Title 雌ラットにおける生殖機能の加齢性変化の発現メカニズムに関する研究( 本文(Fulltext) ) Author(s) 石井, 美沢 Report No.(Doctoral Degree) 博士(獣医学) 甲第395号 Issue Date 2013-09-24 Type 博士論文 Version ETD URL http://hdl.handle.net/20.500.12099/47359 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
1
雌 ラ ッ ト に お け る 生 殖 機 能 の 加 齢 性 変 化 の
発 現 メ カ ニ ズ ム に 関 す る 研 究
2013 年
岐阜大学大学院連合獣医学研究科
(東京農工大学)
石 井 美 沢
2 目次 第 1 章 緒言 ... 5 1-1 加齢に伴う生殖機能の低下 ... 5 1-2 ラット ... 6 1-3 発情周期に伴う生殖関連ホルモンの変動 ... 6 1-4 kisspeptin ... 8 1-5 胚・胎児の分化・発育 ... 9 1-6 本研究の目的 ... 9 第 2 章 雌ラットにおける加齢に伴う生殖機能の変化 ... 11 1. 背景と目的 ... 11 2. 材料及び方法 ... 11 2.1 使用動物 ... 11 2.2 膣スメアの採取及び発情周期判定 ... 12 2.3 交尾率,受胎率及び胚・胎児の発達 ... 12 2.4 統計解析及びデータの評価 ... 14 3. 結果 ... 14 3.1 発情周期 ... 14 3.2 交尾率,受胎率,黄体数及び着床数 ... 15 3.3 胚・胎児の死亡率 ... 15 3.4 胎児の生存,発育及び形態学的異常 ... 16 4. 考察 ... 17 第 3 章 高週齢ラットの妊娠初期及び中期における胚・胎児の生存・発育に関 す る検討 ... 30 1. 背景と目的 ... 30 2. 材料及び方法 ... 30 2.1 使用動物 ... 30 2.2 交配 ... 30 2.3 黄体数及び着床数の算出並びに着床部位の観察 ... 30 2.4 実体顕微鏡下での観察 ... 31 2.5 統計 ... 32 3. 結果 ... 32 3.1 黄体数及び着床数 ... 32 3.2 着床部位の観察 ... 32 3.3 胚の形態学的検査 ... 33
3 4. 考察 ... 34 第 4 章 視床下部 kisspeptin 神経のエストロゲンに対する活性の低下が引き起 こす加齢性 LH サージの消 失 ... 47 1. 背景と目的 ... 47 2. 材料及び方法 ... 47 2.1 使用動物 ... 47 2.2 血漿サンプルの採取方法 ... 48 2.3 血漿中 LH 及びエストラジオール濃度測定 ... 48
2.4 AVPV における Kiss1 mRNA 発現量及び kisspeptin 含有量の測定 ... 48
2.5 GnRH 神経のヒト kisspeptin(Kp-54)に対する反応 ... 49 2.6 卵巣を摘出したラットにおけるエストラジオールによる AVPV の kisspeptin 神経の活性化 ... 49 2.7 OVX+E2 MI における LH サージ測定 ... 50 2.8 ホルモン測定 ... 50 2.9 Real-time PCR ... 50
2.10 Enzyme immunoassay (EIA) ... 51
2.11 抗 kisspeptin モノクローナル抗体の作製 ... 51
2.12 Two-site EIA ... 51
2.13 Kiss1 mRAN 及び cFos の二重染色 ... 52
2.14 細胞数 ... 53
2.15 統計解析 ... 53
3. 結果 ... 53
3.1 LH サージ及び血漿中エストラジオール濃度測定 ... 53
3.2 視床下部 AVPV における Kiss1 mRNA 発現量及び kisspeptin 含有量の測 定 ... 54 3.3 GnRH 神経の Kp-54 に対する反応 ... 54 3.4 卵巣を摘出したラットにおけるエストラジオールによる AVPV の kisspeptin 神経の活性化 ... 54 3.5 OVX+E2 MI における LH サージ測定 ... 55 4. 考察 ... 55 第 5 章 異常発情周期を発症した高週齢ラットの交尾排卵機構に関する検討 .. 68 1. 背景と目的 ... 68 2. 材料と方法 ... 68 2.1 使用動物 ... 68 2.2 交尾能,受胎率,黄体数及び着床数 ... 69
4 2.3 交尾後の LH サージ ... 69 2.4 MI ラットにおけるヒト絨毛性性腺刺激ホルモン( Human Chorionic Gonadotropin: hCG)及び Kp-54 の投与による排卵誘起... 69 2.4 MI ラットの卵胞における LH 受容体遺伝子の発現局在 ... 70 2.5 統計 ... 71 3. 結果 ... 71 3.1 交尾率,受胎率,黄体数及び着床数 ... 71 3.2 MI ラットにおける交尾後の LH サージ ... 71 3.3 MI ラットにおけるヒト絨毛性性腺刺激ホルモン及び Kp-54 の排卵作用.. 72 3.4 MI ラットの卵胞における LH 受容体遺伝子(Lhr)の発現局在 ... 72 4. 考察 ... 72 総合考察 ... 81 総括 ... 89 謝辞 ... 93 参考文献 ... 94
5 第 1 章 緒言 1-1 加齢に伴う生殖機能の低下 生殖機能は,哺乳類を含む全ての動物において子孫を残すために重要な機能 である。哺乳類では,一倍体の生殖細胞が異性の持つ生殖細胞と受精すること で,親とは異なる新たな 2 倍体を持つ受精卵を形成し多様性を獲得することで 進化してきた。 ヒトを初めとする多くの哺乳類は,加齢に伴い生殖機能が低下することが知 られており,月経周期,内分泌,卵細胞の染色体及び胚・胎児に異常をきたす ことが報告されている(1-3)。Menken らの報告によると,ヒトでは 30 歳以上で 受胎率が低下しはじめ,35 歳以上で不妊率が 30-50%となり,平均して 41 歳で 不妊になるという(4)。近年,我が国では女性の社会進出に伴い,合計特殊出生 率(一人の女性が一生に産む子供の平均数)の低下や第一子出生時年齢の上昇 が報告されている(5)。このように子を設けたいと望む年齢が上昇する一方で, 子を設けることができる期間は限られており,産みたい年齢と産める年齢の差 が狭まりつつある。これらの社会的背景から生殖機能の加齢性変化理解し,原 因を解明することは大変重要である。 また閉経は,生殖機能を失うばかりでなく内分泌的変化により様々な疾患が 発症するリスクを増加させる。例えば,卵巣から分泌されるエストロゲンは, 生殖機能に関与する他,血管保護や骨代謝において重要な働きを担っているた め,閉経後に血液中のエストロゲン濃度が低下すると動脈硬化や骨粗鬆症に罹 患するリスクが増加する(6)。 本研究では,加齢に伴い生殖機能がどのように低下していくのかを詳細に観 察し,早期にみられる生殖機能の加齢性変化について原因を解明することを目 的とした。
6 1-2 ラット ラットはヒトと同じく自然排卵動物であり,生殖機能に関するホルモン動態 もヒトと類似する点が多いことから生殖機能の研究にも汎用される。本研究で は,広く普及している系統である Sprague-Dawley(SD)系を実験に用いて,生 殖機能の加齢性変化を調べた。 1-3 発情周期に伴う生殖関連ホルモンの変動 自然排卵動物は,排卵間隔に周期性を示し,ヒトやサルでは約 28 日間隔で 排卵し,ラットでは,4 または 5 日間隔で排卵する。ステロイドホルモンの一 種であるエストロゲンは,卵胞を構成する顆粒層細胞から分泌され,卵胞が成 熟するに従ってその分泌量が増加する。また,卵胞は,排卵すると黄体を形成 しプロゲステロンを分泌するため,排卵後に血液中プロゲステロン濃度が上昇 する。このように,発情周期に伴って卵巣から分泌されるステロイドホルモン の血液中濃度にも周期的な変動が生じ,これはヒトとラットに共通してみられ る。 エストロゲンやプロゲステロンは膣粘膜上皮に作用し,組織学的な変化をも たらすため,膣内膜上皮の形態を反映する膣スメアを観察することで卵巣状態 を非侵襲的且つ継続的に把握することが出来る(7)。この膣スメアを基準に判断 すると,ラットの発情周期は,発情前期,発情期,発情後期及び発情休止期の 4 ステージに分類できる。発情前期の膣スメア像は,角化細胞と有核細胞が混 在することを特徴とし,発情期では多量の角化細胞が観察され,発情後期及び 休止期では角化細胞に加えて白血球が混じるようになる(8)。エストロゲンやプ ロジェステロンの分泌に異常が起こると,膣スメアに影響するため,膣スメア を継続的に観察することで卵巣機能の異常が捕らえられる。
7 ヒトもラットも共通して,卵胞の成熟や排卵は下垂体前葉から分泌される性 腺刺激ホルモンによってコントロールされており,下垂体前葉での性腺刺激ホ ルモン分泌は,その上位の視床下部からの性腺刺激ホルモン放出ホルモン (Gonadotropin-Releasing Hormone :GnRH)によって促進される。さらに,視 床下部と下垂体前葉の機能は卵巣から分泌されるエストロゲン,プロジェステ ロン及びインヒビン等によって調整されている。未成熟な卵胞は下垂体から分 泌される黄体形成ホルモン(Luteinizing hormone:LH)や卵胞刺激ホルモン
(Follicle Stimulating Hormone:FSH)によって発育を促進され,成熟卵胞とな
る。卵胞が発育するにつれ血液中のエストロゲン濃度が増加し,これが引き金 となって視床下部が活性化されると GnRH が一過性に大量に分泌される(GnRH サージ)。GnRH サージが下垂体のゴナドトロフに作用すると LH がサージ状に 分泌され(LH サージ),この LH サージが成熟した卵胞に作用して排卵が誘起 される(9)。血液中のエストロゲン濃度の上昇が LH サージの引き金となること はヒトとラットで共通している。雌ラットの場合,最も血液中のエストロゲン 濃度が増加するのは,発情前期の正午前後で,同日の夕方に LH サージが誘起 され数時間後に排卵が起こる。この血液中エストロゲン濃度の上昇が引き金と なって,視床下部-下垂体系に作用して GnRH と LH サージを誘起する現象をエ ストロゲンの正のフィードバック作用とよぶ。排卵を含む卵巣機能は,視床下 部及び下垂体の支配を受けており,同時に視床下部と下垂体はエストロゲンの 影響を受けている。このような,内分泌学的機構を視床下部-下垂体-性腺軸と よび,視床下部-下垂体-性腺軸いずれかの部位に異常をきたすと,発情周期や 排卵に異常が起こる。雌ラットにおいては,加齢に伴う生殖機能の低下の早期 マーカーとして LH サージの減弱が挙げられる (10-12)。また,ヒトにおいても 閉経前の閉経移行期では LH サージの減弱・消失がみられることが報告されて
8 いる(13)。
1-4 Kisspeptin
kisspeptin はG protein-coupled receptor(GPR) 54 のリガンドとして,2001 年 に武田薬品工業によって発見された GnRH 神経のアクティベーターである(14)。 kissppetin は,神経ペプチドであり Kiss1 遺伝子によってコードされている。近 年,ヒトや齧歯類を初めとする雌性動物で,kisspetin は視床下部において GnRH 分泌を調整するホルモンとしての役割が明らかにされた(15-18)。雌ラットの kisspeptin 陽 性 神 経 細 胞 ( kisspeptin 神 経 ) は , 視 床 下 部 の 前 腹 側 周 囲 核 (anteroventral veriventricular neucleus: AVPV)と弓状核 (arcuate nucleus:ARC) に局在しており(19, 20),AVPV の kisspeptin 神経は,GnRH をサージ状に分泌さ せることで LH サージを制御し,ARC の kisspeptin は,GnRH のパルスジェネレ ータとして働くことで LH パルスを制御している。kisspeptin 神経は,エストロ ゲン受容体アルファ(estrogen receptor alpha:ER)を発現しており(19, 21, 22), Kiss1 mRNA の発現は,エストロゲンによって調節されている。エストロゲンが, AVPV の kisspeptin 神経に作用すると Kiss1 mRNA の発現を上昇させ,反対に ARC の kisspeptin 神経に作用すると発現を抑制する。このため,雌ラットでは,血液 中エストロゲン濃度が上昇する発情前期の午後に AVPV の Kiss1 mRNA 発現量 が増加し(19),エストロゲンの正のフィードバックにより AVPV の kisspeptin 神 経が活性化されると kisspeptin が放出される。AVPV の kisspeptin 神経は,GnRH 神経核が集合する視索前野(preoptic area: POA)に投射しており,放出された kisspeptin は,その受容体である GPR54 を介して GnRH 神経を活性化し,GnRH のサージ状分泌を惹起する(23-26)。この,GnRH サージによって下垂体前葉の性 腺刺激ホルモン分泌細胞から LH がサージ状に分泌される(Fig. 1-1)。 kisspeptin 神経の加齢性変化に関する報告は少なく(27, 28),本研究では生殖機 能の加齢性マーカーの 1 つである LH サージの減弱・消失について kisspeptin 神 経の加齢による機能的変化に着目し研究を行った。
9 1-5 胚・胎児の分化・発育 LH サージにより卵胞から排卵された卵細胞は卵管膨大部で受精し,細胞分 裂を繰り返しながら子宮へと輸送される。後に胚葉へと分化する内部細胞塊を 形成した胚盤胞は子宮内膜に接着すると,胎盤の原基である外胎盤円錐が子宮 内膜へ侵食し,固着する(29)。この一連の現象を着床という。 着床後,胚を形成する細胞は更に増殖を繰り返し,外胚葉,内胚葉及び中胚 葉を形成して各器官へと分化する。器官形成期を経た後も,胎児は成長を続け ラットでは妊娠 22 日目に,ヒトではおよそ 290 日で分娩に至る。 ヒトもラットも母体の加齢より受胎率の低下(4),胚の分化異常及び胎児の発 育遅延(3, 30-32),異常分娩(33)など様々な影響をもたらすことが知られている。 1-6 本研究の目的 加齢により生殖機能がどのように低下するかを理解し,メカニズムを解明す ることは,社会的に極めて重要な課題である。本研究では, SD ラットを用い て, 雌ラットの生殖機能が加齢に伴いどのように変化するか調べた(第 2 章)。 次いで,早期にみられる加齢性変化に着目し,胚の分化異常及び着床不全につ いて検討した(第 3 章)。また,早期にみられる別の加齢性変化である異常発情 周期の発生メカニズムを解明するために,LH サージの消失に着目し,視床下 部 AVPV の kisspeptin 神経の関与について検討した(第 4 章)。最後に,自然排 卵が停止した高週齢のラットについて,交尾排卵の可能性を確かめた(第 5 章)。
10 Figure 1-1:視床下部-下垂体-卵巣軸 ① 卵胞の成長に伴い,エストロゲン濃度が上昇する。 ② エストロゲンの正のフィードバック作用によって AVPV の kisspeptin 神経が活性化さ れる。 ③ kisspeptin は GnRH 神経を刺激し,GnRH のサージ状分泌を誘起する。 ④ GnRH サージにより下垂体前葉の性腺刺激ホルモン分泌細胞から LH がサージ状に分 泌される。 ⑤ LH サージは卵巣の成熟卵胞に作用し,排卵を誘起する。 ⑥ ARC の kisspeptin 神経は GnRH 神経のパルスジェネレータとして働く。
11 第 2 章 雌ラットにおける加齢に伴う生殖機能の変化 1. 背景と目的 ヒトやラットをはじめとする雌性哺乳類では,加齢とともに生殖機能が低下 し,不規則な発情周期,排卵障害及び受胎率の低下などを示すことが知られて いる(11-13, 34-37)。しかし,これらの報告の多くは特定の現象について,若い 女性もしくは雌ラットと高齢の女性もしくは雌ラットを比較したものであり (32, 36, 37),生殖機能の加齢性変化がどの段階から,どのような順番で発現す るかについて言及した報告は乏しい。また,ラットでは系統や飼育条件の違い は生殖機能の加齢性変化に影響を与えるため,異なる施設で異なる系統を用い て得られた結果を比較すことは難しい。よって,生殖機能の加齢性変化がどの ように始まるかを調べるためには,同一系統のラットを用いて同一施設内で経 時的に精査する必要がある。本研究では,同一時期に生産された Sprague-Dawley (SD)ラット(同一ロット)を用いて,春期発動に相当する 6 週齢から半数例 以上で発情周期に異常がみられる 40 週齢の間,発情周期,受胎,胚・胎児の生 存・発育に着目し,加齢に伴う生殖機能の変化を精査した。また,ラットでは 母動物の加齢が胎児の形態にどのような影響をもたらすかについて調べた報告 が乏しいことから,本研究では胎児の外表,内臓及び骨格観察も合わせて実施 した。 2. 材料及び方法 2.1 使用動物 日本チャールスリバー株式会社で生産された Spraque-Dawley 種 Crl:CD(SD) の雌ラット(入荷時週齢 5 週齢,同一時期に生産)及び雄ラット(入荷時週齢 13 週齢,生産及び購入時期は複数にまたがる)を購入し,それぞれ群飼ケージ [1509 cm2 (底面積)×17.4 cm(高さ),3-5 匹/ケージ]及び単飼ケージ[647.7 cm2
12 (底面積)×17.4 cm(高さ)]で SPF 環境下で飼育した。動物室の照明時間は,12 時間/日(点灯時間:7 時~19 時)とし,飼料はγ線照射した固形飼料(CR-LPF, オリエンタル酵母株式会社,東京,日本)を与え,水道水を自由摂取下で飼育 した。雌ラットは入荷から 1 週間の馴化期間を経た後,各実験に使用する際に 単飼ケージに移動した。全ての動物は武田薬品工業株式会社が定める動物の取 り扱いに関するガイドラインに従って取り扱った。 2.2 膣スメアの採取及び発情周期判定 48 匹の雌ラットを用いて,6 週齢から 36 週間(43 週齢まで)毎朝 9-10 時の 間,膣スメアを採取して発情周期を判定した。採取した膣スメアは 5 分間アル コールで固定し,3%ギムザ染色液で染色した後に,顕微鏡下で観察した。発情 周期の判定は以下の通り実施した。 正常発情周期:4~5 日に 1 回発情期が観察される 異常発情周期:正常発情周期以外で,下記の 3 つに分類した。 延長型:発情期の間隔が 5 日以上要する 持続発情型:角化細胞有意なスメア像が 3 日間以上持続する 持続休止期型:休止期が 6 日以上持続する 評価は 2 週間を 1 期間とし,6-7, 8-9, 10-11, 12-13, 14-15, 16—17, 18-19, 20-21, 22-23, 24-25, 26-27, 28-29, 30-31, 32-33, 34-35, 36-37, 38-39, 40-41 及び 42-43 週齢 毎に発情周期を判定した。 2.3 交尾率,受胎率及び胚・胎児の発達 6, 8, 10, 15, 19, 23, 27, 31, 35 及び 40 週齢の未経産ラット(6-27 週齢 N=10/
13 各週齢,31-40 週齢 N=20/各週齢)を用いて,1 週間発情周期を観察した後に, 最長 1 週間に渡って雄(交配時週齢:14-30 週齢)と 1 対 1 で同居させた。交 尾確認は膣栓の有無で行い,膣栓がみられた日を妊娠 0 日とし,週齢ごとに交 尾率を算出した。膣栓が確認された雌は雄から隔離し,単飼ケージで妊娠 20 日目まで飼育した。妊娠 20 日に CO2麻酔下で腹大動脈を切開して安楽死させ, 放血後,肉眼で着床数,死亡胚・胎児数及び生存胎児数を数え,実体顕微鏡下 で黄体数を数えた。胎児の生死は外部からの刺激に対し反応するか否かで判断 した。これらの数値をもとに,週齢ごとに受胎率,平均着床前及び後死亡率を 算出した。各パラメータは以下の式に従って算出した。 交尾率=膣栓が確認された雌数/交配雌数 受胎率=着床が確認された雌数/膣栓が確認された雌数 一腹当たりの着床前死亡率=(黄体数-着床数)/黄体数 一腹当たりの着床後死亡率=(着床数-生存児数)/着床数 着床前・後死亡率は週齢毎に平均値±SD を算出し、週齢の値とした。 死亡胚・胎児は肉眼では胎盤が認められないが子宮壁に着床した形跡が認め られる吸収胚,肉眼で胎盤の形成が認められる胎盤遺残及び四肢の形成が認め られる状態まで発育し死亡した死亡胎児の 3 つに分類した。 生存児は全例について実体顕微鏡下で外表観察を実施し,体重を測定した。 さらに,半数の生存児について内臓観察を,残りの半数について骨格観察を実 施した。内臓観察に用いた生存児に,心臓を拡張したまま固定する目的で 10% マグネシウム溶液を胸腔内に注入し,その後 10%ホルマリンで固定した。作製 した内臓標本は実体顕微鏡下で観察し,Barrow's 及び Wilson 氏の評価方法に
14 基づき内臓異常及び変異を評価した (38, 39)。骨格観察に用いた生存児は,99% エタノールで固定した後に染色液(2 w/v%アリザリンレッド 10 mL+KOH 12.5 g/純水 1L)に 2-3 日間浸漬させた。その後 20,50%グリセリン溶液に浸漬させ 観察に用いた(改 Dawson 氏の方法(40))。作製した骨格標本は実体顕微鏡下で 観察し,骨格異常及び変異を評価し,さらに胎児の発育の指標の一つである仙 尾椎数を数えた。 胎児体重及び仙尾椎数は一腹当たりの平均値を各母動物の値として,週齢ご とに平均値を算出し評価した。 2.4 統計解析及びデータの評価 全データについて,10 週齢を対照群とし統計解析を実施した。発情周期は Wilcoxon 検定を,交尾率及び受胎率は 2x2Fisher の直接確立検定を実施した。 その他のデータについて,Bartlett 検定を実施して分散の均一性を確認し,分散 が均一で無かった黄体数,着床数,着床前死亡率,着床後死亡率,生存児数及 び仙尾椎数はノンパラメトリック Steel 検定を,胎児体重は分散が均一であった ため,パラメトリック Dunnett 検定を実施した。全ての統計解析は SAS function PROBMC (SAS/STAT ソフトウェア: Changes and Enhancements, through Release 6.11. Cary, NC: SAS Institute Inc.)を用いて実施した。
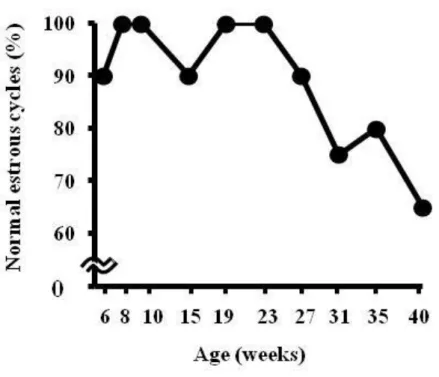
また,雌ラットの週齢と各パラメータの間に相関が認められるか確かめる為 に,相関係数(R2)を算出した。 3. 結果 3.1 発情周期 正常発情周期を示す動物の割合と週齢の間に相関がみられ(R2=0.8133), 雌動物が加齢するに従って正常発情周期を示す動物の割合は減少した。9-10 週
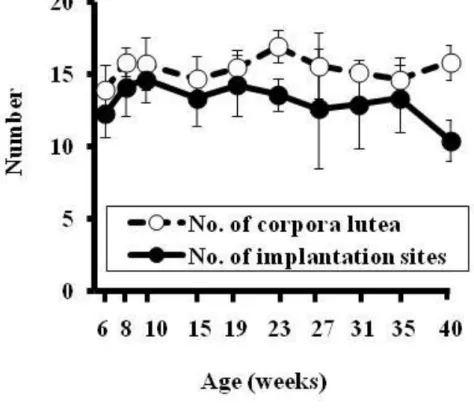
15 齢の間で 100%の動物が正常発情周期を示したのに対して,24 週齢以上の動物 では正常発情周期を示す動物の割合が有意に低下し,24-25 週齢の間で正常発 情周期を示した動物の割合は 81%であり,42-43 週齢の間では 19%であった。 26-27 週齢の間で最も多く認められた異常発情周期の型は延長型であり,次い で持続発情型及び休止期型がみられた。その後,週齢が高くなるにつれ,持続 発情型を示す動物が増え,次いで持続休止期型を示す動物が増加した。41-42 週齢の間では 27%が正常発情周期,6%が延長型異常発情周期,6%が持続発情 型異常発情周期及び 60%が持続休止期型異常発情周期を示した(Fig. 2-1)。 3.2 交尾率,受胎率,黄体数及び着床数 10 週齢のラットでは,交尾前に 100%の動物が正常発情周期を示し,どの週 齢と比較しても有意差は認められなかった(Fig. 2-2)。10 週齢のラットでは交 尾率及び受胎率は 100%であり,どちらも雌ラットの週齢に相関して(交尾率: R2=0.7398,受胎率:R2=0.618)減少した。しかし,どの週齢と比較しても両 パラメータにおいて有意な差は認められなかった(Fig. 2-3)。また,10 週齢の 着 床 数 は 14.6 ± 1.9 で あ り , 雌 ラ ッ ト が 加 齢 す る に 従 っ て 減 少 し た が (R2 =0.3357),どの週齢と比較しても両パラメータにおいて統計学的な有意差 はみられなかった。10 週齢の黄体数は 15.7±1.6 であり,週齢との間に相関は 認められなかった(Fig. 2-4)。 3.3 胚・胎児の死亡率 着床前・後死亡率と母動物の週齢の間に相関がみられ(着床前死亡率: R2=0.4173,着床後死亡率:R2=0.6057),母動物が加齢するに従って両パラメ ータ共に増加した。しかし,10 週齢の着床前・後死亡率(それぞれ 7.0±7.7 及 び 11.1±9.2%)と比較してもどの週齢も統計学的な有意差は認められなかった
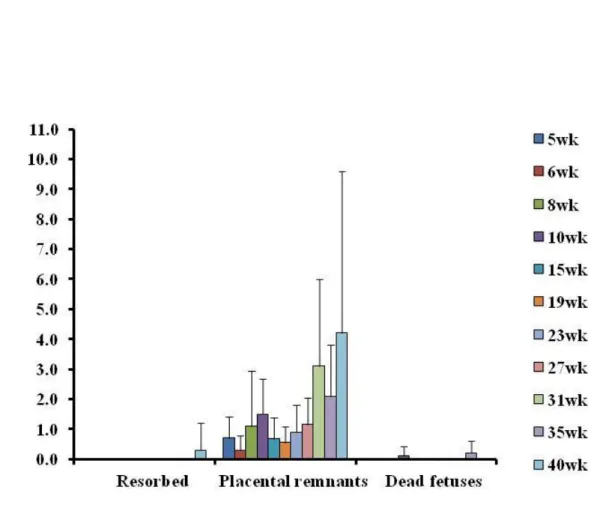
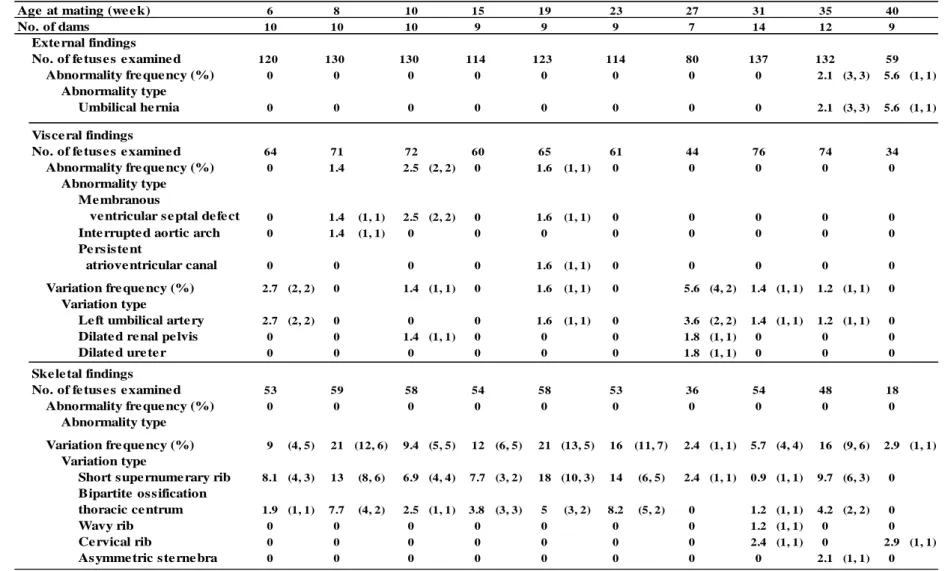
16 (Fig. 2-5)。 死亡胚・胎児の状態を吸収胚,胎盤遺残及び死亡胎児に分類したところ,ど の週齢の母動物においても胎盤遺残が主であり,その数は母動物の週齢に比例 し増加した(R2 =0.6024)。10 週齢でみられた胎盤遺残の割合は 1.5±1.2%であ り,どの週齢と比較しても統計学的な有意差は認められなかった。死亡胎児は 10 及び 35 週齢の母動物で少数みられたが母動物の週齢との間に相関はみられ なかった。また,吸収胚は 40 週齢の動物のみで確認された(Fig. 2-6)。 3.4 胎児の生存,発育及び形態学的異常 平均生存児数は母動物の週齢に相関して減少し(R2 =0.4723),10 週齢の平 均生存児数は 13.0±2.3 であり,40 週齢では 5.9±5.2 と有意に低値であった(Fig. 2-7)。また,胎児体重及び仙尾椎数は母動物の加齢に従いどちらも低下した(胎 児体重(雌/雄):R2 =0.582/0.8095,仙尾椎数:R2=0.697)。10 週齢の母動物から 得られた胎児の平均重量は雄胎児及び雌胎児でそれぞれ 3.67±0.29 及び 3.48± 0.26 g であった。これに対して 35 週齢の母動物から得られた雌胎児の重量は 3.12±0.27 g であり,40 週齢の母動物から得られた雄及び雌胎児の重量はそれ ぞれ 2.99±0.61 及び 3.04±0.57 と 10 週齢の胎児と比べてそれぞれ有意に低値で あった(Fig. 2-8 A)。40 週齢の母動物から得られた胎児の仙尾椎数は 6.7±0.8 であり,10 週齢の母動物から得られた胎児(7.7±10.3)と比較すると統計学的 に有意な低値であった(Fig. 2-8 B)。 外表観察では,35 及び 40 週齢の母動物から得られた胎児で,齊ヘルニアが それぞれ 3/132 例(同週齢内の平均として 2.1%)及び 1/59 例(5.6%)にみら れた。内臓観察では膜性心室中隔欠損が 8,10 及び 19 週齢の母動物から得られ た胎児でみられ,発現頻度はそれぞれ 1.4,2.5 及び 1.6%であり,発現頻度と母
17 動物の週齢に相関がみられなかった。また,骨格観察では 31 及び 40 週齢の母 動物から得られた胎児で頚肋骨がそれぞれ 2.4 及び 2.9%の頻度でみられた (Table 2-1)。 4. 考察 本研究では,雌ラットにおける生殖機能の加齢性変化がいつからどのような 順で認められるかを明らかにする目的で,発情周期を 6-43 週齢に渡り経時的に 観察し,交尾の許容,受胎及び胚・胎児の生存性・発達を 6 週齢から 40 週齢の 雌ラットを用いて検査した。得られた生殖パラメータについて成績が良好であ った 10 週齢を対照群とし,その他の週齢と統計解析を実施した。その結果,24 週齢以上で正常発情周期を示す動物の割合が低下し,35 週齢以上の母動物から 得られた胎児では胎児体重の低下や仙尾椎数の減少が認められ,40 週齢の母動 物では生存児数が減少した。その他,黄体数を除く全てのパラメータ(交尾率, 受胎率,着床数,着床前・後死亡率,胎盤遺残数)に母動物の週齢と相関がみ られた。 雌ラットは 4-5 日の発情周期を示す自然排卵動物であり,膣スメアは卵巣の 内分泌状態(特にエストロゲンとプロゲステロンの分泌)を反映していること から膣スメアを経時的に観察することで排卵周期を知ることができる(7)。 24-25 週齢で最も多くみられた異常発情周期は延長型であったことから,排卵 間隔の延長を示すラットの割合が 24 週齢から増加したことが明らかとなった。 加齢に伴い異常発情周期を示したラットでは,初期に延長型もしくは持続発情 型を示し,その後持続休止期型を示す傾向がみられた。持続発情型及び持続休 止期型異常発情周期を示すラットの卵巣は萎縮し黄体数が減少していること から(41),このような動物では排卵障害が起こり,自然排卵が停止していると
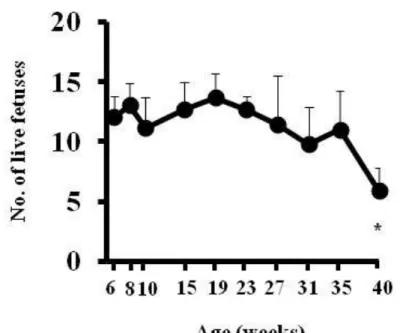
18 推察される。本研究では,42-43 週齢の間で正常性周期を示した動物の割合は 27%であり(Fig. 2-1),この結果は,Watanabe らの結果と概ね一致している(42)。 既報告によると,Long-Evans ラットでは 8 ヶ月齢以上で正常性周期を示した 動物の割合が 50%であり,Jcl:Wistar ラットでは 40 週齢以上で 80%の動物が正 常発情周期を示したとある(36, 37, 42)。また,未経産のラットは経産のラット に比べて早期に異常発情周期を示すことが報告されている(36, 37)。これらの 報告を踏まえると,加齢性にみられる排卵障害の発生時期はラットの系統及び 分娩経験の有無によって異なることが考えられる。SD ラットは他系統と比較 し早期に加齢性の排卵障害を示すことが推測された。 以上の結果から,本研究で使用した Crl:CD(SD)ラットでは 24 週齢から卵巣 機能が低下し始めたものと推察した。 本研究では,10 週齢の動物の交尾率が 100%だったのに対して,35 週齢の動 物では 80%であり,40 週齢の動物では 70%であったことから,交尾率は雌ラ ットの加齢に伴い減少することが示された。また,膣栓が確認出来た高週齢の 動物では,若い動物に比べてケージの底に落ちている膣栓の数が少ない傾向が あり(データ示さず),交尾回数が減少した可能性が示唆された。 本研究では,受胎率及び着床数に 10 週齢の動物と比較していずれの週齢で も統計学的な有意差は認められなかったが,両パラメータとも母動物の週齢と の間に負の相関が認められた。また,生存児数についても母動物の週齢との間 に負の相関が認められ,40 週齢では 10 週齢に比べて有意な低値を示した。着 床前・後死亡率も同様に母動物の週齢との間に負の相関が認められたが,黄体 数と母動物の間に相関が認められなかったことから受胎率,着床数及び生存児 数の減少は着床前・後死亡率の増加によるもと推察された。Matt らは,高週齢 の母動物で一腹児数が減少するのは,正常に発達する初期胚の数が減少するた
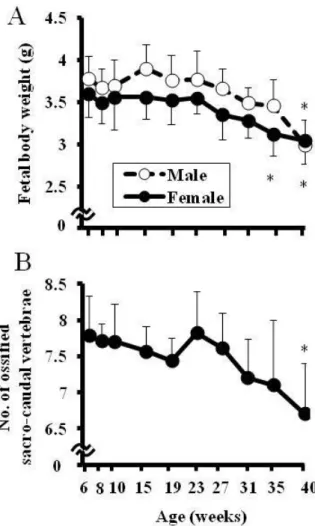
19 めであると報告している(37)。また別の既報告で,9-10 ヶ月齢の動物の受精卵 では,3-4 ヶ月齢に比べて卵割の遅延及び割球の断片化または変性が高頻度で みられることが報告されている(43)。さらに,本研究で死亡した胚・胎児を吸 収胚,胎盤遺残及び死亡児に分類したところ,胎盤遺残数と母動物の週齢に相 関が認められ,着床後の胚は主に妊娠中期で死亡した可能性が考えられた。こ のことから本研究で高週齢の母動物にみられた着床前・後死亡率の上昇は,母 動物が加齢することにより妊娠初期から中期胚の発生・分化に異常があり生存 性が低下した可能性が考えられた。一方,排卵したが受精せず,受胎率が低下 している可能性も否定できないことから,高週齢のラットでみられた受胎率低 下の原因を解明する目的で妊娠初期から中期の胚の発達に着目したさらなる研 究をおこない,その結果を本研究の第 3 章に記述した。また,胚の発生は正常 であるが母動物の妊娠維持機能が加齢により低下している可能性も考えられ, これについては今後の課題と考える。 胎児重量について 10 週齢を対照群とし比較したところ,35 週齢の母動物か ら得られた雌胎児及び 40 週齢の雌雄胎児の重量が有意に低値であった。胎児の 成長の指標として用いられる仙尾椎数は(44), 40 週齢の母動物から得られた胎 児で 6.7 と 10 週齢の母動物と比較して有意に低値であった。したがって,胎児 の発育遅延は胎児体重の減少がみられた 35 週齢以上の母動物で起こるものと 推察した。 外表観察では,35 及び 40 週齢の母動物から得られた胎児で,齊ヘルニアが それぞれ 3/132 例(同週齢内の平均として 2.1%)及び 1/59 例(5.6%)にみら れた。過去 10 年間に同施設で 630 匹の Crl:CD(SD)ラットを用いて胎児の外表 観察を実施したが,齊ヘルニアがみられた胎児は 1 例もみられておらず,本研 究でみられた齊ヘルニアは母動物が加齢したことによるものと考えられた。
20 以上の結果から,雌 Crl:CD(SD)ラットでは加齢により発情周期,交尾率, 受胎率,生存児数及び胚・胎児の発生及び成長に影響がみられ,24 週齢から排 卵障害を示す動物の割合が増加し,生殖機能が減退することが明らかとなった。 本研究では,本章でみられた母動物の加齢に伴う胚の生存性の低下及び加齢 に伴う異常発情周期について原因を解明するため,それぞれ第 3 章及び第 4 章 でさらに検討した。
21 Figure 2-1:雌ラットの加齢による発情周期の変化 6-43 週齢の間膣スメアを採取し,発情周期を観察した(N=48)。 Normal (正常発情周期):4~5 日に 1 回発情期が観察された場合,Prolonged estrous cycles (延長型異常発情周期):発情期の間隔が 5 日以上延長した場合, Persistent estrus (持続発情型異常発情周期):角化細胞有意なスメア像が 3 日間 以上持続した場合,Persistent diestrus (持続休止期型異常発情周期):休止期が 6 日以上持続した場合,*:P<0.05,vs 10-11 週齢
22
Figure 2-2:交配に使用した雌ラットの交尾前の発情周期の加齢性変化 6 から 40 週齢のラットについて,交尾前の 1 週間発情周期を観察した(6 から 27 週齢 N=10/各週齢,31 から 40 週齢 N=20/各週齢)。数値は正常 発情周期を示した雌ラットの割合を示す。
23 Figure 2-3:雌ラットの加齢による交尾率及び受胎率の変化 交配翌日の朝に膣栓が確認された雌ラットについて交尾が完了したと判断し,週例毎に 交尾率(copulation index: 膣栓が確認された雌数/交配雌数×100)を算出した(6-27 週 齢 N=10/各週齢,31-40 週齢 N=20/各週齢)。また,交尾が完了した雌ラットを妊娠 20 日で剖検し,妊娠したラットの割合を算出し受胎率(fertility index: 着床が確認され た雌数/膣栓が確認された雌数×100)とした(6,8,10,15,19,23,27,31,35 及び 40 週齢についてそれぞれ N=10,10,10,10,9,10,9,9,18,16 及び 14)。数値は 交尾または受胎が確認された雌ラットの割合を示す。
24
Figure 2-4:母動物の加齢による黄体数及び着床数の変化
妊娠 20 日目に剖検し,妊娠が確認されたラットについて母動物毎に黄体 数(No. of Corporalutea)と着床数(No. of implantation sites)を数え,週齢 毎に平均値及び SD を算出した(6,8,10,15,19,23,27,31,35 及び 40 週齢についてそれぞれ N=10,10,10,10,9,9,9,7,14,12 及び 10)。数値は平均値±SD を示す。
25
Figure 2-5:母動物の加齢による着床前死亡率及び着床後死亡率の変化 妊娠 20 日目に剖検し,妊娠が確認されたラットについてそれぞれ着床前死 亡率[A,Pre-implantation loss rate, 着床前死亡率=(黄体数‐着床数)/黄体 数×100]及び着床後死亡率[B,Post-implantation loss rate,着床後死亡率=(着 床数-生存胎児数)/着床数×100]を個体ごとに算出し,週齢毎に平均値及 び SD を算出した。6,8,10,15,19,23,27,31,35 及び 40 週齢につい てそれぞれ N=10,10,10,10,9,9,9,7,14,12 及び 10,数値は一腹当 たりの平均値 ± SD を示す。
26
Figure 2-6:母動物の週齢別死亡胚・胎児の分類
妊娠 20 日の剖検時における胚・胎児の死亡を分化・発育の状態から,吸収 胚(Resorption),胎盤遺残(Placental remnant)及び死亡児(Dead fetuses) に分類し,一腹あたりにみられた各死亡胚・胎児の割合を算出した後,週例 毎に平均値及び SD を算出した。6,8,10,15,19,23,27,31,35 及び 40 週齢についてそれぞれ N=10,10,10,10,9,9,9,7,14,12 及び 10, 数値は一腹当たりの平均値±SD を示す。
27
Figure 2-7:母動物の加齢による生存児数の変化
妊娠 20 日に剖検し生存児数(No. of live fetuses)を数え,週例毎に一腹あた りの平均生存児数を算出した。使用した母動物数:6,8,10,15,19,23, 27,31,35 及び 40 週齢についてそれぞれ N=10,10,10,10,9,9,9,7, 14,12 及び 10,数値は一腹当たりの平均値±SD を示す。*:P<0.05 vs 10 週齢
28
Figure 2-8:母動物の加齢による胎児体重及び胎児の仙尾椎数の変化
妊娠 20 日目の剖検で得られた生存児について胎児体重を測定し,週例毎に一 腹あたりの平均胎児体重(Fetal body weight)を算出した(A)。また,半数の 生存児について骨格観察を実施し,仙尾椎数を数え,一腹あたりの平均仙尾 椎数(No. of ossified sacro-caudal vertebrae)を算出した(B)。使用した母動物 数:6,8,10,15,19,23,27,31,35 及び 40 週齢についてそれぞれ N=10, 10,10,10,9,9,9,7,14,12 及び 10,数値は一腹当たりの平均値±SD を示す。*:P<0.05 vs 10 週齢
29 Table 2-1 胎児の外表,内臓及び骨格観察
括弧内は胎児数,母動物数を示す。
Age at mating (we e k) 6 8 10 15 19 23 27 31 35 40
No. of dams 10 10 10 9 9 9 7 14 12 9
Exte rnal findings
No. of fe tuse s e xamine d 120 130 130 114 123 114 80 137 132 59
Abnormality fre que ncy (%) 0 0 0 0 0 0 0 0 2.1 (3, 3) 5.6 (1, 1)
Abnormality type
Umbilical he rnia 0 0 0 0 0 0 0 0 2.1 (3, 3) 5.6 (1, 1)
Visce ral findings
No. of fe tuse s e xamine d 64 71 72 60 65 61 44 76 74 34
Abnormality fre que ncy (%) 0 1.4 2.5 (2, 2) 0 1.6 (1, 1) 0 0 0 0 0
Abnormality type Me mbranous
ve ntricular se ptal de fe ct 0 1.4 (1, 1) 2.5 (2, 2) 0 1.6 (1, 1) 0 0 0 0 0
Inte rrupte d aortic arch 0 1.4 (1, 1) 0 0 0 0 0 0 0 0
Pe rsiste nt
atriove ntricular canal 0 0 0 0 1.6 (1, 1) 0 0 0 0 0
Variation fre que ncy (%) 2.7 (2, 2) 0 1.4 (1, 1) 0 1.6 (1, 1) 0 5.6 (4, 2) 1.4 (1, 1) 1.2 (1, 1) 0
Variation type
Le ft umbilical arte ry 2.7 (2, 2) 0 0 0 1.6 (1, 1) 0 3.6 (2, 2) 1.4 (1, 1) 1.2 (1, 1) 0
Dilate d re nal pe lvis 0 0 1.4 (1, 1) 0 0 0 1.8 (1, 1) 0 0 0
Dilate d ure te r 0 0 0 0 0 0 1.8 (1, 1) 0 0 0
Ske le tal findings
No. of fe tuse s e xamine d 53 59 58 54 58 53 36 54 48 18
Abnormality fre que ncy (%) 0 0 0 0 0 0 0 0 0 0
Abnormality type
Variation fre que ncy (%) 9 (4, 5) 21 (12, 6) 9.4 (5, 5) 12 (6, 5) 21 (13, 5) 16 (11, 7) 2.4 (1, 1) 5.7 (4, 4) 16 (9, 6) 2.9 (1, 1)
Variation type
Short supe rnume rary rib 8.1 (4, 3) 13 (8, 6) 6.9 (4, 4) 7.7 (3, 2) 18 (10, 3) 14 (6, 5) 2.4 (1, 1) 0.9 (1, 1) 9.7 (6, 3) 0
Bipartite ossification
thoracic ce ntrum 1.9 (1, 1) 7.7 (4, 2) 2.5 (1, 1) 3.8 (3, 3) 5 (3, 2) 8.2 (5, 2) 0 1.2 (1, 1) 4.2 (2, 2) 0
Wavy rib 0 0 0 0 0 0 0 1.2 (1, 1) 0 0
Ce rvical rib 0 0 0 0 0 0 0 2.4 (1, 1) 0 2.9 (1, 1)
Asymme tric ste rne bra 0 0 0 0 0 0 0 0 2.1 (1, 1) 0
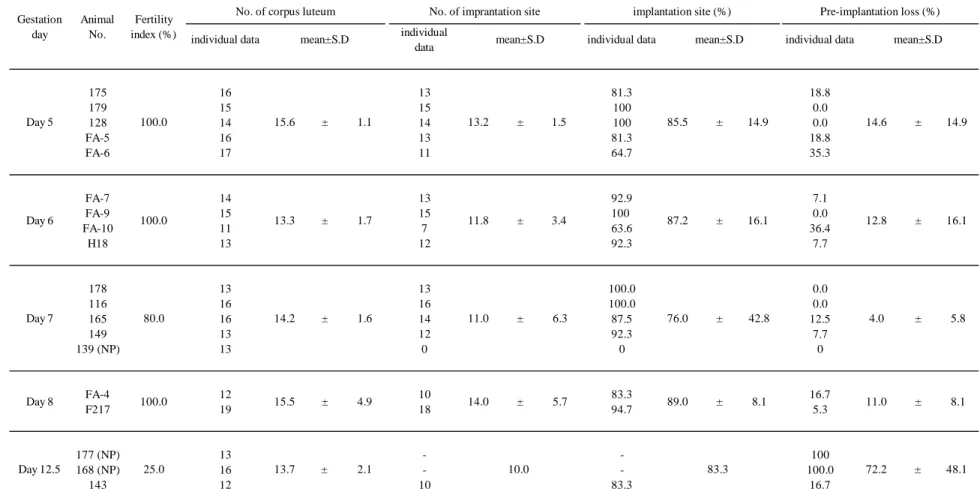
30 第 3 章 高週齢ラットの妊娠初期及び中期における胚・胎児の生存及び発育 1. 背景と目的 第 2 章で,高週齢の母動物で生存児数の減少と受胎率の低下がみられ,これ らは妊娠初期から中期の胚の生存性の低下に起因している可能性が考えられた。 この加齢性変化を詳しく調べるため,高週齢ラットの妊娠初期及び中期の胚の 生存性及び形態について検査した。 2. 材料及び方法 2.1 使用動物 日本チャールスリバー株式会社で生産された Spuraque-Dawley 系の雌ラット (Crl:CD(SD),5-8 週齢)及び雄ラット(13 週齢)を購入し,それぞれ群飼ケ ージ[1509 cm2 (底面積)×17.4 cm(高さ),3-5 匹/ケージ]及び単飼ケージ[647.7 cm2(底面積)×17.4 cm(高さ)]で SPF 環境下で飼育した。動物室の照明時間は, 12 時間/日(点灯時間: 7 時~19 時)とし,飼料はγ線照射した固形飼料(CR-LPF, オリエンタル酵母株式会社,東京,日本)を与え,水道水を自由摂取下で飼育 した。雌ラットは入荷から 1 週間の馴化期間を経た後,各実験に使用する際に 単飼ケージに移動した。全ての動物は武田薬品工業株式会社が定める動物の取 り扱いに関するガイドラインに従って取り扱った。 2.2 交配 または週齢の雌ラットを発情前期の夕方に雄と交配し,翌朝膣栓を 確認して交尾の有無を判定し,膣栓が確認された日を妊娠日とした。 2.3 黄体数,着床数,着床率及び着床前死亡率の算出並びに着床部位の観察 妊娠,,,または日目の時に,子宮外観から着床点を可視化し着床
31 を判定するため 3%エバンスブルーmL/匹(0.9%生理食塩水で調整)を静脈内 投与した。投与時間後に高濃度の CO2 を用いて安楽死し,卵巣及び子宮を摘 出して黄体数及び着床数を数え,受胎率,着床率お呼び着床前死亡率を下記の 式をもとに算出した。 受胎率=着床がみられた雌数/膣栓がみられた雌数×100 着床率=着床数/黄体数×100 着床前死亡率=(黄体数-着床数)/黄体数×100 また,妊娠 6,7 及び 8 日の子宮について着床部位の外観を肉眼で観察した 後に,10w/v%ホルマリンを用いて固定した。ホルマリン固定から少なくとも 1 週間以降に,着床部位を切り出しパラフィンで包埋した。回転式ミクロトーム を用いて 10 m の薄切組織切片を作製し,HE 染色した。作製した組織標本は 光学顕微鏡下で観察した。 群構成 妊娠日齢 5 日目 6 日目 7 日目 8 日目 12 日目 14 週齢(匹数a) 3 4 5 4 3 37~41 週齢(匹数a) 5 4 5 2 4 a:膣栓が確認された動物数 2.4 実体顕微鏡下での観察 黄体数及び着床数の算出並びに着床部位の観察に用いたラットとは別のラッ トを用いて,妊娠 11 日目の胚を実体顕微鏡下で観察した(14 及び 37-41 週齢, それぞれ N=2)。妊娠 11 日に高濃度の CO2を用いて安楽死させ,速やかに子宮
32 を摘出して子宮角に沿って切開した。着床部位を確認した後,ピンセットを用 いてライヘルト膜を破り,胚を摘出した。得られた胚は直ちに 99%エタノール で固定した後,5%グリセリンに置換し実体顕微鏡下で観察した。 2.5 統計 各パラメータについて,妊娠日毎に t 検定を実施した。全てのデータは平均 値±標準誤差で表示し,P 値が 0.05 未満の場合に統計学的に有意な差であると 判定した。 3. 結果 3.1 受胎率,黄体数,着床数,着床率及び着床前死亡率(Table 3-1,及び 2, Fig. 3-1 及び 2) 受胎率は,14 週齢のラットではいずれの妊娠日においても 100%であり,37-41 週齢のラットでは妊娠 5,6 及び 8 日で 100%,妊娠 7 日で 80%であったが,妊 娠 12 日で 33.3%と低値を示した。しかし,着床が確認できなかった 37-41 週齢 のラット(動物番号 139,177 及び 168)でも黄体が観察され,14 週齢のラット と 37-41 週齢のラットで黄体数に差はみられなかった。そのため,37-41 週齢の ラットの着床前死亡率は妊娠 5 から 8 日では 10%前後であったのに対して,妊 娠 12 日で 72.2%と高値を示した。 受胎が確認された動物について着床数及び着床率を算出した結果,いずれの 妊娠日についても 14 週齢のラットと 37-41 週齢のラットの間で差は認められな かった。 3.2 着床部位の観察 着床部位を肉眼で観察した結果,着床部位の大きさが妊娠 5 日目では 14 週齢
33 のラットと 37-41 週齢のラットで違いは認められなかったが,妊娠 6~8 日目で 37-41 週齢のラットの着床部位は 14 週齢のラットに比べて明らかに小さかった。 Figure 3-4 にある代表例について,着床部位間の子宮の太さに対する着床部位の 直径(子宮角に対して垂直方向)の比を,写真をもとに算出した結果,14 週齢 のラットでは妊娠 5,6,7,8 及び 12 日目で 1.0±0.18(測定した着床部位数 13), 1.6±0.18(14),2.1±0.17(13),2.5±0.19(13)及び 3.3±0.8(15)であった のに対して,37-41 週齢のラットでは妊娠 5,6,7,8 及び 12 日目で 1.1±0.1 (13),1.2±0.2(12),1.5±0.3(14),2.0±0.4(10)及び 2.6±0.2(10)と妊 娠 5 日目を除くいずれの妊娠日においても 14 週齢のラットと比べて有意に低値 であった(妊娠日毎に T 検定を実施)。また,着床部位の間隔は 14 週齢のラッ トで概ね均等であったが,37-41 週齢のラットの着床部位の間隔はいずれの妊 娠日でも不均一であった。(Fig. 3-4)。 3.3 胚の形態学的検査 3.3.1 組織学的検査(妊娠 6,7 及び 8 日) 14 週齢のラットから得られた正常胚は,妊娠 6 日目で内部細胞塊(inner cell mass),卵黄嚢腔(yolk-sac cavity)及び外胎盤円錐(ectoplacental cone)の形成 がみられた。また,子宮内膜では細胞が肥大し脱落膜細胞化が認められた(Fig. 3-5)。14 週齢のラットでは子宮内膜上皮細胞の消失が認められ,着床部位周辺 の子宮内に毛細血管の浸潤が多くみられた(Fig. 3-6)。14 週齢のラットから得 られた胚では妊娠 8 日目で外胚葉,内胚葉及び中胚葉が確認出来た(Fig. 3-7)。 これに対して,37-41 週齢のラットから得られた胚では,妊娠 6 日目で内部細 胞塊や外胎盤円錐の区別がつかない構造異常を示す胚(妊娠 6 日,Fig. 3-5B) や発達遅延(妊娠 6 日,Fig. 3-5C)を示す胚がみられ,子宮内膜上皮細胞の単
34 層構造に変化はみられず,脱落膜の形成もみられなかった(Fig. 3-5)。妊娠 7 日目で,一部の子宮内膜の円柱上皮細胞は消失していたものの,14 週齢のラッ トと比べて毛細血管の浸潤が少なく外胎盤円錐内に赤血球の浸潤がみられない 胚が観察された。さらに,胚の大きさも 14 週齢のラットに比べて小さく,外胎 盤円錐も未発達であった(Fig. 3-6)。妊娠 8 日目では,死亡胚が観察された(Fig. 3-7)。 3.3.2 実体顕微鏡下での観察(妊娠 11 日) 14 週齢のラットでは頭褶(head fold),体節(somite)及び心臓原基(heart rudiment)の形成がみられたのに対して(Fig. 3-8 A 及び B),37-41 週齢のラッ トから得られた異常胚では頭褶及び心臓原基が確認できず体節の低形成がみら れた。さらに,37-41 週齢のラットから得られた胚では外胎盤円錐の発生位置 に異常がみられた(Fig. 3-8 C 及び D)。また,別の異常胚では胚の構造が確認 できないものもあった(Fig. 3-8 E)。 4. 考察 ラットでは,排卵後の卵子は卵管膨大部に輸送され妊娠 0 日午前中に卵管膨 大部で受精する。受精卵は卵割を繰り返しながら卵管内を移動し妊娠 4 日目に 子宮に到達する。妊娠 5 日目に,卵巣から分泌されるエストロゲン及びプロゲ ステロンによって胚受容能が整った子宮内膜上皮と接着し,着床が開始する (45)。従って,本研究で観察を開始した妊娠 5 日目では正常な動物において胚 の着床が開始されている胎日齢である。 妊娠初期では着床部位のふくらみが小さく,外観から着床部位を判断するこ とは困難であるが,エバンスブルー溶液を静脈内投与すると着床部位の血管新
35 生が盛んな部位にエバンスブルーが蓄積することで子宮の外観から着床部位を 特定することが出来る。本研究でもこの手法を用いて妊娠初期から中期の着床 部位を特定した。その結果,37-41 週齢のラットの受胎率は,妊娠 5 から 8 日 目に剖検した動物について 80 から 100%と高い値であったが,妊娠 12 日に剖 検した動物については 33%(1/3 例)と低値であった。着床がみられなかった 妊娠 7 日の 1 例(F139)及び妊娠 12 日の 2 例(F168 及び 177)においても黄 体数は減少しておらず,着床前死亡率は妊娠 5 から 8 日で 10%前後であったの に対し,妊娠 12 日では 72%と高値を示した。 着床部位を組織学的に検査した結果,37-41 週齢のラットから得られた胚で は発達遅延及び外胎盤円錐の低形成を特徴とする形態学的な異常がみられた。 正常な胚では,着床開始からすぐに外胎盤円錐の形成が始まり,外胎盤円錐は 子宮内膜へと浸潤し,基底部にある二倍体栄養外胚葉細胞が増殖して,胚体外 外胚葉になり将来胎盤へと分化することから胎盤系の組織幹細胞と考えられて いる(45)。また,本研究では 37-41 週齢ラットで妊娠 8 日目に子宮内膜側に吸収 される中途思われる死亡胚が確認されたことから,これらの異常胚では外胎盤 円錐の低形成により,正常に胚が子宮内膜へ浸潤することができず,胎盤を形 成する前に死亡し,子宮内膜に吸収されたと考えられた。そのため妊娠中期以 降まで着床痕が残らず,妊娠 12 日の剖検では見かけ上受胎率が低下し着床前 死亡率が上昇したと考えられた。 また,37-41 週齢ラットでは着床部位のふくらみが小さく,妊娠 7 日目の子 宮内膜を組織学的に観察すると,毛細血管の浸潤が乏しく脱落膜の形成がみら れなかった。脱落膜とは妊娠の成立に伴い変化し,分娩時に胎盤とともに剥脱 する子宮内膜組織の一部であり,子宮内膜間質細胞が胚の着床によって形態学 的および機能的に分化をする過程を脱落膜化という(46, 47)。この分化過程は絨
36 毛細胞の浸潤,胎盤形成に重要な役割を果たしていると考えられている(48)。 子宮内膜の毛細血管の拡張・新生,間質の浮腫や脱落膜化等は胚が子宮内膜に 浸潤し発育・分化することで進行することから,37-41 週齢ラットでは外胎盤 円錐の異常を伴った胚が子宮内膜へ正常に浸潤できず,脱落膜化が正しく引き 起こされなかったと推察した。 正常な動物では,子宮に到達した胚の着床間隔は均一に保たれるのに対して, 本研究では,37-41 週齢ラットでいずれの妊娠日でも着床間隔が不均等であっ た。着床間隔を決定するメカニズムは未だ明らかとなっていない部分が多いが, アラキドン酸カスケードが重要な役割を果たすことが知られている。COX-2 は アラキドン酸カスケードで遊離アラキドン酸を代謝しプロスタグランディンを 合成する。COX-2 を抑制するためにインドメタシンを投与したラットでは,子 宮の着床間隔に異常がみられることが報告されている(49)。また,近年の研究 でリゾホスファチジン酸(LPA)受容体 LPA3(50, 51)及び細胞質型フォスフォリ パーゼ A2(cPLA2)(52)が着床時期及び配列に関与していることが明らかと なった。LPA3は G タンパク質共役型受容体であり,アラキドン酸カスケード において重要な役割を果たすことが知られている。また,cPLA2はアラキドン 酸代謝と密接に関連しており,生体膜リン脂質からアラキドン酸を選択的に遊 離する。LPA3または cPLA2のホモ欠損マウスでは,着床時期の遅延,着床部 位の配列の異常及び一腹あたりの胎児数が減少することが報告されている(50, 52)。今のところ高週齢のラットの子宮におけるアラキドン酸カスケードに関す る報告はみあたらないが,本研究で明らかとなった加齢に伴う着床間隔の異常 にもアラキドン酸カスケードの異常が関与する可能性が考えられた。 妊娠 11 日の 37-41 週齢ラットから得られた胚を形態学的に観察した結果,外 胎盤円錐の形態異常だけでなく頭褶及び心臓原基の形成がみられなかった。こ
37 のことから,胚の構造異常は特定の器官ではなく,様々な器官にわたることが 確認された。また,別の胚では器官の構造が確認できないものがあったことか ら,胚の構造異常は複数の因子によるもので,同一母体でも胚によって異常な 発生に関わる因子は異なる可能性が考えられた。 以上のように,37-41 週齢のラットでは妊娠初期から中期にかけて着床及び 胚の発育・分化に異常がみられた。しかし,これらの異常が母動物の子宮環境 や内分泌状態と胚のどちらに起因するものなのか明らかではない。本研究でみ られた異常が母動物由来か胚由来か明らかにするには,37-41 週齢のラットと 14 週齢のラットで受精卵の交換移植をする等のさらなる研究が必要である。
38
Table 3-1:14 週齢のラットにおける受胎率(Fertility index),黄体数(No. of corpus luteum), 着床数(No. of implantation site) 及び着床率(implantation rate)
受胎率=着床がみられた雌数/膣栓がみられた雌数,着床率=着床数/黄体数
38
No. of corpus luteum
individual data individual
data individual data individual data
F1 15 14 93.3 6.7 F2 13 13 100.0 0.0 F3 17 12 70.0 29.4 F11 15 14 93.3 6.7 F12 16 14 87.5 12.5 F13 20 20 100.0 0.0 F14 14 12 85.7 14.3 F4 15 14 93.3 6.7 F5 13 11 84.6 15.4 F6 17 17 100.0 0.0 F15 16 16 100.0 0.0 F16 12 12 100.0 0.0 F17 15 14 93.3 6.7 F18 13 11 84.6 15.4 F19 16 15 93.8 6.3 F20 13 13 100 0.0 F7 15 15 100 0.0 F8 21 12 57.1 42.9 F9 20 19 95.0 5.0 Gestation day Animal No. Fertility index (%)
No. of imprantation site implantation site (%) Pre-implantation loss (%)
mean±S.D mean±S.D mean±S.D mean±S.D
Day 5 100.0 15.0 ± 2.0 13.0 ± 1.0 87.8 ± 15.7 12.0 ± 15.4
Day 6 100.0 16.3 ± 2.6 15.0 ± 3.5 91.6 ± 6.5 8.4 ± 6.5
Day 7 100.0 14.6 ± 2.1 14.0 ± 2.5 95.6 ± 6.8 4.4 ± 6.8
Day 8 100.0 14.3 ± 1.5 13.3 ± 1.7 92.9 ± 6.3 7.1 ± 6.3
39
Table 3-2:37-41 週齢のラットにおける受胎率(Fertility index),黄体数(No. of corpus luteum), 着床数(No. of implantation site) 及び着床率(implantation rate)
NP:不妊動物(non-pregnancy),-:不妊動物のためデータ無し,受胎率=着床がみられた雌数/膣栓がみられた雌数, 着床率=着床数/黄体数
39
individual data individual
data individual data individual data
175 16 13 81.3 18.8 179 15 15 100 0.0 128 14 14 100 0.0 FA-5 16 13 81.3 18.8 FA-6 17 11 64.7 35.3 FA-7 14 13 92.9 7.1 FA-9 15 15 100 0.0 FA-10 11 7 63.6 36.4 H18 13 12 92.3 7.7 178 13 13 100.0 0.0 116 16 16 100.0 0.0 165 16 14 87.5 12.5 149 13 12 92.3 7.7 139 (NP) 13 0 0 0 FA-4 12 10 83.3 16.7 F217 19 18 94.7 5.3 177 (NP) 13 - - 100 168 (NP) 16 - - 100.0 143 12 10 83.3 16.7 Gestation day Animal No. Fertility index (%)
No. of corpus luteum No. of imprantation site implantation site (%) Pre-implantation loss (%)
mean±S.D mean±S.D mean±S.D mean±S.D
Day 5 100.0 15.6 ± 1.1 13.2 ± 1.5 85.5 ± 14.9 14.6 ± 14.9 Day 6 100.0 13.3 ± 1.7 11.8 ± 3.4 87.2 ± 16.1 12.8 ± 16.1 Day 7 80.0 14.2 ± 1.6 11.0 ± 6.3 76.0 ± 42.8 4.0 ± 5.8 8.1 11.0 Day 8 100.0 15.5 ± 4.9 14.0 10.0 ± 5.7 89.0 ± Day 12.5 25.0 13.7 ± 2.1 83.3 72.2 ± 48.1 ± 8.1
40 Figure 3-1:14 週齢ラットと 37-41 週齢ラットの受胎率の比較 14 週齢及び 37-41 週齢の妊娠ラットについて妊娠 5,6,7,8 及び 12 日目 に剖検し,受胎率(Fertility index:着床がみられた雌数/膣栓がみられた雌 数)を妊娠日毎に算出した(14 週齢:妊娠 5,6,7,8 および 12 日 N= 3,4,5,4 および 3,37-41 週齢:妊娠 5,6,7,8 および 12 日 N=5,4, 5,2 および 4)。
41 Figure 3-2:14 週齢ラットと 37-41 週齢ラットの着床前死亡率の比較 14 週齢及び 37-41 週齢の妊娠ラットについて妊娠 5,6,7,8 及び 12 日目 に剖検し,着床前死亡率(Pre-implantation loss:(黄体数-着床数)/黄体数 ×100)を妊娠日毎に算出した(14 週齢:妊娠 5,6,7,8 および 12 日 N =3,4,5,4 および 3,37-41 週齢:妊娠 5,6,7,8 および 12 日 N=5, 4,5,2 および 4)。
42
A B C D E
F G H I J
Figure 3-4:14 週齢のラットと 37-41 週齢のラットの子宮
A-E:14 週齢のラット,F-J:37-41 週齢のラット,A・F:妊娠 5 日目,B・G:妊娠 6 日目,C・H:妊娠 7 日目,D・I: 妊娠 8 日目,E・J:妊娠 12 日目,矢印:エバンスブルー陽性(着床部位)
4
43
A B C
Figure 3-5:妊娠 6 日目の胚の組織像
A:14 週齢のラットから得られた正常な胚,ep:ectoplacental cone(外胎 盤円錐), imc:inner cell mass(内細胞塊), ysc:yolk-sac cavity(卵黄嚢空), *脱落膜,B・C:37-41 週齢のラットから得られた異常胚(▲),B:構造 異常を示す胚,C:発達遅延を示す胚
44 Figure 3-6:妊娠 7 日目の胚の組織像 A-C:14 週齢のラット,D-F:37-41 週齢のラット,A・D:着床部位の断面(x4), A’・D’:14 週齢のラットでは着床周囲の子宮内膜に毛細血管(△)の新生が多 くみられるが(A’),37-41 週齢のラットでは毛細血管の新生が乏しい(B’),B・ E(x20):14 週齢のラットでは外胎盤円錐が発達し胚が子宮内膜へ浸潤する様 子がうかがえるが(B,*:脱落膜),37-41 週齢のラットでは外胎盤円錐の発 達が乏しく,子宮内膜への浸潤も 14 週齢に比べて弱い(E)。C・F(x40):14 週齢のラットの外胎盤円錐(ep:ctoplacental cone)には赤血球(△)の浸潤が認 められるが(C),37-41 週齢のラットの外胎盤円錐に赤血球の浸潤は認められ ない(F)。
45
A
B
Figure 3-7:妊娠 8 日目の胚の組織像
A:14 週齢のラットでみられた正常胚,B:37-41 週齢のラットでみられた 異常胚,ep:ectoplacental cone(外胎盤円錐),ec:ectroderm(外胚葉),m:
mesoderm(中胚葉),en:endoderm(内胚葉),ep:ectoplacental cone(外胎
46 Figure 3-8:妊娠 11 日目の胚の写真 A・B:14 週齢でみられた正常胚,B は A を赤矢印方向からみた写真,C-E:37-41 週齢でみられた異常胚,C:正常胚でみられる頭褶,心臓原基及び体節は確認出来 ず,外胎盤円錐は正常胚に比べて小さく形態学的な異常が認められる。D:C を赤 矢印方向からみた写真,E:全体的な構造異常が認められた胚,hf:head fold(頭褶),
47 第 4 章 視床下部 kisspeptin 神経のエストロゲンに対する活性の低下が引き起こ す加齢性 LH サージの消 失 1. 背景と目的 第 2 章で,雌ラットの加齢に伴う生殖機能の低下のうち発情周期の異常が早 期にみられ,排卵障害が起きていることが推測された。LH サージは排卵に必須 であり,LH サージの消失は排卵障害を引き起こす。また,ラットでは LH サー ジの減弱は早期の加齢性マーカーとして知られている(10-12)。しかし,LH サー ジが加齢に伴い減弱する詳細なメカニズムは明かではない。
視床下部前腹側周囲核(anteroventral periventricular nucleus:AVPV)に分布す る kisspeptin 神経は GnRH 神経を活性化して GnRH を放出させることで,LH サ ージを誘起することが最近の研究で明らかとなった(53, 54)。 本章では AVPV に分布する kisspeptin 神経に着目し,雌ラットの老化過程にお ける LH サージ減弱のメカニズムについて検討した。 2. 材料及び方法 2.1 使用動物 日本チャールスリバー株式会社で生産された雌 Crl:CD(SD)ラットを購入し, 単飼ケージ[647.7 cm2(底面積)×17.4 cm(高さ)]で SPF 環境下で飼育した。動物 室の照明時間は 12 時間/日(点灯時間: 7 時~19 時)とし,飼料はγ線照射し た固形飼料(オリエンタル酵母株式会社,東京,日本)を与え,水道水を自由 摂取下で飼育した。実験開始の週間前から単飼ケージにて飼育し,第章と同 様の方法で膣スメアを採取し発情周期を観察した。発情周期及び月齢に基づい て雌ラットを,正常発情周期を示す若いラット(Young rats with Normal estrous cycles: YN,3-4 ヶ月齢),正常発情周期を示す高週齢のラット(Middle-age rats with Normal estrous cycles: MN,9-13 ヶ月齢)及び異常発情周期(持続発情型異常発 情期)を示す高週齢のラット(Middle-age rats with Irregular estrus: MI,9-13 ヶ月 齢)の 3 グループに分けた。全ての動物は武田薬品工業株式会社が定める動物 の取り扱いに関するガイドラインに従って取り扱った。
48 2.2 血漿サンプルの採取方法 無麻酔下で頚静脈からヘパリン加注射筒を用いて採血し,速やかに遠心分離 (g,4℃,2 分間)して血漿サンプルを得た。採取した血漿サンプルはホ ルモン測定に用いるまで-50℃で保存した。 2.3 血漿中 LH 及びエストラジオール濃度測定 YN(N=7)及び MN(N=38)について,発情前期の 16 時,17 時,18 時及び 19 時に採血し,全ポイントのサンプルについて LH 濃度を測定し,16 時のサン プルについてのみエストラジオール濃度を測定した。MI(N=30)は膣スメアに 周期性がみられないため,YN 及び MN と同時刻に採血を実施した。また,MN については採血から 3-4 週間発情期を観察し,採血後 3-4 週間以上正常発情期を 示したラットを MN-Normal(MN-N,N=31)とし,採血後 3-4 週間以内に異常 発情期を示したラットを MN-Irregular(MN-I,N=7)とした。 また,別のラット(YN,MN 及び MI:N=7/群)を用いて発情休止期の 18 時, 発情前期の 8 時,10 時,12 時及び 14 時に採血し,血漿中エストラジオール濃 度の変化を調べた。血漿中 LH 濃度の測定と同様で MI は YN 及び MN と同時に 採血を実施した。
2.4 AVPV における Kiss1 mRNA 発現量及び kisspeptin 含有量の測定
YN,MN 及び MI 群のそれぞれ 9,7 及び 5 例について,Real-time PCR を用い て AVPV の Kiss1 mRNA を測定した。YN 及び MN 群は発情前期の午後に,MI 群は YN 及び MN 群と同じ時間帯に CO2麻酔下で放血し,大脳を摘出した。摘
出した大脳は速やかに脳スライサー(ブレインサイエンスイディア株式会社, 大阪,日本)を用いて前頂 0 mm から尾側へ 2 mm の厚さで切り出した。そして, 直径 1.5 mm の生検パンチ(カイインダストリーズ株式会社,東京,日本)を用 いて,AVPV を含む領域をパンチアウトした。得られた脳サンプルは Kiss1 mRNA を測定するまで-80℃で保存した。残った脳切片は一晩 10%ホルマリンで固定し, その後 4℃で 30%グルコース溶液に 2 日間置換した後に,冠状面で 40 µm の厚さ
49
に薄切した。作製した脳切片を Thionin 染色し,AVPV を含む領域が適切にパン チアウトされているかどうかを確認した結果,全例について適切に AVPV が採 材されていることを確認した。
YN,MN 及び MI それぞれ 7 匹の動物から AVPV を含む脳サンプルを採取し し,enzyme immunoassay(EIA)により kisspeptin 含有量を測定した。剖検時間 及び AVPV を含む脳サンプルの採取は,上記の Kiss1 mRNA 測定と同様に実施 した。
2.5 GnRH 神経のヒト kisspeptin(Kp-54)に対する反応
Kp-54 は Ohtaki らの方法により合成した(14)。YN 及び MN 群は発情前期の 11 時に,MI 群は他の 2 群と同じタイミングで 10 または 100 nmol/kg の kisspeptin を背部に単回皮下投与し,投与前,投与 1,2 及び 4 時間後の血漿中 LH 濃度を 測定した。kisspeptin は 0.9%生理食塩水で溶解し,対照群には 0.9%生理食塩水 を同様に投与した。
2.6 卵巣を摘出したラットにおけるエストラジオールによる AVPV の kisspeptin 神経の活性化
in situ hybridization(Kiss1 mRNA)及び免疫染色(cFos)の二重染色を実施し, AVPV における kisspeptin 神経の活性化の程度を評価した。 YN,MN 及び MI のそれぞれ 4,5 及び 5 例の両側の卵巣を摘出し,その 1 週 間後に粉末のエストラジオール(Sigma-Ardrichi Japan,東京,日本)を充填した シリコンチューブ(株式会社カネカメディックス,大阪,日本,内径 2.0 mm, 外径 3.0 mm,長さ 2.5 cm)を背部皮下に移植した。全ての外科的施術は 2%イソ フルラン麻酔下で実施した。なお,卵巣を摘出しエストラジオールチューブを 移植したラットを以下 OVX+E2ラット(Ovariectomized rats with supplementation of
estradiol-17)と表記した。エストラジオールチューブ移植 3 日後に採血し,血 漿中エストラジオール濃度を測定した。さらに,大脳サンプルを得るため,採 血後の 14-15 時の間にチオペンタール麻酔下で 4%パラホルムアルデヒド
![Figure 2-5:母動物の加齢による着床前死亡率及び着床後死亡率の変化 妊娠 20 日目に剖検し,妊娠が確認されたラットについてそれぞれ着床前死 亡率[A,Pre-implantation loss rate, 着床前死亡率=(黄体数‐着床数)/黄体 数×100]及び着床後死亡率[B, Post-implantation loss rate,着床後死亡率=(着 床数-生存胎児数)/着床数×100]を個体ごとに算出し,週齢毎に平均値及 び SD を算出した。6,8,10,15,19,23,27,31,](https://thumb-ap.123doks.com/thumbv2/123deta/10089002.1489951/26.892.240.615.200.709/Figure動物加齢による着床前死亡率及び着床ラットについてそれぞれ.webp)