細胞周期
cell cycle と細胞分裂
●テーマ 1 セルサイクル制御概観
●テーマ 2 セルサイクル制御発見小史
●テーマ 3 細胞骨格と細胞分裂
● ●参考書
–☞細胞周期の分子生物学
対立的な実験事実
生物群に応じて制御メカニズムは異なるのか
●ドミノモデルの主潮流 出芽酵母の
cdc変異遺伝学:
DNA複製が停止すれば分裂しない
–末端事象の順序の厳密な依存性➜依存経路
●時計モデルの主潮流 カエル等初期胚の卵割:
DNA複製を阻害しても分裂する
–セントラルタイマーによる制御➜独立経路
– ●この対立は
80年代半ばまで続く
モデル対立の統合的解決
制御メカニズムは真核生物に普遍的なものであった
●ドミノモデルからの研究➜
p34
cdc2キナーゼの発見
●時計モデルからの研究➜
MPFとサイクリンcyclinの発見
●MPFの精製➜MPF = p34
cdc2キナーゼ
+ cyclin Bの発見
両モデルの分子的共通性➜両モデルで何が異なるのか?➜
●チェックポイント概念の導入
時計モデルの対象(胞胚期中期まで)はチェックポイント欠落
–➜統合(80年代末)
–➜2
.セルサイクル制御の基本的枠組みの統一見解
2.セルサイクル制御の基本的枠組み
❶セルサイクルエンジン: 自律的に振動し、その位相に応じて末端事象を誘起(ON信号)
❷末端事象:成長、複製、分裂など細胞周期の巨視的事象(最下流の事象)
テーマ2 セルサイクル制御発見小史
●1.
MPF機能とサイクリンB機能・実体の発見
–(い)
MPF機能の発見
–(ろ) サイクリンの発見
–(は) サイクリンB機能の解明
●2.
p34
cdc2キナーゼの機能と普遍性
–(に) 出芽酵母
CDC28がStart、分裂酵母cdc2がStartとM突入に必要
–(ほ)
cdc2遺伝子はp34
cdc2キナーゼをコード
–(へ)
p34
cdc2キナーゼ活性はセルサイクルとともに周期変動
–(と) 出芽酵母cdc2遺伝子は分裂酵母、ヒトにも
。
●3. MPFの精製
–(ち)
MPF=サイクリンB + p34
cdc2キナーゼ
●4. チェックポイント概念の提唱
– (り) ドミノモデルと時計モデルの統合テーマ2 セルサイクル制御発見小史
●1.
MPF機能とサイクリンB機能・実体の発見
–(い)
MPF機能の発見
–(ろ) サイクリンの発見
–(は) サイクリンB機能の解明
☞ECB p615-616 解明への手がかり
● 2.p34cdc2キナーゼの機能と普遍性 – (に) 出芽酵母CDC28がStart、分裂酵母cdc2がStartとM突入に必要 – (ほ) cdc2遺伝子はp34cdc2キナーゼをコード – (へ) p34cdc2キナーゼ活性はセルサイクルとともに周期変動 – (と) 出芽酵母cdc2遺伝子は分裂酵母、ヒトにも。 ● 3. MPFの精製 – (ち) MPF=サイクリンB + p34cdc2キナーゼ ● 4. チェックポイント概念の提唱 – (り) ドミノモデルと時計モデルの統合Maturation Promoting Factor
Mitosis Promoting Factor
成熟分裂促進因子 有糸分裂促進因子(い)MPF機能の発見:顕微注射
GREAT EXPERIMENTS
The discovery of MPF
Yoshio Masui
In: CELLS (B. Lewin ed.)
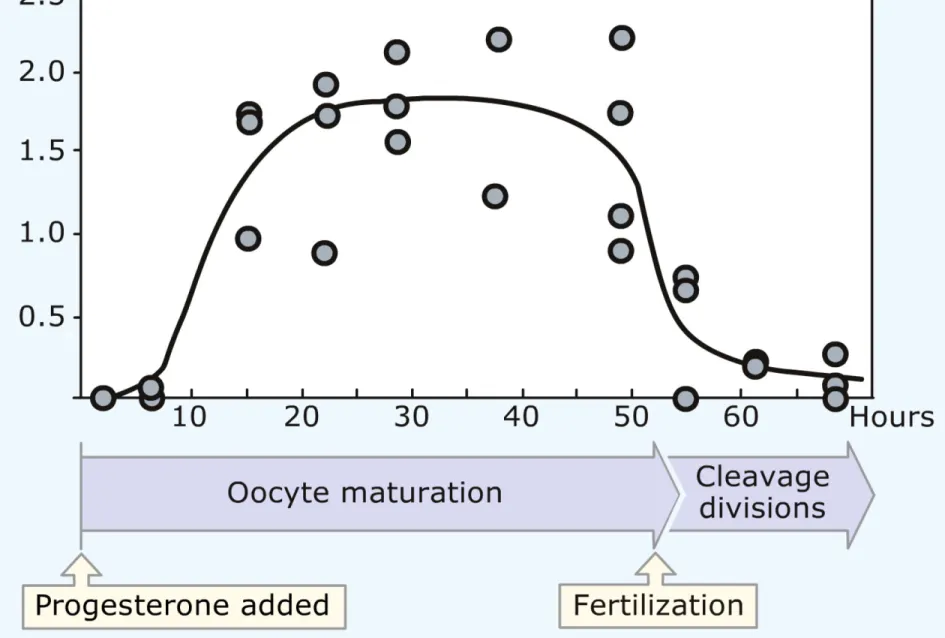
Figure 3 Maturation-promoting factor activity in the oocyte cytoplasm during maturation and early development. The horizontal axis indicates the age of the oocyte (hours since treatment with
progesterone) and the vertical axis the concentration of MPF activity in its cytoplasm. (Fig. 10 of 1582: Masui, Y. and Markert, C. L. (1971). Cytoplasmic control of nuclear behavior during
MPF振動は核・中心小体・ 微小管とは独立に生ずる (1960-1984)。 MPF活性の上昇にも、M 期への突入にもタンパク質 合成は必須である (~1983)
Adapted from Murray, A. W. & Kirschner, M. W. (1989) Dominoes and Clocks: The Union of the Two View of the Cell Cycle. Science 246, 614-621
Prophase arrest 出生時 70万~200万 思春期 4万 G2 arrest 排卵 ~500/生涯 –
細胞周期の分子生物学より
MPFは機能的タンパク質として発見されたが、
その分子的実体は不明のまま残された
●
MPFの機能:
maturation (mitosis) promoting
●
MPFの挙動:M期に最高となる形で周期的に変動
●
●
増井禎夫『細胞と人間のサイエンス』 ☞
http://bit.ly/1doUsD7
● ●「細胞周期の主要な制御因子の発見」
2001年
ノーベル生理学・医学賞
–何故、ノーベル賞受賞とならなかったのか?
–山中博士の
iPS細胞作成の基礎的知見を発見したガー
ドン博士は同時受賞した。
Expt by Gurdon
(1962)
Procedure used to obtain mature frogs from the intestinal nuclei of Xenopus tadpoles. The wild-type egg (with two nucleoli per
nucleus; 2-nu) is irradiated to destroy the maternal
chromosomes, and an intestinal nucleus from a marked (1-nu) tadpole is inserted. In some
cases, there is no cell division; in some cases, the embryo is
arrested in development; but in other cases, an entire new frog, with a 1-nu genotype, is formed. (After Gurdon 1968,Gurdon
1977.)
From: Evidence for Genomic Equivalence
テーマ2 セルサイクル制御発見小史
●1.
MPF機能とサイクリンB機能・実体の発見
–(い)
MPF機能の発見
–(ろ) サイクリンの発見
–(は) サイクリンB機能の解明
● 2.p34cdc2キナーゼの機能と普遍性 – (に) 出芽酵母CDC28がStart、分裂酵母cdc2がStartとM突入に必要 – (ほ) cdc2遺伝子はp34cdc2キナーゼをコード – (へ) p34cdc2キナーゼ活性はセルサイクルとともに周期変動 – (と) 出芽酵母cdc2遺伝子は分裂酵母、ヒトにも。 ● 3. MPFの精製 – (ち) MPF=サイクリンB + p34cdc2キナーゼ ● 4. チェックポイント概念の提唱 – (り) ドミノモデルと時計モデルの統合(ろ)サイクリンの発見:
RIラベル実験
1983 by T. Hunt
機能未知タンパク質の、
セルサイクルに応じたリズミカルな変動
GREAT EXPERIMENTS
The discovery of cyclins
Tim Hunt
In: CELLS (B. Lewin ed.)
ウニ受精卵:連続ラベル実験 受精直後に35Sメチオニンを投与 し、時間を追って反応を止め、SD Sゲル電気泳動にかけてタンパク 質を分離した。 “cyclin”バンドのみ周期的に変 動した。 Evans, T., Rosenthal, E. T., Youngblom, J., Distel, D., and
Hunt, T. (1983). Cyclin: a
protein specified by maternal mRNA in sea urchin eggs
that is destroyed at each cleavage division. Cell 33, 389-396. Google Scholar
●
連続ラベル実験で分かること
–新規合成(
de novo合成)の有無
●合成は定常的だが、分解は周期的
●合成は周期的だが、分解は定常的
合成量
分解量最小
正味合成量の最大
分解量
新規合成量
及び
正味合成量の最大
ウニ受精卵:パルスラベル実験 受精直後から様々な時間に10分だ け35Sメチオニンをパルス投与した。 どの時間にパルス投与しても、 “cyclin”合成量はほとんど同じ ➜cyclin量の周期変動が周期的な cyclin分解に起因することが判明 (もし、特定の時期にだけ合成される なら、その時期だけにラベルcylinが 検出されるはず) Evans, T., Rosenthal, E. T., Youngblom, J., Distel, D., and
Hunt, T. (1983). Cyclin: a
protein specified by maternal mRNA in sea urchin eggs that is destroyed at each cleavage
division. Cell 33, 389-396.
テーマ2 セルサイクル制御発見小史
●1.
MPF機能とサイクリンB機能・実体の発見
–(い)
MPF機能の発見
–(ろ) サイクリンの発見
–(は) サイクリンB機能の解明
サイクリンの機能は(ろ)の段階では不明
①卵細胞抽出液を用いた
in vitro実験系の開発(1983)
②サイクリンB添加によるセルサイクル再開(
1989)
③サイクリンB破壊による間期停止(
1989)
④サイクリンB破壊抑制によるM期停止(M期脱出の抑制)(
1989)
●(は)サイクリンB機能の発見
①卵細胞抽出液を用いた
in vitro実験系の開発
Masui & Lohka (1983)
•On addition, sperm nuclei decondense
then their DNA undergoes an essentially normal round of DNA replication
•Extract can complete 3-5 cycles •Cycle time ~2 x that of intact eggs
(は)サイクリンB機能の発見
②サイクリンB添加によるセルサイクル再開
Murray
& Kirshner (1989)
●
Cyclin synthesis drives the early embryonic cell
cyle. Nature 339: 275-280
Google Scholar
●
サイクリンBだけを合成する条件で、M期突入ができ
る
–
➜
サイクリンB合成はM期突入の十分条件
–
次の図
–
RNaseで、すべてのmRNAを破壊した後
–
➜RNase阻害剤を添加してサイクリンmRNA
を添加
●(は)サイクリンB機能の発見
③
サイクリンB破壊による間期停止
Minshull, Blow & Hunt (1989)
●
Translation of cyclin mRNA is necessary for extract
of Xenopus eggs to enter mitosis. Cell 56: 947-956
Google Scholar
●サイクリンB合成だけを抑制する条件で、
in vitro セル
サイクルは間期に停止する(セルサイクルが止まる)
–
➜
サイクリンB合成はM期突入の必要条件
–
次の図
–
アンチセンスオリゴヌクレオチドでサイクリンB
mRNAを処理し、RNaseHにて、これを特異的
に破壊➜セルサイクルは停止
●(は)サイクリンB機能の発見
④
サイクリンB分解の抑制➜M期脱出抑制
Murray, Solomon & Kirshner (1989)
●
The role of cyclin synthesis and degradation in
the control of maturation promoting factor activity
Nature 339: 280-286
Google Scholar
●
サイクリンB分解だけを抑制する条件で、M期脱出を
抑制できる
–
➜
サイクリンB分解はM期脱出の必要条件
–
次の図
–
サイクリンB
N末端の90個のアミノ酸残基を
欠失させる
mRNAを作成し、in vitroセルサイ
クル系に添加する➜サイクリンは分解されず
に増え続け、M期に突入できるが脱出できな
い
テーマ2 セルサイクル制御発見小史
●1.
MPF機能とサイクリンB機能・実体の発見
–(い)
MPF機能の発見
–(ろ) サイクリンの発見
–(は) サイクリンB機能の解明
●2.
p34
cdc2キナーゼの機能と普遍性
–(に) 出芽酵母
CDC28がStart、分裂酵母cdc2がStartとM突入に必要
–(ほ)
cdc2遺伝子はp34
cdc2キナーゼをコード
–(へ)
p34
cdc2キナーゼ活性はセルサイクルとともに周期変動
–(と) 出芽酵母cdc2遺伝子は分裂酵母、ヒトにも
。
●3. MPFの精製
–(ち)
MPF=サイクリンB + p34
cdc2キナーゼ
●4. チェックポイント概念の提唱
– (り) ドミノモデルと時計モデルの統合2.
p34
cdc2キナーゼの機能と普遍性
酵母の
cdc遺伝学の系譜から
–(に) 出芽酵母CDC28がStart、分裂酵母cdc2がStartとM突入に必要
–(ほ) cdc2遺伝子はp34
cdc2キナーゼをコード
–(へ) p34
cdc2キナーゼ活性はセルサイクルとともに周期変動
–(と) 出芽酵母cdc2遺伝子は分裂酵母、ヒトにも
。
(に) 出芽酵母CDC28がStart、
分裂酵母cdc2がStartとM突入に必要
二種の酵母を用いた遺伝学➜将棋倒しモデル
GREAT EXPERIMENTS
Cell cycle genes
Lee Hartwell
In: CELLS (B. Lewin ed.)
出芽酵母
Saccharomyces cervisiae
温度感受性変異株
temperature-sensitive mutant
(
ts-mutant) の分離
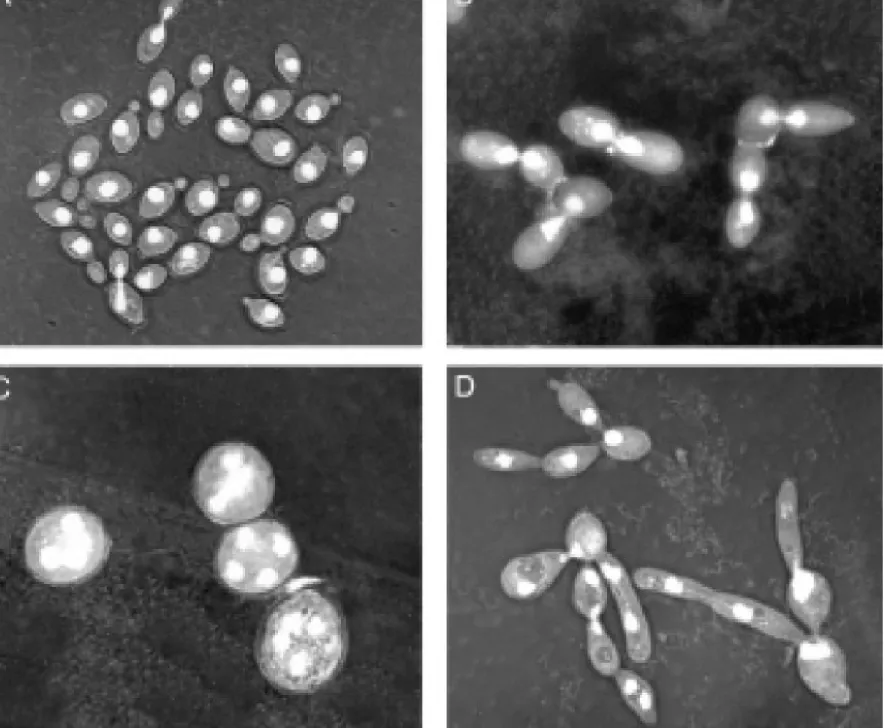
ECB1-35Figure 5. Normal cells and cdc mutant cells several hours after
incubation at the restrictive temperature. (A) wild type, (B) cdc8 (C)
Synopsis of Hartwell’s cell cycle pathway diagram
(~1975)
ファージ形態形成に模した順序的な依存経路(将棋倒しモデル) ● 最重要な遺伝子はcdc28 (Start) ● 出芽、DNA複製、紡錘体極(中心体centrosomeに相当)は互いに独立 ● 有糸分裂サイクルは細胞質分裂や出芽がなくても回転(に) 出芽酵母CDC28がStart、
分裂酵母cdc2がStartとM突入に必要
二種の酵母を用いた遺伝学➜将棋倒しモデル
GREAT EXPERIMENTS
The discovery of cdc2 as the key regulator of the cell cycle
Paul Nurse
In: CELLS (B. Lewin ed.)
WT
分裂酵母
Schizosaccharomyces pombe のセルサイクル
二重変異で正常に戻っている
Nurseらは、cdc2(とcdc13)がM期突入の律速段階であること
(M期突入に必須であること)を解明
Genetic control of cell size at cell division in yeast.
Nature 256, 547-551 (1975)
Google Scholar
まとめ
出芽酵母 CDC28がStartに必要
Hartwell group (1977)
分裂酵母 cdc2 がStartとG2/M転換に必要
Nurse group (1981)
Nurse groupは次に、cdc2遺伝子を単離し、その実体を解明
2.
p34
cdc2キナーゼの機能と普遍性
酵母の
cdc遺伝学の系譜から
–(に) 出芽酵母CDC28がStart、分裂酵母cdc2がStartとM突入に必要
–(ほ) cdc2遺伝子はp34
cdc2キナーゼをコード
–(へ) p34
cdc2キナーゼ活性はセルサイクルとともに周期変動
–(と) 出芽酵母cdc2遺伝子は分裂酵母、ヒトにも
。
cdc2ts
cdc2遺伝子がcdc2ts変異株を相補する遺伝子として単離された。
配列が決められ、タンパク質キナーゼ活性をもつことも判明した。
2.
p34
cdc2キナーゼの機能と普遍性
酵母の
cdc遺伝学の系譜から
–(に) 出芽酵母CDC28がStart、分裂酵母cdc2がStartとM突入に必要
–(ほ) cdc2遺伝子はp34
cdc2キナーゼをコード
–(へ) p34
cdc2キナーゼ活性はセルサイクルとともに周期変動
–(と) 出芽酵母cdc2遺伝子は分裂酵母、ヒトにも
。
出芽酵母 CDC28がStartに必要
Hartwell group (1977)
分裂酵母 cdc2 がStartとG2/M転換に必要
Nurse group (1981)
相補性テストにより
分裂酵母 cdc2 ≒ 出芽酵母 CDC28 (82)
≒ ヒトcdc2 (87)
by Nurse group
Cdc2Pはp34
cdc2キナーゼ。
その活性はセルサイクルに応じて周期的に変動
(by Nurse group, 84-87)
テーマ2 セルサイクル制御発見小史
●1.
MPF機能とサイクリンB機能・実体の発見
–(い)
MPF機能の発見
–(ろ) サイクリンの発見
–(は) サイクリンB機能の解明
●2.
p34
cdc2キナーゼの機能と普遍性
–(に) 出芽酵母CDC28が
Start、分裂酵母cdc2がStartとM突入に必要
–(ほ)
cdc2遺伝子はp34
cdc2キナーゼをコード
–(へ)
p34
cdc2キナーゼ活性はセルサイクルとともに周期変動
–(と) 出芽酵母cdc2遺伝子は分裂酵母、ヒトにも
。
●3. MPFの精製
–(ち)
MPF=サイクリンB + p34
cdc2キナーゼ
4. チェックポイント概念の提唱
–(り) ドミノモデルと時計モデルの統合
Lohka MJ, Hayes MK, Maller JL. (1988) Purification of maturation- promoting factor,
an intracellular regulator of early mitotic events.
Proc Natl Acad Sci U S A . 85 , 3009-13
Google Scholar
Gautier J, Norbury C, Lohka M, Nurse P, Maller J. (1988) Purified
maturation-promoting factor contains the product of a Xenopus homolog of the fission yeast cell cycle control gene cdc2+. Cell 54 , 433-439
●
MPFの実体解明➜二つの潮流が、共通のタンパク質
(セルサイクルエンジン)によって合流した。
●
チェックポイント概念の導入➜時計モデルとドミノモデ
● Masui, Y. (2001) From oocyte maturation to... Differentiation 69: 1-17
● Coudreuse, D. & Nurse, P. (2010) Driving the cell cycle... Nature 468:
1074-9
● Genetics of cell division cycle (ePlantScience.com)
● The control of septum formation in fission yeast (Genes & Dev. 1997. 11:
2939-2951)
● Polo-like kinases: a team that plays throughout mitosis (Genes & Dev. 1998.
12: 3777-3787 )
● Cell Cycle Molecules and Mechanisms of the Buddingand Fission Yeasts
(IN: Meth Mol Biol vol. 296 by Human Press a huge!!! pdf file)