リハビリテーションへの応用を見据えた 脳 - 身体モデルの歩行シミュレーション研究
市村大輔
電気通信大学 大学院 情報理工学研究科 博士 ( 工学 ) の学位申請書論文
2020 年 3 月
リハビリテーションへの応用を見据えた 脳 - 身体モデルの歩行シミュレーション研究
博士論文審査委員会
情報・ネットワーク工学専攻 准教授 山﨑 匡 ( 主査 ) 情報・ネットワーク工学専攻 教授 山本 野人
機械知能システム学専攻 教授 阪口 豊 情報・ネットワーク工学専攻 准教授 龍野 智哉 首都大学東京 システムデザイン研究科
機械システム工学域 教授 長谷 和徳
著作権所有者
市村大輔
2020 年
Simulation study on human bipedal locomotion by a neuro-musculoskeletal model and its application to
rehabilitation Daisuke Ichimura
Abstract
Along with the population aging, the number of patients who need rehabilitation is increasing in many countries. Because the medical conditions and symptoms are different across individuals, therapists have to try various rehabilitation methods until they find an effective treatment for a specific patient. In other words, there are no standard methods in the field of rehabilitation. If we can eliminate the painful trial-and-error processes that coulde take very long time, it will be beneficial for the patients. A potential way to do this is to use computer simulation.
In this study, we focused on bipedal locomotion, that is one of the basic daily activities and its disorder requires rehabilitations. We built two mathematical models for bipedal locomotion, which consists of a neural network model of the brain and a musculoskeletal model of the body. Eventually, we succeeded in simulating bipedal locomotion models for cerebellar injury and hemiplegia after stroke.
In cerebellar injured model, during the initial phase of walking, the cerebellum learned the timing of foot contact on the ground, and after that stable bipedal lo- comotion was realized. We observed that when one hemisphere of the cerebellum was removed, the foot at the injured side was lifted higher than that at the other side. This abnormal foot lifting is consistent with a case study in cerebellar injured patients. This result suggests that the cerebellum plays an essential role in smooth gait control.
Hemiplegia model succeeded in acquiring a stable locomotion. Then, we weakened neural inputs to the muscles on one side leg to simulate a stroke condition. Immedi- ately after the simulated hemiplegia, the model failed to walk. We refitted feedback parameters of reflex or rhythmic locomotion controller to compensate the locomotion.
Stable locomotion was recovered in both conditions. Only in the latter, the neural ac- tivities were abnormal which is similar to chronic stroke patients. This result implies that chronic stroke patients might alter parameters of rhythmic locomotion controller to compensate locomotion.
These results suggest that our simplified brain-body models could simulate patho- logical locomotion in abnormal conditions, therefore computer simulations of rehabil- itations will be possible in future.
概要
超高齢社会において,リハビリテーション (以下リハビリ) は,運動機能の回復のみならず, 認知機能の改善にも効果が期待されている,重要な医療行為である.リハビリは基本的に セラピストの知識や経験に基づく試行錯誤の連続であり,患者の個別因子が強く定量的な 効果測定が難しいため,具体的な方法論が確立されていない.この問題を解決するために は,様々な病態を再現できる数理モデルによる理論的なアプローチが有効であると考えら れる.本研究では脳の神経回路モデルと身体の筋骨格系モデルを統合した脳-身体モデルを 構築し,リハビリを最も必要とする動作の一つである2足歩行に適用した.具体的には小 脳損傷患者の特異的な歩行と,脳梗塞患者の代償歩行をそれぞれ再現することに成功した. さらに,関節トルクやフィードバックパラメータの違いなどで歩行の可否を確認し, それ らの知見を基に臨床現場への提言をした.このような数理モデルによる検証は複雑な病的 歩行のメカニズムの解明,ひいては効果的なリハビリ手法の提案に寄与しうるものである.
目次
第1章 序論 8
1.1 本論文の構成 . . . 10
第2章 基礎知識と手法 11 2.1 歩行の基礎的分類 . . . 11
2.2 生理学的背景 . . . 15
2.3 歩行モデルとシミュレーション手法 . . . 24
第3章 小脳による足底接地情報の遅れ補償を組み込んだ下肢筋骨格系モデルの 歩行シミュレーション 39 3.1 はじめに . . . 39
3.2 方法 . . . 40
3.3 シミュレーション結果 . . . 47
3.4 考察 . . . 52
3.5 むすび . . . 55
第4章 病的状態が2足歩行モデルのmotor moduleに与える影響 56 4.1 はじめに . . . 56
4.2 方法 . . . 57
4.3 結果 . . . 64
4.4 考察 . . . 72
4.5 結論 . . . 76
第5章 総括 77
付録 82
参考文献 97
第 1 章
序論
国連や世界保健機構(WHO)では,高齢化率が7%を超えた社会を「高齢化社会」, 14%
を超えると「高齢社会」, 21%以上では「超高齢社会」と定義している[1]. 2015年に日本 の高齢化率は26%を超えた.また世界的にも今後増加の一途を辿ると見込まれている[2].
こうした高齢者の増加により必然的にリハビリテーション(以下リハビリ)に対するニー ズは高くなっている.特に日本では, 2025年には団塊の世代の全員が75歳以上 (後期高齢 者)に到達することにより, 社会保障費の急増が懸念されるなど超高齢社会への対策は急 務となっている. この2025年問題に対し,厚生労働省は地域包括ケアシステムを立ち上 げ, “高齢者の尊厳の保持と自立生活の支援の目的のもとで,可能な限り住み慣れた地域で, 自分らしい暮らしを人生の最期まで続けることができるよう,地域の包括的な支援・サー ビス提供体制(地域包括ケアシステム)の構築を推進する”ことを掲げている [3]. 地域包 括ケアシステムの構成要素の一つとして,専門的なリハビリが含まれており,リハビリに 対し今後, 社会的な要求が増えていくと予想される.
特にリハビリと関係が深い脳卒中をはじめとする神経損傷患者はその症状が多彩であ り,歴史的に病因–症状関係が調べられてきた[4]. 例えば, 1800年代の終わりから1900年 代に活躍した神経内科医のバビンスキーは数多くの神経徴候を報告し, 現在でも検査・評 価として臨床上使用されている. しかし,彼は診断には非常に熱心であったが治療には関 心がなかったという [5]. これを裏付けるように当時は病因–症状関係を記述する神経症候 学は非常に発展したが, 神経治療学はそれに比べてかなり遅れていた.診断をつけられて も治療法がなく, あったとしても,慢性的な脊髄変性疾患 (脊髄癆)患者に対して行われた 宙吊り療法のようなものが多かった [5, 6]. 宙吊り療法は当時は著効するとされ,多くの患 者が時間をかけ,激痛に耐えこの治療法を施されたが, 現在ではまったく効果がないと実 証され,この治療法は葬られた. このように治療効果の判定は難しく葬られた治療法や薬
剤は少なくない [5]. 医学は日進月歩であるがそのスピードは分野によって差があり,時に は患者に不要な身体的・精神的な苦痛を与えてきた.
このような歴史的な背景もあり,今日では科学的根拠に基づく医療が重視されEvidence Based Medicine (EBM)という理念が掲げられている [7, 8]. しかし,リハビリに目を向 けてみると,必ずしもEBMが確立されているとは言い難い. 脳卒中患者などに対しての 神経系のリハビリは脳科学の基礎研究を基に飛躍的な発展をしているが [9],具体的なリ ハビリ法の提案まではできていない [10]. その要因の一つとして,リハビリの対象となる 患者は再発や合併症などによる複合的な症状を有することが挙げられる. 脳卒中片麻痺の 回復度合いを調べた研究では重症度に関係なく麻痺は回復していくが,そのばらつきは大
きい [11].個人差による影響が著しく強いことを示している. このような個人差を包括し
たEBMの構築が必要であるが,臨床研究のみでは同一の症状を呈する患者を集めること や医師や看護師などが複合的に介入するため, どの治療効果なのか見極めることが非常に 難しい. そのため臨床ではセラピスト個人の知識や経験によってリハビリを提供せざるを 得ない. また臨床現場では,様々なリハビリ手技が提案されている一方,特定のリハビリ手 技が他の治療法と比べ有意差をもって効果があるという報告はされていない [12]. セラピ ストは様々な手法を試してみて効果がありそうなことを経験的に学び,それを患者に提供 していく. この過程は時に長い時間がかかり,また患者に対して苦痛を強いることもある. すなわちリハビリは未だ試行錯誤の連続を抜け出せない.
以上より,より効果的かつ早期に患者の社会復帰や Quality of Life (QoL) を向上させ る, 質の高いリハビリとして,患者個人の特徴や症状に応じたリハビリを提供することが 強く求められている [13]. これを実現させうる方法の一つとして,数値シミュレーション を用い患者個人の病態を再現し, その動作解析を行うことを考えた.
従来より脳や神経系などの複雑なシステムを理解するためには実験的アプローチのみ では不十分であり, 理論やモデルに基づく研究が必要であるとされてきた. 計算機を主な 手段として, シミュレーションなどを用い神経科学を研究するものを計算論的神経科学 (Computational Neuroscience)と呼び,さらに川人は「脳の機能を,その機能を脳と同じ 方法で実現できる計算機のプログラムあるいは人工的な機械を作れる程度に,深く本質的 に理解することを目指すアプローチ」と定義した [14].この考え方はリハビリの分野でも 有効であると考えられ,瀧山は計算論からリハビリにアプローチする有用性を3つ主張し ている [15].
1. モデル上での破壊実験が行えること
2. トレーニング・リハビリ効果の実験が行えること
3. 個人差(パラメータ)の議論が出来ること
これらのことから,理論やモデルによるアプローチはリハビリ分野を大きく飛躍させる方 法論の一つであると考えられる. リハビリの試行錯誤を計算機上でシミュレートし,実際 の患者に適用する前にリハビリの影響・効果を検証・予測することができれば,患者の負 担を大幅に削減することができる. 実際に,運動プリミティブモデルを応用し,脳卒中モデ ルを構築しており,腕のリハビリの有用性を計算機上で検証している [15].
本研究では,脳の神経回路モデルと身体の筋骨格系モデルを統合した脳-身体モデルを構 築し, リハビリ応用を見据えた病的2足歩行シミュレーションを実装することを目的とす る. 具体的には小脳損傷患者の特異的な歩行と,脳梗塞患者の代償歩行をそれぞれ再現す ることを試みた. さらに,関節トルクやフィードバックパラメータの違いなどで歩行の変 化を確認し, それらの知見を基に臨床現場への応用を検討する. 最後に,得られた結果につ いてまとめ,今後の展望を述べる.
1.1 本論文の構成
第1章(本章)では,論文全体に関わる研究の背景,目的について述べた.
第2章では,歩行に関わる生理学的な知識とモデルリング・シミュレーション手法を述 べる.
第3章では,小脳モデルと歩行モデルを接続し,小脳の足底接地のタイミング予測が2 足歩行にどのように影響を与えうるのかを,数値シミュレーションで検討する.
第4章では, motor module制御の 2足歩行モデルを構築し, motor module 変化と脳 梗塞患者の歩行に対して類似性を述べる.
第5章では,得られた結果についてまとめる.
第 2 章
基礎知識と手法
2.1 歩行の基礎的分類
2.1.1 歩行周期
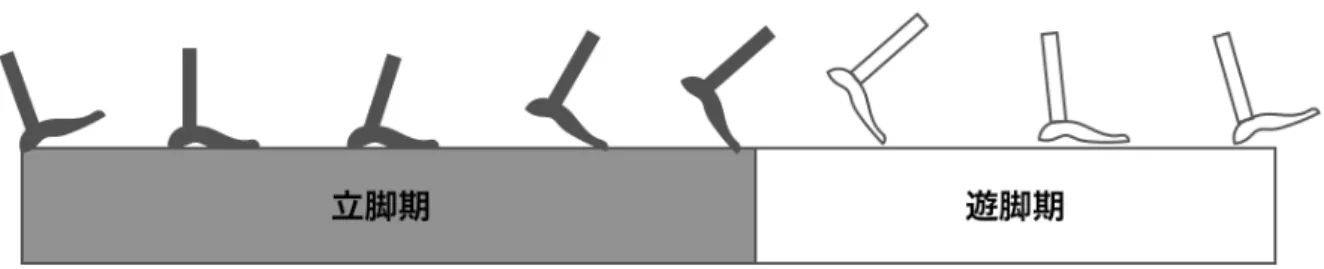
歩行は周期的な動作で身体を前進させる運動である. 一側下肢が持ち上がり新たな支持 面に移動する間,反対側は足を地面に付けて支持することで身体は前進する. その後,下 肢の役割は逆になる.一側下肢から反対側下肢へ重心を移動させるときには, 両足部は地 面に接地している. 一側下肢におけるこの一連の動作は歩行周期という[16, 17]. 一般的に 床接地が歩行周期の開始点とされており,踵接地または初期接地と呼ばれる.
歩行周期の区分
歩行周期は,立脚期と遊脚期の2つの期間に分けられる (図2.1). 立脚期は,足部が接 地しているすべての期間を示す. 立脚期は初期接地から開始され,遊脚期は下肢を前進さ せるために足部が空中にある期間を示す. 遊脚期は,足部が床から持ち上げられるとき (足指離地)から開始される.
立脚期はさらに3つの期間に分けられる. 立脚期の始まりと終わりは,両下肢が接地 している期間(初期両下肢支持期と終期両下肢支持期)を含んでいる. 立脚期の中間部分 は一側下肢のみが接地している(単下肢支持期).
初期両下肢支持は歩行周期の開始であり,初期接地によって両下肢が接地する期間を示 す. 反対側下肢が振り出され,単下肢支持期が始まる. 単下肢支持期の間,全体重は足部 で支持している. 単下肢支持持続時間の増加が安定性の増加を反映すると考えられるの で,下肢の支持能力を評価するもっともよい指標となる.
図2.1 歩行周期の区分.灰色は立脚期の期間,白色は遊脚期の期間を示す. 左から右へ の歩行を表しており,立脚期は初期接地から足指離地まで, 遊脚期は下肢を空中に降り 出してから再び接地するまでの期間である. [16]の図を参考に作成.
終期両下肢支持は,反対側の接地が起こり (反対側の初期接地),そして最初の立脚側下 肢が振り出されるまで (同側の足指離地)続く.
歩行周期タイミング
歩行周期は一般的に,立脚期が約60%,遊脚期が約40%の期間を占めるが, これらの期 間は歩行速度によって異なる. 歩行速度が増加すると立脚期と遊脚期の持続時間の合計は 短縮する. 速度が落ちるにつれて,立脚期と遊脚期の持続時間は次第に長くなる. 立脚期 に関しては,速く歩くと単下肢支持期は長くなり,両下肢支持の期間が短くなる. 逆に,歩 行速度が落ちれば単下肢支持期は短く,両下肢支持が長くなる.
両下肢が支持の役割を交換する両下肢支持期があることは, 歩行の基本的特徴である. 両下肢支持がなくなると,人の移動は走行状態となる.
重複歩
歩行周期は重複歩とも呼ばれる(図2.2). 重複歩は一側下肢の動作に基づいたものであ る. 重複歩の期間は,同側下肢による連続した2回の床接地の間である(例えば,左初期接 地から次の左初期接地). また,重複歩と混合しやすいステップというものがある. ステッ プは両下肢の距離を表す(図2.2). 重複歩(歩行周期)にはステップが2回あり,重複歩の 中間点で反対側の下肢は地面に接地し,立脚期を開始する.
図2.2 重複歩とステップ.左下肢の接地から次の接地までが重複歩であり,右のステッ プは左接地から右接地の距離である. [16]の図を参考に作成。
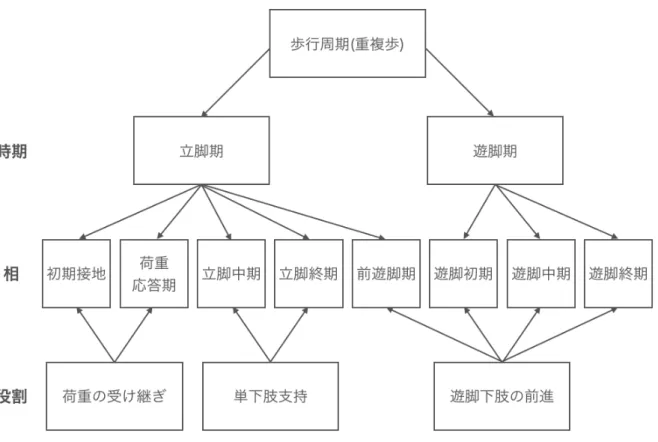
2.1.2 歩行の相
相による歩行分析は,各関節で起こる種々の動作の機能的な意義を評価する. 歩行を相 に分けて分析することで,各関節で同時に起こる運動を全体の下肢機能に関係づけて考え ることができるようになる. これは,能力低下の機能的な影響を解釈するための重要なア プローチである. 1つの歩行相に必要な機能はそれぞれ異なっているため, 1つの歩行相に おいては適切な姿勢でも,別の時点では異常姿勢となる可能性がある. したがって,タイミ ングと関節の角度は非常に重要である. 相の区分を図2.3に示す. ここでは8相に分類さ れ, 機能的な役割として,荷重の受け継ぎ,単下肢支持,遊脚下肢の前進がある.
図2.3 歩行周期の機能的な区分. 時期は,足部接地の有無によって歩行周期の基本的 な区分を示す. 各相は下肢の位置で決められる. 役割は,貢献する機能によって相のグ ループ分けを示す. [16]の図を改変し作成.
荷重の受け継ぎ
荷重の受け継ぎは,立脚期における最初の課題である. これは, 1.衝撃吸収, 2.初期の下 肢の安定性, 3.前進の維持,という3つの機能的な要求が満たされなければならない. 遊 脚期が終わったばかりの不安定な下肢へ急速に体重を移動させなくてはならない. 初期接 地と荷重応答期がこの時期に含まれる.
単下肢支持
反対側下肢が遊脚期を開始すると同時に単下肢支持での立脚期が開始され,これは反対 側下肢が再び床に接地するまで継続する. その間,体重を支持し前進し続ける. 立脚中期 と立脚終期がこれに含まれる.
遊脚下肢の前進
下肢を前進させるという重要な役割を果たすため,立脚期の間から準備姿勢を始める (前遊脚期). その後下肢自体を持ち上げ, 3つの遊脚の相を経て前進して(重複歩距離を完 成させ),次の立脚期の期間に備える. 遊脚下肢の前進には,前遊脚期,遊脚初期,遊脚中 期,遊脚終期の4つの相が含まれる.
2.2 生理学的背景
2.2.1 歩行に関わる中枢神経領域
中枢神経系は基本的に左右対称であり, 2つの主要な部分,脊髄と脳からなる (図2.4).
歩行には多くの中枢神経系が関わるとされるが, その中でも主要な領域について述べ る[18].
前頭葉
頭頂葉
後頭葉 小脳 脳幹
脊髄 大脳
(中脳, 橋, 延髄) 側頭葉
上位中枢
下位中枢
図2.4 運動機能に関わる中枢神経系.中枢神経系は左右対称であり,これは左側から見 た図である.脳は大脳,小脳,脳幹に大きく分けられる.また,大脳の表面は大脳皮質と呼 ばれ,前頭葉,頭頂葉,後頭葉,側頭葉に分けられる. 脳幹は中脳,橋,延髄からなる.
大脳
2つの半球からなり,深くシワの入った外層 (大脳皮質)と深部にある3つの構造物(基 底核, 海馬,扇桃体)が各半球にある. 主に,基底核は運動の制御に関与する. 海馬は記憶の 貯蔵に関係している. 扇桃体は,情動に関係した自律反応と内分泌反応を協調させる. ま た,大脳皮質は4つの脳葉 (前頭葉,頭頂葉,後頭葉,側頭葉)に分けられる. 各脳葉には専 門とする機能がある.前頭葉は主に,短期記憶と将来の行動計画,運動の制御にかかわっ ている. 頭頂葉は,体性感覚,身体イメージの形成,身体イメージの外界との関連づけに 関係している. 後頭葉は視覚をつかさどる. 側頭葉は聴覚をつかさどるとともに,深部に ある海馬と扇桃体を介して学習,記憶,感情にかかわっている.
各大脳半球は主に体の対側の感覚や運動に関与する. 左半身から脊髄に到達した感覚情 報は,右側の神経経路へ交差して大脳皮質に到達する. 同様に右大脳半球の運動野は,左 半身の運動を制御する.
小脳
小脳はヒトでは大脳の背面下部,橋に覆い被さるように位置している. 小脳は運動出力 の細かい調整や運動学習にも関わり,姿勢の維持や頭部,眼球,腕の動きの協調に重要な 役割を果たす. 小脳には大脳半球を含め脳のどの下位区分よりもニューロンの数が多い. しかし,ニューロンの種類は比較的限られているため,内部回路はよく解明されている (図2.5).
小脳回路への入力は橋核などからの苔状線維入力と下オリーブ核からの登上線維入力の 2つがある. 苔状線維は顆粒細胞の樹上突起に興奮性シナプスを作り,その後平行線維を 経由してプルキンエ細胞に興奮性の情報を伝える. 苔状線維系は末梢からの刺激や中枢神 経で生成された行動に関する信号の大きさと持続時間を符号化している. また顆粒細胞は ゴルジ細胞を興奮させ,ゴルジ細胞は顆粒細胞を抑制する再帰型抑制ネットワークである. 抑制性介在ニューロンは平行線維から興奮性入力を受け,プルキンエ細胞を抑制する. 一 方,登上線維は下オリーブ核からなり,末梢と大脳皮質からの情報を小脳へ送る. 低い頻度 でしか発火せず,特異的な情報が来たときのみ発火する.
平行線維と登上線維の同時刺激により長期抑圧(long-term-depression; LTD)と呼ばれ る学習が起こる. この長期抑圧の役割は結果として運動の修正・調整することになる. 運 動が不正確のときに登上線維は教師信号に応答し, その教師信号が生じた運動に関係する 平行線維–プルキンエ線維は抑圧されることになる. 運動を繰り返すと,好ましくない運動 指令を伝える平行線維入力はますます抑えられ,状況に適したものになる. 最終的には登
上線維系で伝えられるエラー信号がなくなり,運動学習が完了する. これにより環境に適 した強度・タイミングで運動制御が可能となる.
CN
PN IO
プルキンエ 細胞
橋核 苔状線維
小脳核
登上線維
下オリーブ核 ゴルジ細胞
分子層介在 ニューロン 顆粒細胞
平行線維
入力 抑制性 出力 入力
興奮性
図2.5 小脳神経回路の模式図.外部からの信号は橋核と下オリーブ核の2カ所から入 る. 出力は小脳核で行う.橋核からの入力は苔状線維を介して顆粒細胞へ伝わる. 顆粒 細胞は平行線維を介してゴルジ細胞,分子層介在ニューロン,プルキンエ細胞を興奮さ せる. また,これらの細胞は顆粒細胞,プルキンエ細胞,小脳核をそれぞれ抑制する. 下 オリーブ核は教師信号を入力とし,登上線維を介してプルキンエ細胞を興奮させる.
脳幹
脳幹は延髄,橋,中脳からなる.脳幹は皮膚や筋肉からの感覚情報を受け取り,筋系に運 動の指令を与える. また,脊髄からの情報を脳に伝達し,脳からの情報を脊髄に伝達する. さらに,網様体を介して覚醒状態を制御する. 脳幹には神経細胞体の集合がいくつかあり,
これらは脳神経核と呼ばれる. 聴覚,平衡感覚,味覚という3つの特殊感覚からの情報を 処理することに特化した脳神経核などがある. 脳幹にも歩行誘発野があるとことが知られ ている. 延髄は脊髄のすぐ吻側の部位で,消化,呼吸,心拍数調節といった重要な自律機 能を担う中枢が複数ある. 橋は延髄の吻側にあり,大脳からの運動に関する情報を小脳に 伝達する. 中脳は橋の吻側にあり,眼の動きや視覚・聴覚反射の協調など,多くの運動・
感覚機能を制御している.
脊髄
脊髄は中枢神経系の中で最も尾側に位置している. 頭蓋底から第1腰惟までのびてい る.脊髄は体幹および四肢の皮膚や関節,筋肉から感覚情報を受け取り,また随意および
反射運動の両方に携わる運動ニューロンを収容している. 歩行のパターンを生み出す中枢 パターン発生器 (Central Pattern Generator; CPG)があることが知られている. 脊髄は 長軸方向に沿って大きさと形が変化しており,これは,四肢や体幹を支配する運動神経が あるかどうかに依存する. 腕や脚を神経支配する位置では厚みを増す.
2.2.2 歩行の神経制御
歩行制御の階層性
ヒトの直立2足歩行の詳細なメカニズムは未だよくわかっていないが, 動物実験やヒト を対象にした電気生理学実験などから様々なことが示唆されている.歩行は外部環境に応 じた適切な調整が必要であり, 脳神経系全般が関与していることが知られている. 特に意 図的な歩行動作は上位中枢系である大脳皮質の関与が強く,それに加えて, 小脳,脳幹部な どが調整系の役割を担う. 下位の中枢系では意図の関与は少なくなり,最終的には脊髄の CPGが上位中枢からの入力により, 歩行リズムパターンを形成していると考えられてい る[19].
上位中枢は大脳運動皮質や小脳をはじめとする脳の多くの領域であるが,歩行において は, 以下のような大きく3つ機能に分類できるとされる[18].
a). 脊髄の歩行中枢を活性化し,歩行を開始させその速度を調整するもの b). 肢からのフィードバック等を受けて歩行を調整するもの
c). 視覚情報をもとに肢の運びを舵取りするもの
a). に関しては歩行誘発野として知られており, 脳幹などに存在していることが確かめら
れている [20]. この歩行誘発野に電気刺激を与えると,刺激パターンとの関連はなく,強度
に依存し歩行を開始し, さらに速度も変化させれることが知られている. b). は主に小脳 が担っていると言われている. 小脳は歩行中,脊髄の CPGや体性感覚系の受容器からの 情報を常時受けている. これらの情報は, ともに苔状線維系として小脳に送られ,顆粒細 胞–平行線維を経由してプルキンエ細胞に伝達される.プルキンエ細胞はこの苔状線維系の 入力を小脳皮質からの出力に変換し, その出力は,小脳核から脳幹–脊髄投射系を介して脊 髄内のニューロン活動をオンラインで調整する (図2.6). 教師信号を用いて長期抑圧が引 き起こされ,制御不良を引き起こすような平行線維–プルキンエ細胞間のシナプス活動を 学習し,調整していくことによって適応的なフィードフォワード制御に貢献していると推 察される. 小脳は歩行における下肢協調を多様に変化する外部環境の中で適応的に制御す るのに重要であると考えられている [21, 22]. c). に関しては後頭葉皮質がその役割を担っ
ていると考えられている. 歩いてる途中に障害物があれば,乗り越えるか迂回しなければ ならない. 四足動物では後肢が乗り越える時には,障害物が視野からなくなっているので, ワーキングメモリーと呼ばれる短期記憶に視野情報を貯蔵し,脚の動きを誘導する. しか し, 後頭葉損傷を受けたネコでは歩行中に障害物をうまく避けきれなくなってしまう [23].
大脳
脊髄
運動系 脳幹
CPG
小脳
体性感覚・CPG情報 教師信号
運動の調整指令
図2.6 小脳と脊髄を中心とする歩行制御回路. 小脳は脳幹,脊髄,大脳との間でネット ワークを構成している. 脳幹,脊髄からの体性感覚情報などを受けとり,姿勢制御や四肢 の運動を調整している.また教師信号を受け取り適応・学習機構として働いている. [22]
の図を参考にし作成.
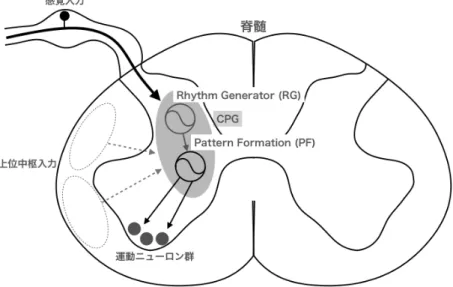
下位の中枢系である脊髄にはCPGがあると考えれらている. 上位中枢や末梢感覚器官 からの入力がなくても, CPG は歩行運動様のにパターン化した交代性の屈筋–伸筋運動 ニューロンの活動のみは生成できる. 上位中枢や末梢からの感覚入力により, CPGは制御 または修飾され,歩行が実現されていると考えられている. CPGモデルとして, 1910年
代にGraham Brownは屈筋と伸筋の収縮は相互抑制する2つのニューロン (ハーフセン
ター)によって制御されているという仮説を提唱し, Anders Lundbergによって継承され
てきた [24]. 2つのハーフセンターは抑制性の結合が徐々に弱まることによって交代制に
活動する. 例えば, 2つのハーフセンターが同時に興奮性入力を受けた場合,屈筋のハーフ センターへの入力が強ければ,屈筋は収縮し,伸筋のハーフセンターは抑制される. そし
て,屈筋からの抑制結合が弱まると伸筋のハーフセンターの出力が大きくなり,今度は屈筋 が抑制され,この抑制結合が弱くなるまで伸筋は収縮する. このハーフセンター仮説は歩 行運動時の交代運動を合理的に説明可能であるが,筋活動と歩行リズムを独立に制御でき ない.しかし,歩行動作では上り坂や下り坂など機能的な柔軟性が求められる. Rybakら (2015)は除脳ネコの実験結果より,タイムキーパーを担うRhythm Generator (RG)ネッ トワークと運動神経群の活動を支配するPattern Formation (PF)ネットワークから成る 2階層CPGモデルを提唱している(図2.7) [25]. このモデルでは, 歩行周期のタイミング や運動神経の活動を独立に制御することができるため, より複雑な運動も説明可能である.
図 2.7 脊髄内の CPG 概略図. Rhythm Generator (RG) とPattern Formation (PF)の2階層ネットワークで構成される. RGがPFに働き,運動ニューロン群の活 動を生成する.上位中枢入力や感覚入力はCPG活動を調整する. [26]の図を参考にし て作成.
ヒトにおけるCPGの存在
このようなCPGに関する研究は四足歩行をするネコなどの動物実験が多いが, ヒトで もその存在は示唆されている. ヒトの新生児は,足を地面につけ身体を保持し,水平に動か すと左右の足に自動的な歩行運動が起こることが知られている (足踏み反射). これは,ヒ トが歩行のための基本的な神経回路が生まれる前から備わっていることを示唆している. またこの反応は大脳を欠損している無脳症の新生児でも観察されることから,脳幹よりも 下部, おそらく脊髄に神経回路があると考えられる [18]. Calancieら (1994)は慢性期の
脊髄損傷患者で,歩行訓練後,歩行に似た下肢のステッピング動作が自分の意志とは無関 係に生じる症例を報告している [27]. これは何らかの原因で脊髄のCPGが活性化されリ ズミックな活動が生じたものと考えられた. Dimitrijevicら (1998)は脊髄損傷患者の脊 髄に電極を挿入し,硬膜外刺激を行うことで歩行様の運動を誘発することに成功した [28].
この結果は脊髄の局所的な部分にCPG が構成されていることを示唆している. 健常者 に対しては,腰部脊髄への磁気刺激により歩行様の運動を誘発できることが報告されてい
る [29]. これらはヒトCPGの存在を強く示唆するものである. さらに,筋電図を用いた
研究では,歩行中に筋は協調して活動するいくつかのモジュール構造があることが報告さ
れている [30–34]. 筋電図は中枢神経系からの刺激を,筋を通して計測するものであり, 中
枢神経系がモジュール構造を用いて歩行運動を生成していることを示唆し, その役割を CPGが担っていると考えられている. このモジュール構造はmotor module (または筋シ ナジー)と呼ばれる. 運動制御ではmotor moduleを用いることで制御すべき自由度を低 次元化していると考えられている. また, motor moduleは神経系の状態を反映している と指摘され,病態の程度を表す指標としても応用が期待されている [35].
図2.8 Motor moduleの概念図.計測された筋活動は筋の重みと,それに対応した時 間パターンに分解できる.中枢神経から脊髄を経て,各筋肉をモジュール化して制御す る. [34]の図を参考にして作成.
2.2.3 神経損傷患者の歩行
歩行障害の要因として,脳,脊髄,骨・関節,末梢神経,筋肉,血管とあらゆる部位の病変 で生じる可能性があり,特に神経損傷患者の場合は前述した神経系の一部の機能欠損だけ では説明できないとされている. それは,単に機能欠落によるものだけでなく,それに応じ た神経再生または代償を伴うものもあるからである. すなわち,歩行に関する健全な神経 系の機能不全の他に,本来持たなかった機能追加 (誤った方向のもを含め)によって,歩行 障害の病態機序は著しく複雑化している [36]. そのため,神経損傷患者の歩行障害につい て不明な点が多い. 歩行障害とそれをもたらす病因がある程度対応しているものとして, 小脳障害,片麻痺,対麻痺,パーキンソン病,下肢筋脱力,多発硬化症などが知られる [36].
今回, 3章と4章では臨床上特徴的な小脳障害歩行と罹患率の高い脳卒中片麻痺歩行をそ れぞれシミュレートした. これらの神経学的な観察による歩様と定量的な歩行評価につい
て概説する.
小脳障害歩行
両脚を開いた開脚歩行と一歩一歩が不安定な歩行から成る. 病状が比較的軽い時は開脚 性にとどまるが,重くなると不安定性が強くなり,遂には歩行不能になることもある. 片側 性小脳障害では患側下肢を高くかつ,前外方に投げ出すように蹴り上げ, 反動的に足を地 面に落とすように下ろす [37]. 3次元歩行解析を用いた定量的な歩行評価では,小脳損傷 患者では肢間協調の障害が生じ, 歩行が不安定になることを報告している [38]. 筋電図を 用いた研究でも,健常者と比べ小脳損傷患者では筋の働くタイミングが変化し, 歩行の不 安定性を示している [39]. また,左右のベルトが異なる速さで動くスプリットトレッドミ ルを用い, 歩行中,健常者では外乱適応するが小脳損傷患者ではそれが生じないことを報 告している [40].
脳卒中片麻痺歩行
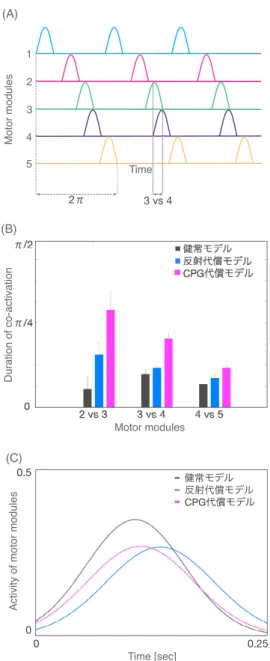
筋緊張が低い弛緩性の場合と筋緊張が高い痙性の場合の 2つに大別される. 弛緩性歩 行は健側下肢を速めに移動し,患側下肢に体重をかける時間を短くする. 患側下肢を前進 させる時には,重みのある物体を引きずるように足底が地に着いたまま, 下肢を前に引き 寄せる.この間,足は前方を向かずに外旋し,最後に腰を回すようにして, 患側下肢を前方 に出す.引きずり歩行 (dragging gait)とも呼ばれる. 一方, 痙性歩行は患側下肢は膝が 伸展し,下肢全体が棒状になるので,足を前進させるのに, 股関節を中心に振り回すよう になる.草刈り歩行または円描き歩行と言われている. 定量的な歩行評価も報告されてい る. 3次元歩行解析を用いた報告では,歩行速度が遅い痙性片麻痺歩行では膝の動きから パターン分類できるとしている [41]. 筋電図を使用した研究では,数理的処理を施すこと
でmotor moduleの時空間構造が調べられており,亜急性期患者では健常者と類似した
motor moduleであるが [42], 慢性期脳卒中患者ではmotor moduleのいくつかは統合さ れ,数が減ってしまうことが報告されている [43].
パーキンソン病歩行
歩幅が短くなる小刻み歩行,一歩目を踏み出すと両足を交互に運んで, 歩行速度が加速 する突進現象,歩行中,急に足底が地面にへばりついたように足が前進できなくなるすく み足歩行などの特徴をもつ.
腓骨神経障害歩行
足を前進させる時に,足先が下垂し地面に引っかかるので,その分膝を高く上げる. その 結果,一歩ごとに膝を高く挙上し,足先が垂れた歩行になる. 足を降ろす時,踵から着地せ ず,地面を叩くように足先から下りる.
3章では両側小脳損傷と片側小脳損傷を,4章では脳卒中片麻痺歩行をそれぞれシミュ レートした.
2.3 歩行モデルとシミュレーション手法
2.3.1 骨格モデル
骨格モデルの概要
1章で述べたように,ヒトの歩行運動は様々な要因から成り立ち,歩行障害を呈すると, さらに複雑化されてしまう.また,個人差も大きく実験・計測による研究だけでは障害の理 解やリハビリへの示唆は難しい. そこで様々な条件を容易に検討でき, 関節トルクや筋張 力から予測的・統合的に運動パターンを生成できる順動力学モデルを用いた.
様々な筋骨格系順動力学モデルが報告されているが [44–54], 今回3章では2次元1質 点4リンクのTagaモデル [44]を基に, 4章ではTagaモデルを拡張した2次元7リンク の筋骨格モデルを構築する.筋モデルは後述する.
Tagaモデルは神経振動子 (Matsuokaニューロン)をつなぎ合わせ,歩行のリズムを生
成(CPG)し, さらに外界からのフィードバックを遅れなしでCPGに与え, 2足歩行を実
現させた. しかし,一般的にはフィードバックには遅れが含まれるので3章では足底接地 のフィードバック情報に遅れを付加し,それを小脳モデルで補償するモデルを構築した. 4章で用いた, 2次元7リンクの筋骨格モデルは様々な方法で歩行制御が提案されている が[46, 48, 49, 51, 52],歩行の頑健性を重視したモデルが多く,特異的な歩行障害を表現でき る柔軟なモデルはなかった. 本研究では,フィードバックパラメータによって歩行パター ンが変化する階層性CPGを有する神経筋骨格系モデルを構築し,歩行制御を行った.
運動方程式の導出は両モデルともニュートン・オイラー法を用いた [44, 45]. ニュート ン・オイラー法は各リンクについて力とトルクの釣り合いから運動方程式を導く. 並進運 動をニュートンの運動方程式,回転運動をオイラーの運動方程式によって記述される. 一
般的なニュートンの運動方程式とオイラーの運動方程式は,それぞれ F =m¨x,
N =Iθ,¨ (2.1)
であり, F は力, mは質量, ¨xは加速度を示す. N はトルク, I は慣性モーメント, ¨θ は回 転加速度を表す.
2リンクモデルの例
2リンクモデルで運動方程式を導く. 図2.9のモデルを使用した. ニュートン・オイラー 法(式(2.1))より運動方程式は,
m1x¨1 =−F1+F3,
m1x¨2 =−F2+F4−m1g,
I1θ¨1 =−F1(l1/2) sinθ1+F2(l1/2) cosθ1
−F3(l1/2) sinθ1+F4(l1/2) cosθ1 +Tr1+Tr2,
m2x¨3 =−F3+F5,
m2x¨4 =−F4+F6−m2g,
I2θ¨2 =−F3(l2/2) sinθ2+F4(l2/2) cosθ2
−F5(l2/2) sinθ2+F6(l2/2) cosθ2
−Tr2−Tr3,
(2.2) と表せる. m1, m2 はLink1とLink2の質量, (x1, x2) と(x3, x4) はそれぞれLink1と Link2の座標, Fi (i = 1, ...,6),は各Linkの力であり, gは重力加速度, I1,I2 はLink1と Link2の慣性モーメント, θ1, θ2 はLink1とLink2の角度, l1, l2 はLink1とLink2の長 さ, Tr1,Tr2,Tr3 は外的なトルクを示している.
また, Link1とLink2は結合されているため運動学的な制約をうける. それらは以下の
ような式で表すことができる:
x1+ (l1/2) cosθ1 =x3−(l2/2) cosθ2,
x2−(l1/2) sinθ1 =x4+ (l2/2) sinθ2, (2.3) これを2階微分すると,
¨
x1−(l1/2) sinθ1θ¨1−x¨3−(l2/2) sinθ2θ¨2 = (l1/2) cosθ1θ˙1+ (l2/2) cosθ2θ˙22,
¨
x2−(l1/2) cosθ1θ¨1−x¨4−(l2/2) cosθ2θ¨2 =−(l1/2) sinθ1θ˙1−(l2/2) sinθ2θ˙22, (2.4)
が得られ,式(2.2)を式(2.4)に代入すると,運動学的な制約を受けた運動方程式が得られ る. 同様の手法で3章で用いるTagaモデルと4章で用いる筋骨格系モデル(図2.10)の 運動方程式を導いた.
(x1, x2) θ1 l1
m1
m2
Link1
g
l2
(x3, x4) θ2
Link2
Link1
Link2
F1
F2
F3 F4 m1g Tr1
Tr2 F3
F4
F5 F6 m2g
Tr2
Tr3
(A) (B)
図2.9 2リンクモデル. (A): 2リンクモデルの座標や位置を示す. (x1, x2), (x3, x4) はLink1とLink2の座標,θ1,θ2はLink1とLink2の角度, l1,l2はLink1とLink2 の長さ,m1, m2はLink1とLink2の質量,gは重力加速度を表す. (B): 各リンクにか かる力とトルクを示す. F1, F2, F3, F4は力,Tr1,Tr2,Tr3,Tr4はトルクを表す.
(A)
(x1, x2)
(x3, x4) (x6, x7)
(x12, x13) (x9, x10) x14
x8
x11 x5
大腿
下腿
足部
(xl, yl) (xr, yr) M
(B)
m1
m2 l1
l2
g HAT
大腿 下腿 足部
(x1, x2)x3
x6 x9
x12 x15
x18 x21
(x4, x5) (x7, x8)
(x10, x11) (x13, x14)
(x16, x17) (x19, x20)
(xrt, yrt) (xrh, yrh) (xlt, ylt)
(xlh, ylh)
lHAT
luthigh llthigh
lushank
llshank mHAT
muleg
mlleg lfoot1
lfoot2 lfoot3
mfoot
足部
g
図2.10 使用した筋骨格モデル. (A): 3章で使用したTaga モデル. 2次元1質点4 リンクである. [44]の図を参考に作成. (x1, x2)は右と左の股関節の座標,(x3, x4), (x6, x7)は右と左の大腿中心の座標,(x9, x10),(x12, x13)は右と左の下腿中心の座標,
(xr, yr),(xl, yl)は右と左の足部の座標,x5,x8は右と左の大腿の角度,x11,x14は右 と左の下腿の角度をそれぞれ表している.M, m1, m2は上半身の重さ,大腿の重さ,
下腿の重さをそれぞれ表している. l1,l2は大腿の長さ,下腿の長さをそれぞれ表して いる.(B): 4章で使用した筋骨格モデル. 2次元7リンク, 18筋を有している. 頭部, 上肢,体幹を合わせて1リンク(Head, Arms, and Torso, HAT)とした. 右上部は拡 大した足部を示す. (x1, x2)はHAT中心の座標,(x4, x5),(x7, x8)は右と左の大腿 中心の座標,(x10, x11),(x13, x14)は右と左の下腿中心の座標,(x16, x17),(x19, x20) は右と左の足部中心の座標,(xrh, yrh),(xlh, ylh)は右と左の踵部の座標,(xrt, yrt), (xlt, ylt)は右と左のつま先の座標をそれぞれ表している. x3, x6, x9, x12, x15, x18, x21はそれぞれHATの角度,右と左の大腿の角度,右と左の下腿の角度,右と左の足部 の角度を表している. mHAT, muleg, mlleg, mfootはそれぞれHATの重さ,大腿の重 さ,下腿の重さ,足部の重さを表している. lHAT,luthigh,llthigh,lushank,llshankはそれ ぞれHATの長さ,大腿上部の長さ,大腿下部の長さ,下腿上部の長さ,下腿下部の長さ をそれぞれ表している.lfoot1,lfoot2, lfoot3はそれぞれ足関節から足部中心までの長さ, つま先までの長さ,踵部までの長さを表している.
2.3.2 筋モデル
4章では筋モデルを使用する. 筋張力は神経系からの刺激によって生じるが,この神経 からの刺激と筋張力の変換は単純ではなく,非線形の力学特性を有している. その特性は
筋の張力–長さ関係,筋の張力–速度関係という2つの基本関係で表される(図 2.11). 筋の 張力–長さ関係は筋張力が筋の長さに影響し,筋には最大張力を発揮しうる最適長が存在 することを示している [55]. 筋の張力–速度関係は筋張力が筋収縮の速度に影響し,筋の収 縮速度が速いほど, 筋は張力を発揮しにくい性質を表している [56]. このような筋の力学 特性を含んでいる以下のような筋モデルを使用した. [46, 49, 51, 57].
Fm′ = ¯FmCE·k(ξm)·h(ηm)·αm+FmPD+FmPE,
k(ξm) = 0.32 + 0.71 exp[−1.112(ξm−1)] sin[3.722(ξm−0.656)], h(ηm) = 1 + tanh(3.0ηm),
FmPD =cPDm L˙m,
FmPE =kPEm {exp[15(Lm−L¯m)]−1}.
(2.5) Fm′ はm番目の筋の筋張力, ¯FmCEは収縮要素 (CE)による最大筋張力, k(ξm)は筋の張 力–長さ関係, h(ηm)は筋の張力–速度関係, αm はα運動ニューロンに対応する刺激信号 (0≤ αm ≤ 1), FmPD と FmPE はそれぞれ粘性と弾性力, ξm と ηm はそれぞれ筋の最適長 L¯mによって正規化された筋の長さと収縮速度である. ξm = Lm/L¯m, ηm = ˙Lm/L¯˙m で あり, Lm, ˙Lm, L¯˙mはそれぞれ筋の長さ, 収縮速度,最大の筋収縮速度である. cPDm は粘性 係数, kmPEは弾性係数である. 各パラメータの値を付録A.1に示す.
図2.11 筋の力学特性.左側は筋の張力–長さ関係を示す.中間的な筋の長さでは張力が 最大になる. 右側は筋の張力–速度関係を示す.ピーク張力は短縮速度が増加するにつれ 減少する. 筋活動する時に筋が引き伸ばされると(伸長)ピーク張力は大きくなる. [18]
の図を参考に作成.
2.3.3 CPG モデル
CPGモデルの概要
1章より, CPGは自律的に活動し,様々な入力を受け,制御,修飾される. 本研究では相 互抑制作用により,自律的なリズムパターンが生成できるMatsuokaニューロンモデルを 使用する [58, 59].
2ニューロンでの例
単純な CPGモデルとして周期的に活動する2 つのニューロンを結合させる. そして, その2つのニューロンは互いに抑制をし,また自身への抑制も行い,リズミックな運動を 生成する. このような数理モデルは以下のように表せる.
τu˙1 =−u1−wy2−βv1+u0, τu˙2 =−u2−wy1−βv2+u0, τ′v˙1 =−v1+y1,
τ′v˙2 =−v2+y2,
yi =f(ui) (f(ui) = max(0, ui)) (i= 1,2), (2.6) この時, ui はi番目のニューロンの状態, yi はi番目のニューロンの出力, viはi番目の ニューロンの自身への抑制をそれぞれ示す. また, u0は外部からの入力, wは結合の重み, τ, τ′は時定数を表す. よって, τ, τ′ は周波数をu0 は振幅の度合いを変化させる. 結果を 図2.12に示す.互い違いに活動が現れる.
3章ではこのMatsuokaニューロンモデルを12個使用し, CPGモデルとし, 4章では 14個使用し, Rhythm Generator (RG)ネットワークとPattern Formation (PF)ネット ワークから成る2階層CPGモデルを構築した.
u 1 u 2 y 1 y 2
(A) (B)
図2.12 抑制的に結合させたMatsuokaニューロン. (A): 2ニューロンを抑制的に結 合させた.互いにかつ自身に抑制をかける. (B): 2ニューロンの活動の例.相互にリズ ムパターンが生成される. 横軸は時間,縦軸はニューロンの活動を示す.
2.3.4 小脳モデル
脳はニューロンを素子として大規模なネットワークを築いている. ニューロンの情報 伝達の単位はスパイクと呼ばれ,スパイクを発射することを発火(fire)と言う. ニューロ ンが単位時間あたり, または単位試行あたりにスパイクを生成することを発火率 (firing
rate)と呼ぶ. この発火率をニューロンの伝達信号として考えるようなモデルをレートモ
デルと呼び, 本研究ではレートモデルである小脳内部時計モデルを使用した [60]. このモ デルでは,小脳の顆粒細胞の発火パターンで時間経過を表現していると提案している. こ のモデルは興奮性の苔状線維入力とゴルジ細胞を介した再帰的な抑制性入力の2つをもつ N 個の顆粒細胞,その投射先である1つのプルキンエ細胞,出力細胞である小脳核からな る(図2.5). 顆粒細胞のダイナミクスは以下のように記述される [60]:
zj(t) = [qj(t)]+, qj(t) =I(t)− 1
N
∑′ j
wGCj,j′
∑t s=1
exp (
−t−s τGC
)
zj′(s−1), (2.7) ここでzj(t)は時間 tでの顆粒細胞j の活動を示しており,qj(t)の正の値で与えられる.
qj(t)は時間tにおける内部状態,I(t)は苔状線維を介しての求心性入力である.wj,jGC′ は 細胞j′ から細胞j へのゴルジ細胞を介した抑制性のシナプス荷重,τGC は時定数をそれ ぞれ示す.顆粒細胞の活動を図2.13に示す. それぞれのニューロンが異なる時間パター ンで活動状態になるが,集団でみると,活動する顆粒細胞集団は一意である.言い換えれば 活動するニューロン集団によって時間経過を表現している. 一方,時刻 t でのプルキンエ
細胞の活動PC(t) は以下で表される.
PC(t) =∑
j
wjPFzj(t)−∑
j
zj(t) + PC, (2.8)
ここで右辺第1項は顆粒細胞から直接投射する興奮性入力,第2項は分子層介在ニューロ ンを介した抑制性入力を表す.分子層介在ニューロンは顆粒細胞から入力を受け取り,プ ルキンエ細胞を抑制する.第3項は定常状態の活動を表す.本論文では顆粒細胞からの入 力の総和に応じてプルキンエ細胞を抑制するものとした.wjPFはプルキンエ細胞と顆粒 細胞j とのシナプス荷重を表す.小脳核の活動CN(t)は式(2.9)で記述される.
CN(t) = {
1 PC(t)< θPC かつ PC˙ <0,
0 それ以外, (2.9)
小脳核はプルキンエ細胞の下流に存在し,プルキンエ細胞の活動が低下し抑制が減弱する とバースト的に強く活動する.その過程を再現するために,通常は一定の強さにあるプル キンエ細胞の活動PC(t)が低下し,閾値θPC を横切った瞬間に活動するものとした.式
(2.9)において,CN(t) = 1となる最初の条件はプルキンエ細胞の活動が閾値を下回って
いること,二番目の条件は閾値を上から下へ横切ることをそれぞれ表している.また,こ のモデルは小脳プルキンエ細胞のLTDを模擬しており,タイミング学習を可能にしてい る.wPFj を以下の式で更新する.
wPFj = max(1, wPFj + PFi(t)wjPFIO(t)), (2.10) PFj(t)は顆粒細胞の活動を表し,zj(t)> 0ならば1,それ以外は0である.IO(t)は登 上線維刺激であり,学習における教師信号を表す.0または1の値を取る.wjPF の学習 は抑制方向に進み,値はゼロへ推移していく.よって,負の方向への変位量に着目するた め,wPFj の上限を1とした.
50
0 10 20 30 40
0 0.2 0.4 0.6 0.8 1.0
ニューロン番号
時間 (秒)
図2.13 顆粒細胞の活動.活動した(zj(t) >0)時刻を線分で表した. 各細胞はそれぞ れ違った形で,活動状態と非活動状態を繰り返す. 1000個の顆粒細胞のうち50個をプ ロットしている.
2.3.5 遺伝的アルゴリズム
遺伝的アルゴリズムの概要
前述したCPGモデルなどでは,実験的に得られないパラメータが多数存在する. これら のパラメータを決定するために4章では遺伝的アルゴリズム (GA: Genetic Algorithms) を使用した. GAとは生物学の遺伝的メカニズム,いわゆる生物進化(自然淘汰・交叉・突 然変異)の現象を,計算機シミュレーションによって模倣した確率的探索・最適化・学習 の一手法である[61].いわゆる, メタヒューリスティクス(meta heuristics)の1つであり, 個体と呼ばれる染色体の集合が外部環境に適応するように,次に示す規則に基づく集団の 構成を世代ごとに生成させるものである.
1. 適合性の高い個体ほど生存率が高い(自然淘汰)
2. 古い個体をもとに新しい個体を遺伝的操作により生成させる(遺伝現象)
GAを組合わせ最適化問題の一解法とした場合,規則1.を確率的探索法,また規則2.を経 験的探索法と見なすことができ, GAは両者の側面を持つメタヒューリスティクス法の1
つである. 大まかな処理の流れを以下に述べていく(図2.14 [61]).
図2.14 基本的なGA処理の流れ.初期解集団を設定し,交叉・突然変異といった遺伝 的操作を経て次世代に残すべき染色体を決定する. [61]の図を参考に作成.
遺伝子表現と染色体集団
実際の生物において,遺伝情報を伝える役目をしているのは染色体 (chromosome),また は個体(individual) である. GAでは,対象問題の解候補を染色体として扱い, 1次元ある いは2次元のビットまたは文字/数値データ列として表現する.これが対象問題の遺伝子 表現(representation),あるいはコード化である. また,このようにコード化された解を遺 伝子型(genotype),実際の解を表現型(phenotype)と呼ぶ. 初期解として,決められた数
(集団サイズ)だけの染色体を問題の制約条件のもとでランダムに生成し, 初期の染色体集 団を作成する.この処理手順をエンコーディングルーチンと呼ぶ.また,染色体から対象問 題の解候補を生成する手順をデコーディングルーチンと呼ぶ.
![図 2.2 重複歩とステップ . 左下肢の接地から次の接地までが重複歩であり , 右のステッ プは左接地から右接地の距離である . [16] の図を参考に作成。 2.1.2 歩行の相 相による歩行分析は , 各関節で起こる種々の動作の機能的な意義を評価する](https://thumb-ap.123doks.com/thumbv2/123deta/7729322.1711518/14.892.117.788.177.386/重複ステップ左下肢接地から接地までステッによる各関節起こる.webp)


![図 4.9 2 足歩行における代償手段の仮説 . 左側は反射フィードバックループである . 患 者が反射ループの調整で 2 足歩行を獲得したとき , 亜急性期脳梗塞患者のように motor module の合併は確認されない [42]](https://thumb-ap.123doks.com/thumbv2/123deta/7729322.1711518/74.892.192.703.174.581/足歩行おける代償手段仮説左側フィードバックループループ足歩行.webp)
