDC を有する Hm 生成菌として、これまでに多くの菌種が報告されている。グ ラム陽性菌では、ピルボイル依存型の HDC を有する(Recsei and Snell, 1985)
Clostridium perfringens(Yoshinaga and Frank, 1982)、Lactobacillus buchneri(Martín et
al., 2005)、Tetragenococcus muriaticus(Kimura et al., 2001)、Streptococcus thermophilus
(Calles-Enríquez et al., 2010)および Oenococcus oeni(Coton et al., 1998)等の複数 の菌種が Hm 生成菌として報告されており、菌種は主にワイン、チーズおよび魚醤 などの発酵食品からの分離が報告されている。グラム陰性菌では、Enterobacter
aerogenes(Enjalbert et al., 1979)、Raoultella planticola(Kanki et al., 2002)、Morganella morgani(川端ら, 1956)を始めとした腸内細菌科菌群や Photobacterium phosphoreum
(Fujii et al., 1997)や Photobacterium damselae subsp. damselae(Kimura et al., 2000; Takahashi et al., 2008)を始めとした海洋性細菌など多様な菌種が Hm 生成菌として 報告されている。これらの菌種は、主に鮮魚や、発酵食品を除く水産食品からの分 離が報告され、ピリドキサール型の HDC を有している(Kamath et al., 1991)。 鮮魚および発酵食品以外の水産食品における Hm 食中毒の主な原因菌としては、 グラム陰性で 30-37ºC が至適温度である中温増殖性を有する M. morganii、E.
aerogenes、R. planticola および R. ornitinolytica 等が報告されてきた(Behling and
Taylor, 1982)。そのため、低温保存・流通下において厳密な温度管理を行う事が Hm
食中毒の予防を行う上で重要だと考えられてきた。しかし、低温貯蔵技術が普及し、 温度管理が徹底的に行われるようになった現在でも、Hm 食中毒は散発的であるも のの発生し続けている(Fig. 2)。
低温性 Hm 生成菌に関する研究は、Okuzumi ら以降複数報告されている(Okuzumi et al., 1981; 与口ら, 1990b; Morii et al., 1995, 2006; Kanki et al., 2007)Okuzumi et al. (1981)は、低温性かつ好塩性 Hm 生成菌をマサバより分離し、N 菌群と命名した。 これら N 菌群の一部は後に P. phosphoreum として同定された(Fujii et al., 1997)。 Morii らは鮮魚より発光細菌として P. phosphoreum を分離し、低温下における魚肉 中での Hm 生成を報告した(Morii et al., 1988)。これまでに低温性 Hm 生成菌の Hm 生成能に環境因子が与える影響に関する研究(栗原ら, 1993a, 1993b; 森井ら, 1994; Morii and Kasama, 1995, 2004)や、HDC 遺伝子(hdc)に関する研究(Kanki et al., 2007; Morii et al., 2006)が報告されている。さらに、P. phosphoreum は、Hm 食 中毒の原因菌として分離された例も報告されている(Kanki et al., 2004)。同様に 低 温 下 で 高 い Hm 生 成 能 を 示 す Hm 食 中 毒 の 原 因 菌 と し て Morganella
psychrotolerans も報告されている(Emborg et al., 2006)。このように一部の低温性
Hm 生成菌に関して報告があるものの、低温性 Hm 生成菌が原因となって報告され た Hm 食中毒発生事例は少数であり、なおかつ食中毒発生事例においては原因菌が
前述のように Hm 生成菌は多様な菌種が報告されており、Hm 食中毒の原因究明 を行う際には原因菌の検出および正確な菌種同定が必要とされている。通常 Hm 生 成菌の同定には、Niven 培地など Hm 生成による培地の pH 上昇を利用した培養法 で検出の後、各種性状試験を利用したキット(api、VIDAS、VITEK など)や 16S ribosomal DNA(rDNA)配列の相同性検索を用いた同定が行われている。しかし、 これらの手法は長い時間を必要とすることから、早急な原因究明を必要とする場合 には、より迅速に行える同定法が必要である。今まで確認されているグラム陰性 Hm 生成菌は、同一のピリドキサールリン酸依存型 HDC を有しており、なおかつ これらの菌種の hdc 遺伝子は 16S rDNA 遺伝子と同等の解析能を有し、同定に利用 する事が可能である事が報告されている(Takahashi et al., 2003)。さらに迅速同定 法に関する研究も single-strand conformation polymorphism(SSCP)法(Takahashi et al., 2003, 2007)が報告されている。しかし、本手法は煩雑な手法を必要とする場合が あることから、さらに簡便な手法が求められている 本研究においては、Hm 食中毒の防止に寄与する事を目的とし、鮮魚における低 温性および中温性 Hm 生成菌の分布を調べると共に、冷蔵保存下における低温性 Hm 生成菌の挙動を調べた。また、hdc 遺伝子を用いた迅速同定法の検討を行った。 以下の検討を行った。第 1 章においては、アジ、サバ、マグロなどの赤身魚を中 心とした魚種を対象に、低温性および中温性 Hm 生成菌の分離・同定を行った。第 2 章においては、第 1 章において分離された菌株のうち、高い Hm 生成能の高い低 温性 Hm 生成菌 2 株を魚肉に接種し、4°C、10°C および 15°C で保存した場合の、 Hm 蓄積量の変化を検証した。第 3 章では、Hm 生成菌の迅速同定を目的に中温性 および低温性 Hm 生成菌の hdc 遺伝子を対象とした PCR-高感度融解曲線解析 (High-resolution melting analysis)の検討を行った。。
本論文の内容の一部は、以下の通り公表済みもしくは公表予定である。 【学術論文】
Yuko Torido, Hajime Takahashi, Takashi Kuda and Bon Kimura. 2012. Analysis of the growth of histamine-producing bacteria and histamine accumulation in fish during storage at low temperature. Food Control. 26, 174-177.
Yuko Torido, Hajime Takahashi, Satoko Miya, Takashi Kuda and Bon Kimura. Distribution of psychrophilic and mesophilic histamine-producing bacteria in retailed fish in Japan. Food Control. 審査中
○通堂裕子, 高橋肇,久田孝,木村凡. 鮮魚におけるヒスタミン生成菌の分布及 び低温下におけるヒスタミン生成菌の挙動について. 日本水産学会(2010 年 9 月、 京都) ○通堂裕子,郷田実穂,高橋肇,久田孝,木村凡.市販鮮魚におけるヒスタミン生 成菌の汚染状況調査と低温性ヒスタミン生成菌の遺伝子解析に関する研究. 日本 水産学会(2012 年 3 月、東京)
Fig. 1 Histidine decarboxylase reaction of Hm producing bacteria.
NH
N
NH
2COOH
NH
N
NH
2Histamine-producing bacteria
Histidine decarboxylase
CO
2
Fig. 2 Numbers of Hm food poisonings and patients in Japan from 2004 to 2013. Referred from database of food poisoning reported from Ministry of Health, Labour and Welfare (http://www.mhlw.go.jp/topics/syokuchu/04.html). 162 111 165 73 438 550 32 194 113 172 0 100 200 300 400 500 600 0 5 10 15 20 25 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 Num b er of p at ients Num b er of Hm f ood p oison in gs Year
第 1 章 鮮魚における低温性および中温性ヒスタミン生成菌の分布
1.1.緒言
Hm 食中毒の多くは水産食品で発生し、主に加工・流通下において温度管理の不 備が発生することによって引き起こされていると考えられている。そのため、Hm 食中毒の予防を行うためには、徹底した温度管理が必要不可欠とされている (Guizani et al., 2005; Kim et al., 2009; Kim et al., 2001, 2002; Tsai et al., 2005)。アメ リカ食品医薬品局(U. S. Food and Drug Administration; FDA)は迅速な冷凍もしくは 華氏 40°C(摂氏 4.4°C)以下での保存を推奨している(FDA, 2011)。近年では、徹 底した温度管理の実施と低温流通の発達によって、Hm 食中毒の発生数は減少した とされているが、現在でも世界中において散発的な発生が報告されている(Lehane and Olley, 2000; 戸田ら, 2009)。現在我が国においても、年数件から十数件の発生 事例が報告されている(Fig. 2)。 そのような近年の Hm 食中毒発生状況から、温度管理における不備の発生以外の 要因が存在している可能性が考えられた。さらに、過去の報告において冷蔵下の水 産食品を対象に Hm 量を調査した結果、31%のサンプルで平均 500 mg/kg 以上の Hm 蓄積が確認された例も存在する事から(Dalgaard et al., 2008)、低温性 Hm 生成菌 が原因となって発生している可能性が考えられた。このように、中温性 Hm 生成菌 のみならず低温性 Hm 生成菌が食品中において Hm 蓄積を引き起こしている可能性 がある上、近年の Hm 食中毒事例の中には、原因菌が明確となっていない事例 (Chang et al., 2008)が存在することから、Hm 蓄積の原因究明が困難な場合が存在 する。 現在まで Hm 食中毒、特に発酵食品を除く水産食品における事例において、主に グラム陰性の Hm 生成菌が原因菌として分離されている。グラム陰性の Hm 生成菌 のうち、中温性 Hm 生成菌としては Morganella morganii、Enterobacter aerogenes、
Raoultella planticola、 R. ornithinolytica(Kanki et al., 2002)、Proteus vulagaris(Kim et
al., 2001)など腸内細菌科菌群に属する菌種や、海洋性細菌である Photobacterium
damselae subsp. damselae や Vibrio parahaemolyticus、V. alginolyticus(Kim et al., 2001)
が分離されている。特に、M. morganii は広範囲の魚種の腐敗サンプルからの分離 例が多数報告されるとともに、高い Hm 生成能を示す(Kim et al., 2003)。そのため、
M. morganii は最も代表的な中温性 Hm 生成菌として考えられている。
一方低温性 Hm 生成菌としては、P. phosphoreum や M. psychrotolerans(Emborg et al., 2005, 2006)が分離されている。P. phosphoreum は低温下において高い Hm 生成
能を示すことが報告されており(Emborg et al., 2005)、0°C においても Hm 蓄積が 確認されたという報告例もある(Morii et al., 1988)。本菌は 15°C 以下では M.
morganii よりも高い Hm 生成能を示す場合があることも知られおり(Lehane and
Olley, 2000)、食中毒の原因菌としても報告されている(Kanki et al., 2004)。しかし
ながらこれまでに報告されている Hm 生成菌における鮮魚および水産食品からの 分離に関する研究の多くは、中温性 Hm 生成菌のみ対象としており(Allen et al., 2005; Kim et al., 2009; Koohdar et al., 2011; Kung et al., 2010; López-Sabater et al., 1994; 与口ら, 1990a)、低温性 Hm 生成菌を対象とした研究は少数だった(Okuzumi et al.,
1981; 与口ら, 1990b)。これらの研究において多様な魚種において同一サンプルか ら低温性および中温性 Hm 生成菌を同時に分離し、遺伝子的手法を用いた同定を行 った例はない。 そのため、本章では、広範囲の魚種を対象とし、低温性および中温性 Hm 生成菌 を培養法によって検出し、16S rDNA 配列を用いた菌種同定を行った。
1.2. 実験方法
1.2.1.食品サンプル 本実験で用いたサンプルは、2008 年 3 月から 2010 年 11 月に東京および埼玉の 小売店で購入した生鮮マグロ(Thunnus spp.)、アジ(Trachurus japonicus)、サバ (Scomber spp.)、カツオ(Katsuwonus pelamis)、サンマ(Cololabis saira)、イワシ (Sardinops melanostictus, Etrumeus teres)、イサキ(Parapristipoma trilineatum)、ブ リ(Seriola quinqueradiata)、メカジキ(Xiphiidae gladius)、トビウオ(Cypseluruspinnatibarbatus japonicus)およびニシン(Clupea pallasii)、計 143 サンプルを用い
た(Table 1)。
1.2.2. 鮮魚からの Hm 生成菌の分離
Hm 生成菌の分離方法は Takahashi et al.(2003)の方法に基づいて行った。すな わち各サンプルの表皮および周辺の魚肉を 10 g を無菌的に測り取り、ヒスチジン ブロス(Bacto Peptone [Becton, Dickinson, and Company(BD)、Franklin Lakes、New Jersey] 10 g、Bacto Yeast Extract [BD] 3 g、glucose [和光純薬工業、大阪] 5 g および L-histidine [和光] 5 g を 50%人工海水 [Artificial seawater: ASW] [NaCl 23.5 g、KCl
(中温性 Hm 生成菌分離用)で培養した。その後、ペーパークロマトグラフィーを 用いて Hm の検出を行った。ペーパークロマトグラフィーは、以下の方法で行った。
培養液5 μL を展開液(100% 1-butanol [国産化学、東京]:ammonium solution [国産化
学]=1:1 で混合)によって濾紙に展開した。室温で濾紙を乾燥させた後、反応液 A
(0.5% sulfanic acid [国産化学]+0.1% HCl: 1% NaNO2を 1:1 で混合)および反応液 B
(Na2CO3飽和溶液)を噴霧し、赤色を示したサンプルを Hm 陽性とした(Miyaki,
1954)。陽性サンプルの培養液を Niven 培地(Bacto Tryptone [BD] 5 g、Yeast extract
[BD] 5 g、L-histidine [和光] 27 g, CaCO3 [関東化学、東京] 1 g、0.4% bromocresol purple
[東京化成工業、東京] 15 mL および寒天 [国産化学] 20 g を 50% ASW 1 L に溶解後、 pH 5 に調整)(Niven et al., 1981)に画線し、30°C、24 時間および 15°C、72 時間 で培養した。周辺の培地が明るい紫色に変色したコロニーのうち、5~10 コロニー を釣菌し、ヒスチジンブロスに接種した。30°C、24 時間もしくは 15°C、48 時間培 養後、5 μL をペーパークロマトグラフィーに供し、Hm が検出されたものを Hm 陽 性菌とした。 1.2.3. 分離菌の Hm 生成能の確認 Hm 陽性菌を 30°C、24 時間および 15°C、48 時間で前培養後、終濃度 106 cfu/mL となるようにヒスチジンブロスに接種し、30°C 分離菌を 30°C、24 時間で、15°C 分離菌を 15°C、48 時間で培養後、0.2-μm フィルター(東洋濾紙、東京)でろ過滅 菌した。ろ液中の Hm 量をチェックカラーヒスタミン(キッコーマン、千葉)を用 いて測定し(Sato et al., 2005)、100 mg/L 以上の Hm 生成が確認されたものを Hm 生成菌とした。その際、同サンプルからの分離菌のうち、同菌種と同定され、なお かつ同等の Hm 生成能を有する菌が 2 つ以上ある場合、それらの菌を同一株とした。 本研究では、30°C で分離された菌株を中温性 Hm 生成菌、15°C で分離された菌株 を低温性 Hm 生成菌とした。 1.2.4. Hm 生成菌の同定 Hm 生成菌の同定は、Takahashi et al. (2003)の手法に沿って行った。すなわち 1.5%
NaCl 添加 Trypticase Soy broth(BD)に分離菌株を接種後、中温性 Hm 生成菌を 30°C、
24 時間、低温性 Hm 生成菌を 15°C、48 時間で培養し、培養液 1 mL を 15,000xg、5 分 、 4°C の 遠 心 で 集 菌 し た 。 得 ら れ た ペ レ ッ ト か ら の DNA 抽 出 は Mag Extractor-Genome(東洋紡、東京)を用いて、添付マニュアルに従って行った。DNA 抽出後、16S rDNA の増幅はプライマー27F(5’- AGAGTTTGATCCTGGCTCAG -3’) および 1492R(5’- GGTTACCTTGTTACGACTT -3’)を用いて行った(Weisburg et al.,
1991)。PCR 組成は Table 2 に示した。PCR 反応は、94ºC 5 分、35 cycle(95ºC 30 秒-58ºC 1 分-72ºC 1 分)および 72 ºC、 7 分とし、Veriti thermal cycler(Life Technologies Corp.、Foster City、California)を用いて行った。PCR 産物 5 μL を、1%アガロース ゲルを用いて 100-bp ladder(GE Healthcare Bio-Science Corp.、Piscataway、New Jersey) と共に 100 V、40 分で電気泳動に供した。電気泳動後はエチジウムブロマイドで染 色後、増幅を確認した。
PCR 増幅産物を Agencount® Ampure(Beckman Coulter Inc.、Brea、California)に て精製後、プライマー27F および 530R(5’- GTATTGCCGCGGCTGCTGGC -3’)を 用いてシーケンス PCR を行った。シーケンス PCR 反応は 96ºC 1 分、25 cycle(96ºC
10 秒-50ºC 5 秒-60ºC 4 分)として行った。増幅産物は Agencount® CleanSEQ(Beckman
Coulter Inc.)を用いて精製後、ABI PRISM® 310 Genetic Analyzer(Life Technologies Corp.)を用いてシーケンシングを行い、400~450 bp の配列決定を行った。得られ た配列は DNA data bank of Japan(DDBJ; 静岡)(http://www.ddbj.nig.ac.jp)の The BLAST 2.0 algorithm を用いて菌種決定を行った。16S rDNA 配列によって菌種の識 別が不可能な菌種に関しては、api 20E, api 50CHE および VITEK(BioMérieux Co. Ltd.、Craponne、Lyon)を用いて菌種を決定した。
1.3.結果
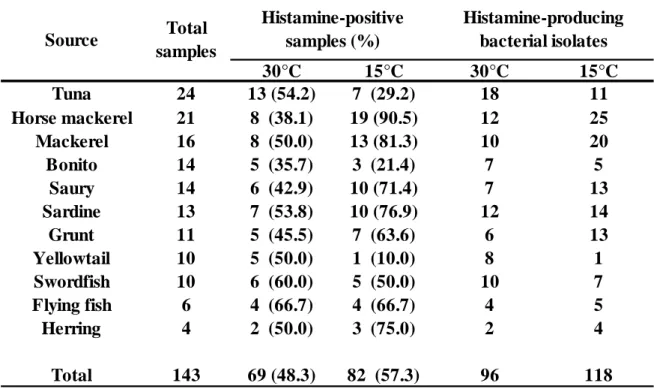
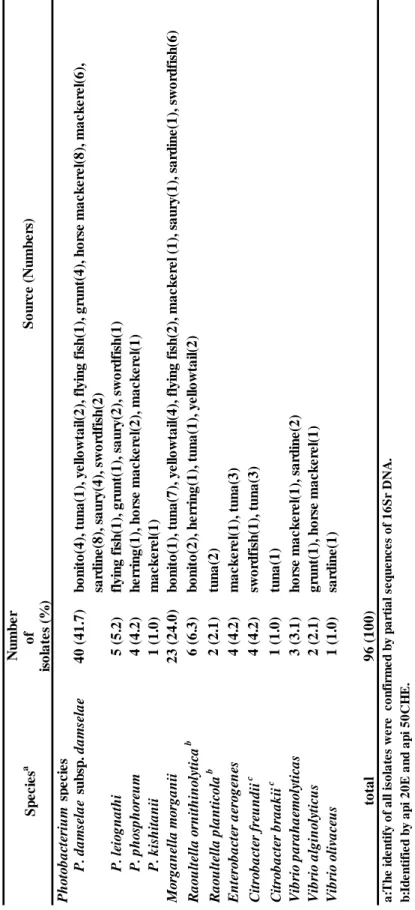
1.3.1. 生鮮魚からの中温性 Hm 生成菌の分離および同定 Table 1 に示すように 30ºC 培養下において 11 魚種 143 サンプル中 69 サンプル (48.3%)の培養液で Hm 生成が示され、その培養液から 96 株の中温性 Hm 生成菌 を分離した。中温性 Hm 生成菌は本実験で用いたすべての魚種から分離され、各魚 種における Hm 生成菌の分離率は 35.7%-66.7%を示した。分離率はトビウオが最も 高く、カツオが最も低かった。分離菌株のうち 50 株(51.0%)は Photobacterium 属 と同定され、内 40 株が P. damselae subsp. damselae、5 株が P. leiognathi、4 株が P.phosphoreum および 1 株が P. kishitanii と同定された。その他の菌株は、M. morganii
23 株、R. ornithinolytica 6 株、Citrobacter freundii 4 株、E. aerogenes 4 株、V.
parahaemolyticus 3 株、R. planticola 2 株、V. alginolyticus 2 株、V. olivaceus 1 株およ
び C. braakii 1 株と同定された(Table 3)。
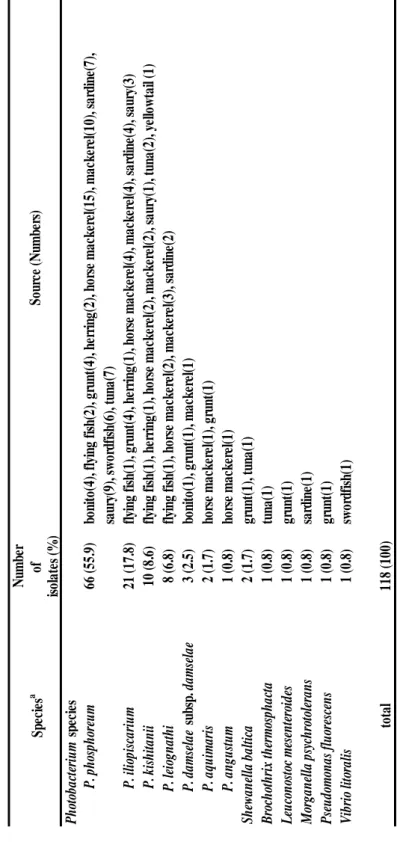
離した。低温性 Hm 生成菌は本実験で用いたすべての魚種から分離され、各魚種に おける Hm 生成菌の分離率は 10.0%-90.5%を示した。アジ・サバで 90.5%および 81.3%と高く、ブリで低かった(Table 1)。分離株は P. phosphoreum 66 株、P.
iliopiscarium 21 株、P. kishitanii 10 株、P. leiognathi 8 株、P. damselae subsp. damselae
3 株、P. aquimaris 2 株、Shewanella baltica 2 株、P. angustum 1 株、M. psychrotolerans 1 株、Leuconostoc mesenteroides 1 株、Brochothrix thermosphacta 1 株、V. litoralis 1 株 および Pseudomonas fluorescens 1 株と同定された(Table 4)。
1.3.3. 分離 Hm 生成菌の Hm 生成量 中温性 Hm 生成菌においては、96 株中 65 株が 1,000 mg/L 以上の Hm 生成を示し (Table 5)、最高値は 4,124 mg/L であった。中温性 Hm 生成菌で最も多量の Hm 生 成を示した菌株は M. morganii であった。その他 8 株が 500–1,000 mg/L、13 株が 200–500 mg/L、10 株が 100-200 mg/L の Hm 生成を示した。R. planticola、E. aerogenes、 R. ornithinplytica および V. alginolyticus は分離菌株数 2-6 株と多くないものの、す べての菌株において 1,000 mg/L 以上の Hm 生成が確認された。一方 C. freundii 4 株 は全て 100-200 mg/L の Hm 生成を示した。 低温性 Hm 生成菌は分離菌株 118 株のうち、22 株が 1,000 mg/L 以上の Hm 生成 を示し、最高値は P. phosphoreum の 2,268 mg/L であった。1000 mg/L 以上の Hm 生 成を示した 22 株中 16 株が P. phosphoreum でその他の菌株は P. kishitanii 3 株、M.
psychrotolerans、S. baltica および P. iliopiscarium 各 1 株であった。その他 27 株が
500–1,000 mg/L の、52 株が 200–500 mg/L の、17 株が 100-200 mg/L の Hm 生成を示 した(Table 6)。
1.4. 考察
本章においては、11 魚種 143 サンプルより中温性および低温性 Hm 生成菌の分 離を試みた。その結果、69 サンプルから中温性 Hm 生成菌が、82 サンプルから低 温性 Hm 生成菌が分離された(Table 1)。Hm 食中毒は、主に中温性 Hm 生成菌が 原因で発生していると考えられてきたが、いくつかの Hm 食中毒の原因菌として低 温性 Hm 生成菌が報告されている。本実験において、低温性 Hm 生成菌が、今回用 いた全ての魚種からの分離された事から、それらの魚種において低温性 Hm 生成菌 も Hm 蓄積を起こす可能性があることが再確認された。 本実験において分離した中温性 Hm 生成菌においては、過去の報告と同様に腸内 細菌科菌群、Photobacterium 属および Vibrio 属などの分離が確認された。その中でも最も多く分離されたのが P. damselae subsp. damselae(41.7%)で、次いで M.
morganii(24.0%)が多かった。本実験で分離された M. morganii は、23 株中 22 株
(95.7%)が 1,000 mg/L 以上の Hm 生成能を示した。M. morganii は以前より高い Hm 生成能を示すことが知られており(Kim et al., 2002, 2003; López-Sabater et al.,
1994)、Hm 食中毒の原因菌としての分離が報告されている(川端ら, 1956)。一方、
P. damselae subsp. damselae は、Hm 食中毒の原因菌としての報告例はないものの、
高い Hm 生成能を有していることは以前より知られていた(Bermejo et al., 2004;
Takahash et al., 2003)。本菌は本実験においても広範囲の魚種から分離され(Table 3)、
なおかつ分離された 40 株中 25 株(62.5%)が 1,000 mg/L 以上の Hm 生成を示した (Table 5)。以上の結果より、中温性 Hm 生成菌においては、M. morganii と同等以 上に P. damselae subsp. damselae に注意が必要と考えられる。
本実験において分離された低温性 Hm 生成菌 118 株中 111 株が Photobacterium 属 で、このうち 66 株が P. phosphoreum と同定された。P. phosphoreum は過去の研究に おいても主な低温性 Hm 生成菌として報告されている(Dalgaard et al., 2006; Fujii et
al., 1997; Morii and Kasama, 2004)。また、本実験において P. phosphoreum に次いで
多 く分離された P. iliopiscarium(17.8%)および P. kishitanii (8.6%)は,P.
phosphoreum の近縁種であり(Ast and Dunlap, 2005)、これまでの研究において Hm
生成菌として報告されていない菌種である。さらにこの 3 菌種において、97 株中 43 株(44.3%)が 500 mg/L 以上の高い Hm 生成が確認された。これらの結果は低 温貯蔵下において、P. phosphoreum およびその近縁種が Hm 蓄積の原因となること を示唆している。また、Photobacterium 属以外にも低温性 Hm 生成菌としては、M.
psychrotolerans、P. fluorescens、 S. baltica などの複数のグラム陰性菌が分離される
と と も に 、 グ ラ ム 陽 性 菌 で あ る Leuconostoc mesenteroides お よ び Brochothrix
thermosphacta が分離された(Table 4)。中でも、M. psychrotolerans、P. fluorescens、 S. baltica は以前より Hm 生成菌として報告されており(Emborg et al., 2005; Rivas et
al., 2005)、本実験で分離されたこれらの菌種のなかには、1,000 mg/L 以上の Hm 生 成を示した菌株も存在した(Table 6)。しかし、本実験においてこれらの菌種やグ ラム陽性菌は Photobacterium 属と比較して分離率が非常に低い。 低温性および中温性 Hm 生成菌双方において優勢であった Photobacterium 属は、 鮮魚において高い頻度で存在しており(与口ら, 1990b)、特に P. phosphoreum は魚 肉腐敗サンプルにおいても優占菌となりやすいことが報告されている(Dalgaard et
合、低温下においても短時間で増殖する可能性が高く、多量の Hm 蓄積を引き起こ すリスクが存在する事が考えられた。
Photobacterium 属は冷凍耐性が低いことから、マグロ、カツオおよびメカジキな
ど漁獲後に長距離輸送する事から冷凍状態で流通されることが多い魚種において (Hiraoka et al., 2004)、分離されにくい可能性が示唆されていた(Fujii et al., 1994)。 しかし本実験では、中温性・低温性 Hm 生成菌双方においてこれらの魚種からの Photobacterium 属の分離が確認されたことから、これらの Hm 生成菌が冷凍工程を 経ても残存することと、流通工程において汚染された可能性も考えられる。 食品中の Hm 量に関する規制については、我が国においては定められていないが、 複数の国において,規制値もしくは基準値が定められている。アメリカにおいて、 FDA はアジ、マグロ、カツオ、シイラなど Hm 食中毒が発生しやすい魚種におい て、50 mg/kg 以上の Hm 蓄積が確認された場合、検査を行った検体および検体と同 一ロットは廃棄することが定められている。FDA はその根拠として 500 mg/kg 以上 の Hm が蓄積された食品を摂取した場合、Hm 食中毒を発症するとしており、検体 の一部分に Hm が 50 mg/kg 以上存在していた場合、同一検体および同一ロット中 に 500 mg/kg 以上の Hm が蓄積している可能性があるとしている(FDA, 2011)。ま た,国際的な食品規格であり、多くの食品製造現場で適応されている Codex Alimentarius Commission(Codex)は、缶詰や冷凍魚といった特定の水産食品にお いて取り扱いおよび衛生基準として、食品中に 200 mg/kg 以上の Hm が存在するべ きではないと定めている(Codex alimentarius, 1981a, 1981b, 1981c, 1981d, 1989a,
1989b, 1995)。低温性 Hm 生成菌において 500 mg/L 以上の Hm 生成能を示す菌株の 割合は、中温性 Hm 生成菌と比較して低いが、18.6%の菌株が 1,000 mg/L 以上の Hm 生成を示した(Table 6)。そのため、たとえ Hm 食中毒が実際に発生しなくと も、水産加工品の海外輸出を行う上での規制対象となる可能性が存在する。この事 から、食品中における Hm 蓄積は、食品衛生上のみならず、経済上の観点からも抑 制が必要であると考えられる。 本章ではヒスチジン含量の高い 11 魚種において、Hm 食中毒の原因となり得る 中温性および低温性 Hm 生成菌が存在している事を示した。現在 Hm 食中毒が発生 した際に、原因菌を調査しない事が多く、原因菌を調査する場合でも中温性 Hm 生 成菌のみを対象として調査を行っている例が多いが(Chang et al., 2008; Chen et al.,
2010)、低温性 Hm 生成菌も同様に原因菌として注意するべきである。さらに、本
実験では P. phosphoreum や M. psychrotolerans など、低温性 Hm 生成菌として報告 のある菌種のみならず、P. iliopiscarium、P. kishitanii および P. aquimaris が Hm 生成
低温下における Hm 生成に関してより詳細な検討を行う必要があると考えられる。 第 2 章においては今回分離された P. phosphoreum および P. iliopiscarium のうち、高 い Hm 生成能を有する菌株を用いて、低温条件下における魚肉中での挙動を調査し た。
Table 1 Prevalence of Hm-producing bacteria in fish samples from retail markets in and near Tokyo Japan.
30°C 15°C 30°C 15°C Tuna 24 13 (54.2) 7 (29.2) 18 11 Horse mackerel 21 8 (38.1) 19 (90.5) 12 25 Mackerel 16 8 (50.0) 13 (81.3) 10 20 Bonito 14 5 (35.7) 3 (21.4) 7 5 Saury 14 6 (42.9) 10 (71.4) 7 13 Sardine 13 7 (53.8) 10 (76.9) 12 14 Grunt 11 5 (45.5) 7 (63.6) 6 13 Yellowtail 10 5 (50.0) 1 (10.0) 8 1 Swordfish 10 6 (60.0) 5 (50.0) 10 7 Flying fish 6 4 (66.7) 4 (66.7) 4 5 Herring 4 2 (50.0) 3 (75.0) 2 4 Total 143 69 (48.3) 82 (57.3) 96 118 Histamine-positive samples (%) Total samples Histamine-producing bacterial isolates Source
Table 2 Components of the PCR mixture for 16S rRNA PCR.
10×buffer (Takara Taq
TM)
5
µL
dNTP Mixture (Takara Taq
TM)
4
Forward Primer
5
Reverse Primer
5
Takara Taq (Takara Taq
TM)
0.25
Distilled water
25.75
DNA template
5
T able 3 I de nti fic ati on a n d pre va len ce of Hm -pr od uc ing bac ter ia l i solate s fr om re tail fish s ampl es incuba ted a t 30°C . N u m b e r o f is o la te s (% ) P h o to b a ct er iu m s p e c ie s P . d a m se la e su b sp . d a m se la e 4 0 (4 1 .7 ) b o n ito (4 ), tu n a (1 ), y e ll o w ta il (2 ), fl y in g fi sh (1 ), g ru n t(4 ), h o rs e m a c k e re l(8 ), m a c k e re l(6 ), sa rd in e (8 ), s a u ry (4 ), s w o rd fi sh (2 ) P . l ei o g n a th i 5 (5 .2 ) fl y in g fi sh (1 ), g ru n t(1 ), s a u ry (2 ), s w o rd fi sh (1 ) P . p h o sp h o re u m 4 (4 .2 ) h e rr in g (1 ), h o rs e m a c k e re l(2 ), m a c k e re l(1 ) P . k is h it a n ii 1 (1 .0 ) m a c k e re l(1 ) M o rg a n el la m o rg a n ii 2 3 (2 4 .0 ) b o n ito (1 ), tu n a (7 ), y e ll o w ta il (4 ), fl y in g fi sh (2 ), m a c k e re l (1 ), s a u ry (1 ), s a rd in e (1 ), s w o rd fi sh (6 ) Ra o u lt el la o rn it h in o ly ti ca b 6 (6 .3 ) b o n ito (2 ), h e rr in g (1 ), tu n a (1 ), y e ll o w ta il (2 ) Ra o u lt el la p la n ti co la b 2 (2 .1 ) tu n a (2 ) En te ro b a ct er a er o g en es 4 (4 .2 ) m a c k e re l(1 ), tu n a (3 ) Ci tr o b a ct er fr eu n d ii c 4 (4 .2 ) sw o rd fi sh (1 ), tu n a (3 ) Ci tr o b a ct er b ra a k ii c 1 (1 .0 ) tu n a (1 ) Vi b ri o p a ra h a em o ly ti ca s 3 (3 .1 ) h o rs e m a c k e re l(1 ), s a rd in e (2 ) Vi b ri o a lg in o ly ti cu s 2 (2 .1 ) g ru n t(1 ), h o rs e m a c k e re l(1 ) Vi b ri o o li va ce u s 1 (1 .0 ) sa rd in e (1 ) to ta l 9 6 (1 0 0 ) a :T h e i d e n ti fy o f a ll i so la te s w e re c o n fi rm e d b y p a rt ia l se q u e n c e s o f 1 6 S r D N A . b :I d e n ti fi e d b y a p i 2 0 E a n d a p i 5 0 C H E . c :I d e n ti fi e d b y V IT E C s y st e m . S p e c ie s a S o u rc e (N u m b e rs )
T able 4 Ide nti fic ati on a n d pre va len ce of Hm -pr od uc ing bac ter ia l isol ates f rom re tail fish s ampl es incuba ted at 15 °C . N um be r of is ol ate s (% ) P h ot ob ac te ri u m s pe ci es P . p h os ph or eu m 66 (5 5. 9) bo ni to (4 ), fl yi ng fi sh (2 ), g ru nt(4 ), h er ri ng (2 ), h or se m ac ke re l(1 5), m ac ke re l(1 0), s ar di ne (7 ), sa ur y(9 ), s w or dfi sh (6 ), tu na (7 ) P . i li op is ca ri u m 21 (1 7. 8) fl yi ng fi sh (1 ), g ru nt(4 ), h er ri ng (1 ), h or se m ac ke re l(4 ), m ac ke re l(4 ), s ar di ne (4 ), s au ry (3 ) P . k is h it an ii 10 (8 .6 ) fl yi ng fi sh (1 ), h er ri ng (1 ), h or se m ac ke re l(2 ), m ac ke re l(2 ), s au ry (1 ), tu na (2 ), y el lo w ta il (1 ) P . l ei og n at h i 8 (6 .8 ) fl yi ng fi sh (1 ), h or se m ac ke re l(2 ), m ac ke re l(3 ), s ar di ne (2 ) P . d am se la e su bs p. d am se la e 3 (2 .5 ) bo ni to (1 ), g ru nt(1 ), m ac ke re l(1 ) P . a qu im ar is 2 (1 .7 ) ho rs e m ac ke re l(1 ), g ru nt(1 ) P . a n gu st u m 1 (0 .8 ) ho rs e m ac ke re l(1 ) S h ewa n el la b al ti ca 2 (1 .7 ) gr un t(1 ), tu na (1 ) Br oc h ot h ri x th er m os ph ac ta 1 (0 .8 ) tu na (1 ) L eu co n os to c m es en te ro id es 1 (0 .8 ) gr un t(1 ) M or ga n el la p sy ch ro to le ra n s 1 (0 .8 ) sa rd in e(1 ) P se u do m on as fl u or es ce n s 1 (0 .8 ) gr un t(1 ) Vi br io li to ra li s 1 (0 .8 ) sw or dfi sh (1 ) to ta l 11 8 (1 00 ) a: T he id en ti fy o f al l i so la te s w er e c on fi rm ed b y pa rt ia l s eq ue nc es o f 16 S r D N A . S pe ci es a S ou rc e (N um be rs )
Table 5 Histamine production by isolated mesophilic HPB. 100-200 200-500 500-1,000 >1,000 R. planticola (2) 2 E. aerogenes (4) 4 M. morganii (23) 1 22 R. ornithinolytica (6) 6 V. alginolyticus (2) 2
P. damselae subsp. damselae (40) 9 6 25
V. parahaemolyticas (3) 1 2 P. leiognathi (5) 3 1 1 P. phosphoreum (4) 1 2 1 V. olivaceus (1) 1 P. kishitanii (1) 1 C. braakii (1) 1 C. freundii (4) 4 total 10 13 8 65
Table 6 Histamine production by isolated psychrophilic HPB. 100-200 200-500 500-1,000 >1,000 M. psychrotolerans (1) 1 P. fluorescens (1) 1 P. kishitanii (10) 3 2 2 3 S. baltica (2) 1 1 P. phosphoreum (66) 8 28 14 16 P. aquimaris (2) 1 1 P. iliopiscarium (21) 1 12 7 1 L. mesenteroides (1) 1
P. damselae subsp. damselae (3) 2 1
B. thermosphacta (1) 1 V. litoralis (1) 1 P. leiognathi (8) 4 3 1 P. angustum (1) 1 total 17 52 27 22 histamine production (mg/L) Strains (No.)
第 2 章 低温性ヒスタミン生成菌 Photobacterium phosphoreum および
P. iliopiscarium の低温保存下における魚肉中での挙動
2.1.緒言
低温性 Hm 生成菌に関する研究は中温性 Hm 生成菌と比べ少ないものの、 Okuzumi et al.(1981)によって「N 菌群」が報告されて以降、様々な研究が報告さ れており、特に P. phosphoreum に関する研究が多い(石本ら, 1994; Kanki et al., 2004; 栗原ら, 1993a, 1993b)。P. phosphoreum は 35ºC においては発育が見られない一方、
4ºC において増殖および Hm 生成を行う事が知られている(Fujii et al., 1994)。また、
本菌は温度帯によっては、主要な中温性 Hm 生成菌として知られる M. morganii よ りも高い Hm 生成能を示すこともあり(Lehane and Olley, 2000)、実際に、Hm 食中 毒の原因菌として報告された例もある(Kanki et al., 2004)。 第 1 章において、鮮魚からの低温性および中温性 Hm 生成菌の分離、同定を行っ た結果、P. phosphoreum に次いで今まで Hm 生成菌として報告されていなかった P. kishitanii および P. iliopiscarium が分離され、これらの中には 1,000 mg/L 以上の高い Hm 生成を示す菌株が存在した。特に P. phosphoreum に次いで分離率が高かった P. iliopiscarium は、なおかつ 4ºC でも増殖が可能である事が報告されている(Rivas et al., 2006)。 本章においては、これらの低温性 Hm 生成菌のうち、低温下において高い Hm 生 成能の高い P. phosphoreum および P. iliopiscarium 菌株を魚肉に接種し、4°C、10°C および 15°C における挙動を検討した。
2.2. 実験方法
2.2.1. サンプルおよび供試菌株 冷凍メカジキ(Xiphias gladius)の切り身を埼玉県の小売店より購入し,供試菌 を接種する直前に解凍し,本実験に用いた。 第 1 章において鮮魚より分離した低温性 Hm 生成菌のうち,500 mg/L 以上の Hm 生成能を示した P. phosphoreum 7 株(4A1525、5G1524、6C1528、6H1523、7D155、 9A154、9D1518)、P. iliopiscarium 2 株(6C1521、7I1519)を用いた。接種実験に は P. phosphoreum 7D155 株および P. iliopiscarium 7I1519 株を用いた鮮魚由来の P. phosphoreum 7 株および P. iliopiscarium 2 株を 15ºC、48 時間で前培 養後、ヒスチジンブロスに接種し、4ºC において培養後 0、72、96 および 120 時間 後に生菌数および Hm 量を測定した。生菌数は、1.5% NaCl 添加 Trypticase Soy Agar (TSA)(BD)に培養液 100 μL を塗末し、15ºC、72 時間培養後に測定した。Hm 量は、菌液 1 mL を 0.2-μm フィルター(東洋濾紙)でろ過滅菌後、チェックカラ ーヒスタミンを用いて測定した。 2.2.3. 魚肉中における挙動 2.2.3.1. 魚肉への接種 冷凍魚肉サンプルを流水解凍後、1 cm×1 cm に切り分け、25 g を無菌的に量り
取った。供試菌株(P. phosphoreum 7D155、P. iliopiscarium 7I1519)を終濃度 104
cfu/g となるように接種し、4ºC、10ºC および 15ºC において保存した。4ºC 保存下におい て 0、3、5、7 日後に、10ºC 保存下においては 0、1、2、3 日後に、15ºC 保存下に おいては 0、12、24、36、48 時間後に一般生菌数、Hm 生成菌数および Hm 量測定 を行った。本実験において、対照群として未接種のサンプルを用いた。未接種サン プルは接種サンプルと同様に各温度で保存後、各項目の測定を行った。実験は同一 サンプルを用いて 3 回行った。 2.2.3.2. 一般生菌数および Hm 生成菌数の測定 サンプル 25 g に対し、225 mL のヒスチジンブロスを加え 10 倍希釈後、ストマ ッカー400(Seward、Worthing、West Sussex)を用いて懸濁した。必要に応じて食 品懸濁液を生理的食塩水で 10 倍に段階希釈後、1.5% NaCl 添加 TSA に 100 μL 塗抹 し、15ºC、72 時間培養後、菌数を測定した。Hm 生成菌数の測定には最確数法(MPN 法)を用いた(Cochran, 1950)。食品懸濁液の各段階希釈液をヒスチジンブロス 3 本に接種し、15ºC、48 時間培養した。培養後、Hm 生成をペーパークロマトグラフ ィーによって確認し、Hm 陽性管数から最確数表を用いて Hm 生成菌数を求めた。 ペーパークロマトグラフィーは第 1 章と同様の方法で行った。 2.2.3.3. 魚肉中における Hm 量の測定 魚肉サンプル 1 g に対し,24 mL の 0.1M EDTA/sodium 溶液(pH 8)を添加し、 20 分間の煮沸を行った。加熱後、冷却したサンプルを 10,000xg、5 分、4ºC で遠心 を行い、上澄みを Hm 量測定に用いた。Hm 量は、チェックカラーヒスタミン(キ
2.3.結果
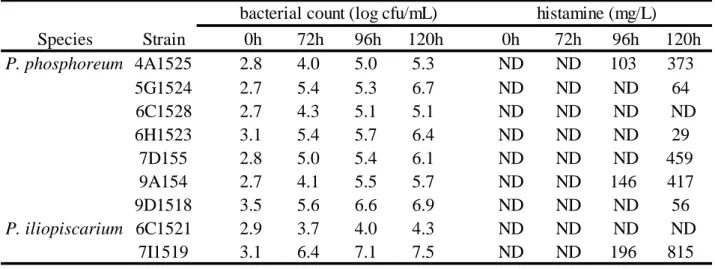
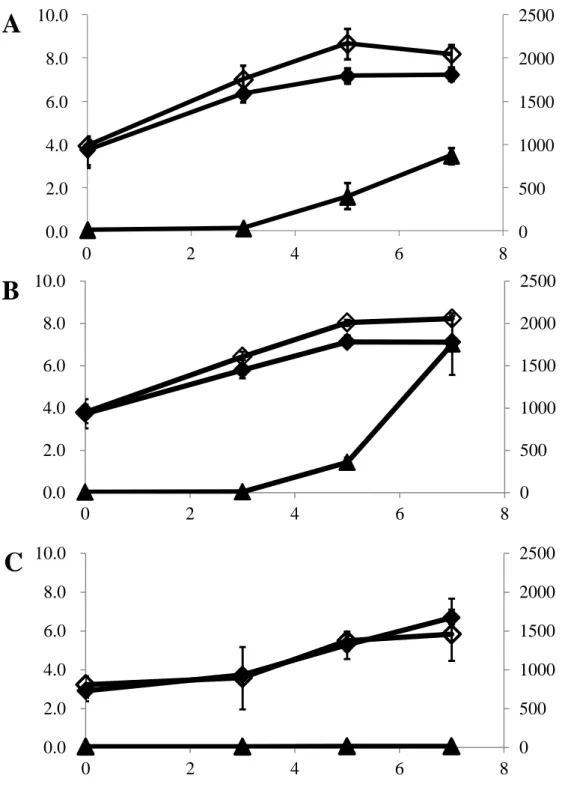
2.3.1. 4°C 培養下における Hm 生成能 第 1 章において鮮魚より分離された P. phosphoreum および P. iliopiscarium から、 9 株の 4ºC 培養下における挙動を調べた(Table 7)。その結果、9 株中 7 株で 120 時 間後に 29-815 mg/L の Hm 生成が確認された。中でも、P. phosphoreum 7D155 が 459 mg/L、P. iliopiscarium 7I1519 が 815 mg/L の Hm 生成を示した。以上の結果より、 7D155 および 7I1519 株を以下の実験に用いた。 2.3.2.魚肉中における低温性 Hm 生成菌の挙動 Fig. 3-5 に低温保存下における魚肉へ Hm 生成菌を接種した場合の Hm 生成菌数、 一般生菌数および魚肉中の Hm 量の変動を示した。 4ºC 保存下において、P. phosphoreum 7D155 を接種したサンプルの Hm 生成菌数 は、培養 3 日目に 106 MPN/g、5 日目までに 108 MPN/g に達した。さらに魚肉中に おける Hm 量は、5 日目に 400 mg/kg、7 日目には 870 mg/kg であった(Fig. 3A)。 P. iliopiscarium 7I1519 を接種したサンプルにおいて Hm 生成菌数は、培養 3 日目に 106 MPN/g、5 日目に 108 MPN/g に達した。Hm 量は、5 日目に 360 mg/kg, 7 日目に は 1,760 mg/kg に達した(Fig. 3B)。両接種サンプルにおいては、7 日目まで魚肉の 変色および腐敗臭が確認されず、腐敗は見られなかった。対照群において Hm 生成 菌数は、7 日目までに 105 MPN/g に達したが、Hm の生成は確認されなかった(Fig. 3C)。 10ºC 保存下において、7D155 を接種したサンプルの Hm 生成菌数は培養 2 日目 に 106 MPN/g に到達した。Hm 量は、2 日目に 100 mg/kg、3 日目に 220 mg/kg に達 した(Fig. 4A)。7I1519 を接種したサンプルにおいても、2 日目に Hm 生成菌数が 106 MPN/g に達した。Hm 量は、2 日目に 140 mg/kg、3 日目には 170 mg/kg であっ た(Fig. 4B)。両接種サンプルは保存 3 日目において僅かに魚肉の色に変化がみら れたものの、腐敗臭は確認されなかった。対照群でも Hm 生成菌数は 106 MPN/g に達したが、Hm 生成は確認されなかった(Fig. 4C)。 15ºC 保存下において、7D155 あるいは 7I1519 を接種したサンプルは共に、Hm 生成菌数が 12 時間後に 106 MPN/g、36 時間後に 108 MPN/g に達した。Hm 量は、 7D155 接種サンプルにおいて 24 時間後には 90 mg/kg、36 時間後には 420 mg/kg、48 時間後には 980 mg/kg であった(Fig. 5A)。7I1519 接種サンプルの場合は、24 時
間後で 170 mg/kg の、36 時間後で 240 mg/kg の、48 時間後で 610 mg/kg の Hm 生成 が確認された(Fig. 5B)。両接種サンプルでは 48 時間後には腐敗が確認された。対
照群では 48 時間に一般生菌数が 108 cfu/g に達したが、Hm 生成菌数は 106 MPN/g であった。しかし、魚肉中における Hm の生成は確認されなかった(Fig. 5C)。
2.4. 考察
本実験において、低温下で高い Hm 生成を示した鮮魚由来の低温性 Hm 生成菌 2 菌種 P. phosphoreum および P. iliopiscarium を解凍メカジキに接種し、4ºC、10ºC お よび 15ºC で保存した結果、4ºC では 5 日後、10ºC では 2 日後、15ºC では P. iliopiscarium 7I1519 において 24 時間後に 100 mg/kg 以上の、P. phosphoreum 7D155 において 50 mg/kg 以上の Hm 蓄積が確認された(Fig. 3A、Fig. 3B、Fig. 4A、Fig. 4B、Fig. 5A、Fig. 5B)。食品中における Hm の規制値や基準値としては、上述したように FDA は 規制値として 50 mg/kg と定めている。本実験では、全ての保存区分の接種サンプ ルにおいて、短時間で 50 mg/kg 以上の Hm 蓄積が確認された。この結果より、本 実験で用いられたような低温性 Hm 生成菌が魚肉中に存在していた場合、短時間で 魚肉中の Hm 量がそれらの規制値・基準値に達する可能性が存在し、Hm 食中毒の 発生しやすい魚種の冷凍魚や加工品の輸出時に影響が出る可能性が考えられた。 4ºC および 10ºC 保存下では、Hm 生成が確認された際、わずかに魚肉の色に変化 が見られたものの、腐敗臭は確認されなかった。特に 4ºC 保存下において、P. phosphoreum 7D155 は、1 週間以内に魚肉中で腐敗に先行して 1,000 mg/kg 以上の Hm 生成を示した。魚肉が腐敗してしまった場合、食用とされる事はないが、低温 保存下において腐敗に先行して Hm が蓄積された場合、食材として用いられるので、 Hm 食中毒の原因となる可能性が考えられる。
本実験では、P. iliopiscarium 7I1519 は P. phosphoreum 7D155 と同様の低温下にお
ける増殖能および Hm 生成能を示した。Hm 生成菌は 106 cfu/ml に達してから Hm 生成する事が報告されている(Takahashi et al., 2003)。本実験の魚肉接種実験でも、 すべての温度区分で Hm 生成菌数が 106 MPN/g 以上に達してから Hm 蓄積が確認さ れた。このことから、低温性 Hm 生成菌が低温下で迅速に増殖することによって、 短時間で Hm 蓄積を行う可能性が示唆された。そのため、本実験で示されたように、 低温下で高い Hm 生成能を示す低温性 Hm 生成菌が存在していた場合、魚肉中で短 時間に多量の Hm 蓄積が発生する危険性が存在する。 マグロやカツオなど通常冷凍状態で流通される魚種が低温性 Hm 生成菌によっ て汚染されていた場合、解凍工程において Hm 蓄積の発生が起こる可能性が報告さ
ン食中毒防止マニュアル)。さらに、冷凍工程などにおいて死滅または VBNC (Viable But Non-Culturable)状態であったとしても、細菌由来の HDC の作用によ って Hm 蓄積が発生する可能性もある。Kanki et al.(2007)は、P. phosphoreum 由 来の HDC は冷凍条件下でも活性を 3 ヶ月以上失わず、なおかつ至適温度において 酵素反応を示し、Hm 生成が行われることを報告している。そのため、冷凍魚にお いて Hm 生成菌が死滅もしくは損傷状態であっても HDC の酵素反応が可能な温度 条件に置かれた場合、食品中における Hm 蓄積が発生すると考えられる。 本章においては、魚肉中に接種した P. phosphoreum および P. iliopiscarium の低温 下における挙動を調査し、低温下においても Hm 蓄積のリスクが存在し、従来 Hm 食中毒の予防手段として考えらえてきた温度管理のみでは食品中における Hm 蓄 積を完全に制御することが困難である事を示した。この事から、Hm 蓄積の予防と 共に、多量の Hm が蓄積した食品が Hm 流通することを防ぐ事が重要だと考えられ る。さらに、食品中における Hm 蓄積や食中毒が発生した際には迅速な原因究明や 対策を行う事が必要とされることから、原因菌となる Hm 生成菌の迅速な同定が必 要となる。そのため、第 3 章においては、低温性 Hm 生成菌も含めたグラム陰性 Hm 生成菌共通の hdc 遺伝子を用いた迅速同定を試みた。
Table 7 Viable count and Hm content in Histidime broth incubated at 4ºC of P.
phosphoreum and P. iliopiscarium isolates used for the screening strong Hm producers.
Species Strain 0h 72h 96h 120h 0h 72h 96h 120h P. phosphoreum 4A1525 2.8 4.0 5.0 5.3 ND ND 103 373 5G1524 2.7 5.4 5.3 6.7 ND ND ND 64 6C1528 2.7 4.3 5.1 5.1 ND ND ND ND 6H1523 3.1 5.4 5.7 6.4 ND ND ND 29 7D155 2.8 5.0 5.4 6.1 ND ND ND 459 9A154 2.7 4.1 5.5 5.7 ND ND 146 417 9D1518 3.5 5.6 6.6 6.9 ND ND ND 56 P. iliopiscarium 6C1521 2.9 3.7 4.0 4.3 ND ND ND ND 7I1519 3.1 6.4 7.1 7.5 ND ND 196 815
Fig. 3 Growth of histamine-producing bacteria (open diamonds), aerobic viable bacteria (closed diamonds) and histamine accumulations (triangles) in swordfish meat at 4°C. A: Inoculated with strain 7D155 (P. phosphoreum), B: Inoculated with strain 7I1519 (P.
iliopiscarium), C: Control, Error bars represent standard deviation of three independent
experiments. 0 500 1000 1500 2000 2500 0.0 2.0 4.0 6.0 8.0 10.0 0 2 4 6 8
A
0 500 1000 1500 2000 2500 0.0 2.0 4.0 6.0 8.0 10.0 0 2 4 6 8B
0 500 1000 1500 2000 2500 0.0 2.0 4.0 6.0 8.0 10.0 0 2 4 6 8C
Fig. 4 Growth of histamine-producing bacteria (open diamonds), aerobic viable bacteria (closed diamonds) and histamine accumulations (triangles) in swordfish meat at 10°C. A: Inoculated with strain 7D155 (P. phosphoreum), B: Inoculated with strain 7I1519 (P.
0 100 200 300 400 0.0 2.0 4.0 6.0 8.0 10.0 0 1 2 3 4
A
0 100 200 300 400 0.0 2.0 4.0 6.0 8.0 10.0 0 1 2 3 4B
0 100 200 300 400 0.0 2.0 4.0 6.0 8.0 10.0 0 1 2 3 4C
Log MP N ( cfu) /g Hist ami ne ( mg/ kg) (( mg/ kg ))Fig. 5 Growth of histamine-producing bacteria (open diamonds), aerobic viable bacteria (closed diamonds) and histamine accumulations (triangles) in swordfish meat at 15°C. A: Inoculated with strain 7D155 (P. phosphoreum), B: Inoculated with strain 7I1519 (P.
iliopiscarium), C: Control, Error bars represent standard deviation of three independent
experiments. 0 200 400 600 800 1000 1200 1400 0.0 2.0 4.0 6.0 8.0 10.0 0 15 30 45 60
A
0 200 400 600 800 1000 1200 1400 0.0 2.0 4.0 6.0 8.0 10.0 0 15 30 45 60B
0 200 400 600 800 1000 1200 1400 0.0 2.0 4.0 6.0 8.0 10.0 0 15 30 45 60C
Hist ami ne ( mg/ kg) (( mg/ kg )) Log MP N ( cfu) /g第 3 章 高感度融解曲線解析(HRMA)を用いた
ヒスタミン生成菌の迅速同定法の検討
3.1. 緒言
食中毒の予防および原因究明を行う上で、原因微生物の特定は必要不可欠である。 現在も微生物検査の微生物同定における公定法としては、現在も基本的に培養法や 性状試験が広く用いられている。しかし、これらの手法は特異的な培地や数日から 一週間程度の時間を必要とする場合があり、なおかつ同種株間においても性状に差 異が出る場合がある(Drancourt et al., 2000)。一方、遺伝子的手法は、培養法と比 較して短時間で同定が可能という利点を持っており、これまでに様々な研究が行わ れてきた(Keer and Birch, 2003)。遺伝子的手法による同定法として、ポリメラーゼ 連鎖反応(Polymerase Chain Reaction: PCR)およびシーケンシングによって得られ た配列を用いた同定が行われており、特に 16S rDNA 遺伝子を始めとする遺伝子配 列の保存性の高い領域を用いたシーケンシングによる菌種同定が広く用いられて いる(Drancourt et al., 2000; Weisburg et al., 1991)。近年、迅速・簡易・低コストな微生物同定の手法として、高感度融解曲線解析法 (High-resolution melting Analysis: HRMA)が注目されている。本手法では、加熱変 性により 2 本鎖 DNA から 1 本鎖へ乖離する際の温度が、その DNA の GC 含量、 長さおよび配列に依存していることを利用し、サンプル間の遺伝子配列の差異を調 べる事ができる(Gundry et al., 2003)。本手法で用いられる SYBR-Green (Gundry et
al., 2003)、RezoLight(Pietzka et al., 2009)および LCGreen(Wittwer et al., 2003)等
の 2 本鎖 DNA に結合して蛍光を発する DNA 結合色素は、2 本鎖 DNA の状態では 蛍光を発するため、DNA の加熱変性による1本鎖への乖離に伴って蛍光量が減少 する。HRMA ではその蛍光の減少をモニタリングすることで得られる乖離温度(融
解温度:Tm 値)および融解曲線から、サンプル間の遺伝子配列の違いを示す事が
可能である(Erali et al., 2008)。
HRMA に関する研究が報告される以前にも、リアルタイム PCR を用いた融解曲 線解析による遺伝子型決定に関する研究が報告されている(Lay and Wittwer, 1997; Ririe et al., 1997)が、これらの手法においては、DNA 増幅後に別の系で DNA 結合 色素を添加してから加熱融解を行っていた。HRMA では閉管系で PCR から融解曲 線解析まで行うことができ(Gundry et al., 2003)、なおかつ、一塩基多型(Single
2003)。
HRMAはその簡便性から、ヒト遺伝子(Vossen et al., 2009)や作物(Koeyer et al., 2010)の変異調査など、異なる遺伝子配列の識別法として様々な分野で用いられて いる。微生物を対象とした研究も、Mycobacterium chelonae/M. abscessus間の迅速同 定に関する研究(Odell et al., 2005)を始めとして、現在まで多くの研究が報告され ており、近年ではウィルスを対象としたHRMA法も報告されている(Lin et al., 2008;
Steer et al., 2009)。微生物におけるHRMAの利用方法としては主に、タイピング法
もしくは迅速同定法としての利用が挙げられる。タイピング法の利用例としては、 複数遺伝子のSNPsを用いたYersinia pseudotuberculosisのタイピング(Souza and Falcão, 2013)や、乳製品腐敗菌であるBacillus licheniformis(Dhakal et al., 2013)に おけるタイピング等が報告されている。さらに、今までに報告されたHRMAを用い た迅速同定法に関する報告としては、16S rDNAを標的遺伝子とした病原菌の迅速 検出に関する報告(Ajitkumar et al., 2012; Cheng et al., 2006; Navrátilová et al., 2013; Robertson et al., 2009; Šimenc and Potočnik, 2011)が多く、近縁種間における迅速同 定に関する報告もされている(Arancia et al., 2011; Gori et al., 2012; Pietzka et al.,
2009; Winchell et al., 2010)。また、今まで報告されたHRMA法では、対象の多くが
Mycobacterium(Alonso et al., 2010; Odell et al., 2005; Pietzka et al., 2009)、ブルセラ菌
(Winchell et al., 2010)やカンジダ菌(Arancia et al., 2011)などの臨床微生物であ ったが、近年、食中毒原因菌(Campylobacter jejuni, Listeria属, 腸管出血性大腸菌) を始めとした食品微生物を対象とした報告も増加している(Jin et al., 2012; Kagkli et
al., 2012; Price et al., 2007)。
本研究の第 1 章および第 2 章において、中温性 Hm 生成菌と同様に、低温性 Hm 生成菌も広範囲の魚種に分布し、なおかつ、低温保存下においても低温性 Hm 生成 菌が短期間で Hm 蓄積を引き起こす可能性がある事を示した。このことから、中温 性のみならず、低温性 Hm 生成菌も Hm 食中毒の原因となり得るリスクを有してい ると考えられる。本章においては、本研究では、新たに Hm 生成菌として分離され た菌種も含めた低温性および中温性 Hm 生成菌の迅速な同定を行うことを目的と
し、中温性 Hm 生成菌 6 菌種(M. morganii、P. damselae subsp. damselae、E. aerogenes、
R. planticola、P. vulgaris、Erwinia spp.)および低温性 Hm 生成菌 4 菌種(P. phosphoreum, P. iliopiscarium, P. kishtianii, P. aquimaris)を対象とし、HRMA による同定法の検討
を行った。