哺乳類受精卵カルシウム振動におけるカルシウム流入の 機能的関与に関する研究
高橋 徹
電気通信大学大学院 電気通信学研究科 博士(理学)の学位申請論文
2013 年 3 月
哺乳類受精卵カルシウム振動におけるカルシウム流入の 機能的関与に関する研究
博士論文審査委員会
主査 白川 英樹 准教授
委員 丹羽 治樹 教授
委員 中村 整 教授
委員 樫森 与志喜 教授
委員 狩野 豊 教授
著作権所有者
高橋 徹
2013 年
On the Role of Calcium Influx in the Generation of Calcium Oscillations in Mammalian Fertilized Eggs
Tooru Takahashi
Abstract
On mammalian fertilization, long-lasting Ca2+ oscillations are induced in the egg by the fusing spermatozoon. While each transient Ca2+ increase in Ca2+ concentration ([Ca2+]) in the cytosol is due to Ca2+ release from the endoplasmic reticulum (ER), Ca2+ influx from outside is required for Ca2+
oscillations to persist. In this study, it was investigated how Ca2+ influx is interrelated to the cycle of Ca2+ release and uptake by the intracellular Ca2+ stores during Ca2+ oscillations in fertilized mouse eggs. In addition to monitoring cytosolic [Ca2+] with fura-2, the influx rate was evaluated using Mn2+ quenching technique, and the change in [Ca2+] in the ER lumen was visualized with a targeted fluorescent probe. It was found that the influx was stimulated after each transient Ca2+ release and then diminished gradually to the basal level, and was demonstrated that the ER Ca2+ stores once depleted by Ca2+ release were gradually refilled until the next Ca2+ transient to be initiated. Experiments altering extracellular [Ca2+] in the middle of Ca2+ oscillations revealed the dependence of both the refilling rate and the oscillation frequency on the rate of Ca2+ influx, indicating the crucial role of Ca2+ influx in determining the intervals of Ca2+ transients. As for the influx pathway supporting Ca2+ oscillations to persist, STIM1/Orai1-mediated store-operated Ca2+ entry (SOCE) may not significantly contribute, since neither known SOCE blockers nor the expression of protein fragments that interfere the interaction between STIM1 and Orai1 inhibited the oscillation frequency or the influx rate. The results in this thesis provide the insight on the molecular mechanisms for the generation of Ca2+ oscillations in fertilized mammalian eggs, in fertilization of other species, or even in other types of cells in general.
哺乳類受精卵カルシウム振動における カルシウム流入の機能的関与に関する研究
高橋 徹
要旨
本論文は、哺乳類受精卵に見られる周期的な細胞質内Ca2+濃度の変動(Ca2+
振動)の発生メカニズムの解明を目指した研究の一環として、マウス受精卵に おける細胞外からの Ca2+流入の活性化の時間経過の詳細を明らかにすること、
Ca2+流入経路の実体およびその制御機構を探究すること、および Ca2+流入が Ca2+振動の発生・維持において担う役割を明らかにすることを目的として行っ た研究の結果をまとめたものである。主として蛍光プローブを用いた細胞生理 学的実験により、Ca2+振動中のCa2+流入速度の変化、および細胞内Ca2+ストア である小胞体内のCa2+濃度変化やそのCa2+流入に対する依存性を解析した結果 から、Ca2+流入はCa2+ストアの再充填速度に影響し、Ca2+振動の周期を規定す る因子として機能すると結論づけている。
まず第1章では、本研究の背景として、各種細胞における細胞機能の調節因 子としての Ca2+の普遍性および哺乳類卵の初期発生をトリガーする因子とし ての重要性について述べている。また、細胞内Ca2+濃度の制御機構として、Ca2+
遊離とCa2+流入という2つの経路の分子的基盤について述べている。哺乳類受 精卵でのCa2+上昇反応の特徴的な時空間的パターンである Ca2+波と Ca2+振動
についての現象論的な記載をし、その発生メカニズムについての現在までの知 見について述べた上で、Ca2+流入の役割と制御機構が未解明であることを指摘 し、それらの探究を本論文の目的として位置づけている。
第2章では、精子によって誘発されるマウス受精卵での Ca2+振動の Ca2+流 入速度に対する依存性を調べた実験について述べられている。受精卵でのCa2+
振動を蛍光性Ca2+プローブfura-2を用いたCa2+イメージングにより経時的に 計測しつつ、外液の Ca2+濃度を変えて Ca2+流入速度を増減させた実験から、
Ca2+振動の頻度はCa2+流入速度に比例して変化するに対し、振幅は全く影響さ れないことが示された。この結果より、Ca2+流入はCa2+遊離後の枯渇した細胞 内 Ca2+ストアを Ca2+遊離が起こり得る元のレベルまで再充填する過程に寄与 しているという仮説を提唱している。
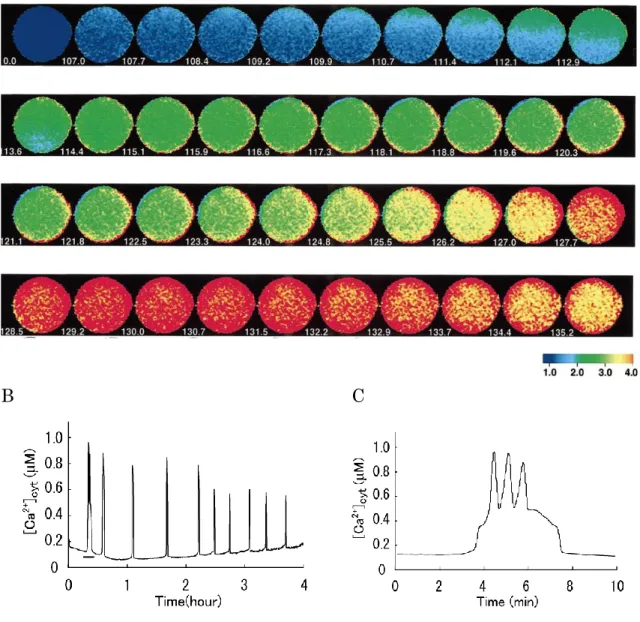
第3章では、受精卵でのCa2+振動時のCa2+流入速度の時間的な変動をMn2+
消光法を用いて解析した実験について述べられている。Mn2+消光法とは、蛍光 性Ca2+プローブであるfura-2がMn2+と結合すると蛍光性を失うことを利用し、
細胞膜のCa2+チャネルを介したMn2+の流入速度を、細胞内のfura-2の蛍光強 度の減少速度として測定するものである。外液に投与した Mn2+の流入速度を Ca2+振動中のCa2+遊離との時間的な関係に注目して解析した結果、流入速度は Ca2+遊離直後に約3倍に増加し、その後およそ 10 分の時定数で元のレベルま で戻ることを明らかにした。このことから、マウス受精卵ではCa2+遊離によっ て一過性に活性化されるCa2+流入経路が存在していると結論づけている。
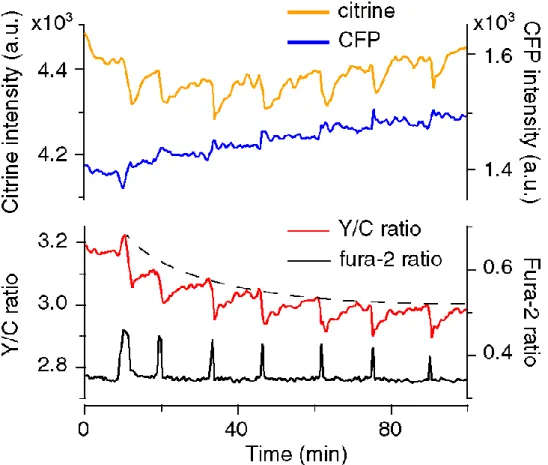
第4章では、受精卵でのCa2+振動時の細胞内Ca2+ストア(小胞体)内のCa2+
濃度変化を測定した実験について述べられている。この実験では、小胞体内腔
への局在化シグナルを付加したタンパク性Ca2+プローブを用いることで、マウ ス卵での小胞体内 Ca2+濃度変化を測定することに初めて成功した。小胞体内 Ca2+濃度は、Ca2+振動開始直後からその基底レベルが徐々に低下すること、ま た個々の Ca2+遊離に伴い一過性に減少した後に約7分の時定数で基底レベル まで戻ることを明らかにした。さらにこのストア再充填の速度は細胞外からの Ca2+流入速度に比例することを示し、Ca2+振動におけるCa2+流入の役割につい て第2章で提唱した仮説を実験的に立証した。
第5章では、受精時の Ca2+振動の維持に関わる Ca2+流入経路として、スト ア作動性Ca2+流入(SOCE)の関与について検証した一連の実験が述べられて いる。これまでマウス卵でのSOCE経路の存在を示した報告があること、また Ca2+振動中の Ca2+流入速度と小胞体内 Ca2+濃度の時間的変化が類似している こと(本論文第3および4章)は、受精卵で Ca2+振動の維持に SOCE が関与 していることを示唆している。しかし、SOCEの阻害剤はCa2+振動の頻度や振 幅に影響せず、さらにSOCEを担うSTIM1/Orai1 タンパク質の相互作用を阻 害するペプチドを細胞質に発現させても、Ca2+振動およびCa2+流入速度は全く 影響されなかった。これらの結果から、マウス受精卵ではSOCEは活性化され ているとしてもその程度はわずかであり、SOCE経路を介したCa2+流入は受精 時のCa2+振動の維持には有意には寄与していないと結論している。
最後に第6章では、本研究で得られた結果および考察をまとめ、今後の展望 について述べられている。
目 次
第1章 背景 ··· 1
1.1 緒言 ··· 1
1.2 細胞内Ca2+による細胞機能の調節 ··· 3
1.3 哺乳類受精卵におけるCa2+の役割 ··· 4
1.4 細胞内Ca2+濃度の制御機構 ··· 8
1.4.1 Ca2+流入経路 ··· 10
1.4.2 Ca2+遊離経路 ··· 12
1.5 哺乳類受精卵におけるCa2+動態 ··· 13
1.5.1 Ca2+波とCa2+振動 ··· 13
1.5.2 Ca2+遊離誘発機構 ··· 15
1.5.3 Ca2+流入経路 ··· 17
1.6 本研究の目的 ··· 18
第2章 受精卵Ca2+振動のCa2+流入依存性 ··· 21
2.1 目的 ··· 21
2.2 方法 ··· 22
2.2.1 成熟卵および精子の採取 ··· 22
2.2.2 Ca2+イメージング ··· 23
2.3 結果 ··· 25
2.4 考察 ··· 28
第3章 受精卵におけるCa2+流入の活性化 ··· 32
3.1 目的 ··· 32
3.2 方法 ··· 33
3.2.1 成熟卵および精子の採取 ··· 35
3.2.2 Mn2+消光法 ··· 35
3.3 結果 ··· 36
3.4 考察 ··· 40
第4章 細胞内Ca2+ストア再充填に対する Ca2+流入の影響 ··· 43
4.1 目的 ··· 43
4.2 方法 ··· 45
4.2.1 RNA合成 ··· 45
4.2.2 未成熟卵および精子の採取 ··· 47
4.2.3 卵細胞へのRNA注入および体外成熟培養 ··· 48
4.2.4 D1ERの細胞内局在の観察 ··· 48
4.2.5 細胞質Ca2+と小胞体内Ca2+の同時測定 ··· 48
4.3 結果 ··· 49
4.3.1 D1ERの細胞内局在 ··· 50
4.3.2 受精時の小胞体内Ca2+濃度の変化 ··· 50
4.3.3 小胞体再充填に対するCa2+流入速度の影響··· 53
4.4 考察 ··· 55
第5章 細胞内Ca2+振動におけるSTIM/Orai系の関与 ··· 59
5.1 目的 ··· 59
5.2 方法 ··· 62
5.2.1 STIM1とOrai1のcDNAクローニング ··· 62
5.2.2 成熟卵および精子の採取 ··· 63
5.2.3 卵細胞へのRNA注入 ··· 66
5.2.4 Ca2+イメージングおよびMn2+消光法··· 66
5.3 結果 ··· 66
5.3.1 ストア作動性Ca2+流入に対するCa2+チャネル阻害剤の影響 ··· 66
5.3.2 Orai1, STIM1の細胞質ドメインペプチドの影響 ··· 67
5.4 考察 ··· 72
第6章 討論および総括 ··· 78
謝辞 ··· 86
文献 ··· 87
第1章 背景
1.1 緒言
カルシウムは骨格を構成する主要元素であるが,細胞内ではカルシウムイオ ン(Ca2+)の形で存在してシグナル伝達物質として機能する.通常,細胞質の Ca2+濃度([Ca2+]cyt)は非常に低く保たれているが,細胞外から様々な刺激を受 けるとしばしば急激に上昇する.この[Ca2+]cytの上昇が細胞内での様々な反応の トリガーとなり,例えばホルモンや消化酵素の放出,筋肉の収縮,遺伝子の発 現などの細胞応答に至る(Campbell, 1983).
哺乳類の卵細胞でも,受精時に精子によって[Ca2+]cytの上昇反応が誘発される.
停止している減数分裂の再開や複数の精子との受精を防ぐ多精拒否の機構にお いて,Ca2+が中心的なシグナル因子としての役割を担っている(Ducibella &
Fissore, 2008).その生理的重要性から,受精卵での細胞内 Ca2+の時間的・空 間的動態およびその制御メカニズムについては,長く研究が行われている.受 精卵での[Ca2+]cyt上昇はまず精子付着点付近から開始し,それが卵内全体に波状 に伝播する(Ca2+波,Miyazaki et al., 1992; Deguchi et al., 2000) [Ca2+]cyt
上昇は一度静止時のレベルに戻るが,以後この一過性の[Ca2+]cyt上昇が数時間に わたって周期的に繰り返される(Ca2+振動,Miyazaki, 1991; Deguchi et al., 2000).個々の[Ca2+]cyt上昇は細胞内 Ca2+ストアからの Ca2+遊離によるもので あることが分かっている(Miyazaki, 1991).すなわち,精子・卵融合後に精子 細胞内のホスフォリパーゼ C(PLC)が卵細胞質に移行してイノシトール三リ
ン酸(IP3)を産生し,小胞体膜上にあるIP3受容体(IP3R)/Ca2+チャネルが開 口して小胞体内腔からCa2+が放出され,[Ca2+]cytが上昇する(Miyazaki et al., 1993; Saunders et al., 2002).増加した細胞質のCa2+は,その後Ca2+ポンプの 作用により小胞体に再び取り込まれ,[Ca2+]cytは速やかに元のレベルに戻る.
一方で,一過性の[Ca2+]cyt上昇が長時間にわたり周期的上昇を繰り返す,即ち Ca2+振動が生じるメカニズムは,未だ解明されていない.種々の体細胞の細胞 応答でもしばしば数十秒周期の Ca2+振動が生じるが(Berridge & Dupont.,
1994),受精卵での10~30分周期のCa2+振動は,これらとは異なるメカニズム
で発生するものと考えられる.これまでの研究により,哺乳類受精卵でのCa2+
振動には細胞外からの Ca2+流入も必要であることが示されている(Igusa &
Miyazaki, 1983; Kline & Kline, 1992).[Ca2+]cyt上昇の際,Ca2+ストアから放 出された Ca2+はすべてストアに再取り込みされるわけではなく,その一部は細 胞外へと排出される.その分の不足したCa2+を補うためにCa2+流入が必要だと 考えられている.また,受精卵では未受精卵に比べて Ca2+流入が促進されてい ることが示唆されているが(McGuinness et al., 1996),Ca2+流入経路の実体や その制御機構は現在全く不明である.
哺乳類受精卵の発生開始は,一度十分大きな[Ca2+]cyt上昇がおこればトリガー され得るので,その意味ではCa2+振動は必須ではない.しかし,Ca2+振動のパ ターンは初期胚における遺伝子発現や発生率に影響する(Ducibella et al.,
2002).従って,正常な Ca2+振動が発生し維持されるための細胞内メカニズム
を解明することは,哺乳類発生学における基礎的知見としてだけでなく,医療 や畜産分野における生殖医学・工学的応用にとっても重要である.本研究は,
Ca2+振動のメカニズムの全容の解明を目指す研究の一環として,マウス受精卵 でのCa2+振動におけるCa2+流入の役割を明らかにすること,Ca2+流入活性化の 時間的変化の詳細を解明すること,およびCa2+振動の維持に関与するCa2+流入 経路の実体とその制御機構を探究することを目的として行った.
1.2 細胞内 Ca2+による細胞機能の調節
Ca2+は最も重要な細胞内シグナル伝達物質の一つである(図1.1)(Berridge &
Dupont, 1994).様々な細胞種において,外界からの刺激に応答して[Ca2+]cytが 上昇する.細胞内の Ca2+は様々なタンパク質に結合することでそれらの機能を 修飾し,その結果として様々な細胞応答を誘引する.
例えば,Ca2+は筋肉細胞の収縮を誘発する(Ebashi & Kodama, 1965).骨格 筋細胞に支配神経からの入力があると,筋細胞の[Ca2+]cyt が急激に上昇する.
Ca2+はアクチンフィラメント結合タンパク質であるトロポニンC に結合する.
Ca2+の結合によりトロポニンCの構造が変化し,アクチンフィラメントとミオ シンの相互作用が可能になることで,筋収縮が起こる.平滑筋細胞では Ca2+は カルモジュリン(CaM)に結合する.CaMは様々な酵素タンパク質やイオンチ ャネルの活性を制御するCa2+結合タンパク質であり(Cheung, 1970; Kakiuchi
& Yamazaki, 1970),平滑筋ではミオシン軽鎖リン酸化酵素(MLCK)を活性 化する(Kamm et al., 2001).MLCKによってミオシン軽鎖がリン酸化される と,アクチンとミオシンの相互作用によって平滑筋が収縮する.
神経細胞,内分泌・外分泌細胞,血液細胞などでは,神経伝達物質,ホルモ ン,酵素などが分泌顆粒の細胞膜への融合,即ち開口分泌(エキソサイトーシ ス)によって細胞外に分泌される.この開口分泌も[Ca2+]cyt上昇によってトリガ ーされる(Augustine, 2001).例えば神経終末のシナプスでは,活動電位によ って[Ca2+]cytが上昇することにより神経伝達物質がシナプス間隙に分泌される.
Ca2+はシナプトタグミンと結合することで,シナプス小胞と細胞膜の融合をト リガーすると考えられている(Yoshihara et al., 2002).一方,後シナプス神経 細胞ではCa2+はシナプスの可塑性の調節に関わる(Lynch, 2004).神経伝達物 質の一種であるグルタミン酸が後シナプス膜上の NMDA 受容体チャネルによ って受容されると,シナプス小頭(synaptic bouton)と樹状突起での[Ca2+]cyt
が上昇する.Ca2+はシナプス小頭の構造変化を誘発し,その結果シナプス伝達 効率の長期増強がおこる(Sweatt, 1999).このシナプス伝達効率の可塑性が,
脳における学習・記憶といった高次機能の基礎となると考えられている.
これら以外にも,細胞内のCa2+は,細胞分裂やアポトーシスの制御(Allen et
al., 2005),免疫応答,さらには遺伝子発現調節を介した細胞分化の調節など,
ミリ秒から時間,日という幅広い時間スケールにわたる,多彩な生理現象の制 御に関わっている(図 1.1).
1.3 哺乳類受精卵における Ca2+の役割
哺乳類受精卵においても細胞内Ca2+は重要な役割を担う(Miyazaki, 2006). 成熟した哺乳類の卵細胞は第二減数分裂中期で細胞分裂が停止している.受精
細胞内 Ca 2+
の上昇 細胞内 Ca 2+
の上昇
神経細胞
シナプス伝達 可塑性
神経細胞
シナプス伝達 可塑性
内分泌細胞 外分泌細胞
ホルモン・酵素 の分泌
内分泌細胞 外分泌細胞
ホルモン・酵素 の分泌
卵細胞
細胞周期再開 多精拒否
卵細胞
細胞周期再開 多精拒否
筋肉細胞 収 縮 筋肉細胞
収 縮
図1.1 細胞内Ca2+が関与する細胞機能
[Ca2+]cyt上昇により引き起こされる生理現象の例を示した.これらの例の他に
も発生・分化,細胞増殖,免疫機能などにも関与している.
時に精子によって[Ca2+]cytの上昇が誘発され,それによって細胞分裂が再開して,
胚発生がスタートする.胚発生開始のトリガーとして Ca2+は必要かつ十分なシ グナル因子である.すなわち,卵細胞内に Ca2+キレート剤をいれることで受精 時の[Ca2+]cyt 上昇を抑制すると,細胞分裂は再開されない(Coticchio et al., 1998).逆に未受精卵の[Ca2+]cytを人為的に上昇させるだけで胚発生が開始され,
2細胞期程度までは進行する(Kono et al., 1996).
哺乳類の未受精卵では,Cdk1/サイクリン B1 複合体(=中期促進因子 metaphase promoting factor; MPF)の作用により分裂周期が停止している.受 精時の[Ca2+]cyt上昇は,以下の様な経路によりサイクリンB1の分解を誘発する ことで細胞分裂を再開させると考えられている(図 1.2 A).まずCa2+を結合し たCaMがCSF(cytostatic factor)を抑制する(Lorca et al., 1993).その結果,
CSFによって抑制されていた後期促進複合体(anaphase promoting complex;
APC)が活性化される(Reimann et al., 2002).活性化されたAPCによってサ イクリ ン B1 がユビ キチン 化さ れ, プロ テアソ ーム によ って 分解さ れる
(Tunquist et al., 2003).
受精卵での[Ca2+]cyt上昇は,複数の精子との受精を防ぐ多精拒否反応を誘発す るためにも必須である(図1.2 B).哺乳類卵の周囲には,糖タンパク質を主成 分とする透明帯と呼ばれる層が存在する.精子は未受精卵の透明帯を通過した 後,卵細胞膜に接着し融合(すなわち受精)する.その後卵の[Ca2+]cytが上昇す るのに伴い,卵表層部にある表層顆粒とよばれる分泌小胞の開口分泌が起こる.
分泌された表層顆粒の内容物のうち,オバスタシンというプロテアーゼが透明 帯の糖タンパク質ZP2を分解する(Burkart et al., 2012).その結果,透明帯
A
第一減数分裂前期
卵核胞 第一極体 第二極体
第二減数分裂中期
(細胞周期は停止)
第二減数分裂後期
受精→Ca2+上昇
成体
卵割
サイクリンB1 Cdk1 抑制
CSF
APC 抑制
MPF
第二減数分裂 中期
第二減数分裂 後期 細胞周期は停止
Cdk1 CSF
APC 抑制
第二減数分裂 中期
第二減数分裂 後期 細胞周期は進行 受精→Ca2+上昇
サイクリンB1 Cdk1 CSF
APC 抑制
CaM
(活性型)
ユビキチン化
分解 ユビキチン
CaM
(不活性型) CaM
(不活性型)
Ca2+
B
受精→Ca2+上昇 精子
卵細胞 精子
卵細胞
表層顆粒 透明帯
卵細胞 オバスタシン
ZP2 ZP2 ZP2 ZP2 ZP2 ZP2
開口分泌
ZP2の分解 透明帯が変性
図1.2哺乳類受精卵におけるCa2+の役割 (A)細胞周期再開の分子機構.(B)多精拒否機構.
の性質が変化し,以後の精子は透明帯を通過できなくなる.
上述以外にも,前核の形成やそれ以後の胚発生,また卵細胞膜の変質を介し た多精拒否応答なども,卵内の Ca2+によって制御されているという報告がある
(Vitullo et al., 1992; Sun, 2003; Liang et al., 2011).
1.4 細胞内 Ca2+濃度の制御機構
細胞外のCa2+濃度([Ca2+]o)はmMのオーダーであるのに対し,[Ca2+]cytは
100 nM程度と,通常は1万分の1以下に低く保たれている.これは主として細
胞外へのCa2+の能動輸送と,細胞小器官である小胞体の内腔へのCa2+の能動輸 送による(図 1.3).細胞膜上には Ca2+ポンプ(Ca2+-ATP アーゼ)が存在し,
アデノシン三リン酸(ATP)の加水分解エネルギーを使って Ca2+を細胞外に排 出している(Vincenzi et al., 1980).また, Na+/Ca2+交換輸送体も,細胞内外 のNa+濃度勾配を駆動力としてCa2+を細胞外に排出する(DiPolo et al., 2006). 一方小胞体膜上にもCa2+ポンプが存在し,Ca2+を細胞質から内腔へ能動的に取 り込 ん でいる(Strehler et al., 2001).そのため小胞体内腔の Ca2+濃度
([Ca2+]ER)は,数10~100 µMと高濃度になっている(Bygrave et al., 1996). 刺激に対する細胞応答の際に,[Ca2+]cytは1〜数M程度にまで急激に上昇す る. [Ca2+]cyt上昇にいたる経路には2通りあり(図1.3),その1つは細胞外か らのCa2+流入である.細胞膜上のCa2+透過性イオンチャネルが開口すると,細 胞内外の大きな濃度勾配に従って Ca2+が急激に細胞内に流入する.もう一つは 小胞体からのCa2+遊離であり,小胞体膜上のCa2+透過性イオンチャネルが開口
PLC PLC
PIP2 DAG
IP3 IP3
IP3受容体
小胞体
Ca2+
TRPCチャネル
Na+/Ca2+交換輸送体
Ca2+ Ca2+
Na+
Ca2+ATPアーゼ /Ca2+ポンプ
Ca2+
Ca2+ 小胞体
(低Ca2+濃度)
Ca2+
STIM
Orai ストア作動性
Ca2+チャネル 電位依存性 Ca2+チャネル
Ca2+
Ca2+ATPアーゼ /Ca2+ポンプ
小胞体
++++ ++++
Ca2+
リアノジン 受容体
Ca2+
図1.3 細胞内Ca2+の調節機構
細胞質のCa2+濃度は細胞外からのCa2+流入と細胞外への排出と,小胞体からの Ca2+放出や小胞体への取り込みにより調節されている.
すると,濃度勾配に従って小胞体内腔から細胞質へと Ca2+が放出される.この ように小胞体は,放出可能なCa2+を蓄えているため,細胞内Ca2+ストアとも称 される.
1.4.1 Ca2+流入経路
細胞膜上に存在する様々なCa2+透過性イオンチャネルが,細胞外からのCa2+
流入の経路として機能し得る(図1.3).
神経細胞や骨格筋・心筋細胞などには,電位依存性Ca2+チャネルが存在する.
膜電位は通常–60〜–90 mV程度に保たれているが,細胞の興奮時に脱分極する とこのチャネルが開き,細胞外からCa2+が流入する.電位依存性Ca2+チャネル は,活性化や不活性化の特性からT型,L型,N型,P/Q型,R型に分類され る(Dunlap et al., 1995).このうちL型Ca2+チャネルは骨格筋,心筋,血管,
脳などの細胞に存在する.骨格筋や心筋細胞では横行小管(T 管)部の細胞膜 に存在し,活動電位による脱分極を検出することで,興奮収縮連関に関わる.
心筋ではL型Ca2+チャネルがCa2+流入を担うとされているが,骨格筋ではL型 Ca2+チャネルと筋小胞体膜上のリアノジン受容体/Ca2+チャネルの直接相互作用 が重要であると考えられている(次節参照).神経細胞のシナプス前膜に存在す る N型Ca2+チャネルは,軸索からの活動電位の伝導に伴って神経末端が脱分極 するとき開口する.N型Ca2+チャネルを介したCa2+流入により,シナプス末端 部の[Ca2+]cytが急激に上昇し,前述のように神経伝達物質の開口分泌がおこる.
TRP(transient receptor potential)チャネルと呼ばれるCa2+透過性チャネ
ルも,Ca2+流入の経路として機能する.TRPは最初ショウジョウバエの光受容 応答変異株の原因遺伝子(trp)として見つかった(Montell & Rubin, 1989). 現在,TRP スーパーファミリーに属するイオンチャネルは,そのアミノ酸配列や 分子構造の類似性からTRPV, TRPC, TRPM, TRPP, TRPML, TRPN, TRPAの 7つに分類されている(Nilius et al., 2008).これらのTRP チャネルは,種々 の刺激によって活性化されることで,細胞における各種センサーとして機能す る.たとえば,TRPV ファミリーは唐辛子の辛み成分であるカプサイシンや熱 刺激(高温)によって活性化される(Caterina et al., 1997).TRPM ファミリ ーは酸化ストレス,温度変化(低温),pH の変化,機械刺激,浸透圧の変化等 で活性化し,Ca2+流入を引き起こす(Hara et al., 2002; Schlingmann et al., 2002; Behrendt et al., 2004).また,TRPCはPLCによるPIP2加水分解時に 生成されたDAGによっても活性化されることが報告されている(Martin et al., 2006).
伸展活性化チャネル(stretch-activated channel; SAC)は,細胞膜が伸ばさ れたときに,チャネルタンパク質にかかる力によって直接活性化される Ca2+透 過性チャネルである(Patel et al., 2010; Del Valle et al., 2012).外部力による 直接的な細胞の圧迫・伸展ばかりでなく,浸透圧変化,細胞分裂や成長,細胞 運動などによって細胞が変形する.これによってSACが開口してCa2+が流入す る.
さ ら に ,Ca2+遊 離 の 直 後 に 活 性 化 さ れ る Ca2+透 過 性 チ ャ ネ ル (Ca2+
release-activated channel; CRAC)も知られている(Hoth et al., 1992).中で
ストア作動性Ca2+流入(store-operated Ca2+ entry; SOCE)が関与する細胞応 答の例が,種々の細胞種において報告されている.細胞内 Ca2+ストアである小 胞体内の Ca2+濃度が Ca2+遊離に伴って低下すると,細胞膜上の Ca2+透過性の ストア作動性チャネル(store-operated channel; SOC)が活性化され,Ca2+が 流入する(Putney, 1990).近年,SOCE の活性化に関わるタンパク質として STIMとOraiが同定された(Liou et al., 2005; Prakriya et al., 2006).STIM は小胞体膜に存在し,[Ca2+]ERセンサーとして働く.[Ca2+]ERが低下するとオリ ゴマーを形成して,細胞膜に存在するCa2+透過性チャネルであるOraiと相互作 用してこれを活性化し,Ca2+流入を引き起こすと考えられている(Liou et al., 2007; Luik et al, 2008; Frischauf et al., 2008).
1.4.2 Ca2+遊離経路
細胞内 Ca2+ストアである小胞体膜に存在する Ca2+放出チャネルは,IP3受容 体(IP3R)/Ca2+チャネルとリアノジン受容体(RyR)/Ca2+チャネルの2種類に 分類される.IP3R/Ca2+チャネルはほぼ全ての細胞に存在すると考えられている
(Mikoshiba, 2007).細胞が外部から刺激を受けると,三量体Gタンパク質など
を介して活性化されたPLCが細胞膜脂質成分であるホスファチジルイノシトー ル二リン酸(PIP2)を加水分解し,細胞膜上にジアシルグリセロール(DAG), 細胞質中にIP3を産生する. IP3がIP3R/Ca2+チャネルに結合するとチャネルが 開口し,小胞体からのCa2+遊離が誘発される(IP3誘発性Ca2+遊離 IP3-induced Ca2+ release; IICR).IP3R/Ca2+チャネルの開口確率は,Ca2+に対して二相性の 依存性を示す(Bezprozvanny et al., 1991). 300 nM以下の低Ca2+濃度域で
は,Ca2+濃度の増加とともにチャネル開口確率が増加する.この性質により,
[Ca2+]cytのわずかな上昇が,IP3R/Ca2+チャネルを感作して更なるCa2+遊離を引 き起こすというポジティブ・フィードバックにより,急激に大きな[Ca2+]cyt上昇 が誘発され得る(Ca2+誘発性Ca2+遊離Ca2+-induced Ca2+ release; CICR).高 Ca2+濃度域ではIP3R/Ca2+チャネルは逆に濃度依存的に抑制されるため,一度上 昇した [Ca2+]cytは,IP3R/Ca2+チャネルへのネガティブ・フィードバックにより 急速に下降することになる
RyR/Ca2+チャネルは骨格筋細胞・心筋細胞や神経細胞などの小胞体に存在す
る.RyR/Ca2+チャネルも Ca2+によって感作される性質があるため(Fabiato, 1983),これらの細胞でも小さな[Ca2+]cyt上昇がトリガーとなってCICRが生じ 得る.心筋細胞では,T管の脱分極によりL型電位依存性Ca2+チャネルが開口 して細胞外から Ca2+が流入し,これが RyR/Ca2+チャネルを介した CICR を誘 発することで,大きな[Ca2+]cyt上昇がおこる(Bers, 2002).一方骨格筋細胞で は,T 管膜上の L 型 Ca2+チャネルの構造変化が筋小胞体膜上の RyR/Ca2+チャ ネルの構造に直接影響を及ぼすことが,Ca2+遊離のトリガーになると考えられ ている(Cheng et al., 1996).またある種の細胞では,環状 ADP リボース
(cADPR)が RyR/Ca2+チャネルに結合して Ca2+遊離を誘発するアゴニストと して働くことが示唆されている(Ogunbayo et al., 2011).
1.5 哺乳類受精卵における Ca2+動態
1.5.1 Ca2+波とCa2+振動
細胞内Ca2+の時間的・空間的動態は,蛍光性Ca2+プローブを用いた画像解析 法(Ca2+イメージング)を用いて,これまで様々な細胞について観察・解析さ れてきている.細胞の一部が局所的に刺激された場合,しばしば[Ca2+]cytはその 刺激部位から生じ,その後細胞全体へと伝搬する.この空間的Ca2+動態はCa2+
波(Ca2+ wave)と呼ばれる.細胞に対する刺激が持続する場合,[Ca2+]cytは増 加したままにはならずに一度減少し,その後周期的な増減を繰り返す.この時 間的Ca2+動態はCa2+振動(Ca2+ oscillations)と呼ばれる.Ca2+波は,局所的 な刺激によるシグナルを細胞内で速やかに伝達し,細胞全体での応答を起こす ための手段であると考えられる.一方 Ca2+振動は,過度の[Ca2+]cyt上昇によっ て Ca2+依存性プロテアーゼ等による自己破壊的な反応が生じることを避けつつ,
長時間にわたって Ca2+シグナルを維持するための応答様式であると解釈される
(Ozil, 1990).また細胞が受容した刺激の強度がCa2+振動の頻度としてコード されること,またCa2+振動の頻度に応じてCa2+依存性酵素の活性が調節される ことも示唆されている(De Koninck & Schulm, 1998; Dupont & Goldbeter, 1998).
哺乳類受精卵でも Ca2+波と Ca2+振動が観測される(Miyazaki et al., 1992;
Deguchi et al., 2000).図1.4はマウス卵の受精時の[Ca2+]cyt上昇の典型例であ るが,他の哺乳類の受精卵も基本的には同様のパターンを示す. [Ca2+]cyt上昇 は卵に精子が融合した部位付近から始まり,数~10秒間程度でCa2+波として反 対側まで伝播する(図1.4A).[Ca2+]cytはその後10~20秒の間さらに上昇する.
[Ca2+]cyt 上昇は数分持続した後,一度元のレベルに戻るが,以後,一過性の [Ca2+]cyt 上昇がほぼ一定の間隔で繰り返し生じる(図 1.4B).2回目以降の
[Ca2+]cyt上昇でも Ca2+波が観察されるが,1回目にくらべて伝播速度が速いこ と,また開始点が徐々に精子融合部位からずれて卵の植物極側半球へ移動する ことが知られている(Deguchi et al., 2000).体外受精で見られるCa2+振動の 周期は卵や精子の状態により大きく異なるが,典型的には10〜30分程度である.
なお,マウス卵での始めの[Ca2+]cyt上昇では,しばしば高濃度域で速い周期の Ca2+振動が観察されるが(図1.4C),以後本論文でいう「受精卵でのCa2+振動」
とは,遅い周期のほうの振動を指すものとする.
1.5.2 Ca2+遊離誘発機構
哺乳類受精卵における最初の[Ca2+]cyt上昇に至るメカニズムは,現在のところ 以下のように考えられている(Malcuit et al., 2006)(図1.5).受精時に精子と 卵の細胞膜が融合して互いの細胞質が通ずると,精子特異的に発現している PLCであるPLC(Saunders et al., 2002)が卵細胞質へと移行する.PLCは,
Gタンパク質やチロシンキナーゼによる活性化作用が無くても,静止時の低い [Ca2+]cytレベルで活性を示す(Kouchi et al., 2005).PLCによって卵内のPIP2
が加水分解され,細胞膜上にDAG と細胞質中に IP3が産出される.IP3は,小 胞体膜上のIP3R/Ca2+チャネルに結合して,Ca2+の放出を誘発する.放出された Ca2+によって IP3R/Ca2+チャネルが感作され,さらなる Ca2+遊離が生じて急峻 な [Ca2+]cyt上昇が引き起こされる.放出されたCa2+が拡散することで近傍の小 胞体上のIP3R/Ca2+チャネルも感作され,Ca2+遊離が誘発される.この過程の繰 り返しにより,[Ca2+]cyt上昇が卵内全体に伝播するCa2+波が形成されると考え
A
B C
図1.4 哺乳類受精卵でのCa2+波とCa2+振動
(A) マウス卵受精時の 1 回目の[Ca2+]cyt上昇の様子.共焦点顕微鏡で測定した
Calcium Green-1 の蛍光変化を擬似カラーで表した.卵の一番上の部分で精子
卵融合が起こっている.左下の数字は経過時間(秒).([Ca2+]cyt上昇が開始した 時刻を0秒とした)(Deguchi et al., 2000より改変)
(B) マウス卵受精時の Ca2+振動.(C) はじめの[Ca2+]cyt上昇.(B)の下線部を引 き伸ばした.
られている(Miyazaki et al., 1992).哺乳類卵にはRyR/Ca2+チャネルも発現し ていることが報告されている(Ayabe et al., 1995).しかしその数は少なく,ま た細胞表側部のみに局在しており,RyR/Ca2+チャネルを介したCa2+遊離は,受 精時の[Ca2+]cyt上昇には有意には寄与していないと考えられる.実際,ハムスタ ー卵を用いた実験では,リアノジンやカフェインが効果が無いことが報告され ている(Miyazaki et al., 1993).IP3R/Ca2+チャネルを特異的に抑制するモノク ローナル抗体によって,受精時のCa2+波もCa2+振動も完全に消失することから,
哺乳類卵での Ca2+遊離は IP3R/Ca2+チャネルのみによるものであると考えられ ている(Miyazaki et al., 1992; Miyazaki et al., 1993).
1.5.3 Ca2+流入経路
マウス受精卵では細胞外からの Ca2+流入も活性化されていることが,Mn2+
消光法(第3章参照)を用いた実験により示唆されている(McGuinness et al.,
1996).また,精子抽出物を卵に注入することによって誘発した Ca2+振動の際
も,Ca2+流入が促進される(Mohri et al., 2001).マウス卵の受精後に外液の Ca2+を除去すると,Ca2+振動は速やかに停止する(Kline & Kline, 1992).また,
ハムスター受精卵のCa2+振動の頻度は,細胞外のCa2+濃度に対して正の依存性 を示す(Igusa et & Miyazaki, 1983).これらの結果は,哺乳類受精卵でのCa2+
振動には Ca2+流入が必須であること,また Ca2+振動の頻度は Ca2+流入速度に 依存することを意味している.McGuinnessらは,Ca2+流入は細胞内Ca2+スト アの再充填に寄与しているという仮説を提唱した(McGuinness et al., 1996).
全てをストアに回収することはできない.再び急激な Ca2+遊離が生じるには,
ストア内のCa2+濃度があるレベルまで回復する必要があり,細胞外からのCa2+
流入はその補充のためのCa2+を供給していると考える(図1.5).
ある細胞に SOCE が存在するかどうかは,タンプシガルジンなどの小胞体 Ca2+ポンプの阻害剤で Ca2+ストアを人為的に枯渇させたときに Ca2+流入が促 進されるか否かで判断されることが多い.マウス卵細胞でも,タプシガルギン 処理によってCa2+流入が増加する(Kline & Kline, 1992; McGuinness et al., 1996). さ ら に 近 年 ,STIM1 と Orai1 が 発 現 し て い る こ と も 確 認 さ れ
(Gómez-Fernández et al., 2009),マウス卵にもSOCE経路が存在すると考え られている.しかし一方で,タプシガルギン処理だけでなく IP3によって Ca2+
遊離を誘発しないとCa2+流入が有意に増加しないという報告もあり(Mohri et al., 2001),マウス卵でのSOCEが活性化するにはCa2+ストア内のCa2+濃度の 大幅な減少が必要である可能性もある.マウス卵にSOCEの経路が備わってい るとしても,それが受精時の Ca2+振動の際に活性化されるか,また活性化され るとしても Ca2+振動の維持に有意に寄与するかという点については検証が必要 である.
1.6 本研究の目的
本研究では,哺乳類受精卵に見られる Ca2+振動の発生メカニズムの解明を目 指した研究の一環として,Ca2+振動に対するCa2+流入の機能的関与の検証と,
Ca2+振動の維持に関わるCa2+流入経路の同定と制御機構の解明を目的として,
PLC
PLC
PIP2 DAG
IP3 IP3 IP3受容体
Ca2+
小胞体 精子
Ca2+
Ca2+
Ca2+
Ca2+流入チャネル
Ca2+ポンプ 卵細胞膜
+
+ ++
Ca2+ポンプ
--
図1.5 哺乳類受精卵での細胞内Ca2+動態の制御機構
以下の研究課題を設定した.
研究課題1 受精卵Ca2+振動のCa2+流入依存性(第2章)
マウス受精卵の Ca2+振動について,細胞外 Ca2+濃度を変えて Ca2+流入速度 を変化させた際の頻度および振幅に対する影響を定量的に評価することで, こ れまでに報告されているCa2+振動のCa2+流入依存性について再検証する.
研究課題2 受精卵におけるCa2+流入の活性化(第3章)
マウス受精卵の Ca2+振動中の Ca2+流入速度を,Mn2+消光法を用いて測定す る.特にCa2+遊離/Ca2+取り込みのサイクルとCa2+流入速度の時間的変動との 関連に注目して解析する.
研究課題3 細胞内Ca2+ストア再充填に対するCa2+流入の影響(第4章)
小胞体内腔にターゲットしたタンパク質性 Ca2+プローブを用いてマウス卵受 精時の[Ca2+]ERの変化を可視化し,細胞内 Ca2+ストアの枯渇~再充填の過程を 測定する.さらにCa2+ストアの再充填速度のCa2+流入速度に対する依存性につ いて検証する.
研究課題4 受精卵Ca2+振動におけるSTIM/Orai系の関与(第5章)
SOC に対する阻害剤を用いて,マウス受精卵の Ca2+振動における SOCE の 寄与について検証する.また,STIM1およびOrai1の細胞質ドメインフラグメ ントの過剰発現の影響を調べることで,STIM/Orai系を介したSOCEの機能的 関与について明らかにする.
第2章 受精卵 Ca
2+振動の Ca
2+流入依存性
2.1 目的
哺乳類受精卵では,精子によって[Ca2+]cytの上昇が誘発される.卵内の[Ca2+]cyt
は数時間 にわた り 一 過 性の[Ca2+]cyt 上昇を 周期的 に繰り 返す (Ca2+振動)
(Miyazaki et al., 1992; Deguchi et al., 2000). [Ca2+]cyt上昇により,減数分 裂中期で停止していた卵の細胞周期が再開される(Hyslop et al., 2004).同時 に,複数の精子との受精を防ぐ応答(多精拒否)も[Ca2+]cyt上昇によって引き起 こされる(Liang et al., 2011).
哺乳類受精卵のCa2+振動での個々の[Ca2+]cyt上昇は,細胞内Ca2+ストアであ る小胞体からのCa2+遊離により起こる(Miyazaki et al., 1991).受精時のCa2+
振動は精子に特異的に発現する PLCの卵細胞への移行により開始されると考 えられている(Saunders et al., 2002).PLCは卵内のPIP2を加水分解して細 胞質中にIP3を産生する.このIP3が小胞体膜上のIP3R/Ca2+チャネルを活性化 することによって小胞体内のCa2+を遊離させる.
一方で,受精時のCa2+振動には細胞外からのCa2+流入も必要であることが知 られている.Igusaらは,ハムスター受精卵で見られる膜電位の周期的な過分極 応答の[Ca2+]oに対する依存性を解析した(Igusa & Miyazaki, 1983).ハムスタ ー卵にはCa2+依存性K+チャネルが存在するので,Ca2+振動の周期は過分極応答 の周期として測定できる.過分極応答の周期は,[Ca2+]oを増加すると短くなり,
[Ca2+]oを低下または除去すると長くなったことから,細胞外からのCa2+流入は
Ca2+振動の維持のために必要であり,またCa2+流入速度が振動頻度に影響する ことが示された.Kline らは,fura-2 による Ca2+イメージングによる測定で,
マウス受精卵での Ca2+振動中に外液の Ca2+を除去すると Ca2+振動が停止する ことを報告しているが(Kline & Kline, 1992),Ca2+振動の頻度や振幅に対する [Ca2+]oの影響について定量的な解析は行っていない.
Ca2+振動の発生・維持におけるCa2+流入の役割を明らかにするためには,先 ずは Ca2+流入が Ca2+振動パターンに与える影響を精査することが重要である.
本研究課題では,マウス受精卵でのCa2+振動をfura-2を用いて経時的に測定し,
そのCa2+流入速度に対する依存性を定量的に評価することとした.実験的には,
Ca2+振動の途中で[Ca2+]oを変えることによりCa2+流入速度を変化させて,その 前後のCa2+振動の頻度および振幅の変化について解析した.
2.2 方法
2.2.1 成熟卵および精子の採取
本実験では,ddY マウス(日本 SLC)のメスマウス(6 週齢以上)とオスマ ウスを用いた.すべての実験は,電気通信大学動物実験指針に従って行った.
成熟卵の採取のために,メスマウスに以下の過排卵処理をした.血清性性腺刺 激ホルモン(ピーメックス,三共エール薬品,5 U)を腹腔内注射し,その 18 時間後にヒト絨毛性性腺刺激ホルモン(ゴナドトロピン,三共エール薬品, 5 U)
を腹腔内注射した.ゴナドトロピン注射の16時間後に卵管から成熟卵を取り出
した.採取した卵は常にM2培地(NaCl 94.6 mM, KCl 4.8 mM, CaCl2 1.7 mM, KH2PO4 1.2 mM, MgSO4 1.2 mM, NaHCO3 5.1 mM, HEPES ( 4-
(2-hydroxyethyl)-1-piperazineethanesulfonic acid) 10 mM, Na lactate 19.6 mM, Na pyruvate 0.5 mM, Glucose 5.6 mM,BSA 4 mg/ml, pH 7.2)の中で 扱った.実験で用いた他の薬品はM2培地に溶解して用いた.卵丘細胞は0.05%
ヒアルロニダーゼによって分解,除去した.
精子はオスマウスの精巣上体尾から採取し,M16培地(NaCl 94.6 mM,KCl 4.8 mM,CaCl2 1.7 mM,KH2PO4 1.2 mM,MgSO4 1.2 mM,NaHCO3 25 mM, Na lactate 17.8 mM,Na pyruvate 0.33 mM,Glucose 5.6 mM, BSA 4mg/ml)
のドロップ(200 L)の中に入れ,5%CO2,37°C のインキュベーター内で培 養した.1時間後,M16ドロップの上方を泳いでいる精子を5 Lまたは10 L を採取し,新たなM16培地のドロップ(200 L)に加えた.以後18 時間以上 培養することで受精能を獲得させた.
2.2.2 Ca2+イメージング
fura-2 AM(同仁)はDMSO(和光)で4 mMに溶解したものをストック溶
液とした.使用時には,これに細胞膜内へのプローブの浸透を高める導入補助 薬であるpluronic F-127(Molecular Probes)(0.05%)を等量混和し,fura-2 AM の最終濃度が2 MになるようにM2培地で希釈した.採取した成熟卵を2M fura-2 AMで37°C,10分間染色し,その後酸性タイロード溶液(NaCl 173 mM,
KCl 7.0 mM,CaCl2 1.6 mM,MgSO4 0.5 mM,glucose 27 mM,HCl 1.6 mM,
PVP-40 0.5 mM,pH 2.5)で約1分処理することで透明帯を除去した.
fura-2の蛍光イメージングには従来型の落射型蛍光顕微鏡(Axiovert S-100, カールツァイス)を用いた.ガラスボトムディッシュに200 µLのM2培地のド ロップを作り,蒸発を防ぐために流動パラフィンで覆った上で,顕微鏡ステー
ジ上で32-34°Cに加温した.透明帯を除去した卵をそのM2ドロップ内におき,
5分程度静置して卵がカバーグラス面に十分付着するのを待った後,前培養した 精子を少量くわえて媒精した.蛍光イメージングには20倍対物レンズ(FLUAR,
NA0.75,カールツァイス)を用い,励起光源として安定化 Xeランプ(75 W,
浜松ホトニクス)を用いた.340 ± 5 nmと380 ± 5 nmのバンドパスフィルタ ー(朝日分光)をフィルター切り替え装置(Labmda10-2,サッターインストル メント)を用いて切り替えることで,340 nmと380 nmの2波長で交互に励起 した.生じた蛍光は,535 ± 12 nmのバンドパスフィルター(オメガオプテイ カル)を通した後,ICCDカメラ(ICCD-350F, ビデオスコープ)によって検出 し,ビデオ信号をフレームキャプチャボード(LG-3, サイオン)により 8 ビッ トでA/D変換してパソコン(PowerMac G3, アップル)に取り込んだ.蛍光 画像は典型的には20 秒間隔で取得した.それぞれの励起波長での蛍光画像をも とに,個々の卵の蛍光強度(F340, F380)を測定し,バックグラウンド減算をし た後,蛍光強度比(F340/F380)を算出して, [Ca2+]cyt の指標とした.なお,フ ィルター切り替え装置の制御,蛍光画像の取得およびその解析は,すべてフリ ーソフトウェアの NIH Image をもとに作成した独自のプログラムによって行 った.
受精によるCa2+振動が安定した後,適当なCaCl2濃度のM2培地をステージ
上のM2ドロップ(1.7 mM CaCl2)に加えることで,[Ca2+]oを変化させた.例 えば,[Ca2+]oを 1/2 にする際には,CaCl2を含まない M2 培地をドロップと等 量(200 µL)加えた.[Ca2+]oを0 にする際には,CaCl2を含まずかつ10 mM EGTAを加えたM2培地を等量(200 µL)加えた.
2.3 結果
マウス受精卵のCa2+振動中に,外液のCa2+濃度を0〜2倍に変化させたとこ ろ,Ca2+振動の頻度の顕著な変化が観察された(図2.1). [Ca2+]oを2倍(3.4 mM)に増加させると頻度が増加したのに対し(図2.1 A), 1/2倍(0.85 mM), 1/4倍(0.43 mM)に減少させると,濃度依存的に頻度が減少した(図2.1B). 一方で Ca2+振動の振幅(個々の[Ca2+]cyt上昇の大きさ)については,いずれの 場合においても[Ca2+]o変更の前後で明確な差異は観察されなかった.Ca2+キレ ート剤であるEGTAを5 mM投与して外液中のCa2+をほぼ完全に除去すると,
直後にCa2+振動は停止することが確認された(図2.1C).
受精卵での Ca2+振動の頻度や振幅は,実験ごとに卵と精子双方の状態に依存 して大きく異なるため, [Ca2+]oの影響を定量的に評価するのにそれらの絶対値 を用いることはできなかった.そこで本解析では,[Ca2+]o 変更の前後1時間に 生じた[Ca2+]cyt上昇の回数を測り,その回数の比(後/前)を計算して,Ca2+振 動の頻度に対する[Ca2+]oの効果の大きさの指標とした(図2.2).同様に振幅に ついても,[Ca2+]o変更の前後1時間に生じた[Ca2+]cyt上昇の大きさ(F340/F380
A
B
C
図2.1 細胞外Ca2+濃度変化の受精時Ca2+振動に対する影響
マウス卵の受精により誘発された Ca2+振動中に[Ca2+]o を変化させたときの Ca2+振動パターンの変化.[Ca2+]oを1.7 mMから3.4 mM(A),0.43 mM(B), 0 mM(5 mM EGTA)(C)に変化させた.
図2.2 Ca2+振動の頻度・振幅の変化の解析方法
[Ca2+]oを変更した前後それぞれ一定時間に生じた Ca2+上昇の回数を測り,そ れらの比 (変更後 /変更前)を計算した.振幅も同様に一定時間に生じたCa2+
上昇の大きさの平均値の比を計算した.



![図 3.4 Ca 2+ 流入速度と[Ca 2+ ] cyt 上昇の関係](https://thumb-ap.123doks.com/thumbv2/123deta/7733542.1711695/49.892.217.680.313.769/図34Ca2+流入速度とCa2+cyt上昇の関係.webp)

![図 4.4 細胞外 Ca 2+ 濃度変更にともなう D1ER と fura-2 のシグナル変化 マウス受精卵の Ca 2+ 振動中に[Ca 2+ ] o を増減したときの[Ca 2+ ] ER と[Ca 2+ ] cyt の;](https://thumb-ap.123doks.com/thumbv2/123deta/7733542.1711695/64.892.179.710.154.964/細胞外+濃度変更ともなうシグナル変化マウス受精+振動中Ca.webp)