沖縄産植物からの抗炎症作用および抗骨粗鬆症作用を有する
天然物の探索と作用機序解明に関する研究
(Studies on Search for Natural Products with Anti-inflammatory and
Anti-osteoporotic Effects from Okinawan Plants
and Their Action Mechanisms)
理学研究科
物質分子系専攻
平成
29 年度
熊谷百慶
Studies on Search for Natural Products with Anti-inflammatory and Anti-osteoporotic Effects from Okinawan Plants and Their Action Mechanisms
Momochika Kumagai
Abstract
In the aging society, utiliging food ingredients that can be ingested on a daily basis have attracted attention for preventing various diseases. The author focused on inflammation, which is known to be deeply involved in a wide range of disease states, and osteoporosis, where the number of patients has significantly increased in recent years in Japan. This thesis describes isolation of biologically active compounds from the natural products in Okinawa Prefecture, focusing on anti-inflammatory effects and anti-osteioporosis effects. Furthermore, to elucidate the molecular mechanism of their pharmacological actions in detail, structure activity relationship studies are also discussed.
This thesis consists of five chapters. The first chapter outlines inflammation and osteoporosis and reports on natural products effective for their improvement. In the second chapter, the author searched for natural products with anti-inflammatory activities from foods or herbal medicines collected in Okinawa. Bioassay-guided fractionation was caried out, and 5,6-Dehydrokawain (DK) and dihydro-5,6-dehydrokawain (DDK) were isolated from Alpinia zerumbet. Furthermore, aculeatin (ACU) and toddaculin (TOD) were obtained from Toddalia asiatica. Among them, detailed action mechanisms of ACU and TOD were examined using LPS-stimulated macrophage-like cell line RAW264 cells. As a result, although ACU and TOD had a similar structure, it was suggested that their mode of action could be different. In the third chapter, anti-osteoporotic effect was examined as a new function of DK and DDK isolated from A. zerumbet. As a result, DK and DDK significantly promoted the differentiation of mouse-derived osteoblastic MC3T3-E1 cells, and for the first time the author clarified the possibility that these compounds are effective for prevention and treatment of osteoporosis. In Chapter 4, as a part of efforts to elucidate the mechanism of action of DK, the author synthesized several partially modifyied DK analogs and clarified structural factors responsible for expression of the DK activity. The author elucidated importance of the A ring of DK, and created new compounds exceeding the activity of DK. Finally, in Chapter 5, the author evaluated the activity of DK molecular probe, and the auther visualized it in cells. DK has spread throughout the living cell including the nucleus and showed the possibility of interacting with a wide range of proteins. The results of these studies are expected to contribute to development of new functional foods and drugs having anti-inflammatory action and anti-osteoporotic action.
略語表
本論文では以下の略語を使用する。 ACU aculeatin
αMEM minimum essential medium eagle, alpha modification BMP-2 bone morphogenetic protein 2
BuOH 1-butanol
cDNA complementary deoxyribonucleic acid COSY correlation spectroscopy
DDK dihydro-5,6-dehydrokawain DK 5,6-dehydrokawain
DMEM Dulbecco's modified Eagle's medium DMSO dimethyl sulfoxide
DNA deoxyribonucleic acid EC50 effective concentration 50
ELISA enzyme linked immuno sorbent assay ERK extracellular signal-regulated kinase ESI electrospray ionization
EtOH ethanol EtOAc ethyl acetate FBS fetal bovine serum
HMBC heteronuclear multiple bond correlation HPLC high performance liquid chromatography HRMS high resolution mass spectrometry HSQC heteronuclear single quantum correlation IL interleukin
IR infrared spectroscopy
J coupling constant (in NMR) JNK c-jun N-terminal kinase LPS lipopolysaccharide
MAPK mitogen-activated protein kinase MeOH methanol
MeCN acetonitrile
mRNA messenger ribonucleic acid
NADH nicotinamide adenine dinucleotide
NMR nuclear magnetic resonance NFκB nuclear factor-kappa B
NOESY nuclear Overhauser effect spectroscopy PBS phosphate buffered saline
RANKL receptor activator of nuclear factor kappa-B ligand RNA ribonucleic acid
RT-PCR reverse transcription polymerase chain reaction rt room temperature
Rt retention time SD standard deviation
SDS-PAGE sodium dodecylsulfate polyacrylamide gel electrophoresis TNF-α tumor necrosis factor alpha
TOD toddaculin UV ultraviolet
目次
第 1 章 序論
1第 2 章 沖縄産植物の抗炎症作用スクリーニングと
活性化合物の単離・同定および作用機序の解明
7 第 1 節 沖縄産植物抽出エキスの NO 産生抑制作用スクリーニング 8 第 2 節 月桃 (Alpinia zerumbet) からの抗炎症活性物質の 単離・構造決定 11第 3 節 サルカケミカン(Toddalia asiatica (L.) Lam.)からの
抗炎症化合物の単離・構造決定 23 第 4 節 Aculeatin と toddaculin の抗炎症作用機序 32 第 5 節 実験項 41
第 3 章 月桃根茎由来 5,6-dehydrokawain と
dihydro-5,6-dehydrokawain の骨代謝改善作用
49 第 1 節 5,6-Dehydrokawain および dihydro-5,6-dehydrokawain の 骨芽細胞分化促進効果 51 第 2 節 5,6-Dehydrokawain の骨芽細胞分化促進作用における 作用機序の検討 55 第 3 節 5,6-Dehydrokawain と dihydro-5,6-dehydrokawain の破骨細胞分化に対する影響 59 第 4 節 5,6-Dehydrokawain 類縁体の合成 61 第 5 節 実験項 63第 4 章 5,6-Dehydrokawain の骨芽細胞分化促進作用および
破骨細胞分化抑制作用に対する構造活性相関
69 第 1 節 5,6-Dehydrokawain 類縁体の合成 71 第 2 節 5,6-Dehydrokawain の骨芽細胞分化促進作用に対する 構造活性相関 74 第 3 節 高活性 5,6-dehydrokawain 誘導体の 骨芽細胞分化促進作用における作用機序 83 第 4 節 5,6-Dehydrokawain 誘導体の RAW264 細胞における 破骨細胞分化抑制作用とその作用機序 87 第 5 節 実験項 97第 5 章 MC3T3-E1 細胞における
5,6-dehydrokawain 標的分子の探索
141 第 1 節 5,6-Dehydrokawain の標的分子同定に使用する 分子プローブの機能評価 142 第 2 節 5,6-Dehydrokawain の細胞内局在 145 第 3 節 実験項 147第 6 章 総括
149参考文献
152本論文の内容に関連する公表論文
157謝辞
1581
第
1 章 序論
高齢化社会の到来により,国民医療費および対国内総生産比, 対国民所得比は 増加の一途をたどっている1。また, 国民医療・介護分野における高齢者や患者 の生活の質の向上にも関心が集まっており,罹患後の適切な治療が重要である ものの, "病気にならないこと" が国民にとって医療費削減のみならず最も根本 的な生活の質改善につながると考えられる。 その解決策の一つとして日常的に摂取できる食品成分を様々な疾病の予防に 役 立 て る た め の 研 究 が お こ な わ れ て い る 2。 温 州 ミ カ ン な ど に 含 ま れ るβ-cryptoxanthin (Fig. 1-1, 1-1) や大豆もやしに含まれる daidzein (Fig. 1-1, 1-2) は,骨代謝改善効果に関する活性 3が着目されている。β-Cryptoxanthin (1-1)を 含有する三ヶ日ミカンは, 2015 年度より施行された消費者庁による食品の機能 性表示制度に初の生鮮食品として登録された。また,大豆もやしについても同 様に骨の健康を維持する働きが認められ,温州ミカンと同時に機能性表示食品 として登録がなされている。このような背景のなか,機能性に関与する成分や その作用機構を明確にすること,さらに安全性を担保することが研究面で強く 求められている2。
2 悪性新生物,心臓病や脳血管疾患など,日本人の主要な死因の下地になって いる病気にメタボリックシンドロームがある。糖尿病や脂質異常症,高血圧な どの生活習慣病と肥満が複合した病態であるメタボリックシンドロームの病態 には,マクロファージが恒常的に過剰に活性化した状態である慢性炎症が深く 関与するといわれている4。マクロファージは体内のあらゆる臓器,組織に局在 し,病原体や異物などを貪食,処理するとともに,病原体構成成分を認識して 炎症応答を惹起する細胞である5。マクロファージは,病原体やウイルス等から 生体を防御するために無くてはならない存在であるが,脂肪組織の分泌する遊 離脂肪酸やアディポサイトカインなどをメディエータ―として異常に活性化す る事で慢性炎症の状態が全身に拡大・波及し,前述の様々な疾病を引き起こす ことが明らかとなっている。マクロファージ自身も,免疫応答の過程において NO をはじめとした多様な炎症性メディエータを産生することが知られている 6。 NO は,内皮細胞由来平滑筋弛緩因子であることが発見されて以来,神経伝達物 質など多くの生理活性を有することが明らかとなっている。一方で,過剰に産 生された NO は,活性酸素である O2-と反応してより酸化力の強い ONOO-となり, DNA の損傷やタンパク質の変性などを引き起こし,炎症性疾患や腫瘍などの危 険性を高めると考えられている(Fig. 1-2)。よって,過剰な NO 産生を抑制する ことは,慢性炎症の改善のみならず,酸化ストレスが要因で引き起こされる様々 な疾病の予防にも繋がると考えられる。
3
NO は細菌の細胞壁成分であるリポポリサッカライド (LPS) などで炎症惹起 されたマクロファージにおいて発現誘導される誘導型 NO 合成酵素 (iNOS)に より,アルギニンから合成されることが知られている(Fig. 1-3)6。
Fig. 1-3. Activation of macrophages by LPS and NO production mechanism.
このため,LPS 刺激をしたマクロファージにおける NO 産生量を測定すること で, 種々のサンプルの抗炎症活性を簡易的に評価する手法が用いられている。 Fig. 1-4 に Griess 法の測定原理を示した。Griess 法では,LPS 刺激によりマクロ ファージが産生した NO が酸化されて生じる NO2-と sulfanilamide (1-3) によっ てジアゾニウム塩化合物 1-4 が生成し,さらに N-(1-Naphthyl)ethylenediamine (1-5)とのジアゾカップリングによって生成したアゾ色素 1-6 の吸光度を測定 することで,NO 産生量を間接的に測定することができる。このような測定系を 用いて,これまでに食品素材や天然物から様々な抗炎症活性物質が報告されて いる (Fig. 1-5) 7-9。
4 NO2- H2N SO2NH2 N N SO2NH2 H H N H2N H N H2N N N SO2NH2 + N-(1-Naphthyl)ethylenediamine (1-5) Sulfanilamide (1-3) 1-4 1-6
Fig. 1-4. Measurement principle of Griess method.
Fig. 1-5. Anti-inflammatory compounds from edible materials.
また,炎症が関与する疾患の一つでもあり,超高齢化社会の到来により患者 数の増加が著しい骨粗鬆症も生活の質を低下させる要因として近年問題となっ ている。骨粗鬆症患者は日本国内で推定 1200 万人を超えており10,今後も増加 の一途をたどると推察されている。健常な骨代謝は,生体において常に一部が 吸収され,また同時にほぼ同等の量の骨が形成されている。この形成と吸収の 持続的な進行により骨は作り変えられるが,この過程をリモデリングと呼ぶ。
5 骨の形成と吸収の量は大人で概ね1 日 500 mg のカルシウムが骨から血中へと骨 の吸収活性により移行し,一方骨の形成に伴いほぼ同量のカルシウムが血中か ら骨へと移行するとされている 11。骨の形成と吸収のバランスの崩れが骨の吸 収側に傾いた状態が長期にわたって持続することで骨量が減少し,骨がその強 度の低下により骨折しやすくなる状態が骨粗鬆症である 10。骨粗鬆症は,骨形 成と骨吸収のバランスがくずれた状態であるが,骨形成と骨吸収は,それぞれ 骨芽細胞と破骨細胞がその役割を担う。よって,骨形成を促進させる化合物 (Fig. 1-6) 12-15や,骨吸収を抑制する化合物 (Fig. 1-7)16-18は骨粗鬆症の予防や治療に 有効な可能性が考えられ,これまでに様々な天然素材から活性物質の探索が行 われている。
Fig.1-6. Osteogenic compounds obtained from natural materials.
Fig.1-7. Osteoclastogenesis inhibitors obtained from natural materials.
本研究では,日常的に摂取できる植物成分から,抗炎症活性および骨代謝改 善作用を持つ化合物を探索し,その作用機序を詳細に調べることでこれらの有 効利用に資する知見を得ることを目的とした。特に,本研究では沖縄に生息す
6 る植物に着目した。沖縄に生息する植物は常に高温多湿や紫外線の影響を受け ており,そのようなストレス環境に適応するために様々な二次代謝産物を産生 すると考えられている。また,沖縄産植物の新たな機能性を発掘することで, 地域資源の六次産業化19につながる可能性があると考えられる。これまでに, 沖縄産植物から調製したエキスに関して各種ガン細胞に対する増殖抑制作用や, マクロファージに対する抗炎症活性が報告されており20,21本州などで通常食用 とされている素材に対して活性が高いことが報告されている21。そこで,本研究 ではこれまでに抗炎症作用について報告の少ない沖縄産植物を中心に収集・調 製した抽出エキスについて,培養細胞を用いて抗炎症活性を有する素材をスク リーニングした。さらに,見出した素材から活性物質を探索し詳細な作用機序 を検討した。見出した活性物質について,新たな機能性として骨代謝改善効果 を検討し,さらに詳細な作用機序解明に向けた検討を行った。
7
第 2 章 沖縄産植物の抗炎症作用スクリーニングと
活性化合物の単離・同定および作用機序の解明
本章では,沖縄県にて収集した機能性食品・生薬素材の中から,抗炎症作用 を有する素材をスクリーニングし,活性の高かった素材から抗炎症作用を有す る化合物の単離・構造決定を試みた。また,得られた化合物の抗炎症活性の作 用機構について詳細に解析した。8
第 1 節 沖縄産植物抽出エキスの NO 産生抑制作用
スクリーニング
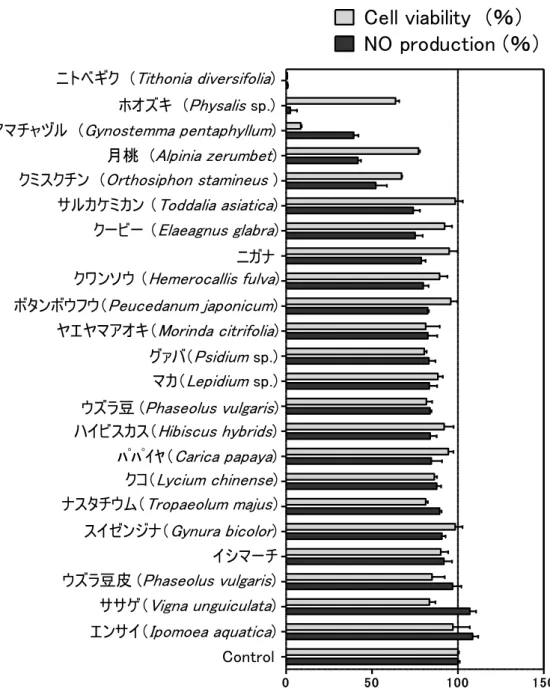
1. 目的 日常的に摂取できる食品や生薬素材から NO 産生抑制作用を有する素材を探 索することを目的とした。 2. 結果と考察 沖縄県にて収集した植物サンプル 23 種の MeOH 抽出物について,RAW264 細 胞を用いた NO 産生抑制効果のスクリーニング試験を行った。各サンプルは DMSO に溶解させ,DMSO の終濃度が 0.1% (v/v) 以下となるように培地に溶解 し,細胞に添加した。また,コントロールには,DMSO のみを 0.1%添加した培 地を細胞に添加した。LPS を添加したコントロールの NO 産生量を 100%とした ときの各サンプルにおける NO 産生量を Fig. 2-1 に示した。 また,同時に Cell counting kit-8 (同仁化学研究所)を使用して細胞毒性試験を行った(Fig. 2-2)。 本キット中には,テトラゾリウム塩(WST-8, 2-1)と電子伝達物質として1-methoxy-5-methylphenazinium methylsulfate (1-Methoxy PMS, 2-3)が含まれてお り,WST-8(2-1)は高い水溶性を有するため生細胞膜を透過せず細胞外に存在 し,乳酸脱水素酵素の補酵素である NADH から 1-Methoxy PMS を介して電子を 受け取ることで還元され, 水溶性の WST-8 ホルマザン(2-2)を生成する。このホ ルマザンの 450 nm の吸光度を直接測定することにより生細胞数を計測すること ができる。細胞毒性試験についてもコントロールにおける 450 nm の吸光度を 100%としたときの各サンプルにおける吸光度を相対値(%)として Fig. 2-1 に 示した。
9 0 50 100 150
NO production (%)
Cell viability (%)
ニトベギク (Tithonia diversifolia) ホオズキ (Physalis sp.) アマチャヅル (Gynostemma pentaphyllum) 月桃 (Alpinia zerumbet) クミスクチン (Orthosiphon stamineus) サルカケミカン (Toddalia asiatica) クービー (Elaeagnus glabra) ニガナ クワンソウ (Hemerocallis fulva) ボタンボウフウ(Peucedanum japonicum) ヤエヤマアオキ(Morinda citrifolia) グァバ(Psidium sp.) マカ(Lepidium sp.) ウズラ豆 (Phaseolus vulgaris) ハイビスカス(Hibiscus hybrids) パパイヤ(Carica papaya) クコ(Lycium chinense) ナスタチウム(Tropaeolum majus) Control エンサイ(Ipomoea aquatica) ササゲ(Vigna unguiculata) スイゼンジナ(Gynura bicolor) イシマーチ ウズラ豆皮 (Phaseolus vulgaris)Fig. 2-1. Screening of natural materials with anti-inflammatory activity. Cells were treated with extracts (50 µg/mL) and LPS (100 ng/mL) for 24 h and NO production was measured using the Griess reagent. Cell viability was measured by WST-8 assay. Data shown are mean ± SD. (n = 3).
10
Fig. 2-2. Measurment principle of cell counting kit-8.
NO 産生抑制試験の結果ニトベギク(Tithonia diversifolia),ホオズキ(Physalis sp.), アマチャヅル(Gynostemma pentaphyllum),月桃 (Alpinia zerumbet),クミス クチン(Orthosiphon stamineus), サルカケミカン(Toddalia asiatica),クービー (Elaeagnus glabra)などに NO 産生抑制効果が認められた(Fig. 2-1)。細胞毒性試 験の結果,最も NO 産生抑制効果の強かったニトベギク(Tithonia diversifolia)は, 50 µg/mL の濃度では顕著な細胞毒性が認められた。また,アマチャヅルは NO 産生抑制効果も強かったが(抑制率 61%), 細胞毒性も強かった。ホオズキ, 月桃,サルカケミカン,クービーは,NO 産生抑制率が細胞生存率よりも高い値 を示した。そこで,細胞毒性が低く NO 産生抑制効果が高いサンプルとして, 月桃, サルカケミカン,ホオズキについては抗炎症作用を有する機能性素材とし て今後研究を進める価値のあるものと判断した。また,文献調査の限り,これ らの素材について抗炎症作用を指標として活性化合物を単離・構造決定した報 告は見当らなかった。
11
第 2 節 月桃 (Alpinia zerumbet) からの抗炎症活性物質の
単離・構造決定
1. 目的 本節では,スクリーニングによって見出された月桃(Alpinia zerumbet)より,抗 炎症作用を有する化合物を単離・構造決定することを目的とした。特に,これ までに成分の分析が十分に行われていない月桃の根茎 (Fig. 2-3)について,琉球 大学名誉教授 多和田真吉博士より琉球大学キャンパス内にて採集されたもの を供与いただき,研究に用いることとした。12
2. 結果と考察
2-1. 抗炎症活性化合物の単離
月桃の根茎 (1.5 kg) を適度な大きさにカットした後に 1 日乾燥させた。3 L の MeOH で 3 回抽出(各 1 日)した後にろ紙でろ過し,40 ℃で減圧濃縮し,MeOH 抽出物を得た(MeOH ext., 21.3 g)。MeOH ext.を 500 mL の水と EtOAc で分配(×3) し,EtOAc 層を合わせて 40 ℃で減圧濃縮し, EtOAc 抽出物を得た (EtOAc ext., 4.7 g)。残りの水層を BuOH で分配 (300 mL×3) し,40 ℃で減圧濃縮して BuOH 抽 出物(BuOH ext., 1.64 g)を得た (Fig. 2-4)。残りの水層を 40 ℃で減圧濃縮して水 抽出物 (Water ext., 12.8 g)を得た。これらの分画物について 50 µg/mL の終濃度に おいて NO 産生抑制試験を行ったところ,EtOAc 抽出物に最も強い NO 産生抑制 効果が認められた(Fig. 2-5)。そこで EtOAc 抽出物を DIAION HP20 (三菱ケミ カ ル) を用いたオー プンカラムクロマト グラフィー (φ4.0×20 cm)に供し, MeCN/Water (2:8), MeCN/Water (4:6), MeCN/Water (6:4), MeCN/Water (8:2), MeCN で溶出した。溶出液を濃縮乾固し, E. Fr-1-5 を得た。得られた E. Fr-1- 5 について NO 産生抑制試験を行ったところ,MeCN 溶出画分 (E. Fr-5)の活性が 最も強かった(Fig. 2-6)。そこで E. Fr-5 の一部(0.93 g)をシリカゲルカラムクロ マ ト グ ラ フ ィ ー (Wakogel N60, φ15 mm×150 mm) に 供 し , Hexane (Hex), Hex/EtOAc (8:2), Hex/EtOAc (5:5)-1, Hex/EtOAc (5:5)-2, EtOAc 各 25 mL で溶出 させた(E. Fr-5-1~5)。活性が強く(Fig. 2-7),収量の多かった Hex/EtOAc (5:5)-2 の一部 (E. Fr-5-3, 59.2 mg)を逆相 HPLC (Detection: 210 nm, Solvent: MeCN/H2O
(6:4), Flow rate: 12 mL/min, Column: Wakosil-Ⅱ5C18RSPrep φ20×250 mm)で分取し (Fig. 2-8),化合物 2-5 (16.7 mg), 2-6 (11.6 mg)をそれぞれ白色固体として単離し た。
13
Fig. 2-4. Separation scheme of A. zerumbet.
Cont rol Me OH ex t. E tOAc ex t. BuOH ex t. Water ex t. 0 50 100 150 NO p ro du c ti o n ( %)
Fig. 2-5. NO inhibitory activities of each extract. Cells were treated with extracts (50 µg/mL) and LPS (100 ng/mL) for 24 h and NO production was measured using the Griess reagent. Data shown are mean ± SD (n = 3).
14 C o n tr o l 25 50 25 50 25 50 25 50 25 50 0 50 100 150 E. Fr-1 E. Fr-2 E. Fr-3 E. Fr-4 E. Fr-5 g/mL
NO
p
ro
du
c
ti
o
n
(
%)
Fig. 2-6. NO inhibitory activities of E. Fr-1~5. Cells were treated with extracts (25 and 50 µg/mL) and LPS (100 ng/mL) for 24 h and NO production was measured using the Griess reagent. Data shown are mean ± SD (n = 3).
C o n tr o l E . F r-5 -1 E . F r-5 -2 E . F r-5 -3 E . F r-5 -4 E . F r-5 -5 0 50 100 150
NO
p
ro
du
c
ti
o
n
(
%)
Fig. 2-7. NO inhibitory activities of E. Fr-5-1~5. Cells were treated with extracts (25 µg/mL) and LPS (100 ng/mL) for 24 h and NO production was measured using the Griess reagent. Data shown are mean ± SD (n = 3).
15
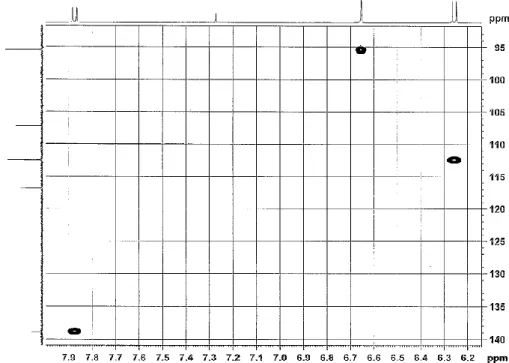
Fig. 2-8. HPLC chromatogram of E. Fr-5-3.
2-2. 化合物 2-5 の構造決定
化合物 2-5 の分子式は,HRMS (ESI): [M+H]+
calcd for C14H13O3, 229.0865;
found, 229.0866 より,C14H12O3であった。IR スペクトル (Fig. 2-9) より,カル
ボニル (1723 cm-1) および芳香環 (3080, 1608 cm-1) の存在が示唆された。 1 H-NMR スペクトルにおいて,δ 7.52-7.35 (5H, m) より 1 置換ベンゼンの存 在が示唆された(Fig. 2-10)。また,3.83 (3H, s) よりメトキシ基,7.50 (1H, d, J = 15.9 Hz), 6.59 (1H, d, J = 16.0 Hz) よりトランスオレフィンの存在が示唆された。 以上の結果から文献検索を行った結果, 化合物 2-5 はこれまでに月桃より単離さ れている styrylpyrone である 5,6-dehydrokawain (DK) と推定した。1 H NMR およ び13 C NMR スペクトル (Fig. 2-11) のケミカルシフトが Itokawa らによる報告値 22と一致した。13 C NMR スペクトルのケミカルシフトを Table 2-1 に示した。
16
Fig. 2-9. IR spectrum of compound 2-5.
Fig. 2-10. Chemical structure and 1H NMR spectrum (CDCl3, 400 MHz) of
17
Fig. 2-11. 13C NMR spectrum (CDCl3, 100 MHz) of compound 2-5.
Table 2-1. Comparison of 13C NMR (δ ppm) spectral data of the literature value with those of compound 2-5 Carbon assignment 5,6-Dehydrokawain * 2-5 1 - - 2 158.5 158.6 3 88.8 88.8 4 171.0 171.1 5 101.3 101.3 6 163.9 164.0 7 118.5 118.5 8 135.6 135.8 9 135.1 135.2 10 127.4 127.4 11 128.8 128.9 12 129.3 129.4 13 128.8 128.9 14 127.4 127.4 4-OMe 55.9 55.9
18
2-3. 化合物 2-6 の構造決定
化合物 2-6 の分子式は,HRMS (ESI): [M+H]+
calcd for C14H15O3, 231.1021;
found, 231.1024 より,C14H14O3であった。IR スペクトル (Fig. 2-12)よりカルボ
ニル (1702 cm-1)および芳香環 (3080, 1648, 1564 cm-1)の存在が示唆された。 1 H-NMR スペクトルにおいて,化合物 2-5 と同様に δ 7.31-7.16 (5H, m) より 1 置換ベンゼン環,5.72 (1H, s), 5.41 (1H, s), 3.77 (3H, s)よりメトキシ基を有する ピロン環の存在が示唆された (Fig. 2-13)。しかし,2-5 においてオレフィン由来 と推察されたシグナルが消失し,かわりにメチレン由来と推察されるシグナル 2.97 (2H, t, J = 7.8 Hz), 2.75 (2H, t, J = 8.0 Hz)が観測されたことから,2-5 の 7,8 位が還元された構造であると推定した。以上より,化合物 2-6 は,これまでに月 桃より単離されている dihydro-5,6-dehydrokawain (DDK)と推定した。1 H-NMR ス ペクトルおよび13 C-NMR スペクトル (Fig. 2-14)が Itokawa らによる報告値22と 一致した。13 C-NMR スペクトルのケミカルシフトを Table 2-2 に示した。
19
Fig. 2-13. Chemical structure and 1H NMR spectrum (CDCl3, 400 MHz) of 2-6.
20
Table 2-2. Comparison of 13C NMR (δ ppm) spectral data of the literature value with those of compound 2-6 Carbon assignment Dihydro- 5,6-dehydrokawain* 2-6 1 - - 2 164.2 164.3 3 87.6 87.7 4 171.0 171.1 5 100.1 100.2 6 164.7 164.9 7 35.8 35.4 8 32.8 32.8 9 139.8 139.8 10 128.2 128.3 11 128.5 128.6 12 126.3 126.4 13 128.5 128.6 14 128.2 128.3 4-OMe 55.8 55.8
* Itokawa H. et al. Phytochemistry 1981, 20, 2503-2506.
2-4. 単離した化合物の抗炎症作用 NO 産生抑制活性を指標として分画を行った結果,化合物 2-5 および 2-6 を単 離した。MS および NMR 解析の結果,これらはこれまでに月桃から報告のある 5,6-dehydrokawain (DK, 2-5)および dihydro-5,6-dehydrokawain (DDK, 2-6)である ことが明らかとなった。単離した DK と DDK について NO 産生抑制試験を行っ た結果,共に NO 産生抑制効果が認められた(Fig. 2-15)。このうち,DK につい ては月桃と同様にショウガ科の植物である Alpinia pricei Hayata から抗炎症作用 を有する化合物として単離されており,LPS 刺激した RAW264.7 細胞に対する NO 産生抑制作用や in vivo における抗炎症作用とその作用機構についての詳細 な検討が報告されている23。本研究において,効果は DK と比較して弱いものの,
21 Cont rol 50 100 50 100 0 50 100 150
(
M)
DK (2-5) DDK (2-6)
**
**
**
NO
p
ro
du
c
ti
o
n
(
%
)
Fig. 2-15. NO inhibitory activities of compounds 2-5 and 2-6. Cells were treated with the compounds (50 and 100 µM) and LPS (100 ng/mL) for 24 h and NO production was measured using the Griess reagent. Data shown are mean ± SD (n = 3). **P < 0.01vs. Control by Dunnett’s test.
今回単離した DK および DDK の類縁体(Fig. 2-16)は,亜熱帯および熱帯域に 生息する Piper methysticum の根や根茎の水抽出エキスから報告されており, kava あるいは kava-kava と呼ばれ,陶酔感をもたらす飲料としてポリネシアやミ クロネシアなどの太平洋の島々の人々に飲用されてきた 24。Kava エキスには抗 不安作用がある事が報告されており 25,この活性に寄与する成分が styrylpyrone であることが知られている。また,月桃に含まれる DK および DDK についても これまでにインフルエンザウイルスの増殖に関与しているノイラミニダーゼの 阻害作用 26 や,糖尿病やアテローム性動脈硬化症の発症を悪化させるといわれ ている終末糖化産物の産生抑制作用 27 などが報告されている。月桃は沖縄地方 では親しまれている機能性食品素材であり,ムーチーと呼ばれる月桃の葉でも ち米を包み蒸したお菓子 26 や,月桃茶として葉の部位を煎じたものが飲用され ている。今後,DK や DDK の新しい機能性を探ることは,これまでにない新た な機能性食品の開発につながる可能性がある。また,その際に安全性や詳細な 作用機構を明らかにすることが求められる。
22
23
第 3 節 サルカケミカン(Toddalia asiatica (L.) Lam.)からの
抗炎症化合物の単離・構造決定
1. 目的 第 1 節のスクリーニングにより,サルカケミカンの MeOH 抽出物が抗炎症作 用を示すことが明らかとなった。しかし,これまでに NO 産生抑制活性を指標 としてサルカケミカンから活性化合物を単離した例は報告されていない。そこ で,サルカケミカンの抗炎症物質同定に向けて,NO 産生抑制作用を指標とした 抽出物の分画と活性物質の単離・構造決定を行うこととした。また,得られた 化合物について作用機序解明を目的として研究を行った。 2. 結果と考察 2-1. サルカケミカンからの抗炎症活性物質の単離 サルカケミカンから抗炎症活性物質を単離同定する目的で,RAW264 細胞に おける NO 産生抑制作用を指標とした分画を試みた (Fig. 2-17)。サルカケミカ ンを MeOH で抽出し,得られた抽出物を Hexane,EtOAc,BuOH,水で分配し た。得られた各抽出物について NO 産生抑制効果を調べたところ,Hexane 抽出 物と EtOAc 抽出物に活性が認められた(Fig. 2-18)。そこで効果の強かった EtOAc 抽出物をさらに DIAION HP20 を担体に用いた逆相オープンカラムクロマトグラ フィーに供し,得られたフラクションの濃縮物(E. Fr-1~5)について,NO 産生抑 制効果を調べたところ,E. Fr-4 に最も強い効果が認められた(Fig. 2-19)。E. Fr-4 について,逆相 HPLC を用いて含有物の分析を行ったところ,保持時間が 6.1 min, 16.8 min, 58.5 min に主なピークが認められた (Fig. 2-20)。他の分画物について も同様に逆相 HPLC で分析したところ,収量の多い H. Fr-3 にこれらと同一の保 持時間のピークが認められた。そこで H. Fr-3 から 16.8 min, 58.5 min のピークに 相当する部分を逆相 HPLC で精製し,化合物 2-12 (199.3 mg), 2-13 (12.8 mg)を得 た。24
Fig. 2-17. Separation scheme of Toddalia asiatica (L.) Lam.
Cont rol 5 10 20 5 10 20 5 10 20 5 10 20 0 50 100 150
Hex. Ext. EtOAc Ext. BuOH Ext. Water Ext.
g/mL
N
O
pr
odu
ct
io
n
(%
)
Fig. 2-18. NO inhibitory activities of separated T. asiatica. extract.
Cells were treated with extracts (5, 10, 20 µg/mL) and LPS (100 ng/mL) for 24 h and NO production was measured using the Griess reagent. Data shown are mean ± SD (n = 3).
25 Cont rol 5 10 20 5 10 20 5 10 20 5 10 20 5 10 20 0 50 100 150 g/mL E. Fr-1 E. Fr-2 E. Fr-3 E. Fr-4 E. Fr-5
NO
p
ro
du
c
ti
o
n
(
%)
Fig. 2-19. NO inhibitory activities of E. Fr-1~5.
Cells were treated with each fraction (5, 10, 20 µg/mL) and LPS (100 ng/mL) for 24 h and NO production was measured using the Griess reagent. Data shown are mean ± SD (n = 3).
26 2-2. 化合物 2-12 の構造決定 化合物2-12 の分子式は,HRMS (ESI): [M+H]+ calcd for C16H19O5, 291.1233; found, 291.1218 より,C16H18O5であった。COSY スペクトルによって C-3 と C-4 および C-1'と C-2'を繋げた(Fig. 2-21)。HMBC スペクトルにおいてメトキシ基の H-9 から C-4a, C-5, C-6 への相関,H-10 から C-6, C-7, C-8 への相関,H-4 から C-2, C-4a, C-8a への相関,H-1'から C-6, C-2', C-3'への相関が観測されたことより, Fig. 2-21 に示した平面構造を確認することができた。文献調査の結果,本化合物は これまでに Ishii らによって単離されていたが28,唯一13 C NMR のケミカルシフ トが記載されている文献29とケミカルシフトの比較を行ったところ(Table 2-3), 文献における帰属表の 3 位と 6 位の帰属が逆になっている可能性が示唆された。 HSQC スペクトルにおいて H-3 (6.25 ppm)と C-3 (112.5 ppm)との相関が確認で きたことより(Fig. 2-22), 今回の帰属が正しいものと思われる。また,比旋光度 の測定値[α]D27 −14.7 (c = 0.1 in AcOEt)が Ishii らの報告値28と一致したことより, C-2'の絶対立体配置を R と推定した。また,NOESY スペクトルの解析結果より, C-4', C-5'位の帰属を行うことができた (Fig. 2-23)。今回の帰属結果と文献値と の比較を Table 2-3 に示す。
27
Fig. 2-22. HSQC spectrum (CDCl3, 600 MHz) of 2-12.
28
Table 2-3. Comparison of 13C NMR (δ ppm) spectral data of the literature value with those of compound 2-12 Carbon assignment Aculeatin * 2-12** 2 161.0 161.1 3 116.7 112.5 4 138.8 138.9 4a 107.1 107.2 5 156.0 156.1 6 112.4 116.8 7 161.7 161.8 8 95.4 95.5 8a 155.1 155.2 9 63.5 63.6 10 56.0 56.1 1' 23.5 23.5 2' 63.1 63.2 3' 59.1 59.1 4' 24.7 24.8 5' 18.9 18.9
* Nyahanga, H. et al. J. Chem. Sci. 2013, 125, 1115-1121.
29 2-3. 化合物 2-13 の構造決定 化合物 2-13 の分子式は,HRMS (ESI): [M+H]+ calcd for C16H19O4, 275.1283; found, 275.1270 より,C16H18O4であった。1H-NMR スペクトルにおける δ 7.87 (1H, d, J = 9.6 Hz, H-4), 6.62 (1H, s, H-8), 6.23 (1H, d, J = 9.6 Hz, H-3)と 2 つのメトキ シ基のケミカルシフトが 2-12 と類似していたことより,2-13 もクマリン骨格を 持つ化合物であり,側鎖部分の構造が 2-12 と異なると推察された(Fig. 2-24)。 さらに,推定した分子式より,2-12 よりも酸素原子が1つ少ない構造が示唆さ れ,1 H NMR スペクトルの δ 5.14 (1H, m, H-2')のシグナルがみとめられたことよ り,化合物 2-12 のエポキシ基がオレフィンとなった構造であると推定した。13 C NMR スペクトルの測定値は Nyahanga らによる toddaculin の報告値28と一致し (Table 2-4),2-13 を toddaculin と決定した。また,NOESY スペクトルの解析よ り,H-4'と H-2'および H-5'と H-1'の相関が観測され,C-4'と C-5'の帰属が明らか となった(Fig. 2-24)。
30
Table 2-4. Comparison of 13C NMR (δ ppm) spectral data of the literature value with those of compound 2-13 Carbon assignment Toddaculin * 2-13** 2 161.3 161.3 3 112.2 112.2 4 139.0 139.0 4a 107.0 107.1 5 155.1 155.2 6 120.2 120.3 7 161.6 161.7 8 95.3 95.4 8a 154.6 154.6 9 63.1 63.1 10 56.0 56.0 1' 22.6 22.7 2' 122.1 122.1 3' 132.0 132.0 4' 25.7 25.7 5' 17.8 17.8
* Nyahanga, H. et al. J. Chem. Sci. 2013, 125, 1115-1121.
** The 13C NMR signals were assigned by the HSQC, HMBC, and NOESY spectra.
2-4. 単離した化合物の抗炎症作用
サルカケミカンから活性を指標として単離した化合物 2-12, 2-13 について NO 産生抑制効果を調べたところ,aculeatin (ACU, 2-12)と toddaculin (TOD, 2-13)に NO 産生抑制活性が認められた (Fig. 2-25) 。サルカケミカンからは,これまで に様々な化合物が報告されており,抗菌活性30,抗マラリア活性29,抗腫瘍活性
31,32,抗糖尿病活性33など多岐にわたる活性が知られている。サルカケミカンの
産生する主要な 2 次代謝産物としては,今回単離同定したクマリン類が知られ
31
する inducible-nitric oxide synthase (iNOS)のタンパク発現抑制効果を,RAW264.7 細胞を用いて調査している36。その中で,ACU および TOD の iNOS 抑制効果お
よび TOD の NO 産生抑制効果については明らかとなっていたが,ACU について の NO 産生抑制効果は調べられていなかった。本研究によって,サルカケミカ ン中の NO 産生抑制物質として ACU が初めて明らかとなった。しかし,これら のクマリンの詳しい作用機序については明らかにされておらず,今後 ACU およ び TOD について抗炎症作用を有する薬剤として利用・開発を進めるためにはさ らなる作用機序の解明が望まれた。 Cont rol 25 50 100 200 25 50 100 200 0 50 100 150 Aculeatin (2-12) Toddaculin (2-13) M
NO
p
ro
du
c
ti
o
n
(
%
)
Fig. 2-25. NO inhibitory activities of compounds 2-12 and 2-13.
Cells were treated with each compound (25, 50, 100, 200 µM) and LPS (100 ng/mL) for 24 h and NO production was measured using the Griess reagent. Data shown are mean ± SD (n = 3).
32
第 4 節 Aculeatin と toddaculin の抗炎症作用機序
1. 目的
本章におけるこれまでの研究により,サルカケミカンに含まれる aculeatin (ACU)と toddaculin (TOD)が RAW264 細胞に対して NO 産生抑制作用を示すこ とが明らかとなった。これらの化合物について,抗炎症作用を有する薬剤とし ての開発を推進するために,より詳細な作用機構の解明が必要である。サルカ ケミカンからは,これまでに様々なクマリン化合物が報告されており,抗菌活 性30,抗マラリア活性29,抗腫瘍活性31,32,抗糖尿病活性33など多岐にわたる活
性が知られている。一方 Watanabe らは ACU と TOD について,in vitro の試験系 において ACU が脂肪細胞の分化促進作用37,TOD が骨代謝改善作用38を有する ことを明らかにしている。ACU と TOD はプレニル側鎖のエポキシドの有無によ り,異なる作用機構を持つことが示唆されており,構造活性相関の観点からも, これら 2 つの化合物の作用機構の違いについて詳細に解析することでクマリン の活性発現機構に関する新たな知見が得られると考えられた。そこで本節では, これらの抗炎症作用に着目し,活性発現機構の違いについて詳細に解析するこ とを目的とした。 今回 ACU と TOD の抗炎症作用機序解析に用いるマウスマクロファージ様細 胞株 RAW264 細胞は,グラム陰性菌の細胞壁成分である lipopolysaccharide (LPS) の刺激により活性化されることが知られている(Fig. 2-26)39,40。その際,LPS が
細胞膜上の Toll-like receptor 4(TLR4)により認識され,Myd88, TRAF6 といった アダプタータンパクの活性化を経由して p38, JNK, ERK1/2 などの MAPK や, IκBα などがリン酸化される。IκBαのリン酸化に伴い核移行し,プロモーター領 域に結合した転写因子 NFκB や,MAPK により活性化された転写因子 AP-1 によ って,iNos, Il1a, Il1b, Mcp1 などの炎症性メディエータの産生が誘導される。本 節では,まず ACU と TOD が LPS 刺激により誘導されるであろう炎症性メディ エータの mRNA 産生を抑制するかを DNA マイクロアレイにより広範に調べた。 その結果,顕著な抑制効果が認められた mRNA について,リアルタイム定量 PCR を用いて検証した。さらにこれらの経路について,ACU および TOD がどのよう に作用するのかを明らかにするため,タンパク質を定量する手法であるウエス タンブロット法や,転写因子の活性を調べる手法であるレポーターアッセイ法
33
を用いて調べることとした。
Fig. 2-26. TLR4 signal transduction pathway.
2. 結果と考察
サルカケミカンより単離した 2 種のクマリン(Fig. 2-27A),ACU および TOD の抗炎症メカニズムに焦点を当てて研究を行った。第 3 節のなかで,ACU およ び TOD が LPS 刺激 RAW264 細胞における NO 産生量を抑制することを明らか にした。そこで,作用機序解明に用いる ACU と TOD の濃度域を確認するため, 細胞毒性を評価したところ,150 μM までの濃度において ACU および TOD が細 胞生存率に影響しないことを確認したため (Fig. 2-27B),以降 100 μM にて作用 機構をより詳細に調べることとした。
34
Fig. 2-27. Chemical structures of aculeatin and toddaculin (A). Effects of aculeatin and toddaculin on cell viability (B) in LPS-stimulated RAW264 cells. Cells were treated with compounds and LPS (100 ng/mL) for 24 h and cell viability was measured by WST-8 assay (B). Data shown are mean ± SD (n = 3).
RAW264 細胞は LPS の刺激により TNF-α や IL-6 といった炎症性サイトカイン を産生することが知られている。そこで,本濃度域において,ACU および TOD は,LPS 刺激により誘導される TNF-α と IL-6 のタンパク産生を抑制するかを調 べたところ,その産生を濃度依存的に抑制した(Fig. 2-28)
35 0 10 20 30 T NF - p ro d u c ti o n ( n g /m L ) LPS - + + + + + + + ACU (M) 25 50 100 TOD (M) - - - - - 25 50 100 a b c d e f g h 0 50 100 150 LPS - + + + + + + + ACU (M) 25 50 100 TOD (M) - - - - - 25 50 100 a b b c c d d d IL -6 p ro d u c tio n ( n g /m L )
Fig. 2-28. Effects of ACU and TOD on LPS-induced TNF-α and IL-6 production in RAW264 cells. Cells were treated with compounds and LPS (100 ng/mL) for 24 h. Then TNF-α and IL-6 content in the culture medium were analyzed using ELISA kit. Data shown are mean ± SD (n = 3). The different letters were significantly different by Tukey-Kramer test (P < 0.05). 次に, RAW264 細胞における LPS 刺激によってどのような炎症メディエータ の遺伝子発現が上昇し,さらに ACU および TOD がどのような遺伝子発現を強 く抑制しているのかを広範に調べるため,DNA マイクロアレイ解析を行った。 その結果,LPS 刺激により顕著に発現上昇が認められた mRNA のうち,ACU と TOD (100 μM)で 24 時間処理するにことにより,炎症性ケモカイン,サイトカ イン(Mcp-1, Il6, Il1a および Il1b)および炎症性酵素(Cox2 および iNos)の発現が顕 著にダウンレギュレートされた(結果は示していない)。これらの遺伝子発現を リアルタイム qPCR で再検証したところ,これらの遺伝子発現が用量依存的に ACU および TOD によって同程度に減少することを確認した(Fig. 2-29)。
36 0 50 100 150 Mcp1 LPS - + + + + + + + ACU( M) 25 50 100 TOD( M) - - - - - 25 50 100 A a b c d e f e f m RNA (% ) 0 50 100 150 iNos B LPS - + + + + + + + ACU( M) 25 50 100 TOD( M) - - - - - 25 50 100 a b b c c c c c m R N A (% ) 0 50 100 150 Il6 C mR N A (% ) LPS - + + + + + + + ACU( M) 25 50 100 TOD( M) - - - - - 25 50 100 a b b c cd d d d 0 50 100 150 Il1a D m R N A (% ) LPS - + + + + + + + ACU( M) 25 50 100 TOD( M) - - - - - 25 50 100 a b c c c c c c 0 50 100 150 Il1b E mRNA (% ) LPS - + + + + + + + ACU( M) 25 50 100 TOD( M) - - - - - 25 50 100 a b bc c c c c c 0 50 100 150 Cox2 F mRNA (% ) LPS - + + + + + + + ACU( M) 25 50 100 TOD( M) - - - - - 25 50 100 a b a b c cd d d
Fig. 2-29. Effects of ACU and TOD on mRNA expression [(A) Mcp1, (B) iNos, (C) Il6, (D) Il1a, (E) Il1b, and (F) Cox2] in RAW264 cells. Cells were treated with compounds and LPS (100 ng/mL) for 24 h, and mRNA levels were measured using qPCR. Data shown are mean ± SD (n = 3). The various letters indicate significant differences in Tukey-Kramer test results (P < 0.05).
以上の結果より, LPS 刺激によるマクロファージの活性化によって生じる炎 症性メディエータの産生を,ACU と TOD が遺伝子レベルで抑制していることが 示唆された。
37 ACU および TOD の抗炎症活性のさらなる作用機構を明らかにするために, ウ エスタンブロットを用いて MAPK リン酸化に対するこれら 2 つの化合物の効果 を評価した(Fig. 2-30)。 その結果,TOD が 15 分で p38 リン酸化を有意に抑制 したが,ACU にはそのような効果はみとめられなかった。 TOD は、15 分およ び 30 分で ERK1 / 2 リン酸化を有意に抑制したが,ACU は有意な効果を示さな かった。 一方 LPS 刺激後 15 分で TOD による JNK のリン酸化促進効果が認め られたが,ACU にはそのような効果は認められなかった。
Fig. 2-30. Effects of ACU and TOD on LPS-induced phosphorylation of MAPKs in RAW264 cells. Cells were treated with compounds (100 µM) and LPS (100 ng/mL) for 15 and 30 min. Then, cells were lysed and analyzed by western blotting. Data shown are means ± SD (n = 3–4). *P < 0.05 vs. LPS-stimulated cells in each time by Dunnett’s test.
38 0 50 100 150
LPS - + + + + + + +
ACU(
M ) 25 50 100
TOD(
M) - - - - - 25 50 100
* *
NF
-
B
a
c
ti
v
it
y
(
%
)
Fig. 2-31. Effects of ACU and TOD on LPS-induced NF-κB activation. Luciferase reporter assay was performed as described in the experimental section . Data shown are means ± SD (n = 3).*P < 0.05 vs. LPS-stimulated cells by Dunnett’s test. 次に,炎症性メディエータの遺伝子発現を制御している転写因子である NF-κB に対する ACU および TOD の効果を調べるために, NF-κB ルシフェラーゼレポー ターアッセイを行った(Fig. 2-31)。 LPS 処理により,NFκB の活性は 10 倍以上 上昇した。 LPS のみを添加したコントロールに対し,TOD は 50 µM 以上の濃 度においてNFκB 活性を有意に抑制したが,ACU には有意な抑制効果は認めら れなかった。 これらの結果から、TOD の抗炎症作用は,p38,ERK1 / 2 および NFκB 経路の 抑制作用を介した経路で発現していると考えられたが,ACU は異なるメカニズ ムで抗炎症作用を示す可能性が示唆された。NFκB 経路は破骨細胞分化にも関与 していることが知られており41,この経路を抑える天然物が破骨細胞分化を抑制 することが報告されている 42。過去の研究より,TOD が破骨細胞に対する分化 抑制効果を有することが明らかとなっている38。今回の結果は, TOD が破骨細 胞分化抑制効果を示すことを支持する結果であると考えられた。ACU と TOD の 構造の違いは,プレニル側鎖のエポキシ基の有無であるが,その部分の違いの
39
みによりこのような作用機構に違いが認められることは興味深い。
ACU と TOD の作用機序の違いの理由をさらに詳しく評価する目的で,これら の化合物の細胞への取り込み量を検討した(Fig. 2-32)。 TOD は,4 時間後(129 pmol / 1.0×10 6細胞)および 24 時間後(177 pmol / 1.0×10 6細胞)の RAW264 細胞で 検出されたが,ACU の取り込み量は,いずれのインキュベーション時間におい ても 20 pmol / 1.0×10 6細胞未満であった。 0 50 100 150 200 250
ACU TOD ACU TOD 4 h 24 h p m o l A C U o r T O D /1 0 6 ce lls N.D. N.D.
Fig. 2-32. Uptake levels of ACU and TOD in RAW264 cells. Cells were incubated with each compound for 4 or 24 h and then extracted and analyzed by HPLC. Data were expressed as mean ± SD (n = 3). N. D. = Not detectable.
TOD および ACU の logP 値がそれぞれ 3.17 および 1.91 であることから(CS Chem Draw Ultra 6.0, Cambridge Soft Corporation, MA, USA), プレニル基のエポ キシ化により疎水性が低下すると考えられる。 フラボノイド類などは,プレニル化を受けることで細胞への浸透性が増加す るとともに,動物試験においては血中における存在時間の延長や組織への蓄積 量の増加が認められることが知られている 43-45。ACU より疎水性の高い TOD は細胞膜を透過し,p38MAPK や ERK1/2 のリン酸化を抑制し,NFκB の転写活 性を抑制することにより抗炎症作用を発現すると予想された(Fig. 2-33)。しかし, ACU は TOD と同様に抗炎症作用を示すものの,その作用機構は TOD とは別の
40
経路を介したものである可能性が示唆された。
Fig. 2-33. Expected action mechanisms of ACU and TOD in this study.
ACU は,マウス由来 3T3-L1 細胞において,細胞表面の β アドレナリンレセプ ターに作用し,脂肪分解促進効果を発現する可能性が示唆されている46。また,
緑茶などに含まれるepigallocatechin gallate (EGCG)が,細胞表面の 67kD ラミ ニンレセプターに結合し抗炎症作用を示すことが報告されている47,48。細胞内へ
の浸透量が少ないと考えられる ACU は,このように細胞膜表面のタンパク質と の相互作用によって抗炎症作用を示している可能性も考えられる。今後 ACU の さらなる作用機序解明が望まれる。
41
第 5 節 実験項
1. 使用機器
マイクロプレートリーダーは,Molecular Devices 社製 Spectra Max M2 を 使用した。高速液体クロマトグラフィー(HPLC)は,分析および分取用ともに島 津製作所製のものを用いた。システムコントローラー (CBM-20A), オートサン プラー (SIL-10AP), デガッサー (DGU-20A), ポンプ (LC-20AR), 紫外可視 検出器 (SPD-20A)で分析および分取を行った。赤外吸収スペクトルの測定は顕 微透過ユニットIRT-5000 を装着した FT/IR-4100 (日本分光)を用いた。核磁気 共鳴 (NMR)装置は,JEOL JNM-ECZ400S, Bruker Biospin Avance III HD 600 を用いた。旋光度計は,JASCO DIP-370 digital polarimeter を用いた。また,高分 解能質量分析 (HRMS)は,Thermo scientific Q exactive を用いて測定した。 2. 試薬
カラムクロマトグラフィーには,Diaion HP-20 (三菱ケミカル), Wakogel 60N (和光純薬工業)を用いた。DMEM 培地 (high-glucose) および penicillin streptomycin は,Sigma-Aldrich より購入した。FBS と GlutaMAXTM は,Life Technologies より購入した。N-(1-Naphthyl)ethylenediamine・2HCl,sulfanilamide, 0.5 mol/L 亜硝酸 Na 溶液,LPS (E.coli O55:B5 由来) は和光純薬工業より購入 した。その他の試薬については,可能な限り最上級のグレードを使用した。 3. 細胞培養
RAW264 細胞は,RIKEN Cell Bank (Tsukuba, Japan)より購入した。細胞 は,10% FBS, 100 units/mL penicillin, 100 µg/mL streptomycin を添加した DMEM 培地にて培養した。サンプルは,DMSO に溶解したものを用い,DMSO の終濃度が0.1% (v/v) 以下となるように培地に添加した。培養中,培地を 2, 3 日おきに交換した。細胞は,5% CO2, 37 ℃の条件で培養した。
4. 第 1 節の実験 4-1.サンプルの調製
42
asiatica), マカ (Lepidium sp.),パパイヤ(Carica papaya), ハイビスカス(Hibiscus hybrids), グァバ(Psidium sp.), クコ(Lycium chinense), クワンソウ(Hemerocallis fulva), ク ミ ク ス チ ン ( Orthosiphon stamineus), ヤ エ ヤ マ ア オ キ ( Morinda citrifolia),アマチャヅル(Gynostemma pentaphyllum)は,2014 年 4 月に沖縄県薬
草協同組合より乾燥物を提供いただいた。ニトベギク(Tithonia diversifolia), ホ オズキ(Physalis sp.), 月桃(Alpinia zerumbet), クービー(Elaeagnus glabra), イ シマーチは,株式会社 比嘉製茶より乾燥物を購入した。ボタンボウフウ (Peucedanum japonicum), ウズラ豆(Phaseolus vulgaris),ウズラ豆皮(Phaseolus
vulgaris),ニガナ,ナスタチウム(Tropaeolum majus),ササゲ(Vigna unguiculata),
エンサイ(Ipomoea aquatica)は,2014 年 4 月にJAおきなわ 中部ファーマーズ マーケット ちゃんぷる~市場にて生鮮物を購入した。生鮮物は,粉砕後,凍 結乾燥した。各サンプルを粉砕後,MeOH で抽出し,ろ過したのちに溶媒を除 去し凍結乾燥した。凍結乾燥後,50 mg/mL の濃度となるよう DMSO に溶解し, スクリーニング用サンプルとした。 4-2. NO 産生抑制試験 RAW264 細胞を 96 ウェルプレート(1.0×105 cells/well)に播種した。各サン プルを終濃度が 50 µg/mL となるよう培地に溶解して細胞に添加し,LPS(100 ng / mL)を添加して 24 時間培養した。 インキュベーション後,採取した培地サン プルを Griess 試薬(1% sulfanilamide,0.1% N-(1-Naphthyl)ethylenediamine・2HCl および 2.5%リン酸)と混合し, 37 ℃, 20 分間インキュベートした。インキュベ ーション後,540 nm での吸光度をマイクロプレートリーダーで測定し, 亜硝酸 ナトリウムを用いた標準曲線と比較した。 細胞生存率の評価のために,水溶性 テトラゾリウム塩(WST)細胞毒性アッセイを,Cell Counting Kit-8(Dojindo) を用いて行った。結果は,LPS 添加したコントロールを 100%とした相対値で表 した。 5. 第 2 節の実験 5-1. 化合物の単離・構造決定 化合物の分離は,第 2 節で述べたとおり行った。NMR の測定溶媒は,重水素 化クロロホルム (TMS 0.05 wt%入り)を用い,内部標準物質として TMS (1 H: 0.00
43 ppm), 重クロロホルム (13C, 77.0 ppm)を用いた。 5,6-Dehydrokawain (2-5): 1H NMR (400 MHz, CDCl3) δ 7.52-7.35 (5H, m, aromatic), 7.50 (1H, d, J = 15.9 Hz, 8-H), 6.59 (1H, d, J = 16.0 Hz, 7-H), 5.95 (1H, d, J = 2.4 Hz, 5-H), 5.50 (1H, d, J = 2.0 Hz, 3-H), 3.83 (3H, s, MeO-); 13C NMR (100 MHz, CDCl3) δ 171.1, 164.0, 158.6, 135.8, 135.2, 129.4, 128.9, 127.4, 118.6, 101.3, 88.8, 55.9; IR (ATR) 3080, 1723, 1639, 1608, 1559, 1148, 958, 890, 832, 755 cm-1; HRMS (ESI): [M+H]+ calcd for C14H13O3, 229.0865; found, 229.0866.
Dihydro-5,6-dehydrokawain (2-6): 1H NMR (400 MHz, CDCl3) δ 7.31-7.16 (5H, m,
aromatic), 5.72 (1H, s, 5-H), 5.41 (1H, s, 3-H), 3.77 (3H, s, MeO-), 2.97 (2H, t, J = 7.8 Hz, CH2), 2.75 (2H, t, J = 8.0 Hz, CH2); 13C NMR (100 MHz, CDCl3) δ 171.1, 164.9,
164.3, 139.8, 128.6, 128.3, 126.4, 100.2, 87.7, 55.8, 35.4, 32.8; IR (ATR) 3080, 1702, 1648, 1568, 1128, 1138, 942, 862, 842, 818, 743 cm-1; HRMS (ESI): [M+H]+ calcd for C14H15O3, 231.1021; found, 231.1024. 5-2. NO 産生抑制試験 RAW264 細胞を 96 ウェルプレート(1.0×105 cells/well)に播種した。各サン プルを終濃度が分画物の場合 25 もしくは 50 µg/mL となるよう培地に溶解して 細胞に添加した。単離した化合物は 50, 100 µM となるように培地に溶解して細 胞に添加した。サンプル添加後,LPS(100 ng / mL)を添加して 24 時間培養し た。培養後の処理は,4-2 と同様に行った。 5-3. 統計解析
統計解析は,GraphPad Prism V5.02 softoware for Windows (Graphpad Softoware) を用い,一元配置分散分析と Dunnett の多重比較検定に供した。5%未満の危険 率で,コントロールとの有意差の有無を判定した。 6. 第 3 節の実験 6-1. 化合物の単離・構造決定 沖縄県薬草協同組合より入手したサルカケミカン(乾燥茎)を,ミルで粉 砕後,100 g を 1 L の MeOH で 80 ℃,2 時間加熱還流抽出した。この操作をも
44 う一度繰り返し,MeOH 抽出液を減圧化で濃縮乾固後,重量を測定した(10.5 g)。 MeOH 抽出物を 100 mL の水で懸濁し,Hexane (100 mL)を加えて,振とう抽出 した。この操作を4回繰り返し,Hexane 抽出液を濃縮乾固後,重量測定した( 1.75 g)。残った水層に EtOAc (100 mL)を加えて振とう抽出し,この操作を4回繰 り返した。さらに,残りの水層に BuOH (100 mL)を加えて振とう抽出し,この 操作を4回繰り返した。残りの水層,EtOAc 抽出液,BuOH 抽出液は,それぞれ 濃縮乾固して重量を測定した (水抽出物: 1.75 g, BuOH 抽出物: 1.12 g, EtOAc 抽 出物: 0.63 g)。
EtOAc 抽出物の一部 (217 mg)を Diaion HP-20 (Mitsubishi Chemical, Tokyo, Japan)を用いたオープンカラムクロマトグラフィーに供し,20% MeCN (E. Fr-1), 40% MeCN (E. Fr-2), 60% MeCN (E. Fr-3), 80% MeCN (E. Fr-4), 100% MeCN (E. Fr-5)で溶出した。溶出液を濃縮乾固後,重量測定した(E. Fr-1: 16.8 mg, E. Fr-2: 56.7 mg, E. Fr-3: 45.6 mg, E. Fr-4: 29.5 mg, E. Fr-5: 25.4 mg)。Hexane 抽出物は, DiaionHP-20(φ 3.0×140 mm)に供し,20% MeCN (H. Fr-1), 40% MeCN (H. Fr-2), 60% MeCN (H. Fr-3), 80% MeCN (H. Fr-4), 100% MeCN (H. Fr-5)で溶出した(各 200 mL)。溶出液を濃縮乾固後,重量測定した(H. Fr-1: 19 mg, H. Fr-2: 28 mg, H. Fr-3: 423 mg, H. Fr-4: 604 mg, H. Fr-5: 112 mg)。H. Fr-3 を逆相 HPLC (Detection: 210 nm, Solvent: MeCN/H2O (50:50), Flow rate: 5 mL/min, Column: Wakosil-Ⅱ
5C18RSPrep φ20×250 mm)で分取し,化合物 2-12 (Rt: 15.6 min,199.3 mg),化合物 2-13 (Rt: 47.7 min, 12.8 mg)をそれぞれ白色固体として単離した。 Aculeatin (2-12): 1H-NMR (600 MHz, CDCl3) δ 7.88 (1H, dd, J = 9.5, 0.2 Hz, H-4), 6.65 (1H, s, H-8), 6.25 (1H, d, J = 9.6 Hz, H-3), 3.91 (3H, s, H-10), 3.87 (3H, s, H-9), 2.98-2.93 (1H, m, H-2'), 2.98-2.86 (2H, m, H-1'), 1.43 (3H, s, H-5'), 1.31 (3H, s, H-4'); 13C-NMR (150 MHz, CDCl3) δ 161.8 (C-7), 161.1 (C-2), 156.1 (C-5), 155.2 (C-8a), 138.9 (C-4), 116.8 (C-6), 112.5 (C-3), 107.2 (C-4a), 95.5 (C-8), 63.6 (C-9), 63.2 (C-2'), 59.1 (C-3'), 56.1 (C-10), 24.8 (C-4'), 23.5 (C-1'), 18.9 (C-5'); IR (ATR) 2917, 1734, 1610, 1456, 1381, 1205, 1130, 1086, 822, 774 cm-1; HRMS (ESI): [M+H]+ calcd for C16H19O5, 291.1233; found, 291.1218; [α]D27 −14.7 (c = 0.1 in AcOEt).
Toddaculin (2-13): 1H-NMR (600 MHz, CDCl3) δ 7.87 (1H, d, J = 9.6 Hz, H-4), 6.62
(1H, s, H-8), 6.23 (1H, d, J = 9.6 Hz, H-3), 5.14 (1H, m, H-2'), 3.89 (3H, s, H-10), 3.83 (3H, s, H-9), 3.35 (2H, d, J = 6.6 Hz, H-1'), 1.78 (3H, s, H-5'),1.68 (3H,s, H-4');
45 13 C-NMR (150 MHz, CDCl3) δ 161.7 (C-7), 161.3 (C-2), 155.2 (C-5), 154.6 (C-8a), 139.0 (C-4), 132.0 (C-3'), 122.1 (C-2'), 120.3 (C-6), 112.2 (C-3), 107.1 (C-4a), 95.4 (C-8), 63.1 (C-9), 56.0 (C-10), 25.7 (C-4'), 22.7 (C-1'), 17.8 (C-5'); IR (ATR) 2959, 1733, 1610, 1564, 1458, 1381, 1356, 1309, 1204, 1136, 1088, 1011, 952, 917, 823 cm-1; HRMS (ESI): [M+H]+ calcd for C16H19O4, 275.1283; found, 275.1270.
6-2. NO 産生抑制試験 RAW264 細胞を 96 ウェルプレート(1.0×105 cells/well)に播種した。各サン プルを終濃度が分画物の場合 5, 10, 20 µg/mL となるよう培地に溶解して細胞に 添加した。単離した化合物は 25, 50, 100, 200 µM となるように培地に溶解して細 胞に添加した。サンプル添加後,LPS(100 ng / mL)を添加して 24 時間培養し た。培養後の処理は,4-2 と同様に行った。 7. 第 4 節の実験 7-1. 細胞毒性試験 RAW264 細胞を 96 ウェルプレート(1.0×10 5cells/well)に播種した。次に各種濃 度で ACU または TOD 処理し, LPS (100 ng / mL)を添加して 24 時間培養した。 イ ンキュベーション後, Cell Counting Kit-8(Dojindo)を用いて, 水溶性テトラゾリ ウム塩(WST)細胞毒性アッセイを行った。
7-2. ELISA 法
RAW264 細胞を 24 ウェルプレート(5×105 cells/well)に播種した。次に各種濃度 で ACU または TOD 処理し,LPS(100 ng / mL)を添加して 24 時間培養した。 イ ンキュベーション後, 採取した培地中の TNF-α および IL-6 の量を Mouse TNF-α ELISA MAXTMと Mouse IL-6 ELISA MAXTM (Biolegend)を用いてそれぞれ定量し た。
7-3. DNA マイクロアレイおよび定量的 RT-PCR
RAW264 マクロファージ細胞から, TRIzol 試薬(Life Technologies)および RNeasy Mini Kit(Qiagen)を用いて Total RNA を抽出した。抽出した Total RNA を用いて Biotin 標識 RNA を作製した後, 183 個の遺伝子を搭載したフォーカス ト DNA マ イ ク ロ ア レ イ 分 析 (Genopal Mouse Mouse innate immunity Chip,
46
Mitsubishi Chemical, Tokyo, Japan)を, 製造者のプロトコールに従って実施した。 リアルタイム定量 RT-PCR のために, random primers および PrimeScript reverse transcriptase (Takara Bio)を用いて cDNA を合成した。 mRNA 発現量の測定には, Fast SYBR Green Master MixTM (Applied Biosystems)を用い,Tabele 2-5 に示した
遺伝子特異的プライマーで標的 cDNA を増幅させ,StepOnePlusTMリアルタイム
PCR システム (Applied Biosystems)にて測定を行った。各種 mRNA 発現量は,比
較 Ct 法により,コントロールの mRNA 発現量に対する各試験区の mRNA 発現量 を mRNA 発現率として算出した。内部標準として β-actin の mRNA 発現量で標準 化を行った。
Table 2-5. Primer pairs used in this study
Gene name Primer Primer sequence (5' to 3')
Actb Forward CATCCGTAAAGACCTCTATGCCAAC
Reverse ATGGAGCCACCGATCCACA
Mcp1 Forward GCATCCACGTGTTGGCTCA
Reverse CTCCAGCCTACTCATTGGGATCA
Nos2 Forward GGAATGGAGACTGTCCCAGCA
Reverse GTCATGAGCAAAGGCGCAGA
Il6 Forward CAACGATGATGCACTTGCAGA
Reverse CTCCAGGTAGCTATGGTACTCCAGA
Il1β Forward TCCAGGATGAGGACATGAGCAC
Reverse GAACGTCACACACCAGCAGGTTA
Il1α Forward TGGTTAAATGACCTGCAACAGGAA Reverse AGGTCGGTCTCACTACCTGTGATG Cox2 Forward GTGTGCGACATACTCAAGCAGGA
Reverse TGAAGTGGTAACCGCTCAGGTG
7-4. NFκB レポーターアッセイ
既 製 品 の 手 順 書 に 従 い , Lipofectamine LTX お よ び PLUS Reagents (Life Technologies Japan) を用いてホタル pGL4.32 /NFκB プラスミドおよび Renilla
47
pGL4.73 / SV40 対照プラスミド(Promega, Tokyo, Japan) を RAW264 細胞へトラ ンスフェクションした。トランスフェクション後の RAW264 細胞(5.0×104
cells/well) を 96 ウェルプレートへ播種し,16 時間培養した。 培養後,細胞を 異なる濃度の ACU および TOD で処理し,次いで LPS(100 ng/mL)を添加した。 6 時間後,細胞を溶解し,Dual luciferase reporter assay system(Promega)を用い てルシフェラーゼ活性を測定した。 値は対照ルシフェラーゼ活性によって標準 化し,コントロールの値を 100%とした相対値で表した。
7-5. ウエスタンブロット
RAW264 細胞を ACU および TOD(100 µM)で 1 時間処理し, 次いで LPS (100 ng /mL)にそれぞれ 15 および 30 分間暴露した。細胞を PBS で 2 回洗浄し,cOmpleteTM Lysis-M および Phos-STOPTM(Roche Diagnostic, Tokyo, Japan)を用いて溶解させた。 溶解物を 13,000 rpm で 5 分間遠心分離し,上清を回収した。タンパク質含量は, Pierce BCA Protein Assay Kit (Thermo Scientific, Rockford, USA)を用いて測定した。 ライセートを 13,000 rpm,4 ℃で 5 分間遠心し,上清を回収した。上清のタンパ ク量は,Pirce BCA Protein Assay Kit (Thermo Scientific)を用いて定量し,サンプル 間で均一となるよう適宜希釈した。ライセートを 2-mercaptoethanol を 5%含有さ せた 2×Lamimli Sample Buffer (Bio-Rad)に溶解させ,100 ℃で 5 分間変性させた。 同量のタンパクを SDS-PAGE にて分離し,PVDF メンブレン (Merck Millipore) に転写した。2% ECL Blocking reagent (GE Healthcare)でブロッキングを行った後, TPBS で各種一次抗体を希釈し,4 ℃で一晩インキュベートした。その後,メン ブレンを TPBS にて 4 回洗浄し,2 次抗体(GE Healthcare)にて室温,1 時間反応 させた。反応後,さらに TPBS で 4 回洗浄し,ECL Prime Western Blotting Detection Reagent (GE Healthcare)を添加し,LAS-4000 Luminescent Image Analyzer (GE Healthcare)を使用して検出した。タンパクの定量は,Multi Gauge version 3.11 software (Fujifilm Life Science)を用いて行った。
7-6. 細胞内取り込み量の定量
RAW 細胞 (3.0×106細胞)を 25 cm2のフラスコに播種し,100 μM の ACU およ び TOD と共に 4 時間および 24 時間インキュベートした。 インキュベーション
48 後,細胞を PBS で 2 回洗浄し,回収した細胞数を血球計算盤を用いて計測した。 1,000 rpm で 2 分間の遠心分離後, 1 mL の MeOH を添加し,細胞を数秒間超音 波処理により粉砕抽出した。 溶液を 10,000 rpm で 10 分間遠心分離し,収集し た上清を N 2ガスで乾燥させた。 乾燥物を 200 μL の HPLC 移動相に溶解し,0.2 μm のメンブレンフィルターを通して濾過した。 次に, 20 μL のサンプルを C18 カラム(Wakosil-Ⅱ5C18RS-Prep, 4.6×250 mm, 和光純薬工業株式会社)に注入し た。 移動相は 50%アセトニトリル(ACU)または 70%アセトニトリル(TOD)を 使用した。 流速は 1 mL/min, UV 波長 310 nm における吸収を検出した。ACU お よび TOD の標準品で検量線を作成し,ピーク面積値を定量に用いた。 7-7. 統計処理
統計解析は,GraphPad Prism V5.02 softoware for Windows (Graphpad Softoware) を用い,一元配置分散分析と Dunnett もしくは Tukey-Kramer の多重比較検定に 供した。5%未満の危険率で,コントロールとの有意差の有無を判定した。