学位論文
神経障害性疼痛の治療薬に関する研究
Studies on the analgesics for the treatment of neuropathic pain
2017 年
i 目次 諸言 ... 1 第一章 神経障害性疼痛への炎症反応の関与 ... 4 1.1 実験材料・方法 ... 5 1.1.1 実験動物 ... 5 1.1.2 被験薬 ... 5 1.1.3 神経障害性疼痛モデルの作製 ... 5 1.1.4 疼痛行動の評価 ... 7 1.1.5 実験プロトコール ... 8 1.1.6 組織標本の作製 ... 10 1.1.7 ヘマトキシリン・エオジン染色 ... 10 1.1.8 免疫染色 ... 10
1.1.9 Total RNA の抽出及び逆転写反応による cDNA の合成... 10
iii 略語表
本文中では以下の略語を用いる。
BCTC N-(4-tertiarybutylphenyl)-4-(3-chloropyridin-2-yl)tetrahydropyrazine-1(2H)-carbox-amide CCI chronic constriction injury
CGRP calcitonin gene-related peptide CHO chinese hamster ovary
DRG dorsal root ganglion
EC50 50% effective concentration GFAP glial fibrillary acidic protein HE hematoxylin-Eosin
HIV human immunodeficiency virus
Iba 1 ionized calcium-binding adapter molecule 1 IgG immunoglobulin G JWH133 (6aR,10aR)-3-(1,1-Dimethylbutyl)-6a,7,10,10a-tetrahydro-6,6,9-trimethyl-6H-dibenzo[b,d]pyran IL-1 interleukin 1 KO knockout LSP lipopolysaccharide
MCP-1 monocyte chemotactic protein 1 NF200 neurofilament 200
NGF nerve growth factor PCR polymerase chain reaction PKA protein kinase A
PKC protein kinase C
PSL partial sciatic nerve ligation PWT paw withdrawal threshold QOL quality of life
RTX resiniferatoxin
TNF-α tumor necrosis factor alpha
1 諸言 疼痛は、一般的には痛みと呼ばれ、学術的には「組織の実質的あるいは潜在的な傷害に結 びつくか、このような傷害を表す言葉を使って述べられる不快な感覚及びその情動体験」と 定義される1) 。疼痛を誘発する侵害刺激は、末梢の侵害受容器で感知され、電気的興奮(活 動電位)へと変換される。この活動電位は、Aδ 線維及び C 線維の 2 種類の一次求心性感覚 神経から後根神経節(dorsal root ganglion、DRG)を経て脊髄後角へと伝達される。これら一
3
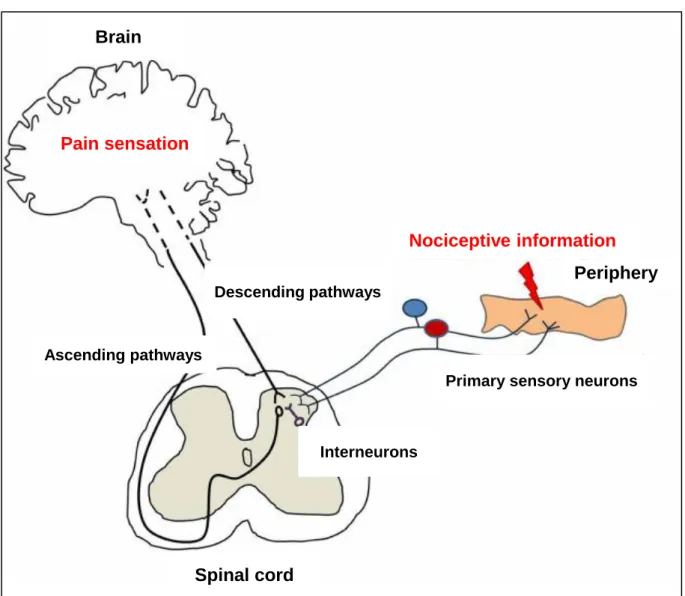
Figure 1 Schematic diagram of pain pathways involved in pain transmission and modulation. Nociceptive information is transmitted from the periphery to the spinal dorsal horn by primary sensory neurons. At the spinal level, these neurons transmit nociceptive information to second order neurons (“Ascending pathways”) through the release of neurotransmitters like the excitatory amino acids glutamate and aspartate, calcitonin gene-related peptide, substance P galanin and neuropeptide Y. In the brain, the nociceptive information is then perceived as a pain sensation. The transmission of nociceptive information at the spinal level is modulated by interneurons (mainly inhibitory) through the release of opioid peptides and GABA and also by supraspinal descending neurons (“Descending pathways”) through the release of serotonin and noradrenaline. Descending pathways may inhibit or enhance nociceptive transmission from the spinal cord. Figure 1 modified from Tavares et al2).
Primary sensory neurons
5 1.1 実験材料・方法 本実験計画は、帝人ファーマ株式会社 生物医学総合研究所の動物実験委員会にて審査され た。その後、承認されたプロトコールに基づいて本実験が実施された。 1.1.1 実験動物 雄性 ddY 系マウス(16-18 g、日本 SLC、浜松)を用いた。実験に使用するまで明暗サイク ル 12 時間(明期 7:00-19:00)、室温 23±1℃、湿度 52±2%の一定環境下で動物を飼育した。 飼養中の動物には水及びマウス用固型飼料を自由に摂取させた。 1.1.2 被験薬 免疫グロブリン製剤(乾燥スルホ化人免疫グロブリン、献血ベニロン®)は財団法人化学及 血清療法研究所より分与された。免疫グロブリン製剤は付属の溶解液で溶解させ、50 mg/mL 溶液を調製した。 1.1.3 神経障害性疼痛モデルの作製
神経障害性疼痛モデルとして坐骨神経絞扼モデル(chronic constriction injury、CCI モデル)、 坐骨神経部分結紮モデル(partial sciatic nerve ligation、PSL モデル)及び脊髄神経結紮モデル
(spinal nerve ligation、SNL モデル)が汎用されている25)。これらモデルは、後肢を支配する
6
Figure 2 Schematic of representative experimental models of neuropathic pain 25).
The schematic illustrates the three main rodent models of neuropathic pain associated with direct nerve injury; the L5-L6 spinal nerve ligation (SNL) model, the chronic constriction injury (CCI) of the sciatic nerve model, and the partial sciatic nerve ligation (PSL) model.
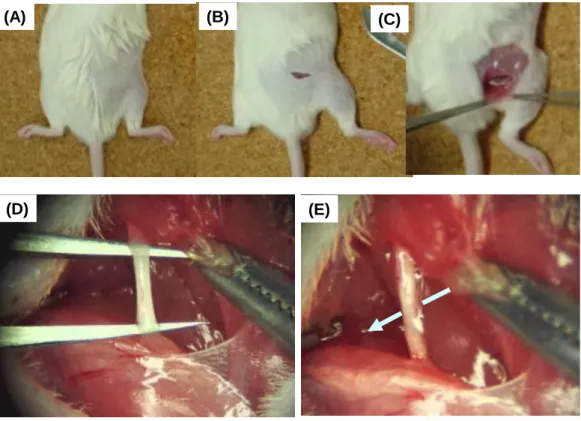
Figure 3 Surgical procedures for the mice with PSL.
(A-E) The surgical procedures for the mice with PSL were shown. The mice were anaesthetized with isoflurane in oxygen during surgery. Partial sciatic nerve ligation (PSL) was induced by the sciatic nerve ligation with 9-0 silk suture around approximately one-half the diameter of the nerve under light microscope.
(A)
(D) (E)
7 1.1.4 疼痛行動の評価 神経障害性疼痛モデルの疼痛行動を評価するため、刺激に対する逃避行動を測定した。測 定方法は Tal 及び Bennett らがラットで報告した方法27) を一部変更して行った(Figure 4)。 マウスを床面が網目状になった実験架台に乗せ、約 1 時間実験環境に馴化させた。その後、 直径の異なる 9 本の von Frey(VF)フィラメント(Touch-Test Sensory Evaluator Instruction、 North Coast Medical Inc.、San Jose、CA、USA)(0.008、0.02、0.04、0.07、0.16、0.4、0.6、 1.0 及び 1.4 g)を用いてマウス後肢足底を刺激した。刺激は、最も細い VF フィラメントから 順に行った。逃避行動が認められた最小の VF フィラメント刺激を、そのマウスの疼痛閾値 (paw withdrawal threshold、PWT)とした。正常マウスは、通常、0.4 g 以上の VF フィラメン ト刺激で逃避行動を示す。一方、坐骨神経束を結紮されたマウスは、正常マウスでは逃避行 動を示さない細い VF フィラメント刺激でも逃避行動を示すようになる。本章では疼痛閾値 が 0.04 g 以下になることを神経障害性疼痛発症と判定し、本実験で疼痛モデルとして使用し た。
9
Figure 5 Experimental protocols of immunoglobulin G treatment for the neuropathic pain. PSL; partial sciatic nerve ligation, IgG; immunoglobulin G, PWT; paw withdrawal threshold, i.p.; intraperitoneal administration
Day1 Day0
(PSL or sham) Day14
Sampling
Protocol 1: Histopathological examination of sciatic nerve and spinal cord in mice with PSL. Day5 Sampling Sampling Day5 Day3 Day0
(PSL or sham) Day14 Day8
Day-1 Day10
PWT measurement
Vehicle or IgG, i.p.
Day21 Day-2
Protocol 2: Effects of IgG on the development of neuropathic pain in mice with PSL.
Day18 Day30
Day-1 Day11
PWT measurement Vehicle or IgG, i.p.
Day36 Day24
Day0
(PSL or sham) Day15
Protocol 3: Effects of IgG on neuropathic pain in mice with PSL.
Day1 Day0
(PSL or sham) Day-1
Vehicle or IgG, i.p.
Sampling
Day3 Day0
(PSL or sham) Day-1
Vehicle or IgG, i.p.
Day5 Sampling Protocol 4: Effects of IgG on mRNA expression of inflammatory cytokines in mice
10 1.1.6 組織標本の作製 PSL マウス及び Sham マウスをイソフルランによる深麻酔下、100 mL の 10 mmol/L phosphate-buffered saline(pH7.2-7.4)(PBS)を経心的に灌流し、組織内の血液を除去した後、 4%パラホルムアルデヒド含有 PBS 100 mL で灌流固定した。灌流固定後、右大腿部を切開し、 坐骨神経束(結紮部位を含む)を約 1 cm 採取した。脊椎を切り出し、腰髄(第四から第六腰 髄:L4-L6)も採取した。採取した組織は、パラホルムアルデヒド固定液でさらに一晩固定し た。組織を水洗後、それぞれ 70%、80%、95%及び 100%のエタノール中で組織を浸漬するこ とで脱水させた。その後、キシレン及びパラフィンに浸漬し、パラフィンブロックを作製し た。このブロックをミクロトームで薄切することで、5 μm の薄切切片を作製した。 1.1.7 ヘマトキシリン・エオジン染色 薄切切片をエタノールに浸漬し、脱パラフィン処理を行った。その後、切片を流水中で水 洗し、ヘマトキシリン及びエオジン染色液の順に浸漬し、ヘマトキシリン・エオジン(HE) 染色を行った。染色した切片はエタノールで脱水後、キシレンで透徹し、カバーガラスで封 入した。 1.1.8 免疫染色 薄切切片をエタノールに浸漬し、脱パラフィン処理を行った。その後、切片を流水中で水 洗した。抗 Iba1 抗体を用いた染色は、抗原賦活化のため、10 mmol/L クエン酸緩衝液(pH 6.0) で加熱処理(98℃、20 分間)を行った。10 mmol/L PBS(pH7.4)で洗浄後、内因性ペルオキ シダーゼを不活性化させるため、3%過酸化水素水で 5 分間処置した。
Dako Antibody Diluent(Agilent Technologies、Santa Clara、CA、USA)に溶解させた一次抗 体(下記)を室温で 50 分間反応させた。続いて、二次抗体(下記)を室温で 30 分間反応さ せた。切片を PBS で洗浄し、DAB(3,3'-diaminobenzidine、ニチレイバイオサイエンス、東京) で発色させた。切片を水洗後、ヘマトキシリン液で核染色を行った。染色された切片は、エ タノールで脱水後、キシレンで透徹し、カバーガラスで封入した。 一次抗体 抗 GFAP ウサギポリクローナル抗体(1.5 倍希釈、ニチレイバイオサイエンス) 抗 Iba1 ウサギポリクローナル抗体(2000 倍希釈、和光純薬工業、大阪) 二次抗体 ヒストファイン シンプルステインラット MAX-PO(MULTI)(希釈なし、ニチレイバイ オサイエンス)
1.1.9 Total RNA の抽出及び逆転写反応による cDNA の合成
11
央の皮膚を切開して坐骨神経束を採取し、ISOGEN®
(ニッポンジーン、東京)を満たした 1.5 mL チューブに浸漬させた。組織が小さいため、2 匹分の組織を 1 つのチューブに入れ 1 サン プルとした。ISOGEN を用い、total RNA を抽出後、260 nm の吸光度の値から RNA 濃度を算 出した。
各サンプル当たりの total RNA 濃度が 12 μL 当たり 1 μg となるよう RNase-free water で希釈 した。Omniscript RT kit(QIAGEN、Valencia、CA、USA)、Random Primer(最終濃度 1 μmol/L、 タカラバイオ、滋賀)及び RNase OUT Recombinant Ribonuclease Inhibitor(QIAGEN)を混合、 PTC-100® Thermal Cycler(Bio-Rad Laboratories, Inc.、Hercules、CA、USA)を用い、37℃で 60 分、93℃で 5 分の条件にて転写反応を行い、cDNA を合成した。
1.1.10 Real-time PCR による発現解析
各サンプル当たりの cDNA 量が、14.4 μL 当たり 500 ng となるよう RNase-free water で希釈 した。希釈した cDNA、SYBR Green®
(Applied Biosystems、Waltham、MA、USA)及び Forward / Reverse primer(最終濃度各 100 nmol/L、Table 1)を混合した。リアルタイム PCR システム (Applied Biosystems 7500)を用い、以下の条件にて real-time PCR を行い、Ct 値を算出した。
Step 1 50.0℃、2 min、1 回 Step 2 95.0℃、10 min、1 回
Step 3 95.0℃、15 sec 及び 60.0℃、1 min のサイクルを 40 回繰り返す Step 4 95.0℃、15 sec、60.0℃、1 min、95.0℃、15 sec、60.0℃、1 min、1 回
各サンプルの標的遺伝子の Ct 値は内在性遺伝子(GAPDH、TaqMan®
Rodent GAPDH Control Reagents、Thermo Fisher Sci. Inc. Waltham、MA、USA)の Ct 値で補正し相対値を算出した。 具体的には標的遺伝子の Ct 値から内在性遺伝子の Ct 値を減じて⊿Ct 値を算出、2 を底とし て-⊿Ct 値の累乗を算出した。
Table 1 The primer sets for PCR.
Genes Forward primer Reverse primer
TNF-α AAGCCTGTAGCCCACGTCGTA GGCACCACTAGTTGGTTGTCTTTG
IL-1β TGTGAAATGCCACCTTTTGA AGCTTCTCCACAGCCACAAT
MCP-1 AACTGCATCTGCCCTAAGGTCT GGAAAAGGTAGTGGATGCATTAGC
12
1.1.11 統計処理
13
1.2 結果
1.2.1 PSL マウスの疼痛閾値の経時推移
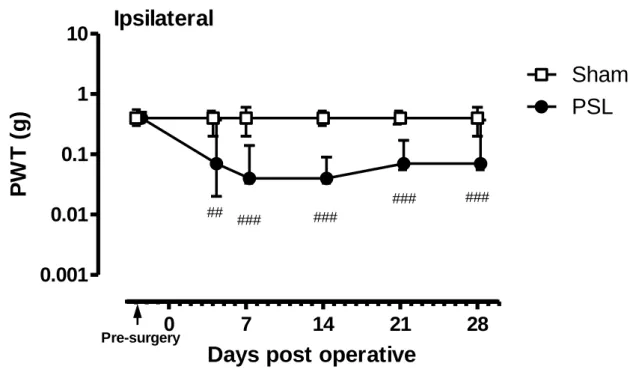
PSL マウス及び Sham マウスの疼痛閾値の経時変化を Figure 6 及び Figure 7 に示す。Figure 6 には神経結紮側の足(ipsilateral)の疼痛閾値を示す。PSL マウスの疼痛閾値は、結紮後 4 日 目以降低下し、その後 28 日目まで低値の状態が維持された。つまり、結紮前は約 0.4 g であ ったのに対し、結紮後 4 日目では約 0.07 g(P<0.01)に、結紮後 7 日目では約 0.04 g(P<0.001) に低下した。その一方、Sham マウスの疼痛閾値は手術前後で変わらず約 0.4 g のままであっ た。このように PSL マウスは疼痛閾値が低下し、正常動物では逃避行動を示さない弱い VF フィラメント刺激でも逃避行動を示すようになったことから、神経障害性疼痛を発症してい ることが示された。
Figure 6 Changes in the PWT on the ipsilateral paws of mice with PSL.
Graph shows temporal changes in the PWTs on the ipsilateral paws of mice with PSL (n=19) and sham (n=10). Data are expressed as the median force (g) required to induce paw withdrawal, and vertical bars represent 1st and 3rd quartiles. P<0.01 and P<0.001 vs. before surgery (Dunnett’s test).
0
7
14
21
28
0.001
0.01
0.1
1
10
Sham
PSL
Days post operative
14
Figure 7 に神経を結紮しなかった側の足(contralateral)の疼痛閾値を示す。PSL マウスの疼 痛閾値は、手術前後で変わらず約 0.16 g から約 0.4 g となった。なお、Sham マウスの疼痛閾 値も、偽手術前後で変化しなかった。
Figure 7 Changes in the PWT on the contralateral paws of mice with PSL.
Graph shows temporal changes in the PWTs on the contralateral paws of mice with PSL (n=19) and sham (n=10). Data are expressed as the median force (g) required to induce paw withdrawal, and vertical bars represent 1st and 3rd quartiles.
0
7
14
21
28
0.001
0.01
0.1
1
10
Sham
PSL
Days post operative
15
1.2.2 PSL マウスの坐骨神経及び脊髄の組織学的変化
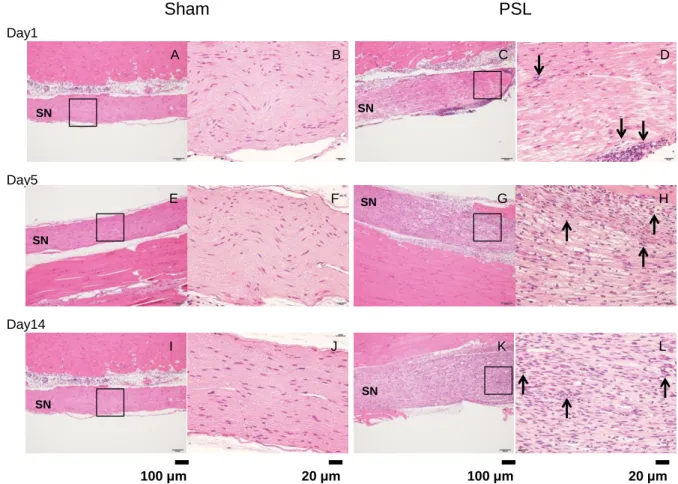
神経障害性疼痛モデルの経時的な組織学的変化を把握するため、神経結紮後 1(Day1)、5 (Day5)及び 14 日目(Day14)に、坐骨神経及び腰部脊髄を採取した。PSL マウス及び Sham マウスの坐骨神経の HE 染色像を Figure 8 に示す。Sham マウスと比較して、PSL マウスの坐 骨神経では、神経結紮後 1、5 及び 14 日目で、ヘマトキシリン濃染の核を有する小円形から 類円形の細胞の増加、すなわち、好中球及びリンパ球等の細胞の集簇が視認された(↑で示 す)。
Figure 8 HE staining of the sciatic nerve in sham and PSL mice.
16
PSL あるいは Sham マウスの術後の坐骨神経へのマクロファージの浸潤を観察するため に、マクロファージマーカーIba1 の免疫染色を行った。染色像を Figure 9 に示す。PSL マウ スの坐骨神経では、Sham マウスのそれと比較して、神経結紮後 1、5 及び 14 日目で、Iba1 陽 性細胞数の増加が視認された(↑で示す)。
Figure 9 Iba1 immunohistochemistry of the sciatic nerve in sham and PSL mice.
17 PSL あるいは Sham マウスの術後の脊髄後角でのアストロサイトの活性化を観察するた めに、アストロサイトマーカーGFAP の免疫染色を行った。染色像を Figure 10 に示す。PSL マウスの脊髄後角では、神経結紮後 1、5 及び 14 日目で、GFAP 陽性細胞が強陽性を示した (↑で示す)。また、Sham マウスの脊髄後角でも、偽手術後 1 日目で GFAP 陽性細胞が強陽 性を示したが、5 及び 14 日目では示さなかった。
Figure 10 GFAP immunohistochemistry of the lumbar spinal cord in sham and PSL mice. The spinal cords of PSL and sham mice were harvested at 1, 5 and 14 after surgery (Day1, 5 and 14). GFAP, an astrocyte maker, staining was performed to examine pathological changes after PSL (Day1: A-D, Day5: E-H and Day14: I-L). Right side photo indicates higher magnification of the area
surrounded with a box in the left side photo. Ipsilateral means ipsilateral side of PSL, and contralateral means contralateral side of PSL. Allows indicate GFAP positive cells.
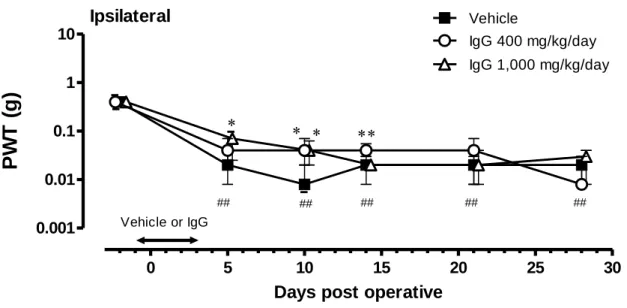
18 1.2.3 PSL マウスの神経障害性疼痛に対する免疫グロブリン製剤前投与の効果 神経障害性疼痛の発症に対する炎症反応の関与を調べるため、免疫グロブリン製剤(400 あるいは 1,000 mg/kg/day)を神経結紮前日からその 3 日後まで、1 日 1 回腹腔内投与した。 疼痛閾値の変化を Figure 11 及び Figure 12 に示す。 神経結紮側の足(Ipsilateral)の疼痛閾値は、結紮前(免疫グロブリン製剤投与前)の値よ りも、結紮後 5 日目から低値を示した。つまり、結紮前の疼痛閾値は約 0.4 g であったのに対 し、結紮後 5 日目で約 0.02 g、結紮後 10 日目では約 0.008gとなり(それぞれ P<0.01)、弱 い VF フィラメント刺激でも痛みを感じるようになったことが示された。
Figure 11 Effects of IgG on the development of neuropathic pain in mice with PSL.
Vehicle or IgG (400 or 1,000 mg/kg/day) was intraperitoneally administered to mice with PSL from Day-1 to Day3. Data are expressed as the median force (g) required to induce paw withdrawal, and vertical bars represent 1st and 3rd quartiles (n=14-15/group). P<0.05, P<0.01 vs. vehicle, and
P<0.01 vs. before surgery (Dunnett’s test). 0 5 10 15 20 25 30 0.001 0.01 0.1 1 10 IgG 400 mg/kg/day Vehicle ## ## ## ## ## IgG 1,000 mg/kg/day Ipsilateral
Days post operative
19
免疫グロブリン投与群の動物の疼痛閾値は、溶媒対照群のそれよりも結紮後 5(Day5)、 10(Day10)及び 14 日目(Day14)で高値となった。溶媒対照群、免疫グロブリン 400 mg/kg/day 群及び 1,000 mg/kg/day 群の疼痛閾値は、結紮後 5 日目では約 0.02 g、約 0.04 g(P>0.05)及び 0.07 g(P<0.05)、結紮後 10 日目では約 0.008 g、約 0.04 g(P<0.05)及び約 0.04 g(P<0.05)、 結紮後 14 日目では約 0.02 g、約 0.04 g(P<0.01)及び約 0.02 g(P>0.05)であった。これらの 結果から免疫グロブリン製剤の前投与は、PSL マウスの疼痛閾値の低下を軽減することが示 された。
Figure 12 Pre-treatment of IgG attenuated development of neuropathic pain in mice with PSL. In order to make Figure 11 clear, the PWTs were plotted Day5, 10 and 14, respectively. Bars represent median, and vertical bars represent 1st and 3rd quartiles (n=14-15). P<0.05, P<0.01 vs. vehicle, and P<0.01 vs. before surgery (Dunnett’s test).
20 1.2.4 PSL マウスの神経障害性疼痛に対する免疫グロブリン製剤後投与の効果 神経障害性疼痛を発症した状態での炎症反応の関与を調べるため、免疫グロブリン製剤 (400 あるいは 1,000 mg/kg/day)を、神経障害性疼痛発症後の結紮後 11 日から 15 日目まで、 1 日 1 回腹腔内投与した。疼痛閾値の変化を Figure 13 及び Figure 14 に示す。 神経を結紮した側の足では、結紮前の疼痛閾値は約 0.4 g であった。結紮後 11 日目(免疫 グロブリン製剤投与前)では約 0.008 g(P<0.01)となり、神経障害性疼痛の発症が確認され た。
Figure 13 Effects of IgG on neuropathic pain in mice with PSL.
Vehicle or IgG (400 or 1,000 mg/kg/day) was intraperitoneally administered to mice with PSL from Day11 to Day15. Data are expressed as the median force (g) required to induce paw withdrawal, and vertical bars represent 1st and 3rd quartiles (n=12/group). P<0.05 and P<0.01 vs. before surgery (Dunnett’s test). 0 5 10 15 20 25 30 35 0.001 0.01 0.1 1 10 IgG 1,000 mg/kg/day Vehicle IgG 400 mg/kg/day Ipsilateral ##
Days post operative
21 本実験条件での免疫グロブリン製剤投与後の疼痛閾値は、いずれの測定時点でも変化しな かった。溶媒対照群、免疫グロブリン 400 mg/kg/day 群及び 1,000 mg/kg/day 群の疼痛閾値は、 結紮後 18 日目(Day18)で約 0.008 g、約 0.014 g 及び約 0.008 g、結紮後 24 日目(Day24)で 約 0.02 g、約 0.014 g 及び 0.02 g、結紮後 30 日目(Day30)で約 0.04 g、約 0.03 g 及び 0.03 g、 結紮後 36 日目(Day36)で約 0.02 g、約 0.04 g 及び約 0.04 g であった(いずれも P>0.05)。 これらの結果から後投与された免疫グロブリン製剤は、PSL マウスの神経障害性疼痛を改善 しないことが示された。
Figure 14 Post-treatment of IgG failed to attenuate the maintenance of neuropathic pain in mice with PSL.
In order to make Figure 13 clear, the PWTs were plotted Day18, 24, 30 and 36, respectively. Bars represent median, and vertical bars represent 1st and 3rd quartiles (n=12).
22 1.2.5 PSL 後 1 日目の坐骨神経での炎症性サイトカイン遺伝子発現に対する免疫グロブ リン製剤の効果 免疫グロブリン製剤前投与による神経障害性疼痛軽減作用の機序を調べるため、坐骨神経 での炎症性サイトカイン遺伝子発現を調べた。免疫グロブリン製剤(1,000 mg/kg/day)を神 経結紮前日からその 1 日後まで、1 日 1 回腹腔内投与したときの坐骨神経束での炎症性サイ トカイン mRNA 発現レベルの変化を Figure 15 に示す。
PSL/溶媒対照群の動物の TNF-α、IL-1β、MIP-1α 及び MCP-1 の mRNA 発現レベルは、sham/ 溶媒対照群のそれよりも高値となった(TNF-α:P<0.01、IL-1β、MIP-1α 及び MCP-1:P<0.001)。 一方、PSL/免疫グロブリン群では、TNF-α 及び MCP-1 の mRNA 発現レベルの上昇が一部抑 制された(P<0.05)。IL-1β 及び MIP-1α の mRNA 発現レベルに、免疫グロブリン製剤の前投 与による影響は認められなかった。
Figure 15 Effects of IgG on mRNA expression of the inflammatory cytokines on Day1 in mice with PSL.
Vehicle or IgG (1,000 mg/kg/day) was intraperitoneally administered to mice with PSL from Day-1 to Day1. The mice were sacrificed on Day1, and total RNA was isolated from sciatic nerves. The mRNA expression levels of TNF-α, IL-1β, MIP-1α and MCP-1 were measured using qRT-PCR. The amount of the target mRNA relative to the expression of GAPDH mRNA is expressed as the mean±SEM (n=10). P<0.05, P<0.01 and P<0.001 (Tukey’s test).
23 1.2.6 PSL 後 5 日目の坐骨神経での炎症性サイトカイン遺伝子発現に対する免疫グロブ リン製剤の効果 免疫グロブリン製剤(1,000 mg/kg/day)を神経結紮前日からその 3 日後まで、1 日 1 回腹腔 内投与し、結紮後 5 日目の坐骨神経束での炎症性サイトカイン mRNA 発現レベルを調べた。 結果を Figure 16 に示す。 PSL/溶媒対照群の動物の TNF-α 及び MCP-1 の mRNA 発現レベルは、sham/溶媒対照群の動 物のそれらよりも高値であった(TNF-α:P<0.05、MCP-1:P<0.001)。一方、PSL/免疫グロ ブリン群では、MCP-1 の mRNA 発現レベルの上昇が抑制される傾向にあった(P=0.0542)。 TNF-α の mRNA 発現レベルに及ぼす免疫グロブリン製剤投与の影響は観察されなかった。な
お、IL-1β 及び MIP-1α の mRNA 発現レベルに、坐骨神経結紮による影響は認められなかった。
Figure 16 Effects of IgG on mRNA expression of the inflammatory cytokines on Day5 in mice with PSL.
Vehicle or IgG (1,000 mg/kg/day) was intraperitoneally administered to mice with PSL from Day-1 to Day3. The mice were sacrificed on Day5, and total RNA was isolated from sciatic nerves. The mRNA expression levels of TNF-α, IL-1β, MIP-1α and MCP-1 were measured using qRT-PCR. The amount of the target mRNA relative to the expression of GAPDH mRNA is expressed as the mean±SEM (n=10). P<0.05 and P<0.001 (Tukey’s test).
26 第二章 神経障害性疼痛への TRPV1 チャネルの関与
第一章で、神経障害性疼痛の病態では、末梢神経及び脊髄で炎症反応が関与していると考 えられることを示した。この前章の結果から、神経障害性疼痛への障害局所の炎症反応の寄 与は、疼痛の発症期で高く、かつ維持期では低くなると考えられた。
27 2.1 実験材料・方法 本実験計画は、ファイザー株式会社 名古屋研究所及び帝人ファーマ株式会社 生物医学総 合研究所の動物実験委員会にて審査された。その後、承認されたプロトコールに基づいて本 研究が実施された。 2.1.1 実験動物 雄性 SD 系ラット(251-293 g、日本チャールスリバー)及び雄性 ddy 系マウス(第一章と 同じ)を用いた。ラットは、実験に使用するまで明暗サイクル 12 時間(明期 7:00-19:00)、 室温 23±2℃、湿度 55±15%の一定環境下で動物を飼育した。飼養中の動物には水及び固型飼 料を自由に摂取させた。 雄性 ddy 系マウスは、第一章と同様の環境下で飼育した。 2.1.2 被験薬 選択的 TRPV1 チャネル遮断薬の BCTC(N-(4-tertiarybutylphenyl)-4-(3-chloropyridin-2-yl) tetrahydropyrazine-1(2H)-carbox-amide)はファイザー株式会社 名古屋研究所で合成あるいは ENZO® Life Science(Farmingdale、NY、USA)から購入した。カプサイシン誘導体である resiniferatoxin(RTX)は Sigma(St Louis、MO、USA)から購入した。BCTC は、0.5%メチル セルロース溶液/Tween80(95:5 v/v)に懸濁させ経口投与した。Resiniferatoxin は、エタノー ル/Tween80/生理食塩液(10/10/80 v/v/v)に溶解させ皮下投与した。
2.1.3 神経障害性疼痛モデルの作製
2.1.3.1 坐骨神経絞扼モデルの作製
神経障害性疼痛モデルでの TRPV1 チャネルの分布を調べるため、ラットで文献報告の多い 坐骨神経絞扼モデル(chronic constriction injury、CCI モデル)を作製した43)。ラットを、ペ ントバルビタールナトリウム(60 mg/kg)腹腔内投与で麻酔した。左側大腿部中央の皮膚を 切開し、坐骨神経束を露出させた。4-0 絹糸を用いて、当該神経束を 4 回結紮し、その後皮膚 を縫合した。偽手術(sham)は坐骨神経剥離まで行い、神経束を結紮せずに皮膚を縫合した。 2.1.3.2 PSL モデルの作製 第一章と同様に、マウスを用いて作製した。 2.1.4 疼痛行動の評価
CCI ラットの疼痛行動の評価は、Field らの方法44)に従って実施した(Figure 17)。ラット
28
行動が認められた最小の VF フィラメント刺激を、そのラットの疼痛閾値(paw withdrawal threshold、PWT)とした。疼痛閾値が 2 g 以下のラットを神経障害性疼痛発症と判定し、本実 験で疼痛モデルとして使用した。
PSL マウスの疼痛行動の評価は、第一章と同様に行った。
Figure 17 Schematic of measurement of paw withdrawal threshold (PWT) using von Frey (VF) filaments in rats. 2.1.5 免疫染色用組織標本の作製 CCI ラットをペントバルビタールナトリウム(70-80 mg/kg)腹腔内投与により麻酔し、200 mL の 100 mmol/L PBS(pH7.4)で経心的に灌流することにより、組織内の血液を除去した。 続いて、300 mL の 4%パラホルムアルデヒド含有 PBS で灌流固定した。灌流固定後、腰部脊 椎を切り出し、第四及び第五腰椎部(L4 及び L5)の後根神経節(DRG)を単離し、4%パラ ホルムアルデヒド含有 PBS で一晩固定した。次に、10、15 及び 20%のスクロース含有 PBS の順に、組織を浸漬させた。水溶性包埋剤の OCT compound を用いて、液体窒素下で組織ブ ロックを凍結させ、凍結ブロックを作製した。この凍結ブロックからクリオスタットを用い て、8 μm の薄切切片を作製した。 2.1.6 免疫染色
一次抗体の非特異的反応を抑制するために、10% normal goat serum 及び 0.3% Triton X-100 を含む CAS BLOCK(ZYMED Laboratories、South San Francisco、CA、USA)(以下、blocking buffer)で 2 時間処置した。その後、免疫蛍光染色を行った。下に示す一次抗体を blocking buffer で希釈し、それぞれ室温で一晩反応させた。反応後、切片を PBS で洗浄した。その後、blocking buffer で希釈した二次抗体を、室温で 2 時間反応させた。切片を PBS で洗浄後、カバーガラ スで封入した。組織切片の観察は共焦点レーザー顕微鏡( LSM5 PASCAL; CarlZeiss 、 Őberkochen、Germany)を用いて行った。
一次抗体
29 二次抗体
rhodamine red-X conjugated donkey anti-goat immunoglobulin G ( 3 μg/mL; Jackson ImmunoResearch Lab. Inc.、West Grove、PA、USA)
30
Figure 18 Experimental protocols of TRPV1 ligands treatment for the neuropathic pain. CCI; chronic constriction injury of sciatic nerve, PSL; partial sciatic nerve ligation, RTX; resiniferatoxin, PWT; paw withdrawal threshold, p.o.; per os, s.c.; subcutaneous injection
2.1.9 統計処理
疼痛閾値は各群の中央値で示した。また、25 及び 75%反応値を算出した。神経障害性疼痛 の発症確認のため、Sham/溶媒対照群と CCI/溶媒対照群あるいは PSL/溶媒対照群の比較を、 Mann-Whitney’s U-test(Figure 20 及び Figure 22)あるいは Wilcoxon’s test(Figure 24)で行っ た。BCTC の評価は、溶媒対照群と BCTC 投与群で、Dunn’s test(Figure 20 及び Figure 21) あるいはDunnett’s test(Figure 22 及び Figure 23)で行った。Resiniferatoxin の評価は、溶媒対 照群と resiniferatoxin 群で、Mann-Whitney’s U-test で行った(Figure 24 及び Figure 25)。いず れの検定も有意水準は 5%とした。 1 h 0 CCI or PSL or sham 4 h 2 h BCTC, p.o.
Protocol 6: Effects of BCTC on neuropathic pain in rats with CCI or mice with PSL.
PWT measurement 2 weeks
Day 1
CCI or sham RTX, s.c. Day 4 Day 7
PWT measurement 2 weeks
Protocol 7: Effects of RTX on neuropathic pain in rats with CCI.
CCI 2 weeks
Protocol 5: Histopathological examination of dorsal root ganglion neurons in rats with CCI.
31
2.2 結果
2.2.1 CCI ラットの L4-L5 後根神経節での TRPV1 チャネルの分布
TRPV1 チャネルの組織学的解析結果を Figure 19 及び Table 2 に示す。CCI ラットの後肢を
32
Figure 19 Double-labeling immunohistochemistry of TRPV1 and NF200 in L4-L5 DRG neurons from rats with CCI.
The L4-L5 DRG neurons were obtained from CCI rats at 2 weeks after surgery. (A)
NF200-immunoreactive neurons (green). (B) TRPV1-immunoreactive neurons (red). (C) Merged image: double-labeled neurons (arrowheads) appear as yellow staining. Scale bar: 50 μm.
Table 2 The number of DRG neurons that express TRPV1 and/or NF200 in rats with CCI.
(A) Positive for TRPV1 only, (B) TRPV1 and NF200 double-labeled, (C) Positive for NF200 only.
33
2.2.2 CCI ラットの神経障害性疼痛に対する BCTC の効果
神経障害性疼痛への TRPV1 チャネルの関与を調べるため、選択的 TRPV1 チャネル遮断薬 の BCTC を CCI ラットに投与し、疼痛閾値の変化を調べた。結果を Figure 20 及び Figure 21 に示す。BCTC 投与前の CCI ラット及び Sham ラットの疼痛閾値は、それぞれ約 0.6 g 及び 約 16 g であった。Sham ラットの疼痛閾値よりも CCI ラットのそれは低く(P<0.01)、CCI ラットでも PSL マウスと同様に神経障害性疼痛の発症が示された。
Figure 20 Effects of BCTC on neuropathic pain in rats with CCI.
Baseline PWT to VF filaments were determined before drugs administration, and PWTs were reassessed at each time points. Results are expressed as the median force (g) required to induce paw withdrawal, and vertical bars represent 1st and 3rd quartiles (n=8/group). P<0.01 vs. CCI/vehicle group (Mann-Whitney’s U-test). P<0.05, P<0.01 vs. CCI/vehicle group (Dunn’s test).
34 BCTC 経口投与後(10 あるいは 100 mg/kg)、CCI ラットの疼痛閾値は、投与 1 時間後に 溶媒対照群、BCTC 10 及び 100 mg/kg 投与群で、それぞれ約 0.6 g、約 1.4 g(P<0.05)及び 約 2 g(P<0.01)になった。さらに投与 2 時間後では、それぞれ約 0.6 g、約 1 g(P>0.05)及 び約 2 g(P<0.01)になった。これらの結果から、BCTC は CCI ラットの神経障害性疼痛を 軽減することが示された。
Figure 21 BCTC attenuated the maintenance of neuropathic pain in rats with CCI.
In order to make Figure 20 clear, the PWTs (g) were plotted before, 1, 2 and 4 hour after BCTC administration. Bars represent median, and vertical bars represent 1st and 3rd quartiles (n=8).
P<0.05, P<0.01 vs. CCI/vehicle group (Dunn’s test).
35 2.2.3 PSL マウスの神経障害性疼痛に対する BCTC の効果 PSL マウスの神経障害性疼痛に対する TRPV1 チャネル遮断薬 BCTC の効果を Figure 22 及 び Figure 23 に示す。BCTC 投与前の PSL マウス及び Sham マウスの疼痛閾値は、それぞれ約 0.008 g 及び約 0.4 g であった。Sham マウスの疼痛閾値よりも PSL マウスのそれは低値であ った(P<0.001)。
Figure 22 Effects of BCTC on neuropathic pain in mice with PSL.
Baseline PWT to VF filaments were determined before drugs administration, and PWTs were reassessed at each time points. Results are expressed as the median force (g) required to induce paw withdrawal, and vertical bars represent 1st and 3rd quartiles (n=10/group). P<0.001 vs.
36
BCTC 経口投与後(10、30 あるいは 100 mg/kg)、PSL マウスの疼痛閾値は、投与 1 時間後 に、溶媒対照群、BCTC 10、30 及び 100 mg/kg 投与群で、それぞれ約 0.02 g、約 0.014 g(P>0.05)、 約 0.02 g(P>0.05)及び約 0.07 g(P<0.05)となった。これらの結果から、BCTC は PSL マウ スでも CCI ラットと同様に神経障害性疼痛を軽減することが示された。
Figure 23 BCTC attenuated the maintenance of neuropathic pain in mice with PSL. In order to make Figure 22 clear, the PWTs (g) were plotted before, 1, 2 and 4 hour after BCTC administration. Bars represent median, and vertical bars represent 1st and 3rd quartiles (n=10).
P<0.05 vs. PSL/vehicle group (Dunnett’s test).
37
2.2.4 CCI ラットの神経障害性疼痛に対する resiniferatoxin の効果
神経障害性疼痛への TRPV1 チャネルの関与を調べるため、神経障害性疼痛を発症した CCI ラットに、カプサイシン誘導体 resiniferatoxin(RTX)0.3 mg/kg を皮下投与し、投与後 1、4 及び 7 日目に疼痛閾値を測定した。結果を Figure 24 及び Figure 25 に示す。
Figure 24 Effects of resiniferatoxin on neuropathic pain in rats with CCI.
38
CCI ラットの疼痛閾値は、投与前(Day 0)のそれと比べ、投与後 1 日目(Day 1)から 4 日目(Day 4)は変化しなかった。一方、投与後 7 日目(Day 7)の CCI ラットの疼痛閾値は、 溶媒対照群及び resiniferatoxin 投与群で、それぞれ約 0.6 g 及び約 4 g(P<0.05)となり、 resiniferatoxin 投与群で高値を示した。Sham ラットに resiniferatoxin を投与しても、投与後 1 日目から 7 日目で、疼痛閾値は変化しなかった。これらの結果から、resiniferatoxin は BCTC と同様に CCI ラットの神経障害性疼痛を軽減することが示された。
Figure 25 Resiniferatoxin attenuated the maintenance of neuropathic pain in rats with CCI. In order to make Figure 24 clear, the PWTs (g) were plotted before (Day0), 1, 4 and 7 days after resiniferatoxin (RTX) injection. Bars represent median, and vertical bars represent 1st and 3rd quartiles (n=10). P<0.05 vs. PSL/vehicle group (Mann Whitney’s U-test).
39 2.3 考察 正常ラットの後根神経節細胞を用いた検討で、TRPV1 チャネルは主に無髄 C 及び有髄 Aδ 線維に発現していることが報告されている47, 48)。本実験では、神経障害性疼痛モデルの後根 神経節での TRPV1 チャネルの分布を観察した。免疫染色の結果、TRPV1 チャネルは主に NF200 陰性の小型及び中型の神経細胞に発現していた。様々な研究報告で、NF200 は後根神 経節細胞の A 線維マーカーと見なされている49-52) 。Lawson 及び Waddell は、抗リン酸化 NF200 抗体で染色した後根神経節細胞を神経伝導速度から分類し、NF200 陽性及び陰性細胞が、そ れぞれ有髄 A 線維及び無髄 C 線維となることを報告した53)。本実験結果は、正常ラットを用 いたこれら報告と一致したことから、TPRV1 チャネルは、主に無髄 C 線維に発現すると考え られた。この無髄 C 線維は痛覚伝達を担うことから、TPRV1 チャネルは神経障害性疼痛の伝 達に関与すると考えられた。 BCTC は、高い親和性を有する TRPV1 チャネル選択的遮断薬である54)。本研究で、BCTC を CCI ラットあるいは PSL マウスに投与したところ、それらの神経障害性疼痛の症状が軽減 された。続いて神経障害性疼痛での TRPV1 チャネルの関与についてさらに検討するため、カ プサイシン誘導体の resiniferatoxin を用いた。Resiniferatoxin はカプサイシンよりも強力かつ 持続的な TRPV1 チャネル活性化作用を有する55) 。この TRPV1 チャネルの持続的な活性化は、 一過性に感覚神経を興奮させるものの、その後 TRPV1 チャネルによる一次感覚神経の伝達を 持続的に低下させる55)。この現象は、一次感覚神経の脱感作と考えられている55) 。VF フィラ メント刺激に対する疼痛閾値の測定で、BCTC 投与の結果と同じく、投与 7 日目の CCI ラッ トの疼痛閾値の低下すなわち痛覚過敏が軽減された。これらの結果から、TRPV1 チャネルは、 神経障害性疼痛の維持に関与することが示唆された。 末梢神経の障害局所で産生、遊離された炎症性メディエーター及び炎症性サイトカインは、 一次感覚神経を直接興奮させて疼痛を惹起させるとともに、細胞内シグナル伝達経路を介し て TRPV1 チャネルの機能変化を引き起こす56)。炎症性メディエーターは、その受容体を介し
42 3.1 実験材料・方法 本実験計画はファイザー株式会社 名古屋研究所の動物実験委員会にて審査された。その 後、承認されたプロトコールに基づいて本研究が実施された。 3.1.1 実験動物 雄性 ddy マウスは、第一章と同様の動物を使用した。カンナビノイド CB2受容体 knockout (KO)マウスは、C57BL/6 系統を用い、Buckley らの方法83) に従って作製した。 3.1.2 被験薬 カンナビノイド CB2受容体刺激薬の JWH133 ((6aR,10aR)-3-(1,1-Dimethylbutyl)-6a,7,10,10a-tetrahydro-6,6,9-trimethyl-6H-dibenzo[b,d]pyran) は Sigma より購入した。JWH133 は DMSO に溶解させた。 3.1.3 神経障害性疼痛モデルの作製 第一章と同様に、マウスを用いて PSL モデルを作製した。 3.1.4 疼痛行動の評価 第一章と同様に実施した。 3.1.5 脊髄くも膜下腔内投与 脊髄カンナビノイド CB2受容体の神経障害性疼痛に対する効果を調べるため、被験薬を脊 髄くも膜下腔内に投与した。被験薬の脊髄くも膜下腔内投与は、Hylden 及び Wilcox らの方法 84)に従って実施した。イソフルラン麻酔下、マウスを腹臥位とし、背部をバリカンで剃毛し た。腰椎部を指で保定し、第 5 腰椎と第 6 腰椎の間にマイクロシリンジの針を挿入した。被 験薬を、5 μL/mouse の容量で、約 1 分間かけて投与した。 3.1.6 Total RNA の調製及び発現ベクターの作製
RNeasy® Mini kit(QIAGEN)を用いてマウス脳より total RNA を取得した。Oligo(dt)12-18 primer(Invitrogen、Carlsbad、CA、USA)を用いて poly(A)+ RNA を調製、SuperScript First Strand
Synthesis(Invitrogen)を用いて逆転写し、cDNA を合成した。カンナビノイド CB1及び CB2
受容体の cDNA は、Taq DNA polymerase 及び下記プライマー(Table 3)を用いて PCR で増幅 した(PCR の条件は以下の通り)。核酸の塩基配列を確認するため、PCR 産物は、TOPO™ TA cloning kit(Invitrogen)を用いて、pcDNA2.1-TOPO ベクターに挿入した。塩基配列の確認は
東洋紡ジーンアナリシスにて実施した。また、カンナビノイド CB1及び CB2受容体 cDNA を
pcDNA3.1(+) ベクターに組み入れ、発現用ベクターを作 製した(mCB1/pcDNA3.1(+) 及び
43 PCR 条件
(mCB1)
Step 1 96.0℃、5 min、1 回
Step 2 96.0℃、30 sec、60.0℃、30 sec 及び 72.0℃、1 min のサイクルを 30 回繰り 返す
(mCB2)
Step 1 94.0℃、2 min、1 回
Step 2 94.0℃、15 sec、53.0℃、30 sec 及び 68.0℃、2 min のサイクルを 33 回繰り 返す
Table 3 The primer sets for PCR.
Genes Sense primer Antisense primer
CB1 5'-ACCATGAAGTCGATCTTAGACGG C-3' 5'-TCACAGAGCCTCGGCAGA-3' CB2 5'-GCCACCATGGAGGGATGCCGGG AGAC-3' 5'-TCCAGCCAACCAGCATTCC-3' 3.1.7 細胞培養及びトランスフェクション
チャイニーズハムスター卵巣(CHO)細胞を Ham's F12 medium(10 % FBS、2 mmol/L% HEPES、100 units/mL penicillin 及び 100 μg/mL streptomycin 含有)で培養した。FuGENE6(Roche Applied Science 、 Indianapolis 、 IN 、 USA ) を 用 い て 、 そ れ ぞ れ マ ウ ス カ ン ナ ビ ノ イ ド CB1/pcDNA3.1(+)及び CB2/pcDNA3.1(+)を CHO 細胞にトランスフェクションし、カンナビノ イド CB1及び CB2発現細胞を調製した。
3.1.8 cAMP 評価
96 ウェルプレートに、カンナビノイド CB1あるいは CB2受容体発現 CHO 細胞を、それぞ
れ 5x103
cells の濃度で播種した。次に、被験物質を溶解させたアッセイ buffer(20 mM HEPES、
44 3.1.9 実験プロトコール 本章での実験プロトコールを Figure 26 に示す。 Protocol 8: 脊髄カンナビノイド CB2受容体の関与を調べるため、神経結紮 2 週間後に神経 障害性疼痛を発症した PSL マウスに JWH133 を脊髄くも膜下腔内投与し、その 0.5、1 及び 2 時間後に疼痛閾値を測定した。 Protocol 9: カンナビノイド CB2受容体 KO マウス及び wild-type マウスを用いた検討では、 神経結紮 2 週間後に神経障害性疼痛を発症した PSL マウスに JWH133 を脊髄くも膜下腔内投 与し、その 0.5、1 及び 2 時間後に疼痛閾値を測定した。
Figure 26 Experimental protocols of CB2 agonist treatment for the neuropathic pain.
PSL; partial sciatic nerve ligation, PWT; paw withdrawal threshold, i.t.; intrathecal injection, CB2 KO; cannabinoid CB2 receptor knockout
3.1.10 統計処理
疼痛閾値は各群の中央値、25 及び 75%反応値を算出した。神経障害性疼痛の発症確認のた め、Sham/溶媒対照群と PSL/溶媒対照群(Figure 28)あるいはモデル作製前とモデル作製後 (Figure 29)の比較を、Mann-Whitney’s U-test で行った。JWH133 の評価は、溶媒対照群と JWH133 投与群で、Dunn’s test(Figure 28)、あるいは Mann-Whitney’s U-test(Figure 29)で 行った。いずれの検定も有意水準は 5%とした。 0.5 h PSL or sham 2 h 1 h JWH133, i.t.
Protocol 8: Effects of i.t. administration of JWH133 on neuropathic pain in mice with PSL.
PWT measurement 2 weeks
PSL JWH133, i.t.
PWT measurement
2 weeks
Protocol 9: Effects of JWH133 on neuropathic pain cannabinoid CB2receptor knockout mice with PSL.
0.5 h 1 h 2 h CB2KO mice
45 3.2 結果 3.2.1 JWH133 のマウスカンナビノイド CB2及び CB1受容体への刺激作用 JWH133 はヒトのカンナビノイド CB2受容体刺激薬と報告されている 85)。しかしながら、 マウスのカンナビノイド CB2受容体に対し、JWH133 が選択的な刺激作用を発揮するか否か については不明である。カンナビノイド CB2及び CB1受容体は、ともに Gi/oタンパク質共役 型受容体で、受容体刺激により cAMP の産生が抑制される。そこで cAMP 産生能の減弱を指 標に、マウスのカンナビノイド CB2及び CB1受容体に対する JWH133 の刺激作用を評価した (Figure 27)。 JWH133 はマウスカンナビノイド CB2受容体に対して刺激作用を示し、濃度依存的にホル スコリン誘発 cAMP 産生を抑制した。その EC50値は 63 nM、最大抑制率はほぼ 100%に達し た。JWH133 は、マウスカンナビノイド CB1受容体に対しても刺激作用を示し、濃度依存的 にホルスコリン誘発 cAMP 産生を阻害した。その EC50値は 2500 nM、最大抑制率は約 94%で あった。これらの結果から、JWH133 はマウスカンナビノイド CB1受容体よりもカンナビノ イド CB2受容体に対して、約 40 倍の高い親和性を発揮することが示された。
Figure 27 The ability of JWH133 to activate mouse cannabinoid CB2 and CB1 receptors.
46 3.2.2 PSL マウスの神経障害性疼痛に対する JWH133 の効果 JWH133 はマウスカンナビノイド CB2受容体に親和性の高い刺激薬であることが確認され た。JWH133 の PSL マウスの神経障害性疼痛に対する効果を Figure 28 に示す。 神経障害性疼痛を発症した PSL マウスに、JWH133(50 あるいは 100 nmol/mouse)を脊髄 くも膜下腔内投与したところ、PSL マウスの疼痛閾値は、投与後 0.5 時間で、溶媒対照群、 JWH133 50 及び 100 nmol/mouse 群で、それぞれ約 0.04 g、約 0.16 g(P<0.05)及び約 0.16 g(P<0.05) になった。このことから JWH133 は神経障害性疼痛を軽減することが示された。
Figure 28 Effects of i.t. administration of JWH133 on neuropathic pain in mice with PSL. The PWT to VF filaments were determined in sham and PSL mice before, 0.5, 1 and 2 h after administration. For i.t. administration, drugs were administered into the subarachnoid space through the intervertebral foramen between L5 and L6. Data are expressed as the median force (g) required to induce paw withdrawal, and vertical bars represent 1st and 3rd quartiles (n=6-8/group). P<0.01,
P<0.001 vs. PSL/vehicle group (Mann-Whitney’s U-test). P<0.05 vs. PSL/vehicle group (Dunn’s test). 0.0 0.5 1.0 1.5 2.0 0.001 0.01 0.1 1 10 ## ## ## ##
Sham / vehicle (DMSO), i.t. PSL / vehicle (DMSO), i.t.
48
Figure 29 Effects of JWH133 on neuropathic pain in cannabinoid CB2 receptor knockout mice
with PSL.
(a) The PWT to VF filaments were determined pre-surgery, and before, 0.5, 1 and 2 h after drug administration. Data are expressed as the median force (g) required to induce paw withdrawal, and vertical bars represent 1st and 3rd quartiles (n=8-10/group). (b) The PWTs (g) were plotted 1 hour after JWH133 administration. Bars represent median, and vertical bars represent 1st and 3rd quartiles (n=8-10). P<0.05 vs. PSL/vehicle group (Mann-Whitney’s U-test). P<0.05, P<0.01 vs. CB2 KO/PSL mice (Mann-Whitney’s U-test). N.S., Not significant
(b)
1 h after JWH133 administrationP
W
T
(
g
)
0.001 0.01 0.1 1 10 Vehicle JWH133 Vehicle JWH133 Wild-type / PSL CB2 KO / PSL
# N.S. -5.00 0.001 0.01 0.1 110 Wild type PSL / vehicle, i.t. (9)
50 内投与は、VF フィラメント刺激で誘発される二次神経細胞の広域作動性神経細胞の発火を抑 制することを報告し、カンナビノイド CB2受容体が脊髄での神経伝達を直接制御することを 示した89) 。カンナビノイド CB2受容体刺激薬が、カプサイシン刺激による培養後根神経節細 胞からの CGRP の遊離を抑制することも報告されている90)。これらの報告から、カンナビノ イド CB2受容体は、グリア細胞だけでなく、脊髄での神経伝達も制御する可能性が考えられ た。 本研究で、PSL 作製前のカンナビノイド CB2受容体 KO マウスの疼痛閾値は、wild-type マ ウスのそれと同様のものであった。カンナビノイド CB2 受容体は主に脾臓あるいは胸腺とい った免疫系組織に分布し、サイトカインの発現及び免疫機能の調節を行うとされており 73)、 正常状態での痛覚伝達に対するカンナビノイド CB2受容体の寄与は低いと考えられた。本章 での結果は、同様にカンナビノイド CB2受容体 KO マウスを用いた Racz らの報告と一致する 67) 。その一方で、Ibrahim らは、カンナビノイド CB2受容体 KO/PSL マウスで、熱刺激に対す る逃避行動(熱刺激に対する反応潜時)が増悪傾向、つまり神経障害性疼痛の症状が増悪さ れたことを報告した91) 。しかしながら、本実験では、カンナビノイド CB2受容体 KO/PSL マ ウスの疼痛閾値の低下は、wild-type マウスのそれと同程度であり、神経障害性疼痛の症状の 増悪は認められなかった。Ibrahim らは熱刺激に対する反応潜時を測定したため、この評価法 の違いがカンナビノイド CB2受容体 KO マウスの疼痛閾値の変化に対する差異になったと考 えられた。 神経障害性疼痛モデルの脊髄では、カンナビノイド CB2受容体の発現増加に加えて、内因
51
54
Figure 30 Schematic representing an overview of the stages of the neuro-inflammation and development of neuropathic pain.
(1) Infection, tissue injury, or stress can trigger the immune response. This leads to the activation and proliferation of resident immune cells (e.g. mast cells and macrophages). Activated immune cells release inflammatory mediators (e.g. IL-1β, TNF-α, MCP-1, ATP, PGE2 and PGI2), which mediate recruitment of circulating immune cells. (2) Inflammatory mediators can modulate neuronal excitability and synaptic function for instance in the somatosensory system leading to peripheral sensitization. Nociceptor inputs can trigger a prolonged increase in the excitability and synaptic efficacy of secondary neuron, and activate glial cells (microglia and astrocyte), which lead central sensitization.
56 引用文献
1. Pain terms: a list with definitions and notes on usage. Recommended by the IASP Subcommittee on Taxonomy. Pain. 1979;6:249.
2. Tavares I and Martins I. Gene Therapy for Chronic Pain Management, Gene Therapy - Tools and Potential Applications, Dr. Francisco Martin (Ed.). 2013.
3. Bloom JR, Stewart SL, Chang S and Banks PJ. Then and now: quality of life of young breast cancer survivors. Psychooncology. 2004;13:147-60.
4. Spiegel D, Sands S and Koopman C. Pain and depression in patients with cancer. Cancer. 1994;74:2570-8. 5. 日本神経治療学会治療指針作成委員会. 標準的神経治療:慢性疼痛. 神経治療. 2010;27:593-622. 6. Bouhassira D, Lanteri-Minet M, Attal N, Laurent B and Touboul C. Prevalence of chronic pain with neuropathic characteristics in the general population. Pain. 2008;136:380-7.
7. Classification of chronic pain. Descriptions of chronic pain syndromes and definitions of pain terms. Prepared by the International Association for the Study of Pain, Subcommittee on Taxonomy. Pain Suppl. 1986;3:S1-226.
8. Treede RD, Jensen TS, Campbell JN, Cruccu G, Dostrovsky JO, Griffin JW, Hansson P, Hughes R, Nurmikko T and Serra J. Neuropathic pain: redefinition and a grading system for clinical and research purposes. Neurology. 2008;70:1630-5.
9. Gilron I, Watson CP, Cahill CM and Moulin DE. Neuropathic pain: a practical guide for the clinician. CMAJ. 2006;175:265-75.
10. 小川節郎、井関雅子、菊地臣一. わが国における慢性疼痛および神経障害性疼痛に関する大規模実態調
査. 臨床整形外科. 2012;47:565-574.
11. Quasthoff S and Hartung HP. Chemotherapy-induced peripheral neuropathy. J Neurol. 2002;249:9-17. 12. 大石了三、江頭伸昭. 抗がん剤による末梢神経障害. 福岡医学雑誌. 2013;104:171-180.
13. 日本ペインクリニック学会神経障害性疼痛薬物療法ガイドライン改訂版ワーキンググループ. 神経障 害性疼痛薬物療法ガイドライン 改訂第 2 版. 真興交易(株)医書出版部. 2016 年 7 月.
14. Tsuda M, Shigemoto-Mogami Y, Koizumi S, Mizokoshi A, Kohsaka S, Salter MW and Inoue K. P2X4 receptors induced in spinal microglia gate tactile allodynia after nerve injury. Nature. 2003;424:778-83.
15. Dworkin RH, O'Connor AB, Backonja M, Farrar JT, Finnerup NB, Jensen TS, Kalso EA, Loeser JD, Miaskowski C, Nurmikko TJ, Portenoy RK, Rice AS, Stacey BR, Treede RD, Turk DC and Wallace MS. Pharmacologic management of neuropathic pain: evidence-based recommendations. Pain. 2007;132:237-51.
16. OCHI Yasuo HT, SUZUKI Misaki, ARAKAWA Akio and SUZUKI Makoto Mechanism of Action and Clinical Efficacy of Pregabaline (Lyrica Capsule), a Novel Analgesic for Peripheral Neuropathic Pain. Japanese Journal of Pharmaceutical Palliative Care and Sciences. 2011;4:53-64.
17. Ochi Y, Harada T, Kikuchi C and Arakawa A. Pharmacology profile and clinical findings of pregabalin (Lyrica capsule). Nihon Yakurigaku Zasshi. 2010;136:165-74.
57
19. Fallon MT. Neuropathic pain in cancer. Br J Anaesth. 2013;111:105-11.
20. Nakagawa T, Isami K, Haraguchi K, So K, Asakura K, Shirakawa H and Kaneko S. Roles of transient receptor potential melastatin 2 expressed on immune cells in neuropathic pain. Yakugaku Zasshi. 2014;134:379-86.
21. Misawa S. Pathophysiology of neuropathic pain: Na+ channel and hyperexcitability of primary afferents. Brain Nerve. 2012;64:1249-53.
22. Kobayashi Y, Kiguchi N, Saika F and Kishioka S. Exploration of novel therapeutic targets for neuropathic pain based on the regulation of immune cells. Nihon Shinkei Seishin Yakurigaku Zasshi. 2015;35:65-72.
23. Horiuchi A, Abe Y, Miyake M, Osuka Y and Kimura S. Natural human IgG inhibits the production of tumor necrosis factor-alpha and interleukin-1 alpha through the Fc portion. Surg Today. 1993;23:241-5.
24. Kishimoto C, Takada H, Kawamata H, Umatake M and Ochiai H. Immunoglobulin treatment prevents congestive heart failure in murine encephalomyocarditis viral myocarditis associated with reduction of inflammatory cytokines. J Pharmacol Exp Ther. 2001;299:645-51.
25. Starowicz K and Przewlocka B. Modulation of neuropathic-pain-related behaviour by the spinal endocannabinoid/endovanilloid system. Philos Trans R Soc Lond B Biol Sci. 2012;367:3286-99.
26. Malmberg AB and Basbaum AI. Partial sciatic nerve injury in the mouse as a model of neuropathic pain: behavioral and neuroanatomical correlates. Pain. 1998;76:215-22.
27. Tal M and Bennett GJ. Extra-territorial pain in rats with a peripheral mononeuropathy: mechano-hyperalgesia and mechano-allodynia in the territory of an uninjured nerve. Pain. 1994;57:375-82.
28. Perkins NM and Tracey DJ. Hyperalgesia due to nerve injury: role of neutrophils. Neuroscience. 2000;101:745-57.
29. Zuo Y, Perkins NM, Tracey DJ and Geczy CL. Inflammation and hyperalgesia induced by nerve injury in the rat: a key role of mast cells. Pain. 2003;105:467-79.
30. Kim CF and Moalem-Taylor G. Detailed characterization of neuro-immune responses following neuropathic injury in mice. Brain Res. 2011;1405:95-108.
31. Kumar V and Sharma A. Neutrophils: Cinderella of innate immune system. Int Immunopharmacol. 2010;10:1325-34.
32. Moalem G, Xu K and Yu L. T lymphocytes play a role in neuropathic pain following peripheral nerve injury in rats. Neuroscience. 2004;129:767-77.
33. Wagner R and Myers RR. Endoneurial injection of TNF-a produces neuropathic pain behaviors. Neuroreport. 1996;7:2897-901.
34. Zelenka M, Schafers M and Sommer C. Intraneural injection of interleukin-1b and tumor necrosis factor-alpha into rat sciatic nerve at physiological doses induces signs of neuropathic pain. Pain. 2005;116:257-63.
35. Abbadie C, Lindia JA, Cumiskey AM, Peterson LB, Mudgett JS, Bayne EK, DeMartino JA, MacIntyre DE and Forrest MJ. Impaired neuropathic pain responses in mice lacking the chemokine receptor CCR2. Proc Natl Acad Sci U S A. 2003;100:7947-52.
58
2008;21:570-9.
38. Narita M, Yoshida T, Nakajima M, Narita M, Miyatake M, Takagi T, Yajima Y and Suzuki T. Direct evidence for spinal cord microglia in the development of a neuropathic pain-like state in mice. J Neurochem. 2006;97:1337-48. 39. Watkins LR, Milligan ED and Maier SF. Spinal cord glia: new players in pain. Pain. 2001;93:201-5.
40. Ohsawa M, Yamamoto S and Ono H. Contribution of the sensitization of supraspinal nociceptive transmission in chronic pain. Yakugaku Zasshi. 2014;134:387-95.
41. Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD and Julius D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389:816-24.
42. Woolf CJ and Ma Q. Nociceptors--noxious stimulus detectors. Neuron. 2007;55:353-64.
43. Bennett GJ and Xie YK. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 1988;33:87-107.
44. Field MJ, Bramwell S, Hughes J and Singh L. Detection of static and dynamic components of mechanical allodynia in rat models of neuropathic pain: are they signalled by distinct primary sensory neurones? Pain. 1999;83:303-11. 45. Li HS and Zhao ZQ. Small sensory neurons in the rat dorsal root ganglia express functional NK-1 tachykinin receptor. Eur J Neurosci. 1998;10:1292-9.
46. Ma QP. Expression of capsaicin receptor (VR1) by myelinated primary afferent neurons in rats. Neurosci Lett. 2002;319:87-90.
47. Valtschanoff JG, Rustioni A, Guo A and Hwang SJ. Vanilloid receptor VR1 is both presynaptic and postsynaptic in the superficial laminae of the rat dorsal horn. J Comp Neurol. 2001;436:225-35.
48. Guo A, Vulchanova L, Wang J, Li X and Elde R. Immunocytochemical localization of the vanilloid receptor 1 (VR1): relationship to neuropeptides, the P2X3 purinoceptor and IB4 binding sites. Eur J Neurosci. 1999;11:946-58. 49. Rose KE, Lunardi N, Boscolo A, Dong X, Erisir A, Jevtovic-Todorovic V and Todorovic SM. Immunohistological demonstration of CaV3.2 T-type voltage-gated calcium channel expression in soma of dorsal root ganglion neurons and peripheral axons of rat and mouse. Neuroscience. 2013;250:263-74.
50. Lever IJ, Robinson M, Cibelli M, Paule C, Santha P, Yee L, Hunt SP, Cravatt BF, Elphick MR, Nagy I and Rice AS. Localization of the endocannabinoid-degrading enzyme fatty acid amide hydrolase in rat dorsal root ganglion cells and its regulation after peripheral nerve injury. J Neurosci. 2009;29:3766-80.
51. Yu L, Yang F, Luo H, Liu FY, Han JS, Xing GG and Wan Y. The role of TRPV1 in different subtypes of dorsal root ganglion neurons in rat chronic inflammatory nociception induced by complete Freund's adjuvant. Mol Pain. 2008;4:61. 52. Ishikawa T, Miyagi M, Ohtori S, Aoki Y, Ozawa T, Doya H, Saito T, Moriya H and Takahashi K. Characteristics of sensory DRG neurons innervating the lumbar facet joints in rats. Eur Spine J. 2005;14:559-64.
53. Lawson SN and Waddell PJ. Soma neurofilament immunoreactivity is related to cell size and fibre conduction velocity in rat primary sensory neurons. J Physiol. 1991;435:41-63.
59
55. Kissin I and Szallasi A. Therapeutic targeting of TRPV1 by resiniferatoxin, from preclinical studies to clinical trials. Curr Top Med Chem. 2011;11:2159-70.
56. 富永真琴. TRP チャネルと痛み. 日本薬理学雑誌. 2006;127:128-132.
57. Constantin CE, Mair N, Sailer CA, Andratsch M, Xu ZZ, Blumer MJ, Scherbakov N, Davis JB, Bluethmann H, Ji RR and Kress M. Endogenous tumor necrosis factor a (TNFa) requires TNF receptor type 2 to generate heat hyperalgesia in a mouse cancer model. J Neurosci. 2008;28:5072-81.
58. Ji RR, Samad TA, Jin SX, Schmoll R and Woolf CJ. p38 MAPK activation by NGF in primary sensory neurons after inflammation increases TRPV1 levels and maintains heat hyperalgesia. Neuron. 2002;36:57-68.
59. Xiao L, Cheng J, Zhuang Y, Qu W, Muir J, Liang H and Zhang D. Botulinum toxin type A reduces hyperalgesia and TRPV1 expression in rats with neuropathic pain. Pain Med. 2013;14:276-86.
60. Kanai Y, Nakazato E, Fujiuchi A, Hara T and Imai A. Involvement of an increased spinal TRPV1 sensitization through its up-regulation in mechanical allodynia of CCI rats. Neuropharmacology. 2005;49:977-84.
61. Gunthorpe MJ and Chizh BA. Clinical development of TRPV1 antagonists: targeting a pivotal point in the pain pathway. Drug Discov Today. 2009;14:56-67.
62. Masuda T, Tsuda M, Yoshinaga R, Tozaki-Saitoh H, Ozato K, Tamura T and Inoue K. IRF8 is a critical transcription factor for transforming microglia into a reactive phenotype. Cell Rep. 2012;1:334-40.
63. Chen Y, Willcockson HH and Valtschanoff JG. Influence of the vanilloid receptor TRPV1 on the activation of spinal cord glia in mouse models of pain. Exp Neurol. 2009;220:383-90.
64. Clark AK, Yip PK, Grist J, Gentry C, Staniland AA, Marchand F, Dehvari M, Wotherspoon G, Winter J, Ullah J, Bevan S and Malcangio M. Inhibition of spinal microglial cathepsin S for the reversal of neuropathic pain. Proc Natl Acad Sci U S A. 2007;104:10655-60.
65. DeLeo JA and Yezierski RP. The role of neuroinflammation and neuroimmune activation in persistent pain. Pain. 2001;90:1-6.
66. Puffenbarger RA, Boothe AC and Cabral GA. Cannabinoids inhibit LPS-inducible cytokine mRNA expression in rat microglial cells. Glia. 2000;29:58-69.
67. Racz I, Nadal X, Alferink J, Banos JE, Rehnelt J, Martin M, Pintado B, Gutierrez-Adan A, Sanguino E, Manzanares J, Zimmer A and Maldonado R. Crucial role of CB2 cannabinoid receptor in the regulation of central immune
responses during neuropathic pain. J Neurosci. 2008;28:12125-35.
68. Walker JM and Hohmann AG. Cannabinoid mechanisms of pain suppression. Handb Exp Pharmacol. 2005:509-54.
69. Pacher P, Batkai S and Kunos G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol Rev. 2006;58:389-462.
70. Egertova M and Elphick MR. Localisation of cannabinoid receptors in the rat brain using antibodies to the intracellular C-terminal tail of CB. J Comp Neurol. 2000;422:159-71.
71. Bridges D, Rice AS, Egertova M, Elphick MR, Winter J and Michael GJ. Localisation of cannabinoid receptor 1 in rat dorsal root ganglion using in situ hybridisation and immunohistochemistry. Neuroscience. 2003;119:803-12.