せん断応力と一酸化窒素が肝細胞アンモニア代謝機 能に及ぼす影響
隅井, 干城
https://doi.org/10.15017/1807019
出版情報:Kyushu University, 2016, 博士(工学), 課程博士 バージョン:
権利関係:Fulltext available.
せん断応力と一酸化窒素が
肝細胞アンモニア代謝機能に及ぼす影響
平成29年3月
九州大学大学院 工学府 機械工学専攻
隅井 干城
i
目次
第
1
章 序論... 1
1.1
本研究の背景... 1
1.2
本研究の目的 ... 31.3
本論文の構成 ... 4第
2
章 肝臓に関する基礎事項と従来研究... 5
2.1
緒言 ... 52.2
肝臓 ... 52.2.1
肝臓の構造... 5
2.2.2
肝臓の機能 ... 82.2.3
肝細胞索と肝臓類洞微小構造 ... 82.3
微小環境に含まれる細胞... 11
2.3.1
肝細胞 ... 112.3.2
星細胞 ... 132.3.3
類洞内皮細胞... 14
2.4
せん断応力 ... 152.4.1
せん断応力とは ... 152.4.2
生体内の血管におけるせん断応力... 16
2.4.3
せん断応力説とせん断速度説 ... 162.5
生体内での一酸化窒素の役割 ... 182.6
生体外における肝細胞機能向上に関する従来研究... 20
2.6.1
肝細胞と星細胞の同時培養 ... 202.6.2
肝細胞と内皮細胞の同時培養 ... 202.6.3
せん断応力負荷による肝細胞機能の変化... 21
2.6.4
一酸化窒素負荷による肝細胞機能の変化 ... 222.7
本論文の研究意義 ... 222.8
結言... 23
第
3
章 生体内肝臓構造模擬モデルによる肝細胞アンモニア代謝機能の変化... 25
3.1
緒言 ... 253.2
実験方法... 25
3.2.1
細胞培養 ... 253.2.2
コラーゲンゲルの作製方法... 26
3.2.3
肝臓構造模擬モデルの作製 ... 263.2.4
細胞へのせん断応力負荷方法... 29
3.2.5
実験装置の全体図... 32
3.2.6
肝細胞機能の測定 ... 343.2.7
一酸化窒素濃度測定と一酸化窒素産生阻害実験... 34
3.2.8
統計処理... 36
3.3
実験結果 ... 373.3.1
培養モデルの細胞数と細胞画像... 37
3.3.2
静置培養におけるアンモニア代謝量... 39
3.3.3
せん断応力負荷におけるアンモニア代謝量 ... 403.3.4
せん断応力負荷時における一酸化窒素産生量... 42
3.3.5
一酸化窒素産生阻害剤によるアンモニア代謝量の変化... 44
3.4
考察 ... 463.4.1
静置培養におけるアンモニア代謝量の変化... 46
3.4.2
せん断応力負荷時におけるアンモニア代謝量の変化... 47
3.4.3
各モデルにおける一酸化窒素産生量とアンモニア代謝量の変化 ... 483.4.4
一酸化窒素産生阻害によるアンモニア代謝量の変化... 49
3.5
結言... 51
第
4
章 肝細胞単一モデルにおける一酸化窒素と肝細胞アンモニア代謝機能の関連性.... 52
4.1
緒言... 52
4.2
実験方法... 52
4.2.1
一酸化窒素負荷装置の作製 ... 524.2.2
せん断応力負荷と一酸化窒素負荷の条件... 58
4.2.3
一酸化窒素濃度の測定... 58
4.2.4
細胞培養方法 ... 584.2.5
細胞状態の観察方法と生存率測定方法... 60
4.2.6
肝細胞機能の測定... 60
4.2.7
統計処理 ... 604.3
実験結果... 61
4.3.1
一酸化窒素負荷装置による一酸化窒素透過濃度の計算値と測定値... 61
4.3.2
一酸化窒素とせん断応力負荷による細胞への影響 ... 634.3.3
静置培養における一酸化窒素がアンモニア代謝量に与える影響... 65
iii
4.3.4
せん断応力負荷時において一酸化窒素がアンモニア代謝量に及ぼす影響... 66
4.4
考察 ... 674.4.1
負荷装置による一酸化窒素供給濃度の評価... 67
4.4.2
細胞に負荷される一酸化窒素流束... 68
4.4.3
実験による細胞への影響 ... 694.4.4
静置培養における一酸化窒素とアンモニア代謝量の関連性... 70
4.4.5
せん断応力負荷における一酸化窒素とアンモニア代謝量の関連性... 71
4.4.6
一酸化窒素負荷による肝細胞アンモニア代謝機能向上のメカニズム ... 724.5
結言... 74
第
5
章 肝細胞と星細胞の2
種培養モデルにおける一酸化窒素の影響... 75
5.1
緒言 ... 755.2
実験方法... 76
5.2.1
使用する細胞と細胞培養方法... 76
5.2.2
肝細胞と星細胞の2
種培養モデル作製方法 ... 765.2.3
一酸化窒素負荷条件と培養液による流れ負荷... 78
5.2.4
細胞の免疫染色による画像撮影... 78
5.2.5
肝細胞機能測定 ... 795.2.6
肝細胞増殖因子の産生量測定... 79
5.2.7
一酸化窒素負荷による星細胞の状態変化... 79
5.2.8
一酸化窒素負荷と肝細胞増殖因子投与が代謝機能向上に及ぼす影響 ... 805.2.9
統計処理... 80
5.3
実験結果... 81
5.3.1
肝細胞と星細胞の2
種培養モデル画像 ... 815.3.2
一酸化窒素が2
種培養モデルにおけるアンモニア代謝機能に及ぼす影響.. 83
5.3.3
肝細胞増殖因子産生における一酸化窒素の影響... 85
5.3.4
一酸化窒素負荷による星細胞の状態変化 ... 875.3.5
一酸化窒素と肝細胞増殖因子による代謝機能向上変化... 90
5.4
考察... 93
5.4.1
肝細胞と星細胞の2
種培養モデルにおける一酸化窒素負荷の影響 ... 935.4.2
一酸化窒素負荷による肝細胞増殖因子産生量の変化... 94
5.4.3
一酸化窒素負荷による星細胞の状態変化... 94
5.4.4
一酸化窒素負荷と肝細胞増殖因子がアンモニア代謝機能に及ぼす影響 ... 955.4.5
一酸化窒素と肝細胞増殖因子が代謝機能を向上させるメカニズム... 96
5.5
結言... 99
第
6
章 総括... 100
第
7
章 付録... 106
7.1
せん断応力負荷条件... 106
7.2
流束計算に用いた2
重境界膜理論 ... 110参考文献
... 114
発表目録
... 122
謝辞
... 125
1
第 1 章 序論
1.1 本研究の背景
肝臓は
500
以上の機能を持ち,アンモニアを無害な尿素にする解毒機能やア ルブミン産生など生命維持に必要な物質の産生を行う,人体の中でも重要な臓 器の一つである.それゆえ,長期間に及ぶ重篤な肝不全は生命維持に対して致 死的な状態を引き起こし,これにより日本において毎年約50,000
人が死亡して いる.現在までにおける重篤な肝不全に対する最も有効な治療法は肝移植であ るが,肝移植を必要とする患者に対するドナー不足や高額な医療費,移植後の 拒絶反応などに対するケアなど肝移植には様々な問題が存在する.特にドナー 不足は深刻な問題であり,近年日本では年間約400
件しか肝移植が行われてい ない[1].そこで,移植するまでの肝機能の補助,もしくは肝臓の代替品として人工肝 臓が用いられている.人工肝臓は患者の血液を装置に流し込むポンプと,肝臓 の機能を担うバイオリアクターで成り立っており,バイオリアクター中には肝 細胞が充填されている[2].この肝細胞は解毒などの肝機能を司る細胞である.
バイオリアクターの中に患者の血液を流すことで,解毒や必要物質の交換など が行われる.また近年,人工肝臓の他に肝臓再生医療も盛んに行われており,
生体外で小型肝臓を作製したという報告がある[3].しかし,生体内から生体外 に取り出した肝細胞は機能や生存期間が大幅に低下するため,肝臓の代用品と しては不十分である[4].また,機能が低下した肝細胞を使用すると,人工肝臓 や再生肝臓の巨大化・コスト増大という問題が発生する.したがって,生体外 で肝細胞の機能や生存期間を生体内のレベルにまで戻すことは,人工肝臓や肝 臓再生医療の改善につながる重要な研究である.そこで生体外で肝細胞機能を 向上させるために,肝臓構造やせん断応力などの力学刺激と,一酸化窒素など の化学刺激が肝細胞に与える影響について着目がされている.
生体内の肝臓における血管周辺の構造は
Fig. 1-1
の示すようになっており,血 管を構成する内皮細胞の外側に星細胞が,星細胞の外側に肝細胞が存在する立 体構造となっている.生体内の肝臓は肝機能を司る肝細胞だけではなく,複数 の細胞が整列した配置で存在することによって3
次元構造を成している.また 血管には血液が流れており,血流によって発生したせん断応力が内皮細胞に刺 激を与えている.Fig. 1-1より,生体内の肝細胞は他の細胞と接触しかつ,血流 の影響を受けていることが分かる.したがって,肝細胞以外の細胞や血流が生 体内における高い肝細胞機能発現を引き起こしていると推測され,従来研究に おいて肝細胞と肝細胞以外の細胞を共に培養する研究,肝細胞に血流を模した せん断応力を負荷する研究が行なわれた[5].
しかし,これら従来研究では肝細胞 とそれ以外の細胞をただ混合させ培養する方法や,2
種類の細胞を3
次元に立体 的培養する方法など,Fig. 1-1の様な3
種細胞による肝臓構造が考慮されていな かった.また,生体内では血流によるせん断応力は血管を構成する内皮細胞に のみ負荷されており,従来研究におけるせん断応力を直接肝細胞に負荷する研 究では,生体内における血流の影響を及ぼす箇所について考慮されていなかっ た.したがって,生体内における高い肝機能発現の要因を特定するためには,生体外で
3
種細胞の肝臓構造を模擬し,肝細胞以外の細胞とせん断応力が肝細 胞に与える影響について調査することが重要だと考えられる.Fig. 1-1
生体内における血管周辺の肝臓構造3
肝臓構造以外の肝細胞機能を高める要因に一酸化窒素(
NO
)が上げられる.NO
は生体内の様々な機能に対し働く物質であり,肝臓にも影響を与えている.従来研究において,肝炎症を起こしたラットに体内
NO
濃度上昇物質を添加す ると肝炎症が減少し[6]
,生体外に取り出した肝細胞にNO
濃度上昇物質を付加 した場合も,肝細胞のアンモニア代謝によって生成される尿素の合成量が上昇 したという報告がある[7].一方で,生体外においてNO
上昇物質を肝細胞に付 加した時,肝細胞機能が減少したという研究も存在する[8]
.この様にNO
の肝 細胞機能に与える効果について2
面性が報告されている.この原因は,NO
負荷 方法が細胞によるNO
産生であり,安定して定量的なNO
濃度を細胞に負荷する ことが出来なかったからだと考えられる.細胞によるNO
産生は,温度や酸素 濃度などの培養環境や細胞の培養日数による細胞状態に大きく左右され,その ため各研究において細胞が産生するNO
濃度が異なっていたと考えられる.肝 細胞機能向上はNO
濃度に依存して変化すると推測されるが,NO
濃度と肝細胞 機能向上の関連性を調査するためには,従来の細胞によるNO
産生負荷方法で は不可能である.したがって,肝細胞に負荷するNO
濃度を調節し負荷する濃 度が定量的に分かる装置を用いて,肝細胞機能向上とNO
濃度の関連性を調査 することが重要だと考えられる.1.2 本研究の目的
生体外で肝細胞の機能を高める方法を見つけることは,人工肝臓の小型化や コスト削減,肝臓再生医療の発展に役立つ研究である.肝細胞機能向上のため に肝臓に含まれる肝細胞以外の細胞や血流によるせん断応力が着目されている が,生体内の肝臓
3
次元構造に着目した研究はない.また,細胞が産生するNO
による肝機能向上も行われているが,肝細胞に負荷するNO
濃度を定量的に調 整する方法がなく,肝細胞機能向上とNO
濃度の関係性が明らかになっていな い.そこで本研究では
2
つの目的を設定した.1
つ目は生体内の肝臓3
次元構造を 模擬した肝臓構造模擬モデルを生体外で構築し,生体内の肝臓構造が肝細胞機 能向上,特にアンモニア代謝機能向上に与える影響についての調査である.こ れにより生体内における高いアンモニア代謝機能発現の要因を特定する.2
つ目 は細胞に負荷するNO
濃度を定量的に調節できる装置を作製し,肝細胞単一モ デルに取り付け,NO
濃度とアンモニア代謝機能向上との関連性についての調査である.これにより
NO
がアンモニア代謝機能向上に与える影響を明らかにす る.本研究より生体内の肝臓3
次元構造がアンモニア代謝機能を向上させるメ カニズムを解明し,代謝機能が高い人工肝臓の作製方法を提案できると考える.1.3 本論文の構成
第
1
章では本研究の背景および目的について述べた.第2
章では本研究を理 解する上で必要な肝臓に関する知識やせん断応力,一酸化窒素などの基礎事項 を述べ,生体外におけるアンモニア代謝機能を含む肝細胞機能向上の従来研究 について紹介する.第3
章では,生体内の肝臓構造がアンモニア代謝機能向上 に及ぼす影響について調査する.そのために,生体内の肝臓3
次元構造を模擬 した培養モデルを,生体外で作製し,内皮細胞にのみせん断応力を負荷する.第
4
章では,肝細胞単一モデルにおけるアンモニア代謝機能向上とNO
濃度の 関連性について調査する.この関連性を調査するために,細胞に負荷するNO
濃度を調節する負荷装置を作製する.第5
章では,肝細胞と星細胞の培養モデ ルに異なるNO
濃度を負荷し,NO
と星細胞がアンモニア代謝機能向上に与える 影響について調査する.第6
章は総括であり,各章の結言についてまとめを行 い,本研究で明らかになったことを記述する.そして,本研究結果を応用した 人工肝臓を提案する.5
第 2 章 肝臓に関する基礎事項と従来研究
2.1 緒言
本章では,本研究を理解する上で必要となる基礎事項と肝細胞機能向上に関 する従来研究について説明する.生体内の肝臓には肝細胞の他に複数の細胞が 存在しており,それらの細胞が秩序良く配列することで肝臓構造を成している.
また,肝臓に張り巡らされた血管には血液が流れており,細胞に酸素や栄養を 運ぶと同時に血管にせん断応力という力学刺激を与えている.このせん断応力 が細胞に様々な影響を及ぼしている.体内で発生した一酸化窒素(
NO
)は臓器 に影響を与えており,臓器の機能向上や細胞死の抑制などに関わっている.こ の様に本研究で用いられる細胞や力学刺激,NO
は生体内において様々な役割を 担っている.そのため,これらの基礎知識を説明することで,本研究をより深 く把握することができると考える.また,生体外における肝細胞機能向上に関 してどのような従来研究が行われているかを紹介し,本研究を行う意義につい て説明する.2.2 肝臓
2.2.1 肝臓の構造
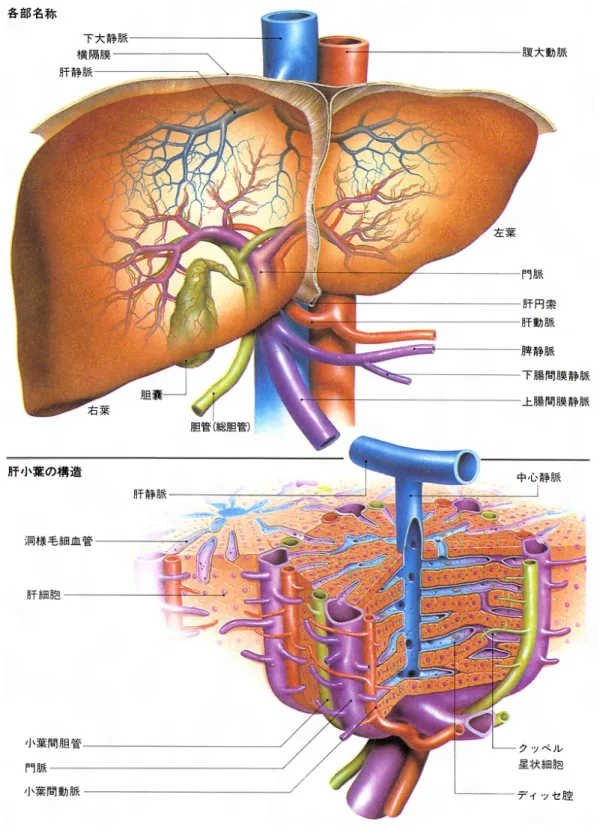
ヒト生体内の肝臓は,腹部の右上に位置する.肝臓は右葉と左葉の二葉から 成り立っている(Fig. 2-1)
[9].肝臓の最小単位は肝小葉であり,直径 0.7~2.0 mm,
高さ
0.5
~2.0 mm
の多角形状の形態をしている(Fig. 2-
1a)
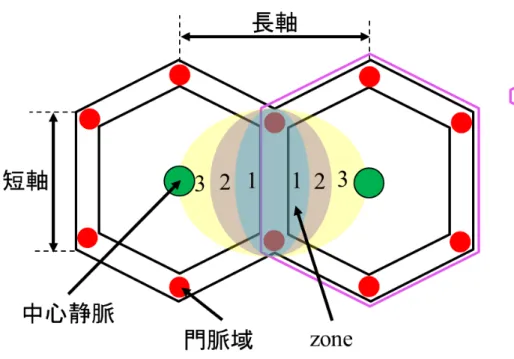
.肝小葉の中心には中 心静脈があり,肝細胞索と呼ばれる肝細胞の列が放射状に並んでいる.血液は 門脈と小葉間動脈から類洞と呼ばれる肝臓の毛細血管へ流れ込み,最終的に中 心静脈に流れる.この間に肝細胞と血液の間で物質の交換や代謝,血液の浄化 などが行われている.肝小葉3
つのzone
に分けることができる(Fig. 2-1b
)[10]
. 肝動脈と門脈周辺をzone1,肝静脈周囲を zone3, zone1
とzone3
の間がzone2
である.
zone1
からzone3
に行くに従い,酸素や栄養素の供給量が減少していく.Fig. 2-1a
肝臓構造と最小単位の肝小葉(“小学館・日本大百科全書”から転載)
7
Fig. 2-1b 肝小葉における zone
分類(
Godoy et. al., 2013
を参照)2.2.2 肝臓の機能
肝臓の機能は主に次の
3
つである[10, 11]
.(1)
代謝機能肝臓における代謝機構は,糖質・脂肪・アミノ酸・ビタミンの代謝を行って いる.代表的なものとしてブドウ糖の分解による
ATP
の産生や血漿蛋白である アルブミンの産生,ビタミンA
の貯蓄が挙げられる.(2)
排泄機構ビリルビンは脂溶性のタンパク質であり,それが血漿タンパクであるアルブ ミンと結合して肝臓まで運ばれる.そこで肝細胞により極性基を付加され,胆 汁中に分泌できるようにグルクロン酸と抱合し,抱合ビリルビンという形の水 溶性に変換して,胆汁に分泌させている.尿素の合成も肝臓で行われ,95%の アンモニアが尿素となる.不要になったタンパク質が脱アミノ化されてアンモ ニアが生じ,肝臓でアンモニアから尿素へと変換されている
[12]
.(3)
解毒機能肝臓は,体に入った異物をさまざまな化学反応で,毒性が少なくかつ排泄し やすい水溶性物質に変え,尿中や胆汁中に排泄させている.また,肝臓の類洞 にあるクッパー細胞は,門脈から肝臓内に入った毒素や異物を貪食し,血流循 環の解毒作用を行っている.
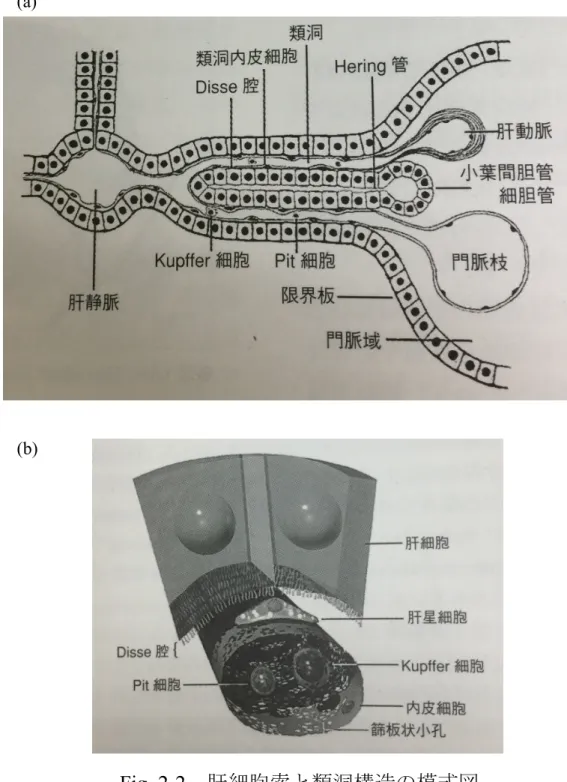
2.2.3 肝細胞索と肝臓類洞微小構造
肝小葉を構成する肝細胞索を
Fig. 2-2a
に示す[13]
.肝細胞索は放射状の列を形 成し中心静脈へと向かっている.放射状に並んだ肝細胞索の間は類洞と呼ばれ ている.類洞における微小構造をFig. 2-2b
に示す[5, 13].血管である類洞の周り に肝細胞が存在している.肝臓では類洞と肝細胞の間にDisse
腔という空間があ り,この空間には肝星細胞が内皮細胞と肝細胞の足場として存在する.類洞内 にはクッパー細胞やPit
細胞など様々な細胞が存在する.Disse 腔と類洞に含ま れている様々な細胞が肝細胞と相互作用を行っている[14]
.肝細胞を実質細胞,その他の細胞を非実質細胞と呼ぶ.また,隣接する肝細胞間には毛細胆管が存 在し,肝細胞で産生された胆汁がこの胆管内を通る.本研究では類洞微小構造
9
を生体内の肝臓微小構造と定義し,研究に用いた.
次の項にて,肝臓微小構造に含まれる各細胞のうち本研究で使用する
3
つの 細胞の機能について解説する.Fig. 2-2 肝細胞索と類洞構造の模式図 (a)
肝細胞索(“肝臓病学”から転載)(b)
類洞構造の3
次元模式図(“肝臓病学”から転載)(b)
(a)
11
2.3 微小環境に含まれる細胞
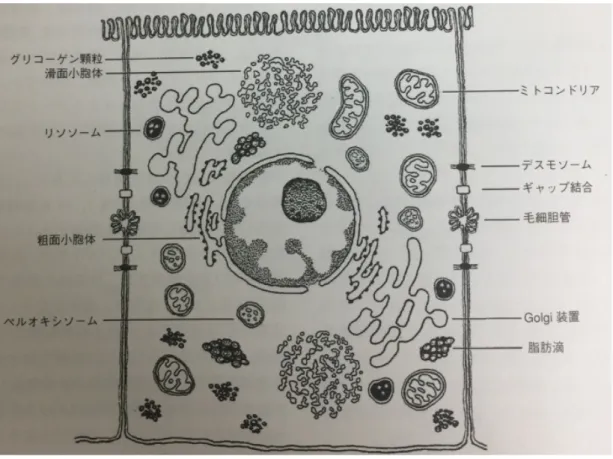
2.3.1 肝細胞
アンモニア代謝やアルブミン産生などの肝臓の機能を司るのがこの肝細胞で ある.肝細胞は直径
15
~30 µm
の多面体の細胞であり,隣接する肝細胞との間 に毛細胆管を形成する.肝細胞の形態をFig. 2-3
に示した[13].肝細胞は肝臓の容積の
85%,総細胞数の 65%を占めている[15].肝細胞はアルブミンや α
フェトプロテイン(
AFP
),グリコーゲン,サイトケラチン(CK
)8
,18
などのタン パク質を発現する.肝細胞は上皮細胞でありながら,血管との間に基底膜を欠 き,Disse腔に向けて多数の微絨毛を出して,表面積を拡大している.また肝細胞は
Disse
腔と類洞側に面する面(血管極)と毛細胆管に面する面(胆管極)の2
種類の極性を有している.それぞれの細胞膜に特定のタンパク質のレセプター を持ち,類洞面においては,アルブミンと低密度リポタンパク(LDL)を,毛 細胆管膜には胆汁を分泌している.肝細胞は肝小葉の中で機能の不均一性を示 し,類洞側では栄養や酸素を取り込む機能が優位であり,中心部では胆汁や代 謝産物を分泌する機能が優位である.この不均一性からも肝細胞は非常に機能 分化に優れた細胞であると言える.この様に優れた機能を司る肝細胞であるが,生体内から生体外に肝細胞を取 り出してしまうとこれらの機能と生存期間が大幅に低下してしまう[16-18].そ のため生体外で肝細胞を用いた研究が行いにくく,未だに肝細胞に関して未知 な部分が多い.
Fig. 2-3
肝細胞の構造(“肝臓病学”から転載)
13
2.3.2 星細胞
星細胞は
Disse
腔に存在する細胞で,多くの脂肪滴を細胞質内に持つ.この細 胞は類洞内皮細胞の外側面に沿って細長い樹枝状の細胞突起を伸展し,その一 部は肝細胞間を通り隣の類洞外側に達している.隣接する星細胞はこの細胞突 起を介して互いに接着する.星細胞は類洞壁細胞の13%を占めている[19].さら
にその構造的特徴から細胞収縮により類洞の血流調節を司っている.星細胞の脂肪滴は大量のビタミン
A
を含んでおり,生体内に存在するビタミン
A
の80~90%を貯蔵している[19].星細胞が線維化すると細胞内の脂肪滴は
減少し,細胞内には発達した粗面小胞体とその細胞周囲には多くのコラーゲン 束が見られる.このことから,星細胞はビタミン
A
の代謝と細胞外マトリクス の産生に関与していることが明らかとなった.また星細胞の線維化は活性化状 態と呼ばれ[20]
,星細胞が静止化状態から活性化状態になることで肝硬変などの 炎症に繋がる[21].星細胞はさまざまな肝細胞の増殖を亢進させる生理活性物質を産生する.ヘ パ リ ン 結 合 増 殖 因 子 や , イ ン ス リ ン 様 増 殖 因 子 に 加 え , 肝 細 胞 増 殖 因 子
(Hepatocyte Growth factor; HGF)産生が認められている[22].
2.3.3 類洞内皮細胞
肝臓内の血管は類洞と呼ばれており,類洞を構成する細胞は類洞内皮細胞と 呼ばれている.類洞内皮細胞と血管内皮細胞には大きく異なる点が
2
つ存在す る.1
つは基底膜とよばれる足場が存在しないこと[23]
,もう1
つは孔が多数存 在することである.細胞は細胞外基質で構成される基底膜上に接着することで 自身の位置を維持している.しかし,肝臓ではこの細胞外基質が非常に少なく,類洞内皮細胞の外側には直接星細胞が接着している
[24]
.この細胞外基質が少な い原因は解明されていないが,細胞外基質が増えると肝臓が炎症を起こし肝硬 変に繋がるという報告がある[25].したがって,肝臓の性質を保つために細胞外 基質が少ないという可能性が考えられる.類洞内皮細胞は核を伴った細胞質と,小孔(約
100 nm
)を伴った師板(sieve plate
)とよばれる約0.1 µm
厚の細長く伸 展した突起からなりたっている(Fig. 2-2).この小孔は類洞と肝細胞間における 血漿成分の直接的なやり取りを可能にし,類洞とDisse
腔との間の浸透圧差をな くし,また肝細胞がDisse
腔に分泌した物質の類洞血中への移行を可能にしてい る.一方,栄養脂肪を運搬するキロミクロンは小孔の大きさに比して大きいた め,そのままでは小孔を通過できない.このように師板に存在する小孔はフィ ルター機能も果たす[26]
.15
2.4 せん断応力
従来,せん断応力は物理学,特に流体力学あるいは水力学で使用されてきた が,近年,生体内の流れや変形に関連した現象を扱うバイオレオロジーや生理 学の領域でも登場することが多くなってきた.これは,血液の流れ(血流)に より生じるせん断応力が血管壁に作用し,血管の機能に影響を与える可能性が あるなど,生体内における意義が注目されるようになったからである.
2.4.1 せん断応力とは
血管壁には血流によるせん断応力と血圧による張力・貫壁性応力などの力学 的応力が作用している.せん断応力は血管壁の内面に存在している内皮細胞に 対して働き,内皮細胞を血流方向にゆがませる性質を持つことが知られている
[27]
.せん断応力を作用させたときの培養内皮細胞の形状変化を顕微鏡で観察し たところ,流れ方向に歪むことが確認されている(Fig. 2-4)[28].Fig. 2-
4 シェアストレスによる内皮細胞の変形(“シェアストレスと内皮細胞”から転載)
2.4.2 生体内の血管におけるせん断応力
血管の径や,そこを流れる血液の速度は場所により異なるので,血管壁にか かるせん断応力も違ってくる.ヒトの生理条件下の大動脈では
1
~2 Pa
,一方,静脈では
0.1~0.6 Pa
のせん断応力が血管壁に作用していることが分かっている[29].
生体の血管内で生じているせん断応力を生体内で測定することは難しい.ま た血液速度などから大まかに計算することはできるが,それを厳密に評価する のは難しい.これは,血管は弾性があり,まっすぐな導管ではなく
3
次元的に 複雑な形態をしているからである.場所によっては血管が湾曲し多くの分岐に なっているため,血液の流れが非常に複雑化しており,血管にかかるせん断応 力の分布も複雑になる.そこで,血管に負荷されるせん断応力を計算するため にシミュレーションが主に用いられている.肝臓の血管においても直接せん断応力を計測することは難しく,シミュレー ションのみが行われている[30, 31].これらの結果より,肝臓の血管では
0.2 Pa
から
0.6 Pa
のせん断応力が負荷されていることがわかる.したがって,本研究では肝臓内のせん断応力を
0.6 Pa
とし,このせん断応力の大きさを実験に用い ることとした.2.4.3 せん断応力説とせん断速度説
せん断応力を負荷した際の肝細胞代謝において考えなければならないのが,
流体中の物質が近接する壁面上で消費される場合の拡散境界層(diffusion
boundary layer)と壁近傍のせん断速度(shear rate)との関係である[32].アンモ
ニアが拡散によって膜上に到達すると別の物質に変換されてしまう場合,細胞 膜近傍の溶液中濃度には勾配が生じる.この濃度勾配の生じる層を拡散境界層 と呼ぶ.この境界層の厚dは,流れが速いほど小さく,壁面上のせん断速度gの(-1/3)
乗に比例することが分かっている.δ = α D 𝛾
&
'
(2-1)
17
ただし,
D
は溶液中の拡散係数,a
は溶液の状況に関係ない定数,g
はせん断速 度,である.拡散境界は,溶液内物質の壁面への拡散を阻害する障壁となるの で,例えばアンモニアの細胞膜への単位時間当たりの到達量m
は障壁の厚さd に反比例する.m = β D
𝛿 ∆C (2-2)
ここで,bはある定数,⊿Cは流体中央部と壁面部での濃度差,となっている.
(2-1)
式と(2-2)
式より,せん断速度が大きい場合には境界層d
が薄く,アンモニアの到達量
m
が多くなるため,より大きなアンモニア代謝が起きると考えられ る.これを「せん断速度説」と呼ぶ(Fig. 2-5a).一方,機械的な細胞のせん断変形を主体にした考え方を「せん断応力説」と 呼ぶ.流れによって生じた壁せん断応力が細胞のせん断変形を引き起こし,機 械受容体の応答によってアンモニア代謝が起きるとする仮説である(Fig. 2-5b).
この
2
つの説のどちらが適用されるかを調査するためには,別の実験が必要 となる[33].(a) (b)
Fig. 2-5 せん断速度説とせん断応力説の図解
(a)
せん断速度説,(b)
せん断応力説2.5 生体内での一酸化窒素の役割
単純な
2
原子分子の一酸化窒素(NO)は,思いがけない多彩な能力を生体内 で発揮している.現在多くの研究者がNO
の挙動に注目している.NO
の生理作用に関しては,1970
年代後半から1980
年代初頭にMurad, Ignaro,
Furchgott
らが萌芽的研究を報告しており,それぞれニトログリセリン,グアニル酸シクラーゼ,血管内皮由来弛緩因子を研究対象にしていた.
1986
年,1987
年にNO
が血管を拡張させる因子であることを確認し,これらの成果からMurad,
Ignaro, Furchgott
らは1998
年にノーベル生理学賞を受賞した.この結果をはじめとし,様々な分野で
NO
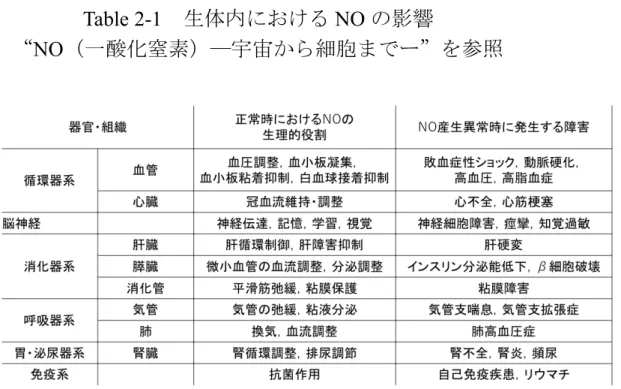
に関する研究が行われた.現在までにTable 2-1
のよう な効果があることが分かっている[34].この表から分かる通り,NOには2
面性 が存在する.この2
面性についてはまだ詳しく解明されていない.生体内で
NO
を産生させるために必要な物質がアルギニンというアミノ酸で ある.アルギニンは内皮細胞に取り入れられ,酵素によってNO
へと変換され る.しかし,アルギニンは非必須アミノ酸なため体内にあまり取り込まれない ため,外部から摂取する必要がある[35]
.また,内皮細胞が血流によって発生す るせん断応力の影響を受け,NOを産生し始めることが報告されている[36].組織内の
NO
の寿命は0.1 s
から5 s
程度だと報告されている[34].非常に寿命が短い
NO
であるが,NO
の拡散係数と寿命から計算すると生体内における拡散距離は
0.1 mm
から0.5 mm
となっている[34].細胞の平均的な大きさが10 µm
であることから,NOはかなり遠くの細胞にまで影響を与えることができる.
肝臓において
NO
は重要な役割を担っており[37]
,細胞死の抑制[38, 39]
,肝臓 再生[40],肝臓障害の予防[41]などの働きを行っている.しかし一方で,肝臓に 害を与えているという報告もある.Sesti 氏らは肝細胞にNO
産生促進剤を投与 すると,細胞死が引き起こされると報告している[8]
.また,NO
はミトコンドリ アに働きかけ,細胞死を引き起こすという研究も存在する[42].したがって,NO
は肝臓にてポジティブな働きとネガティブな働きの2
つの効果があると考えら れている.19
Table 2-1
生体内におけるNO
の影響“
NO
(一酸化窒素)―宇宙から細胞までー”を参照2.6 生体外における肝細胞機能向上に関する従来研究
2.2,2.3,2.4,2.5
にて肝臓に関する基礎知識と,生体内におけるせん断応力と
NO
の役割について説明した.生体内の肝臓は多くの細胞が配列し立体構造 をなしている.肝臓機能を司る肝細胞は生体内において様々な細胞と相互作用 し,せん断応力やNO
ガスなどの影響を受けていることが分かった.したがっ て従来研究において,これら生体内における肝細胞に影響を与えている因子の 解明を行うことで,生体外で肝細胞の機能を高める方法発見に繋がるのではな いかと考えられた[43].そこで,本項では肝細胞機能向上解明に関する従来研究 について紹介する.2.6.1 肝細胞と星細胞の同時培養
肝細胞と直接接触している星細胞を同時に培養することで,生体外で肝細胞 機能が向上するのではないかという研究が行われている[18, 44-46].ここでは
Pan
氏らの研究とKrause
氏らの研究について紹介する.Pan
氏らは肝細胞と星細胞の2
種培養モデルを2
種類作製した.1つは肝細胞 と星細胞を混ぜ合わせ,単層になるように播種したモデルであり,もう1
つは 肝細胞と星細胞の直接的な接触がないように離して培養し,物質のやり取りだ け培養液を通して行うことが出来るモデルである.その結果,ミックスしたモ デルより直接接触のないモデルのほうが高い肝細胞機能発現を示した.Krause 氏らは肝細胞と星細胞を混ざらない様に平面上に培養し,直接接触のあるモデ ルと接触しないモデルの2
種類を作製した.その結果,直接接触のモデルが接 触ないのモデルより高い肝細胞機能を示した.2.6.2 肝細胞と内皮細胞の同時培養
肝細胞と直接接触していないが,血流によるせん断応力を受け様々な物質を 産生する内皮細胞[47]を肝細胞と同時に培養することで,生体外における肝細胞 機能向上に繋がるのではないかという研究が行われている[48-52].ここでは
Ohno
氏らの研究とKang
氏らの研究について紹介する.Ohono
氏らはセルシートを用いて実験を行った.セルシートは東京女子医大の岡野光夫教授らが開発した特殊な温度感受性シートであり,このシート上で
21
細胞を培養し特定の温度をシートに加えると,細胞を単層で培養皿からはがす ことが出来る.この技術により
3
次元的に複数の細胞を直接接触で重ねること が可能となる.肝細胞と内皮細胞の細胞シートをそれぞれ作製し,肝細胞シー トの上に内皮細胞のシートを重ね,物理的に両者が接触している2
種培養モデ ルを作製した.その結果,肝細胞機能の向上と内皮細胞機能の向上が共に見ら れた.Kang氏らはMicro Electro Mechanical Systems
技術を用いて,微小2
種培 養モデルを作製した.薄いポリマー膜の上下に肝細胞と内皮細胞を播種し,膜 の上下に培養液が流れる流路を設置した.ポリマー膜には小孔があり,両者の 産生した物質が通過できるようになっている.培養液を流さず静置培養した場 合,2
種培養モデルは肝細胞のみモデルより尿素合成量が少なかった.そこで,内皮細胞側の流路に培養液を流すと,
2
種培養モデルの尿素合成量が肝細胞のみ モデルより高くなった.2.6.3 せん断応力負荷による肝細胞機能の変化
血液を流すせん断応力は細胞に様々な影響を与えており,人工肝臓に血液を 流す際せん断応力により肝細胞に何らかの影響が発生していると考えられる[2,
4, 53, 54].そこで特にせん断応力と肝細胞機能向上に関して様々な研究が行われ
ている[55-59]
.細胞にせん断応力を負荷する装置は
2
種類あり,回転円板型負荷装置と平行 平板型負荷装置である.回転円板型負荷装置は,細胞を培養した円形のdish
の 中に円錐板が存在している形である.この円錐を回すことで,dish
の底面に存 在する細胞に一様な特定のせん断応力が負荷されることになる.しかし,この 装置作製には精密な工作技術が必要であり,また円錐が細胞に接触し傷害や剥 離を起きてしまう可能性がある.またdish
の中央付近では培養液の量が少なく なるため,酸素や栄養供給に問題が生じ,せん断応力以外の影響が出てしまう 危険性がある.さらに培養液を循環させることができず酸素や栄養素が枯渇す る問題も生じる.一方で平行平板型負荷装置は,ある距離を離して平行に並べ た2
枚の平板の間に培養液を灌流させるものである.この装置は平板の面積を 大きくすることで大量の細胞にせん断応力を負荷することができる.しかしこ の装置は培養液を灌流させるため多くの培養液を必要とし,装置全体も大型に なるため温度やpH
調整が問題となる.ここではこれらの負荷装置を用いたTorii
氏らの研究とTanaka
氏らの研究について紹介する.Torii
氏らは回転円板型負荷装置を用いて,肝細胞に30 rpm
(0.05 ~ 2 Pa
)のせ ん断応力を負荷した.すると120 rpm
や0 rpm
の時より高いアルブミン産生を示 した.一方で,Tanaka氏らはMicro Electro Mechanical Systems
技術を用いて幅250 µm
深さ100 µm
のマイクロ平行平板型負荷装置を作製した.その結果,せ ん断応力が大きくなるにつれアルブミン産生量が低下し,同時に肝細胞1個の 形状も細長く変化する傾向が見られた.2.6.4 一酸化窒素負荷による肝細胞機能の変化
NO
が細胞機能に影響を与えることが知られて以降,様々な分野でNO
と細胞 機能の研究が行われている.肝細胞とNO
に関する研究も行われている[7, 8, 60].従来研究において細胞に
NO
を負荷する方法は2
つ存在し,細胞にNO
を産生さ せる方法とNO
キャリアーを投与する方法である.ここではNagao
氏らの研究と
Chimenti
氏らの研究について紹介する.Nagao
氏らは培養した肝細胞に0.2 mM
のアルギニンを12
時間投与したところ,尿素合成が増加したと報告している.
Chimenti
氏らは10 µM
のNO
キャリ アーを2
時間肝細胞に投与したところ,尿素合成量が増加した.しかし,100 µM
と
150 µM
のNO
キャリアーを2
時間投与したところコントロールより減少した.また,肝細胞機能が低下した条件では細胞の生存率も低下した.
2.7 本論文の研究意義
生体外で肝細胞機能を向上させるために,生体内の肝臓構造に着目して様々 な研究が行われた.しかしこれら従来研究で明らかになっていない事柄が
2
つ 存在する.(1)生体内の
3
次元構造と肝細胞機能向上の関係性が明らかになっていない.(2)NOと肝細胞機能向上の関係性がはっきりと解明されていない.
従来研究は生体内の肝臓構造に着目はしているが,
3
種細胞による3
次元立体 肝臓構造と肝細胞機能向上の関連性についての研究は行われていない.肝細胞 と星細胞,肝細胞と内皮細胞といった,2
種類の細胞培養は行われているが,実 際の肝臓では3
種細胞が同時に存在している.2
種培養の各細胞培養配置も生体 内の構造と異なっており,肝細胞は内皮細胞と接触しておらず,星細胞とは直 接接触している.さらにこれらの細胞は縦に3
次元的に重なり合っている.肝23
細胞に直接せん断応力を負荷しているが,生体内では内皮細胞にのみせん断応 力は負荷されている.また近年マイクロデバイスを用いる研究が多いが,肝細 胞はある一定密度で集合していないと機能が低下するとの報告がある[61].将来 的に人工肝臓や肝臓再生に応用するためには,高い細胞密度で実験し,正確な 肝細胞機能変化を調査する必要がある.したがって,生体内の肝臓
3
次元構造 が肝細胞機能に与える影響について研究するためには,肝臓構造を模擬した3
次元培養モデルを生体外で作製し,せん断応力を内皮細胞にのみ負荷するよう に設計することが重要である.NO
と肝細胞機能向上に関する研究が行われているが,NOは機能向上させる と述べている従来研究と,NO
は機能低下させると述べている従来研究がある.この様に真逆の研究結果が出てしまったのは,細胞に負荷する
NO
濃度を調節 できていないからだと考えられる.細胞に産生させたNO
やNO
キャリアーから 乖離したNO
の濃度は,細胞の状態や温度などの培養条件に依存してしまう.そのため,同じ条件で実験することが難しく細胞に負荷される
NO
濃度を正確 に把握しにくい.また,培養液中に存在するNO
は酸素と反応し酸化してしま い,酸化したNO
は細胞に悪影響を与える.NO
を酸化させずに細胞に負荷する ことが重要である.以上より,NO
と肝細胞機能向上の関連性を研究するために は,負荷するNO
濃度を正確に制御でき,NO
を酸化させずに細胞に直接負荷で きる装置を新たに作製する必要がある.生体外で肝臓構造を模擬する培養装置と細胞に負荷する
NO
濃度を調節する 装置を作製することで,生体内の肝臓3
次元構造が肝細胞機能向上に与える影 響を解明し,NO
と肝細胞機能向上の関連性を明らかにしたいと考えている.2.8 結言
肝臓には様々な細胞が存在しており,実質細胞(肝細胞)と非実質細胞(星 細胞,内皮細胞)が相互作用しあっていることが分かった.また,血流によっ て発生するせん断応力や細胞が産生した
NO
が細胞や人体に様々な影響を与え ていることも分かった.これらの知見から,従来研究において生体内の肝臓に 着目して様々な肝細胞機能向上の研究が行われた.しかし,これらの従来研究 では解明されていないことが2
点ある.1
つは「生体内の肝臓3
次元構造が肝細 胞機能に与える影響」であり,もう1
つは「NOと肝細胞機能向上の関連性」で ある.これらの2
点に関する研究が行われていないのは,研究を行うための適切な実験装置が存在していないからである.
したがって,次の章からは新しい実験装置を作製し,この
2
点を解明する研 究を行っていく.本研究の結果から,生体外で肝細胞機能,特にアンモニア代 謝機能をより向上させる新しい手法が見つけられると考えている.25
第 3 章 生体内肝臓構造模擬モデルによる肝細胞アンモ ニア代謝機能の変化
3.1 緒言
本章では,生体内の肝臓構造がアンモニア代謝機能向上に与える影響につい て明らかにすることを目的とした.第
2
章で説明した通り,生体内の肝臓は肝 細胞・星細胞・内皮細胞が3
層に重なっている立体構造を成している.また血 管内皮細胞には血流によって発生したせん断応力が負荷されている.そこで,この肝臓
3
次元構造が生体内における高い肝機能発現に関与していると推測し,生体外でせん断応力を含む肝臓構造を模擬した培養モデルを構築し,肝臓構造 がアンモニア代謝機能向上に与える影響について調査した.この結果から,ア ンモニア代謝機能向上に関与する要因の特定を行った.
3.2 実験方法
3.2.1 細胞培養
内皮細胞にはラット肺毛細血管内皮細胞(
Rat Lung Microvascular Endothelial
Cells, DS
ファーマー, Japan),星細胞には不死化したラット星細胞 (RI-T,
HSRRB),そして肝細胞には不死化したラット肝細胞 (RHT33, RIKEN, Japan)を
用いた.これらの細胞は肝臓構造模擬モデルを作製するまで別々に培養された.Dulbecco’s Modified Eagle Medium(DMEM, Invitrogen, Japan)に 10%ウシ胎児血清 (Fetal Calf Serum, Biological Industries, USA)と 1%抗生物質-抗真菌剤 (Penicilin, Streptomycin, Amphotericin B,
食塩水, Invitrogen, Japan)
を添加したものを細胞培 養液として使用した.細胞培養は温度37℃,湿度 100%,CO
25%,Air 95%のイ
ンキュベーター内で行った.φ60 mm セルカルチャーディッシュ (BD Falcon,USA)
の上に細胞を播種した.セルカルチャーディッシュ上に細胞を播種する前 に,ディッシュをコラーゲンコーティングした.コーティングの手順としては,まずラット尾コラーゲンⅠ溶液 (BD Falcon, USA)を
0.02 N
酢酸で50 µg/mL
に希釈し,この溶液を
1dish
につき2 mL
滴下した.そしてディッシュの蓋を半開き にしてクリーンベンチ内で6
時間乾燥した後,リン酸緩衝液(phosphate buffersaline; PBS; Nissui Pharmaceutical, Japan)2 mL
で2
回洗浄した.3.2.2 コラーゲンゲルの作製方法
細胞を
3
次元立体構造に培養するために,コラーゲンゲルを使用した.細胞 の上にコラーゲンゲルを播くことで,1
層目細胞の上に2
層目細胞を播種するこ とが可能となり,3
次元肝臓構造模擬モデルを作製することができる.コラーゲ ンゲルの作製手順を以下に示す.なお作業はすべて氷上で行った.1. 10×PBS,ラット尾コラーゲンⅠ溶液,1 N NaOH,滅菌水の量を下記の式
より算出する.・
10×PBS
の量(ml)=
最終溶液量(mL)/10
・コラーゲン溶液量(mL)
=(最終溶液量(mL)×最終コラーゲン濃度 (3.0mg/mL)) /コラーゲンの濃度
(mg/mL)
・1N NaOHの量(mL)= 加えるコラーゲン溶液量(mL)×0.023
・滅菌水量(mL)=最終溶液量-コラーゲン溶液量-10×PBS-1N NaOH溶液量
2.
氷上で冷やした遠沈管に10×PBS
を加える.3. 10×PBS
に1N NaOH
を加える.4. 10×PBS
と1N NaOH
に滅菌水を加える.5.
氷上で遠沈管内の溶液を混合する.6.
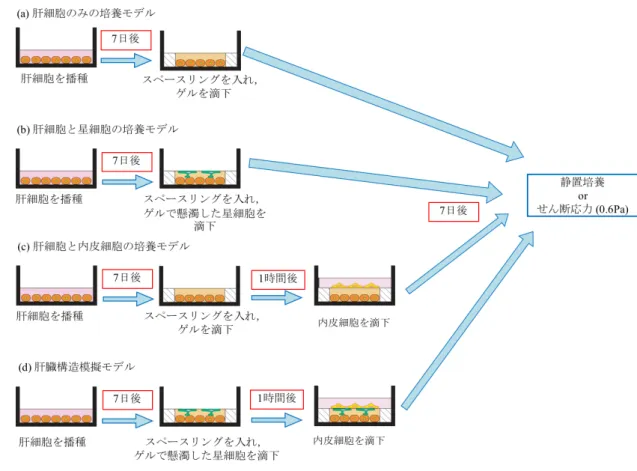
コラーゲン溶液量を加え,混合する.使用するまで氷上で保存する.3.2.3 肝臓構造模擬モデルの作製
模擬モデルの作製は
φ60 mm
セルカルチャーディッシュ上で作製した.コラー ゲンコーティングしたディッシュ上に肝細胞,星細胞,内皮細胞をFig. 3-1d
の 手順で培養し,肝臓構造模擬モデルを作製した.まずφ60 mm
ディッシュに肝細 胞を播種した.7 日後,培養液を取り除き肝細胞の上からスペースリング(
SUS304,
外径50 mm,
内径40 mm,
厚さ100 µm;
ミクロ精工, Japan
)を設置し た.次にコラーゲンゲル溶液450 µL
で懸濁した星細胞をスペースリング内に滴 下し,あふれたコラーゲンゲル溶液は取り除いた.37℃で1
時間インキュベー27
トしてコラーゲンをゲル化させた後,内皮細胞をゲル上に播種した.
7
日間培養 したのち,作製した3
種細胞培養モデルをせん断応力負荷装置に取り付け,内 皮細胞にのみせん断応力を負荷した.また比較対象として,肝細胞のみモデル(
Fig. 3-1a
),肝細胞と星細胞のモデ ル(Fig. 3-1b),肝細胞と内皮細胞のモデル(Fig. 3-1c)を作製した.さらに,せ ん断応力の効果を調べるために,せん断応力を負荷しない静置培養下で各モデ ルの機能向上を調査した.Fig. 3-1 肝臓構造模擬モデルの作製手順
29
3.2.4 細胞へのせん断応力負荷方法
3.2.3
で作製した各モデルに0.6 Pa
せん断応力を負荷するために,平行平板型フローチャンバーを作製した(
Fig. 3-2
).Fig. 3-2a
の寸法で作製したフローチャンバーを
Fig. 3-2b
の様にディッシュの上から被せた.ディッシュとチャンバーの間に
1 mm
シリコンシートを噛ませた.フローチャンバー装着後の断面図はFig. 3-2c
に示した.チャンバーと細胞培養面の間に,高さh = 0.3 mm
,幅b = 20
mm
の流路ができており,このスペースを培養液が流れることで細胞にせん断応 力が負荷される.この時の培養液流れはポアズイユ流れを想定した.(a) (b)
Fig. 3-2
フローチャンバー概要図(a)
フローチャンバの寸法(b)
フローチャンバーとディッシュの装着(c)
フローチャンバー装着後の装置全体断面図(c)
31
作製したフローチャンバーによって発生するせん断応力は
Navier-Stokes
の運 動方程式から求められ,以下の式となる.計算式の詳細は付録に記載する.τ = 6𝜇𝑄
ℎ
3𝑏 (3-1)
ただし,各物性値は
µ:培養液の粘性係数;0.85 × 10
-3Pa・S h:
流路の高さ;0.3 × 10-3m
b
:流路の幅;20 × 10
-3m
となっている.第
3
章の実験では,負荷するせん断応力の大きさを生体内と同じ0.6 Pa
とす る.したがって,流量Q
は0.212 × 10
-6m
3/ s
となった.またこの時の流速
V
は𝑉 = 𝑄
𝑏ℎ (3-2)
で求まり,
0.6 Pa
のときは0.353 m / s
となった.このときレイノルズ数Re
はRe = 𝑉ℎ
𝜈 (3-3)
で表され,この時
ν
は動粘性係数(0.85 × 10-6m
2/ s)であり,0.6 Pa
のレイノル ズ数は125
となった.一般的にレイノルズ数Re
が2,000
を超えると乱流になる といわれている.したがって本実験に用いた装置は層流であった.3.2.5 実験装置の全体図
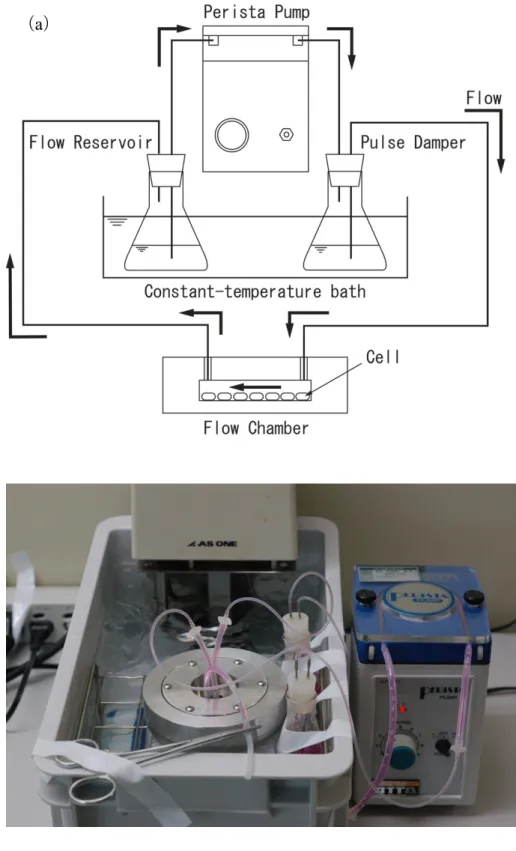
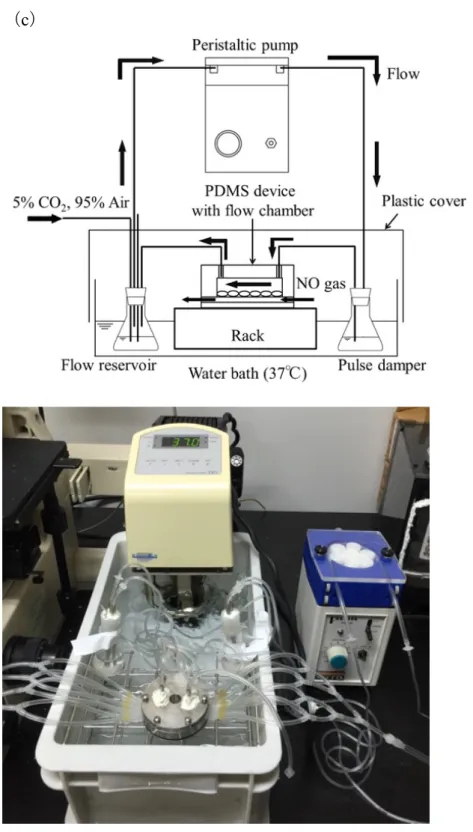
せん断応力負荷回路は細胞培養ディッシュを組み込んだチャンバーと,リザ ーバー,パルスダンパー,ペリスタポンプをそれぞれシリコンチューブで連結 して構成した (Fig. 3-3).流量はペリスタポンプの出力により調節した.パルス ダンパーはペリスタポンプによる拍動を抑えるために用いた.細胞を適切な温 度に維持するため,灌流液は
37
℃の恒温槽で温めた.また,静置培養はフロー チャンバーを組み込まず,インキュベーター内で実験を行った.静置培養,せん断応力負荷実験共に実験期間を
24
時間とし,実験後培養液を 回収し肝細胞機能の変化を測定した.33
(
a
)Fig. 3-3
実験装置全体図(a) 模式図
(