海水養殖魚の品種改良に関する研究
144
0
0
全文
(2) 博士学位論文. 海水養殖魚の品種改良に関する研究. 平 成 9年 9 月. 近畿大学水産研究所 (主査:熊井英水教授). 村 田. 修.
(3) 海水養殖魚、の品種改良に関する研究 近畿大学水産研究所 村田修 (主査:熊井英水教授). S t u d i e sont h eB r e e d i n go fC u l t i v a t e dMarineF i s h e s. OsamuMurata G r a d u a t eS c h o o lK i n k iU n i v e r s i t y D i v i s i o nofA g r i c u l t u r a lS c i e n c e ( A d v i s o r :P r o f .HidemiKumai) S e p t1 9 9 7 ラ. ラ. i n k iU n i v e r s i t yt of u l l f i l lt h er e q u i r e m e n t s S u b m i t t e dt ot h eG r a d u a t eS c h o o lK f o rt h eD o c t o r a t eD e g r e e . ラ.
(4) 海水養殖魚の品種改良に関する研究 S t u d i e sont h eB r e e d i n go fCu 1 t i v a t e dM a r i n eF i s h e s. 村田修. 目次. 第 I章 第 E章. 農畜産物および養殖魚における品種改良の意義と歴史. iqJnhU. 市. 緒論. 選抜による品種改良. I- 1.選抜育種マダイの成長. II-1-3. 考 察 I- 2 . 選抜育種マダイのアイソザイムによる集団解析. 司. I- 1- 2 . 結果. 30234916 11111122. II-l-1.材料および方法. II-2-1.材料および方法 I- 2- 2 . 結果 II-2-3. 考察 第皿章. 人工交雑による品種改良. 国一l.交雑魚の成長. 外部形態および環境ストレス耐性. m-l-1 . マダイ×クロダイ. 皿 -1-1-2 . 結果および考察. m-1-2 . マダイ×ヘダイ. 円 。。。 つ u つω つ山内δ. 国一 1-1-1.材料および方法.
(5) III-1-2-2. 結果および、考察 ill-1-3. イシダイ×イシガキダイ III-1-3-1.材料および方法 I I- 1- 3- 2 . 結果 III-1-3-3. 考察 国 -1-4 .. プリ×ヒラマサ. 皿-1-4-1.材料および方法 皿-1-4-2. 結果および考察 皿 -1-5 . カンノ tチ×ヒラマサ. III-1-5-1.材料および方法. 351129223999344489991 3344445555556666666677. III-1-2-1.材料および方法. I I- 1- 5- 2 . 結果および考察 皿 -2 . 交雑魚の生殖腺成熟. III-2-1 . マダイ×クロダイおよびマダイ×ヘダイ 国一. 2-1-1 . 材料および方法. III-2-1-2. 結果 皿-2-1-3. 考察. III-2-2.. イシダイ×イシガキダイ. III-2-2-1 . 材料および、方法. 皿-2-2-2.. 結果および考察. 1. ill-2-3-1 . 材料および方法. ょ. III-2-3. プリ×ヒラマサおよびカンパチ×ヒラマサ.
(6) ヴ i. OO. III-3-1.材料および方法. ヴ t. III-3. 交雑魚のアイソザイムおよび核型分析. 1i. 結果および考察. 777788. 田 -2-3-2.. III-3-2. 結果 ヴ t. 00. 皿 -3-3 . 考察 第町章導入による品種改良. N-l. 日本マダイと香港マダイとの成熟・産卵期の比較. N -2-2 . 結果および考察. つU A T Q U. 町一 2-l.材料および方法. つd. N-2. 日本マダイ×香港マダイ. Quod. N -1- 2 . 結果および考察. 889999. N - l -l.材料および方法. 第 V章 染 色 体 操 作 に よ る 品 種 改 良 V -l.マダイの三倍体. V -1- l.材料および方法. 98. V -1- 2 . 結果. 99. V-1-3. 考察. 107. V-2. イシダイの三倍体. 要約. 108. V -2-l.材料および方法. 108. V -2-2 . 結果. 109. V-2-3. 考察. 118 120.
(7) Summary. 123. 謝辞. 127. 文献. 128.
(8) 緒論 国連食糧農業機関 (FAO) の世界漁獲統計 (1994)によると,世界の総漁. 958万トンに達し過去の生産量をさらに更新した。しかし, FAO 業生産量は 10, の予測によれば世界の総漁獲量は水産資源の状況などから今後 11, 000万トンが 限界のようである. O. 一方. 日本の総漁獲量はかつては1.000万トン台を続け世. 界第一位であったが, 1989年に首位の座を中国に譲って以来年々減少し,今 日ではチリに次いで第四位である. O. 今後も増加が見込めないことから,これか. らの日本の漁業は水産増養殖に期待するところが大きい。 わが国の海面養殖漁業の生産量は年々増加し, 1994年には 130万トン,総漁 業生産量の 16%,生産額で 6, 300億円,総生産額の 26.4%を占めてこの 20年間 で約 2倍となり,海水養殖業は遠洋漁業を上回るまでに成長した。その主な原 因は,資源培養型漁業に基づく海水魚類養殖の発展によるもので, 1928年に プリ養殖に着手した野網和三郎によって開発された。その後, 1954年に原田 輝雄によって網いけす式養殖法が考案され,従来の池中養殖から養殖場を内湾 海面に設置することができるようになって当時停滞していたプリ養殖が急速に 進展した. O. そして,マダイ. カンパチ,ヒラマサ,シマアジさらにはクロマグ. ロなどについても天然幼稚魚を飼い慣らして家魚化することが可能となった。. 1965年頃からのわが国の高度経済成長に伴う海産魚の. 海水養殖業の発展は. 需要増大が一要因となっている。 なお,淡水養殖は 1, 000年以上の歴史があるのに比べ,海水養殖の産業とし ての歴史はまだわずか 30年余りでしかない。そして海水魚養殖の発展初期には 天然種苗が用いられたため年による豊凶の差が大きくてその入手が不安定で あった。そこで必然的に養殖対象魚の種百生産技術の開発が切望され,近年は 長足の進歩を遂げつつある. O. さらに,野生動植物の飼育栽培においては,それが人類にとって望ましい産 業品種となるように多年に亘って改良することで,農畜産物の育種には多くの 成果が得られている. O. しかし,魚類における品種改良の歴史は栽培植物および. 家畜に比べまだ著しく短い。しかし,水産養殖業の将来の一層の進展を計るた めには,養殖対象魚の品種改良によって,成長,外観,肉質,環境ストレスお よび病気に対する耐性など経済的特質の向上を期待する必要がある. O. 以上の観点から,本研究では海水養殖対象魚として重要なブリ属,タイおよ びイシダイ科魚類、を中心に,選抜育種,交雑育種,導入育種および染色体操作 による品種改良を行い. 作出魚の養殖効果を検討した。.
(9) 第 I章では,農畜産物および養殖魚における品種改良の意義と歴史について 述べた。 第 E章では,成長の早い養殖マダイ種苗を生産することを目的として,選抜. 5年以上に宣って試み,その聞の選抜効果を調べるとともに選抜マダイ 育種を 2 のアイソザイムによる集団解析を試みた。 第皿章では,マダイ×クロダイ,マダイ×ヘダイ. イシダイ×イシガキダ. イ,プリ×ヒラマサおよびカンパチ×ヒラマサ雑種第一代の特性を明らかにす る目的で,これらの交雑魚の成長,生残率,外部形態および環境ストレス耐性 をそれぞれの親魚種と比較した。また,各交雑魚の生殖腺の発達状態を調べ, さらにアイソザイムおよび、染色体の分析を行った。 第 W章では,香港近海で漁獲されたマダイと日本の養殖マダイとの成熟およ び産卵時期を比較するとともに,それらの正逆両交配を行い成長および形態を 比較した。 第 V章では,染色体操作によって三倍体マダイおよび三倍体イシダイを作出 し,成長および体成分組成に及ぼす不妊化の影響などについて調べた。. 2.
(10) 第 I章. 農畜産物および養殖魚における品種改良の意義と歴史. 野生の動物や植物を人工的に飼育栽培しその一生を管理することが可能にな ると,次にその動植物の遺伝質を改善し人類の希望する有用な新しい品種に改 良しようと試みるがこれが品種改良である. O. 農業では農作物の栽培を始めた大昔の頃から品種改良が続けられ,耐寒,耐 暑,耐病性などのストレスに耐えるものや環境適応性の異なる品種などを選択 して栽培しやすくし,一定の土地からより多くの生産物を得ること,すなわち 多収性作物への改良が行われてきた. O. このようにして. イネやコムギでは. 1950年代から,半嬢性品種の育成が中国,韓国,アメリカ合衆国など各国で 開始されるようになり,種間の交雑やハイブリッドF1の利用によって多収安定 化に富んだ新品種が誕生した。また,近年ではバイオテクノロジーの急速な進 展によヮて細胞培養や DNA組換えによる新植物の作出に関する研究が盛んに行 われている o. 1 ). 畜産業では野生動物を飼い慣らして家畜化し. それらの動物のもっている性. 質や能力を引き出し,経済性の優れた家畜の共通要素である繁殖性,生残率, 飼育しやすさ,飼料利用性および病気に対する自然抵抗性などの諸能力を備え た上で,量的,質的に優れた乳,肉,卵,毛皮などの生産性の向上を目的とし た品種改良が行われてきた。. 2 ). 家畜では交雑育種が主であり,例えば,肉用牛. の黒毛和牛は日本の在来種にブラウン・スイス. デボン. ショートホーンなど. が人工交配され改良されてできたものであり,交配の種類には属間,種間,品 種間における遠縁交配や,純粋,系統,近親交配による近縁交配がある. O. さら. ) に,最近は, F1雑種強勢が多く利用されるようになってきた。 3. 以上のように,農業,畜産業の分野における品種改良は次に述べる水産業の 分野に比べて格段に長く古い歴史をもって推移してきたことは周知の通りであ. 剣く る 水産業の中でも魚類養殖は,コイ,キンギョ,ニジマスなどの一部の j O. 魚を除くと,養魚の歴史が浅く,対象魚には野生種がそのまま養殖されている 場合が多い。魚類養殖の現況をみると,養魚環境の悪化による成長速度の鈍化 や魚病の発生,人件費や飼料コストの高騰などによる収益低下などの問題を生 じ,その発展には種々の対策が望まれる. O. 野生種を改良し,成長,形状(姿) ,色調,肉質,繁殖力,飼料効率,耐病 性,環境ストレス耐性など種々の形質において優れた品種を作出することはそ の打開策として極めて重要である. O. 本研究を進めるにあたり,これまでに取ら. 3.
(11) れてきた魚類育種の概要を述べると次の通りである. O. ( 1)選抜育種:魚類の選抜育種による品種改良のほとんどは,コイ. ωr p i o , 4 ) ニジマス. , 8 ) t s h a w y t s c h a. Oncorhynchus m y k i s s , 57). ギンザケ O n c o r h y n c h u s k i s u t c h, 9 ). マスノスケ. C y p r i n u s. Oncorhynchus. アメリカナマズ. Jc l t αl u r u s. p u n c t α ,t u s , 10-12) ナイルテラビア O r e o c h r o m i sn i l o t i c u s, 13,14 ) カダヤシ G ambusi α 6) q f f i n i s15)などの淡水魚で試みられている。 H o r s t g e n S chwark は. " p a n s i z e "ニジ. マスの成長に関する選択実験において平均実現遺伝率は 0 . 1 5であったと報告し ている o H e r sh b e r g e rら9)によれば, 10年間選抜した大西洋サケでは,奇数年お. . 3 2および よび偶数年に成熟する親魚の成長に対する遺伝率はそれぞれ1.22士 0 0 . 8 1+0.30であヮた o B o n d a r ilO)は,体重に対するアメリカナマズの実現遺伝率 は0 . 1 0で雄親あるいは雌親の分散より求めた遺伝率よりも低かったと報告して いる. O. 遺伝率はその値が高いほど選抜の効果が早く得られて効率がよく, 0 . 2. 以上あればそれを推定した集団のその形質の選抜効果があるといわれている. O. 1 6 )しかし,海水魚の選抜育種に関する研究はまだほとんどみられない。 1 7 ). ( 2 )交雑育種:育種の一つに選抜された系統を交雑して雑種強勢をはかり,. その中から優秀なものを再び選抜して回定化を行う方法がある. O. また,種間交. 雑や属間交雑を行い,これまでにない種を作り出すことが試みられているが, 和田 16)は育種学的には同一種内の異なる系統を交雑する方法が,形質の固定や 品種の育成には長い目でみれば有利であろうと述べている. O. コイ科魚類の交雑. として古くから試みられているキンギョは,フナの交雑種であって多くの品種 がある. O. シュプンキン,キヤリコ,キンランシなどはこの交雑によって得られ. たものである o. 18, 1 9 )また,コイとフナとの雑種の成長はコイよりも劣るがフ. ナよりも良く両親の中間的であることが知られている 0 1 8,. 19)サケ科魚類で. は,寺尾 20)は,サケとヒメマスの正逆交雑Flについて,広食性,棲息環境条件 の雌雄ともに成熟したので、 F の幅広い抵抗性に優れ, F1 1同志の交雑と戻し交雑 を行った結果,交雑魚 F2の卵径は不同で、あり,発生・発育能力が低く,発生速 度の変異の幅も著しく大きかったことを報告している. O. なお,サケ科魚類の交. 雑については内外を問わず多くの研究がなされているがここでは省略する. O. そ. の他の淡水魚類について鈴木 21),皆森 22, 23)は,シマドジョウ×マドジョウの 交雑魚は発生が正常で雑種強勢となったがその逆の組み合わせは雑種致死,崩 壊がみられたと報告している. O. 海水魚類としては,藤田 24)が について報告している. O. また. トラフグ属数種の人工交配と数種幼生の飼育. 荒川および吉田 25)は,マダイ X チダイの交雑魚. 4.
(12) は母系であるマダイに近い測定形質を示したが,体高,頭長,眼寓長,吻長お よび上顎長はマダイより短く,体色はマダイに比べて黒色が薄く鮮やかな赤色 を呈したと報告している. O. このように,海水養殖魚に関する報告は淡水魚類の. それに比べてまだ著しく少ない。. ( 3 ) 導入育種:日本のキンギョは 1 502年に中国から渡来した。 1895年には吏に中 国からリュウキン,デメキン,そして,戦後米国よりコメットが, 1961年に は中国キンギョと称される数種のキンギョが移入され,. 26.27). 日本産キンギョ. 種類が知ら の多くはこれらから淘汰あるいは交雑により作出されたもので約 20 れている o. 28). ニジマスは 1877年に米国カリフォルニア州から発眼卵が初めて. 日本に移植された。 29.30) コイは 1 904年にドイツから体長約 6cmのカワゴイ雌. 4尾とカガミゴイ雄 1尾が移植され 1906年にカワゴイに日本ゴイの雄を交配 したものが飼育された。. 3 1 ). これら以外にもソウギョ,レンギョ,ベヘレイな. ど多くの淡水魚が外国より日本に移植された。 2 ) 一方,海水養殖魚においても東南アジア産ハタ・クエ類,中国産スズキ, 3. 韓国産マダイ,香港産マダイなどの移植が試みられているが,それらの研究報 告はまだほとんどみられない。 ( 4 )染色体操作:染色体操作による三倍体は不妊による成長の促進や寿命の. 延長を期待して,淡水魚ではサケ科魚類およびアユ,海水魚ではマダイ,イシ ダイ,ヒラメなどに関する若干の報告があるが,養殖魚として全雌三倍体,異 質三倍体などを作出して実用化されているのはアユ,サクラマス,アマゴ,ヤ マメなどに過ぎない。 3335) 海水魚としては,ヒラメは雌の方が雄より成長が早い 36)ので,田畑 37)および 山本38)らが雌性発生ヒラメの生産について報告しており,実用化に向かってい る唯一の魚種である. O. しかし. ヒラメ以外の海水魚の倍数体の実用化に関する. 研究報告はまだほとんどみられない。. 5.
(13) 第 E章 選 抜 に よ る 品 種 改 良. n-1.. 選 抜 育 種 マ ダ イ の 成 長 39). マタ明イ P αr g r u sm a j o rの本格的な養殖は, 1965年頃から天然種苗を用いて行わ れるようになヮた。しかし. マダイの漁獲量が減少しはじめ,資源管理面 4042). から人工種苗の生産が必要となった。山下43)による天然親魚からの採卵,稚魚 までの育成,原因 44)による養成親魚からの採卵の成功などを通じて,マダイの 大量種苗生産技術 45)の途が聞かれたが,マダイの養殖生産量の増加に伴って, 養殖マダイの市場価格が低下し,現在では,商業生産に適した形質を持った種 苗を作出する必要が生じてきた。 著者共は,マダイにおいて天然種苗よりも成長の速い人工種苗を大量生産す ることを目的として, 1964年以来,成長を主な指標とした選抜育種を試み, 46) 現 在,. 8 世代目. 飼育中である. O. (1993~1995 年産,以下,. その結果. GIV ( I V世代目. GV I I Iと略記)の選抜マダイを. 1975~1977 年産)頃から世代を. 重ねるに従って,親魚の体重が増加するとともにより成長の速い種苗が得られ るようになった。以下にその研究方法と結果について述べる. O. II-l ー 1. 材 料 お よ び 方 法. ( 1 ) 親魚の養成と選抜 初代親魚 (1964年)から GV I I.V I I I親魚 (1989~1991 年産)に至るまでの 選抜育種の過程,ならびに各世代種苗の生産に用いられた親魚数およびその雌 i g .I I 1に示した。本研究所選抜マダイの創始集団は,主に 1964年前後 雄比を F. に和歌山県白浜沿岸で漁獲された天然幼魚を飼いつけ. 親魚として養成したも. のである o 1963年には約 200尾 , 1964年には約 500尾が導入され,それらは導 入時期の違いから 3集団に分類された o 1968年から 1970年にかけてそれらの 個体群より採卵,人工癖化飼育した GIの個体群より,成長,体色および体形 の優れた個体を選抜し次世代の親魚とした。 GIの生産に用いた雌雄親魚の数 は不明である. O. 人工鮮化から親魚に至るまで、の基本的な選抜方法を F i g .I I 2に. 示した。陸上水槽において全長 20mm前後になヮた稚魚を海面網いけすに沖出 しして約 3週間飼育し. 全長40mm前後になった頃に第 l回目の選別を行い,. その中の大型個体が全長 70mm前後の種苗サイズになったところで,第 2回目 の選別を行い大型魚のみを養成した. O. 単年度の飼育尾数は年次によりやや異な. 6.
(14) 礼. τ. 品 r. 。 品 司 令. 品事. AF也市暗唱 田平伊. J. 。、 h. 小。川ノ. 、 ,. ﹀し円. F i g .1 1 -1 .T h eh i s t o r yo fs e l e c t i v eb r e e d i n gi nr e ds e ab r e a m . N u m e r a l so na r r o w sshown u m b e ro f b r o o d s t o c ku s e df o rp r o d u c t i o no f n e x tg a n e r a t i o n .Romann u m e r a l s i np a r e n t h e s e ss h o wg e n e r a t i o no ft h ep o p u l a t i o n s .. 付制. 干れ禅述肉ゆ翫司抑﹁同. ﹁ぺ. 抑琳国抑制嚇 hF. τ. o難. δ卸業協剛淫ゆ命。 UOμ♂ 津 川 間 分 灘 品 一 園 田. 、 寸. 汁温い津市一隅!庁決議 (N0185 臼)打鵡﹁パゆ回一浮附若干れし汁 σ州議匂宮内論汁. H. w﹁ぺ鈴森川向 uo か MOJOM 同調部隊d J VO汁。き Nh 相理蝋h J ニ片町長ーピハ同﹁汁事、舟A 鴻 ωE回︿守備浬州訪 汁腿d wm ・骨 4 険3爾 斗 ﹁ 汁 菌 薪 時 嗣 鉢 了 世 述 通 費 ヰ斗 3隔持前津一件 ω G o g o嗣 H﹁汁 o μ Q J↑付申神州司油 打 完 胡 ﹁ 汁 。 中δ. ♀ 25♂ 15.
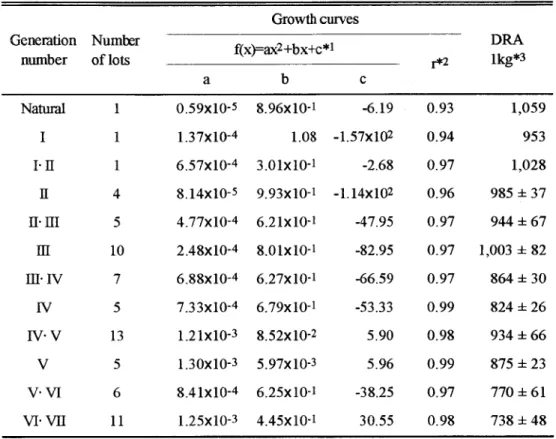
(15) 魚水槽への収容尾数は産卵水槽の容量によって異なるが 101 5 0尾程度とし た 。 e 、 出 Q J v. y. un. Jo. S. Hu. U. ・. Gd. h. obwu. 均台. J u v e n i l e s ( n e t c a g e s ). J u v e n i l e s ( n e t c a g e s ). yno. 也 m 山 ∞内問. Lm. AU. ぅ“. n u. n u. bρu m u N. 1 s t s e l e c t i o n. 一一件肱. , に. 相 Th. T . L . :~20mm. N u m b e r :800 ∞ ,o. ∞. A g e :50~7 0d a y s T .L . :20 4 0 mm. Ag e :0~50 d a y s. 2nd s e l e c t i o n. s. 的町. 4. ∞. 班. m d. 4J. EA 咽. 3 町 eL山 σむ 司 A 刷. 、Z. mm. 町. LU. yo r b -. L. 陥. Fm. A. Ag e :O~2y e a r s 500 N u m b e r :12,. e d + ・ ・ ‘ EA、‘,ノ. 出 、. uco. 日. A. dp. 4. c. 、 ‘. 〆'E. A d u l t s ( n e t c a g e ). 03 、 d ‘ ・ 川市町. Broodstock ( t a n konl a n d ). uvi o Y. 3rd s e l e c t i o n. s e l e c t i o n. F i g . I I ・2 .A句r p i c a lc y c l eo fs e l e c t i o np r o c e d u r eo fr e ds e ab r e a m . ( 2 ) 選 抜 育 種 マ ダ イ 4歳親魚の体重測定. 選抜世代の経緯とその選抜方法については Figs. II-1 および~I-2 に示したとお りであるが,これらの各世代親魚から生産された稚魚は次世代の親魚を選抜す るために 6歳魚まで飼育した. O. しかし. 種苗生産に供した親魚の多くは 4歳か. ら 5歳であった。そこで,選抜世代回数と親魚体重との関係を知るために,各 選抜世代の 4歳親魚群から採卵に供するために最終選抜した親魚 10尾の体重を 各世代とそのロット毎に測定し比較した。データは,各世代毎の 1 8グルー プの平均値として示した。 ( 3 ) 選抜育種マダイの癖化後の体重測定. 選抜育種マダイの世代数とそれぞれの解化後の成長との関係を知るために, 生産ロットが少なかった天然魚. GIおよび GI.I Iを除き. その他の各世代に. 0 1 0 0尾体重測定を,稚魚では沖出し時, ついては 4ロット以上の個体群の 1. 選別時などに,成魚では出荷時などにそれぞれ行った。各世代のすべてのロッ トの測定値をプロットした. O. また,各ロットの成長曲線より商品サイズの 1kg. に達するまでの平均時間を算出し. 世代聞で比較した。. 8.
(16) 2000 N a t u r a l. 1000. 4. 内. -nu nunu n u n u. 500. 1000. 1500. 500. 1000. 1500. 500. 1000. 1500. 500. 1000. 1500. 500. 1000. 1500. 1000. 1500. GI. 1000. 司 'h. -nu nunu n u n u. 1000. OJ V. G. q4. nunu n u n u. ∞. h4. 'nu n-nu n u n u. (凶)五凶む診 h-uO﹄ωE ω k r︿. 1000. ・. G VVI. 1000. 。 o 2000. GVI ・ V I I. 1000 -nu n u. 500. D a y sa f t e rh a t c h i n g F i g . I I ・3 .Gr owthc u r v e so fe a c hg e n e r a t i o no ft h er e ds e abreamproduced bys e l e c t i v eb r e e d i n g . Gandromann u m e r a l sshowt h enumbero fg e n e r a t i o n s .. 9.
(17) ( 4 ) データ解析. デ}タは平均値および標準偏差 (mean士SD) で表示した。各世代聞の商品サ イズに達するまでの期間を Welch法47)により有意差検定した。 遺伝率は次式により求めた。 H=Y ( a 土1)- Ya Yap-Ya. Y ( α+1 ),a 世代目の選抜親魚より生産されたけ 1世代目の成長曲線より算出し. た 4歳魚の平均体重 ,Ya,a世代目の成長曲線より算出した 4歳魚の平均体 重 ,Yap,a世代目の選抜された 4歳親魚の平均体重.. Tablel l -1 .Thea v e r a g ebodyw e i g h to f4 y e a r o l d b r o o d s t o c ko f r e ds e abreamu s e df o rs e l e c t i v eb r e e d i n g Year. G e n e r a t i o n number. 1 9 7 2. Bodyw e i g h t * 包). 1 ) 2000( ラ. 1 9 7 5. l '1 1. 2, 440( 1 ). 1976. I I. 2, 7 3 5( 2 ). 1 9 7 8. I I . I I I. 600( 1 ) 2,. 1 9 8 0 1 9 8 1. I I I. 3, 089士 2 4 1( 4 ). 1 9 8 3. I I I .IV. 656土 1 6 7( 3 ) 3,. 1 9 8 4 1 9 8 5. I V. 3, 627士 405( 5 ). 1 9 8 6 1 9 8 7. IVV. 3, 48 8士 630( 8 ). 1989. V. 075( 2 ) 4,. 1 9 9 1. VVI. 1 0 9土 215( 4 ) 4,. 1994. V I .V I I. 009( 1 ) 5,. *Means土 SD.Numeralsi np a r e n t h e s e sshownumbero f t h e b r o o d s t o c kg r o u p s . ll-1-2. 結 果. ( 1 ) 選 抜 世 代 回 数 と 4歳親魚の体重との関係 I 1に示した。 1968年に天然親魚から採 各世代の 4歳親魚の魚体重を Table I. 卵した GIの 4歳親魚の平均体重は 2, OOOgで、あヮた。 GI I I( 1975~ 1977年産). 10.
(18) の 4歳親魚の体重は 3 , 089+2 4 1 g ( n = 4 )となり, GIに比較して1.5倍になった。 次に, GV.V I( 1 9 8 6 1 9 8 8年産)のそれは 4, 109+215g(n=4)となり GIの. 2 . 1倍となヮた。さらに, GVI.V I I( 1 9 8 91 9 9 2年産)のそれは 5 , 009g、 で2 . 5 倍となった。. Table1 1 2 .Th egrowthc u r v eandt h ea v e r a g ed a y sr e q u i r e df o ra 伐a i n i n g t h e c o m m e r c i a l 1 ya v a i l a b 1 es i z eo ft h ee a c hg e n e r a t i o no fr e ds e abream Growthc u r v e s G e n e r a t i o n Numrer number o f l o t s. 2 f ( x ) = a x +bx+c判 a. r * 2. DRA l k g * 3. c. b. 6 . 1 9. 0 . 9 3. , 059 1. 1 .0 8 1 .5 7 x l 0 2. 0 . 9 4. 953. 2 . 6 8. 0 . 9 7. 1 , 028. 4. 2 8 . 1 4 xl O -5 9 . 9 3 xl O -1 1 . l4 x10. 0 . 9 6. 985土 37. 5. . 2 1 xl O ・1 O -4 6 4 . 7 7 xl. 4 7 . 9 5. 0 . 9 7. 944土 67. 1 0. 48xl O ・4 8 2. . 0 1 xl O -. 8 2 . 9 5. 0 . 9 7. 1 .0 03土 82. 皿. I V. 7. . 2 7 xl O ・1 6 . 8 8 xl O -4 6. 6 6. 59. 0 . 9 7. : 1 :3 0 864:. I V 日T . V. 5. 7. 33xl O ・4 6 . 7 9 xl O -1. 5 3. 33. 0 . 9 9. 824土 26. l 3. 1 .21xl O ・3 8 . 5 2 xl O ・2. 5 . 9 0. 0 . 9 8. 934土 66. V. 5. 1 . 3Ox1 0 ・ 3 5 . 9 7 xl O ・3. 5 . 9 6. 0 . 9 9. 875土 2 3. VVI. 6. 41 xl O ・4 6 . 2 5 xl O -1 8.. 3 8. 25. 0 . 9 7. 1 770土 6. V I 'V I I. 1 1. 1 .25xl0 ・ 3 4. 45xl O -1. 3 0 . 5 5. 0 . 9 8. 738土 48. Naωral. 1. 59xl0 ・ 5 8 . 9 6 xl O ・1 0.. I. 1. 1 .37xl O ・4. 1 .I I. 1. O ・4 3 . 0 1 xl O ・l 6 . 5 7 xl. E. I I I I I 皿. 幽. * 1f ( x ) = b o d yw e i g h t( g ) ;x=d a y sa f t e rh a t c h i n g . * 2C o r r e 1 a t i o nc o e f f i c i e n . t * 3Thea v e r a g ed a y sr e q u i r e df o ra t t a i n i n g血ebodyw e i g h to f1kgwhichwas d e t e r m i n e dbyt h egrowthc u r v eo fe a c h1 0 . t. ( 2 ) 選抜世代回数と瞬化後の成長との関係. 各選抜世代マダイのすべてのロットの鮮化後の体重変動をプロットして F i g .. I I 3に示した。商品サイズの 1kgに達するまでの日数は,世代数が進むにつれ て短くなった。各世代の成長曲線および商品サイズに達するまでの平均日数を Table I I 2に示した。ロット毎の成長曲線より求めた 1kgに達するまでの平均. 日数は, G I Iでは 985土 3 7 ( n = 4 ), G I.I I Iでは 944士6 7 ( n = 5 )であったが, GV. .V Iでは 770土 6 1 ( n = 6 ), GV I.V I Iでは 738士4 8 ( n = 1 1 )となり, GI I I以下と GV .V Iおよび刀 VI.VIIとの聞にはいずれも有意差が認められた ( p< 0 . 01 )。. 11.
(19) ( 3 ) 実現遺伝率. 各世代 4歳魚の魚体重の実現遺伝率を T a b l eI I 3に示した。 GI-GII, GI. II-GI I.I I I, GII-G I I Iおよび GIII.IV-GN・Vの実現遺伝率は比較的低. く,それぞれ0 . 1 3,0 . 0 7,0 . 0 8および0 . 1 4であり,これらの世代では選択反応 は小さかった. O. しかし, GI I.III-GI I.I V, GIII-GIV, GIV-GVおよび. GV .VI-GVI.V I Iでは,選抜効果は高く,実現遺伝率はそれぞれ, 0.68, 0 . 5 8,0 . 3 5および0 . 7 8であった。 2)o T a b l el l 3 .R e a l i z e dh e r i t a b i l i t y( h f bodyw e i g h ta t4y e a r so fa g ei ne a c h g e n e r a t i o no f r e ds e abream G e n e r a t i o n s. h 2. GI-GII. 0 . 1 3. ・ 1 1- G1 1 . 1 1 1 GI. 0 . 0 7. GII-GIII. 0 . 0 8. G1 1 . 1 1 1- GI I I . I V. 0 . 6 8. GIII-GIV. 0 . 5 8. Grn.IV-GIV ・ V. 0 . 1 4. GIV-GV. 0 . 3 5. GIV.V-GV ・ VI. 0 . 2 0. GV ・ VI-GVI.VII. 0 . 7 8. M e a n : ! : :SD. 0. 33土 0 . 2 8. ll-1-2. 考 察. 著者らはマダイの 25年以上に亘る選抜育種に取り組み のような結果を得た。まず 増加するかを調べた. O. その成長について次. 選抜によヮてマダイ 4歳親魚の体重がどのように. その結果, GIにおける 4歳親魚の体重に比べて, G I I I. におけるその体重は1.5倍 , GV.VIには 2 . 1倍,さらに GVI.VIIには 2 . 5倍と なり,選抜世代回数を重ねる毎にその 4歳親魚の体重は増大することが分かヮ た。この結果は正の選択反応のひとつであるとみなすことができる. O. 次に,こ. れら各世代の親魚から作出したマダイ種苗の成長は明らかに世代を重ねる毎に 早くなり,商品サイズの 1kgに到達するまでの日数が短縮された。マダイ養殖 の最も基本的な形は. 当歳魚種苗を購入して 3ヶ年間養殖し,体重 1kgに仕上. 12.
(20) げることである. O. しかし,ここで選抜育種された V I,四世代目の稚魚は約 2ヶ. 年間でその大きさに達する. O. これは,養殖業者が種苗 (6-8cm) として購入. してから商品サイズに仕上げるまでの実質養殖期間は約 l年 7ヶ月間でよいこ とを意味する D H u l a t aら14)はナイルティラピアで成長を指標とした集団選抜を 2世代に亘り. 行い,もとの集団に比べ成長が向上しなかったことを報告している. o. Huang. and L i a o13)は,ナイルテイラビアを用いた成長率に関する l世代の集団選抜で. は実現遺伝率は低く,ティラピアの成長を向上させる方法としては集団選抜は 有効でないことを示唆した。本研究においても 3世代目以下では実現遺伝率は 比較的低く,ナイルテイラピア 13, んどみられなかヮた。しかし. 1 4 )と同様に初期世代の成長率の向上はほと. GI I I.IVあるいはそれ以降の世代 (GIV.Vを. 除く)では,実現遺伝率は 0.2以上の値を示し,選抜効果が明確に認められ た 。. 本研究においては. 4歳親魚の平均体重は GVI'VIIまで増加し続けた。. 実現遺伝率もまた減少しなかった。従ヮて. 本実験に用いたマダイは選択限界. に達していなかヮたと考えられる. 本実験はマダイ養殖の創始期から. 始めたものであり. 4 4 ). O. さらに. 本実験を遂行する間にも養殖技術は顕著に進歩した。. 3世代目以前にみられた低い実現遺伝率の一部は. 4 5 ). 未熟な養成技術によるのか. もしれない。 DonaldsonandO l s o n5)によれば. 2 3年間に亘って選抜育種したニジマスは成. 長率のみならず早期成熟,高水温耐性,抗病性および産卵数の増大など養殖魚 として大変有用な形質を獲得したという. O. マダイについても選抜育種により成. 長だけでなく,生残率,色・形,早期成熟,産卵数,卵質,ストレス耐性,耐 病性,肉質その他の形質に優れた品種改良を達成するために,今後一層の量 的,質的形質の向上に努力したいと考えている. C. さらに,マダイの人工種苗生. 産においては形態異常魚の出現が深刻な問題のひとつとなっており,形態異常 魚は出荷前に処分される D 形態異常魚,特に短躯魚が選抜育種により生産され たマダイにみられる点は選抜法との係わりにおいて検討しなければならない課 題である. O. I I- 2. 選 抜 育 種 マ ダ イ の ア イ ソ ザ イ ム に よ る 集 団 解 析 48). ll-lで述べたように. 選抜育種された親魚から得られたマダイ稚魚の成長. は世代を重ねる毎に明らかに速くなり,商品サイズの l k gに達するまでの養殖. 13.
(21) 日数が短縮された. O. しかし,選抜育種が数代繰り返されると. 優良形質の固定. 化が進むと同時に,遺伝的変異の減退や近交度の上昇に伴う劣性有害遺伝子の 顕在化などの影響が危倶される。魚類において近親交配により形態異常魚が出 現することが実際に報告されている o. 49-51). また,サケ・マス類では人工集団. における遺伝的な変化について多くの報告がある。. 52-57). しかし,我国で最も. 重要な養殖魚種の一つであるマダイについては,人工種苗集団の遺伝的形質に ついての研究は著しく少ない。 として用いられており 非常に重要である. 59). 58-60). マダイの養殖には選抜育種系の種苗が種. 選抜育種マダイの遺伝的形質について調べることは. O. マダイ種苗生産の初期において前轡症や短椎症などの脊椎異常魚がみられて いる o. 61,6 2 )本研究所においては,. の出現率は 5 . 8 3 3 . 7 %で. 1984年から 1993年の間のマダイ形態異常魚. 1993年には形態異常魚の 5 2 . 7土 17.9%を短躯魚が. 占めた。しかし,前轡症および短椎症はそれぞれ短躯魚の 1 2 . 1および13.6%に 過ぎなかった。(未発表)従って,最近の問題は顕著な脊椎異常を伴わない短 躯魚の出現である. O. 集団の遺伝的組成を把握するための一般的方法として,電気泳動法により検 出されるアイソザ、イムなどの分子多型がその遺伝標識として用いられる. O. この. アイソザイム多型を利用する方法は,多型の出現頻度から集団が保有する遺伝 子の頻度と変異の割合を直接的に推定できる点で優れている. O. また,集団の近. 交度についても遺伝子型の出現頻度から間接的に推定することができる. O. そこ. で本研究では,本学水産研究所において長年選抜育種されたマダイ種苗につい て,今後におけるその遺伝的側面の質的な改善を図るための基礎的知見を得る ためにアイソザイム分析による集団遺伝学的解析を行った。. II-2ー 1. 材 料 お よ び 方 法 ( 1 ). 供試魚. 選抜育種の過程および本実験に用いた個体群の由来を F i g .I I 4に示した。. 1968年から 1970年にかけて天然親魚より採卵 目の個体群より,それぞれ成長. 人工解化飼育した人工 I世代. 体色および体形の優れた個体を選抜し次世代. の親魚とした。これらの親魚群より得た仔魚を同様に選抜して次世代の親魚に する方法を繰り返した。採卵は自然産卵により行ヮたため,実際に次世代の生 産に貢献した親魚の数は不明であり, F i g .I I 4に示した数よりも少ないと考え られる o. 60). 1993年に生産された個体群のうち. 14. 1982年に解化した親魚(収.
(22) S V ¥. ' 9 3 ( V I I, V i l l ). AR. 3 4. U可 .. , .a色体伊九可. ' 9 3 ( V I I, V I I I ) 1 9 8 J l. 1 9 9 0 A 1 . 2 ' 9 3 ( V I I , V I I I ). 19908. ♀25♂ 1 5. A v h r. 叫有. 。 可 出. ♀ 25♂ 25. O. ー. .. ( ] 1. 匂. fAVAF. 凶. 本 -Fh. dFe. , 色 、. 、 戸 。. ♀1 2♂ 12. F i g .1 1 4 .T h eh i s t o r yo fs e l e c t i o nb r e e d i n gi nr e ds e ab r e a mu s e df o rp o p u l a i t o ng e n e t i ca n a 1y s i s . Numeralsona r r o w sshownumbぽ o f b r o o d s t ∞ ,kusedforproductionofnextgeneration.Romannumeralsi np a r e n t h e s e s showg e n e r a t i o no ft h ep o p u l a t i o n s ..
(23) T a b l e 1 1 4 .S a r n p l ef i s he x a m i n e da n dt h e i rs i z e h a r a c - Number Body F o l k Body Body Pop u 1 a t i o n so f C 血( c m ) e i g h t( g ) l e n g t h ( c m ) l e n g t h ( c m ) d e p t e r o f f i s h w b r o o d s t o c k. BLIBD*l. Normal 1 9 8 2 ( ♀ ,1 2 ;♂ ,1 2 ) S h o r t 時. 53. 6.0+1.1吃. 6.6+0. 4 5 . 1 2.56+0.05 . 1土0 . 5士0 . 3 2. 56. 5.8+1.3. 6.3+0. 4 5.3+0. 4 2.2+0.2 2.37+0.07. Normal. 51. 6.5+1.8. 6.9+0.6 5.7+0.5 2.2+0.2 2.56+0.05. S h o r t. 56. 7.3+2. 4. 7.0+0.8 5.8+0.7 2.4+0.3 2. 42+0.06. Normal. 52. 7.0+1. 4. 7 .1+0. 4. 5.9+0.3 2 . 3士0 . 1 2.58+0.06. S h o r t. 55. 7. 4 土 1. 4. 7 .0+0. 4. 5.9+0. 4 2. 4 士0 . 1 2. 4 2+0.05. 1 9 8 9 ( ♀ ,3 2 ;♂ ,1 町 1 9 9 0 A l ( ♀ ,5 3 ;♂ ,2 η. ー ー ー ー ・ ー ー ー ー ー ー ー ー ー ー ー ー ー ー ー ー ・ ・ ・ ・ ・ ・ ・ ー ー ー ー ー ー ー ー ー ー ー ・ ー , ー ー ー ー ー ー ー ー ・ ・ ・ ・ ー ー 圃 圃 園 田 ・ ・ ー ー ー ー ー ー ー ー ー ー ー ・ 圃 ー ー ー ー ー ・ 司 ・ ・ ー ー ・ ・ ー ー ー ー " ・ ・ ー ー ー ー ・ ー ー ー 圃 圃 ・ ・ ・ 自 国 圃 圃 圃 司 咽 咽 ・ 1 明治A -2 ( ♀ ,5 3 ;♂ ,2 η. Normal. 60. . 3 6 . 2土 1. 6.9+0. 4 5.7+0. 4 2 . 2土0 . 1 2.59士0.06. S h o r t. 52. 6.2+1.2. 6 . 7士0. 4 5.5+0.3 2 . 3土0 . 1 2. 41土0.06. 65. 5.8+1.0. . 1土0 . 1 2 . 6 1+0.06 6 . 8士0. 4 5.6+0.3 2. 64. 6.3+1.3. 6.6+0.9 5 . 6士0. 4 2.3+0.2 2. 41+0.05. Norm a 1 明均B 1 ( ♀ ,2 5 ;♂ , 25 ) S h o r t * 1B odyleng 也! B o d y d e p t h *2恥1 e a n : tSD 可. S h o r ts t a t u r e(Bodyl e n g 血!Bo dydep 由< 2 . 5 ) .. 容親魚数:♀. 12 ♂ , 1 2 ) より生産された lロット ( 2群) , 1989年産親魚. ( ♀ , 32 ♂ , 18) より生産された 1ロット ( 8 9群) , 1990年産親魚(♀,. 53; ♂ , 27) より生産された 2ロット (90A-1および90A-2群)および同年産 , 25) より生産された lロット (90B群)の計 5ロットをそ 親魚(♀, 25 ♂ れぞれ正常魚と短躯魚とに分類しサンプリングした。本調査に用いた各サンプ ルは 82群のみが人工生産 V.VI世代目であり. 89および90群はいずれも 1986. 年産の異なる親魚群に由来する人工生産刊・四世代目の種苗である. O. 各ロット. のサンプル数およびそのサイズを Table I I 4に示した D 体高に対する体長の比 が2 . 5に満たないものを短躯魚とした. O. また,軟 X線写真により,正常魚に骨. 格異常魚が混入していないことおよび短躯魚に標形成異常. 前轡症および短椎. 症が含まれていないことを確認した。各ロットのサンプルは,採取後直ちに氷 殺し, -30Cのフリーザーで分析まで保存した。 o. ( 2 ) 電気泳動. 電気泳動法により検出した酵素および遺伝子座,並びに検出に用いた部位お よび、緩衝液を Table I I 5に示した. O. 電気泳動法. 16. 染色法および、電気泳動バンド.
(24) パターンの判読は谷口,岡田 63)および、Taniguchi and Sugama64)に従った。検出 した 12酵素の泳動像に基づき 16遺伝子座を同定した。遺伝子座の表記法は, Shakleee t .a . 16 5 )に準拠した O. Table 1 1 5 .Th el i s to fe 皿 y mesexamined , l o c id e t e c t e d , t i s s u ea s s a y e da n db u f f e rs y s t e m su s e d f o re l e c t r o p h o r e s i si nr e ds e abreams a m p l e s Enz yme Commission. I ρ c u s. T i s s u e. B u f f e r *. A l ∞ ,holdεhydrogenase(ADH) α G l y ∞rophosphatedehydrogenase( α, -GPD). 1 . 1 . 1 . 1. Adh*. l i v e r. T-C. 民 a m i n o t r a n s f e r a s e(AAT) A s p a r t a. 2 . 6 .1 . 1. A αt 2申. l i v e r. C-APM. ST) E s t e r a s e(E. 3 .1 .1 . 3. E s t *. l i v e r. 1B E. G l u c o s e p h o s p h a t ei s o m e r a s e( G P I ). 5. 3 . 1 .9. i v e r , Muscle C-APM G p i l * L. I s ∞i t r a t ed e h y d r o g e n a s e( I D 町. 1 . 1 .1 . 4 2. I d h l *. l i v e r. La c t a t ed e h y d r o g e n a s eι D 町. 1 . 1 . 1 .2 7. Ld h 2 *. L i v e r , Muscle C-APM. Mal a t ed e h y d r o g e n a s e(恥1OH). 1 . 1 . 1 .3 7. Mdh-2* Muscle. 恥 匂l a 飽d e h y d r o g e n a s eNAD pt-仏1E). 0 1 . 1 . 1 .4. P h o s p h o g l u c o m u t a s e伊GM). 2 . 7 . 5 . 1. Pgm-l* l i v e r. C-APM. 6 P h o s p h o g l u c o n a t ed e h y d r o g e n a s e(6-PGD). 1 . 1 . 1 .44. 6 P g d 串. l i v e r. C-APM. S o r b i t o ld e h y d r o g e n a s e(SD H). 1 . 1 . 1 . 14. Sdh*. l i v e r. T-C. Enz yme( a b b r e v i a t i o n ). 1 . 1 . 1 .8. αG p d l * l i v e r. Me-l*. Muscle. C-APM. C-APM. C-APM C-APM. T-C , T r i s c i t r i ca c i dpH8 .0 ;C-APM , C i t r i ca c i d a m i n o p r o p y lm o r p h o l i n epH6 . 0 ;TBE, T r i s b o r a t eEDTAp H 8 . 9 .63,64) 合. ( 3 ) データ分析. 各個体群について Ha 紅r 匂 dy 待値との間の x が2検定を行つた. O. また,変異性の指標である 1遺伝子座当たりの. 平均対立遺伝子数,多型的遺伝子座率および平均ヘテロ接合体率を算出し,天 然集団と比較した D なお 天然集団は本実験で選抜育種に用いた祖先集団 が,和歌山県田辺湾周辺のものを中心に構成されていたため,三重および高知 4 )を参照することにした。さらに,各ロットの正常魚と における既報のデータ 6 短躯魚との間および正常魚各ロット間で、の対立遺伝子頻度の異質'性に関するが 検定を行った。. 17.
(25) T a b l e1 1・6 .A l l e l ef r e q u e n c i e sa tp o l戸no 中h i cl o c i岨 dc h i s q u a r ev a l u e sf o rd i f f e r e n c eb e t w e e no b s e r v e d祖 de x p e c t e df r e q u e n c i e so fg e n o t y p e si n n o r m a l組 ds h o r ts t a 加r er e ds e ab r e a mp o p u l a t i o n ss p a w n e df r o md i f f e r e n tg r o u p so fc u l t u r e db r o o d s t ∞k. 0.384 0.07. N o r m a l S h o r t 0.049 0.080 0.284 0.196 0.667 0.723 0.04 2 . 8 2. 0.529 0.74. 0.655 0.87. 0.679 0 . 3 2 1 2 . 9 1. 0.696 0.304 1 . 2 8. 0 . 8 2 1 0.179 0 . 5 1. 0 . 7 2 1 0.279 0.52. 0.827 0.173 2. 40. 0.692 0.308 5 . 0 3b. 0 . 7 3 1 0.269 1 . 5 6. 0.854 0.146 1 . 9 0. 0.859 0 . 1 4 1 1 . 7 1. 0.082. 0.050. 0.087. 0.185. 0 . 2 1 1. 0 . 2 4 1 0.304 0. 455 3 . 4 9. 0.205 0.009 0.616 0.170 0.50. 0.077. 0.198 0. 406 0.396 2 . 5 9. 0.137 0.010 0.637 0.216 0.16. 0.529 0.394 0.74. 0.327 0 . 5 9 1 0 . 0 1. 0.450 0.500 O. 0.365 0.548 4 . 1 1b. 0 . 4 3 1 0.385 6 . 1 1. 0.375 414 0. 3 . 1 4. 卯. 0.548 0.298 0.154. 0.518 0.345 0.136. 20 χ 2. 0 . 6 0. 1 . 7 8. 0.049 0.196 0.392 0.375 0.078 0.107 0 . 3 2 1 0. 480 1 2 . 3 1a 0. 48. 0.040 0.380 0.370 0.210 7 . 6 9. 0.036 0.300 0. 409 0.255 2.87. 0.025 0.350 0.325 0.300 2.94. 0.039 0.363 0.363 0.235 6 . 3 6. 0.078 0.539 0.313 0.070 0 . 1 6. 0.063 0.547 0.328 0.063 0 . 8 8. 0.795 0.205. 0.873 0.127. 0.929 0 . 0 7 1. 0.635 0.365. 0.627 0.373. 0.717 0.283. 0.647 0.353. 8 0 χ 2. 0.840 0.132 0.028 1 . 9 3. 1 . 8 0. 0.05. 2.07. 8.7~. 2 . 3 2. 3 . 2 1. 4 . 2 3b. 0.672 0.320 0.008 0 . 0 0. 0.648 0.336 0 . 0 1 6 4 . 6 0b. 1 2 0 1 0 0 χ 2. 0.226 0.774 1 . 8 1. 0.269 0.732 0. 48. 0.137 0.863 1 . 5 1. 0.134 0.866 2.65. 0.038 0.962 0 . 0 8. 0.055 0.945 0 . 1 8. 0.067 0.933 0 . 3 1. 0.069 0 . 9 3 1 0 . 2 8. 0.092 0.908 0 . 6 7. 0.070 0.930 0 . 3 7. A l l e l e -6 0 1 ω. N o r m a l 0.057 434 0.. S h o r t 0.125 0. 49 1. 1 3 0 χ2. 0.509 8 . 3 1a. A αt 2申. 1 0 0 2 0 0 χ 2. 0.585 0. 41 5 0. 41. E s t *. 1 2 5 1 1 0 1 0 0 ο 9 χ 2. Adh*. ー.. αコ. 1 2 0 1 0 0 M e 1 *. 1 0 0 P g m 1 ' ド. 6 P g d 申. 1 2 9 0 A l N o r m a l S h o r t 0.106 0.036 0.365 0.309. 1 2 9 0 A 2 N o r m a l S h o r t 0.100 0.087 0.333 0.250 0.567 0.663 0 . 0 2 0 . 0 0. l . 2 8 2 L o c u s. 卯. l . 2 8 2. a , bS i g n i f i c a n t l yd i f f e r e n tb e t w e e no b s e r v e da n de x p e c t e df r e q u e n c i e so fg e n o t y p e s( a,p<O.O1 ;b,pく 0 . 0 5 ) .. 陪 2 Q 己 N o r m a l S h o r t 0.069 0.039 0.492 0.430 0.438 0 . 5 3 1 0.76 0 . 2 8.
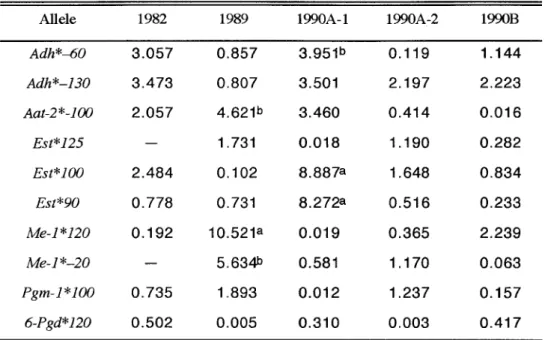
(26) II-2-2. 結 果 変異のみられた遺伝子座における対立遺伝子頻度および遺伝子型頻度の観察 値と期待値との聞の χ2 検定の結果を Table I I 6に示した。変異の認められた遺 伝子座は, Adh' ,Aat-2' ,Es , ' t Me-1' ,Pgm-1' ,6 P g d * 1 の 6遺伝子座であり, T a b l e 1 .G巴n e t i cv a r i a b i l i t yi nn o r m a lands h o r ts t a t u r ep o p u l a t i o n sspawnedfromd i f f e r e n tg r o u p so f 1・7 c u l t u r e db r o o d s t o c ka n dn a t u r a lp o p u l a t i o n so fr e ds e abreamb a s e don1 6l o c io fi s o z y m e C h a r a c t e r s P o p u l a t i o n o r s a m p l e s l o c a t i o n s. 1982. Average h e t e r o z y g o s i t y Ho. He. Ho!He. Normal. 53. 16. 1.625. 0.375. 0.375. 0.182 0.180 1.010. S h o r t. 56. 1 6. 1 . 5 6 3. 0.375. 0.375. 0.193 0.187 1 .032. 1 Norma. 5 1. 16. 1.688. 0.375. 0.375. 0.167 0.156 1 . 0 7 1. S h o r t. 56. 16. 1.688. 0.375. 0.375. 0.152 0147. Normal. 52. 16. 1.625. 0.375. 0.313. 0.199 0.172 1.159. S h o r t. 55. 16. 1.625. 0.375. 0.313. 0.169 0.159 1.065. 1989. 1990A-1. Averag 巴 Number P r o p o r t i o no f Number o f number p o l y m o r p h i cl o c i o fl o c i samples 11 e l e s o fa e d examined p* P examm p e rl o c u s. 1.035. ・ . . . . . _ ー . . . . . . . . . ・ ・ ・ ・ ・ ・ ー . . . . . _ . . . . . . . _ ・ ・ ・ ・ ・ ・ . . . . ‘ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ー . .. ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・・ ・ ・ . . . . . . . _ _ . . . . . . . . . _ _ . . . . . . . . . . _ . . . . . E. 1990A-2. 1990B. Normal. 60. 16. 1.625. 0.375. 0.375. 司 。 186 0 . 1 7 1. S h o r t. 52. 16. 1.625. 0.375. 0.375. 0.189 0.169 1.119. Normal. 65. 16. 1 . 6 8 8. 0.375. 0.375. 0.1830.166 1 . 1 0 1. S h o r t. 64. 16. 1.625. 0.375. 0.375. 0.170 0.162 1.046. Mie. 100. 16. 2.313. 0.688. 0.313. 0.179 0.167 1 . 0 7 1. Kochi. 137. 16. 2.625. 1 . 0 0 0. 4 38 0.. 0.1830.180 1.016. 1.088. a 1* l N a t u r. e f e rt ot h ed a t ao fMiea n dK o c h ii n1 9 8 7 .64) *1R P :丹o p o r t i o no fp o l y m o r p h i cl o c i . 戸. P r o p o r t i o no fp o l y m o r p h i cl o c i (maximuma l l e l ef r e q u e n c y< 0 . 9 5 ). H o : Av e r a g eh e t e r o z y g o s i t yo b s e r v e d H e : Av e r a g eh e t e r o z y g o s i t ye x p e c t e d .. 他の 10遺伝子座はいずれのロットにおいても全て単型であった。各サンプル集 団における遺伝子型の観察値と期待値との聞の χ2検定の結果, 82群正常魚の Adh*,89群短躯魚の Me-1*,90A-1群正常魚の Pgm-1¥90A-2群正常魚の Aat-2やおよび、短躯魚の E s t *とMe-1" ,90B群短躯魚の Me-1*にそれぞれ有意差が. . 0 5 )。 認められた(p<0.01,0 各ロットの正常個体群における 1遺伝子座当たりの平均対立遺伝子数,多型 的遺伝子座率および平均ヘテロ接合体率を. 既報の天然集団におけるそれぞれ. と比較してTableI I 7に示した。 16遺伝子座中の 1遺伝子座当たりの平均対立. 19.
(27) 遺伝子数(以下, A / Lと記す)では,天然集団の三重および高知における値が. .625であるのに対して,各ロットでは1.6251 .688で それぞれ 2.313および2 あり,いずれのロットにおいても明らかに低い値を示した。また,低頻度の変. .688,高知では 16 異を含めた全ての多型的遺伝子座率 (P)においても,三重で 0 遺伝子座全てで変異が確認されているが,各ロットでは低頻度の多型は全く認 められず,変異のみられた遺伝子座は前述の 6遺伝子座のみであったため,そ の値はいずれのロットでも主対立遺伝子頻度が0 . 9 5以下の多型的遺伝子座率( P っとほぼ同じ 0.375であヮた。しかし. P事についてのみ比較した場合,その値. は天然集団とほぼ同じであった。また,目。においても, 89群でやや低い値が みられたことを除けば,天然集団とほぼ同じかあるいはそれ以上の高い値を示 した O 近親交配の度合いを示す lつの指標として用いられる Ho/He で、は,全 てのロットで 1以上であり,近親交配の特徴であるホモ接合型過剰は認められ なかった。また,その値は 82群を除いて天然集団よりも高く, 90A-1群で特 に高い値を示した. O. 正常魚と短躯魚との問では, A / Lでは 82群と 90B群で. 正常魚が短躯魚よりも高い値を示したが,多型的遺伝子座率では各ロットとも 両個体群の聞に差はみられなかった。また,平均ヘテロ接合体率では, Hoで , 90A-l群および9GB群で は89群. Ho/Heで、は 90A-l群および90B群で. それぞれ正常魚が短躯魚よりも高い値を示した。そのうち, 90B群短躯魚の H. o/Heは l以下となり,ホモ型過剰を示したが,. その他の群では Ho および、 Ho. /Heについて短躯魚の方がそれぞれ高い値を示した。 各ロットの 6遺伝子座における各対立遺伝子頻度より,正常個体群を用いて. I 8に示し 行った各ロット間でのそれの異質性に関する χ2検定の結果を Table I た。その結果, 82群とその他の群との間で 8 9対立遺伝子に, 89群と 90A-1 および90B群との間でそれぞれ 6および 9対立遺伝子に, 90A-1群と 90B群 との問で 4対立遺伝子にそれぞれ有意差が認められた ( pく0.0010 . 0 5 )。ま た,9GA-1群と 90A-2群との間には有意差は認められず,出荷サイズの種苗 では,産卵日の違いによる影響はみられなかった。 正常魚と短躯との聞の対立遺伝子頻度の異質性に関する Table I I 9にまとめた. O. x2検定の結果を. 正常魚と短躯魚との間では, 89群で Aat-2*100,Me-. 1*120,Me-1*-20の 3対立遺伝子に, 90A-l群でAdh -60,Est*100および 虐. E s t * 9 0の 3対立遺伝子にそれぞれ有意差がみられた(pく 0 . 0 0 1,0 . 0 5 )。. 20.
(28) T a b l e1 1 ・ 8 .C h i s q u a r eva 1 u e so fh e t e r o g e n e i t yt e s tf o ra 1 l e l ef r e q u e n c i e sa t6polymo 叩h i c l o c i b e t w e e nnorm a 1r e ds e abreamp o p u l a t i o n ss p a w n e df r o md i f f e 民 n tg r o u p so fc u l t u r e d b r o o d s t o c k C o m b i n a t i o no f. A a t 2ヰ. Adh*. E s tキ. p o p u l a t i o n -100. s a m p l e s. -130. -1ω. 125. 15.59ga 19.48B a 11.16ga 7.954 b. ' 8 2 x ' 8 9. C 5.049. 5.299 C. 2.786. ' 8 2 x ' 9 0 A l. 1.029. 0.079. C 4.297. ' 8 2 x ' 9 0 B. 0.799. 1 . 1 8 1. a 21.582. ' 8 9 x ' 9 0 A l. 1.542. C 4.066. 0.157. 1.965. 8. 41 4 b. 0.936. ' 8 9 x ' 9 0 B 明. ' A l x ' 9 0 B. ' 9 0 A l x ' 9 0 A2 司. a 10.295b 11.978. s a m p l e s. 8. 47G b 22.893a. 3.201. 0.001. 21.78sa 28.27Qa. 0.151. 0.033. 1.025. 2. 489. b 7.732. 1.280. 9.76G b 7.621b. 3.785. 1 . 8 9 1. 6.239 C. 5.676 C. 2.229 0.022. 0.252. 0.322. 0.233. 0.689. 1.386 2.517. C o m b i n a t i o no f p o p u l a t i o n. S り. lω. 110. Me-1* 120. 100. 90. -20. Pgm-1牢. 6-Pgd 中. Iω. 100. ' 8 2 x ' 8 9. 60.96Qa. 2.018. 2.845. a 65.554. 0. 457. 2.767. ' 8 2 x ' 9 0 A l. 62.78sa. 1.529. a 12.38G. 24.34G a. a 1 1. 413. a 16.048. ' 8 2 x ' 9 0 B. 61.69ga. a 13.587. b 7.887. b 7.608. b 8.653. b 8.123. ' 8 9 x ' 9 0 A l. 0.096. 0.031. 24.79aa. 16.30aa. 15.65Qa. C 6.303. ' 8 9 x ' 9 0 B. 0.789. C 4.914. 18.83aa. 50.62Qa. 12.564 a. 1.160. 1. 416. C 5.703. 0.823. b 9.587. 0.353. 2.630. 0. 488. 2.302. 1.720. 0.874. 明. ' A l x ' 9 0 B. ' 9 0 A l x ' 9 0 A 2. 0.398. 0.212. , b, cS i 伊i f i c a n t l yd i f f e r e n tbe 何 回nd i f f e r e n tp o p u l a t i o n s( a , pく 0 . 0 0 1 ;b, pく 0 . 0 1 ;c, pく 0 . 0 5 ) . a. 1 I-2-3. 考 察 人工種苗集団において,アイソザイム遺伝子を遺伝標識とした対立遺伝子頻 度の変動や遺伝的変異の減退は. 様々な魚種で例がみられている o 52-58,60,63,. 6 6 7 1 ) マダイにおいても,その人工種苗集団の遺伝子頻度は,天然集団の遺伝. 子頻度に比べて有意に異なヮていたことがいくつかの研究により報告されてい. 2 1.
(29) Table 1 1 ・9 .C b i s q u a r ev a 1 u e so fh e t e r o g e n e i t yt e s tf o ra 1 1 e l ef r e q u e n c i e sa t6 p o l y m o r p b i cl o c ib e t w e e nnonn a 1a n ds h o r ts t a t u r er e ds e abreamp o p u l a t i o n s A l l e l e. 1 9 8 2. 1 9 8 9. 1 9 9 0 A l. 1 9 9 0 A 2. 1 9 9 0 B. Adh* -6 0. 3.057. 0.857. 3.951b. 0.119. 1.144. Adh*-130. 3. 473. 0.807. 3.501. 2.197. 2.223. A a t 2 * 1α3. 2.057. 4.621b. 3. 460. 0. 414. 0.016. 1 . 7 3 1. 0.018. 1.190. 0.282. E s t * 1 2 5 E s t * lω. 2. 484. 0.102. a 8.887. 1.648. 0.834. E s t * 9 0. 0.778. 0.731. a 8.272. 0.516. 0.233. Me-l*120. 0.192. 10.521a. 0.019. 0.365. 2.239. 5.634 b. 0.581. 1.170. 0.063. Me-l*-20 Pgm-l*lω. 0.735. 1.893. 0.012. 1.237. 0.157. 6-Pgd*120. 0.502. 0.005. 0.310. 0.003. 0.417. , bS i 伊i f i c a n t 1 yd i f f e r 四 tb e t w 田 nn onn a 1a n ds h o r ts t a t u r e p o p u l a t i o n s( p < O . O I, a. るo 58,60,63,69)今回,正常個体群を対象に調査を行った各ロットの遺伝的変異 の保有量についても,既報の天然集団におけるデ}夕 64)に比べ. 1遺伝子座当. たりの平均対立遺伝子数,および多型的遺伝子座率(P)について明らかな変異 の減退がみられた。また,その値は,前述のいずれの種苗生産機関の人工種苗 6 ' 7 1 ) しかし,これまで で観察されている値よりも低い水準であった。 58,60,63,6. にマダイで観察されてきた事例と,今回の調査結果とでは,本実験のマダイ種 苗が長期間にわたり選抜されてきた選択育種系であるという点で大きく異なっ ている. C. アユの人工種苗集団では,導入後の世代数が長いほどその遺伝的変化. の程度が大きくなっていたこと,また,選択育種系のそれでは,十分な親魚数 が確保されていたにも関わらず急激な遺伝子頻度の変化が認められたことが報 告されている o 67, 7 0 ) このような現象については. 単純な遺伝子の機会的浮動. の他に,種苗生産の過程で生じる無意識的あるいは意識的選択の影響が関与し ているものと考えられている. O. したがヮて,今回のマダイ種苗にみられた対立. 遺伝子や多型的遺伝子座の減少に関しても,種苗生産の創始期,もしくは長年 の継代交配の中で生じた少数親魚からの生産による低頻度遺伝子の消失に加 え,成長や形態に関して毎代強度の育種的選抜を行ってきたことに伴う変化で あることが示唆される. O. また,もう一つの特徴として,全体的な変異幅の減退とは逆に,多型の認め. 22.
(30) られた 6遺伝子座では天然集団と同等,もしくはそれ以上の高い水準で変異を 保持していたことが挙げられる 期待値との聞の. O. 各サンプル集団における遺伝子型の観察値と. x2検定の結果,82群正常魚の Adh"でみられた有意差について. は,ホモ接合型の個体が多くみられたホモ型過剰によるものであったが,その 他の群でみられた有意差は,いずれもヘテロ接合型過剰によるものであった。 さらに,いずれの個体群においても Ho/Hdまl以下の値を示さず,近親交配の 指標となるホモ型過剰はみられなかった。近交度の上昇が遺伝的変異の減退を 伴う場合,通常,平均ヘテロ接合体率は多型的遺伝子座率とともに減少し,. H. o/Heは l以下のホモ型過剰の状態を示す傾向にある。 Sugama e t .a . 160)はマダ イ人工種苗において遺伝子型の実測頻度とそれらの期待値が一致しない場合, その原因はいずれもホモ接合体過剰であヮたことを報告している おけるマダイ種苗について. O. 本実験に. 全てのロットでその Ho/Heが l以上となったこ. とは,これらの遺伝子座のヘテロ型個体に何らかの競争力の優勢が存在してい たか,あるいはサンプリング時以前の選別によりホモ型個体が除外されていた 可能性を示唆している. O. 実際に. 1994年に生産された種苗の 4ロットにおい. A a t 2 " ,Est~および、 6-Pgd") に基づく Ho は,大型 て , 3アイソザイム遺伝子座 ( 個体群では 0.375土 0.004であったが. ランダムサンプリング群では 0.303土. 0.001で,大型個体群の方が有意に高い値を示した (p<0.05) 閉鎖された集団内 0. において近親交配を防止,あるいはその影響を軽減させるための育種プログラ ムとして, Kincaid49)は,大規模なランダム交配集団を用いること,体系的な 系統交配を行うこと. 血縁関係のない親魚との雑種系統を形成させることの 3. ワの条件を提案している o F i g .I I 4より,本研究におけるマダイの祖先集団は 少なくとも 3種類の天然集団から成り立っており 混合を重ねてきていることがわかる. O. 以後それらの系統は複雑な. このことが結果的に. 親魚数の減少によ. る近親交配の進行を防ぎ,元来高頻度の変異を保持していた遺伝子座の変異保 有量を維持させてきた要因の一つであると考えられる. O. 一方,各正常個体群におけるロット聞での対立遺伝子頻度の異質性について の検定では,同一親魚群で産卵日の異なる 90A-1群と 90A-2群を除く全ての 組合せでその差が有意と判定された。特に,世代の異なる 82群とその他の群と の聞では. 2遺伝子座で対立遺伝子に置換が認められた。このことは,. 親魚数が他の親魚群に比べ著しく少なかったことに加え の由来の違いに関係するものと思われる. o. 82群の. 両世代間での系統上. Fig. I I 4より,本調査に用いた. 1989年産と 1990年産の両親魚 (6 ・7世代目)は,いずれも 1986年産の親魚. 23.
(31) 群(5 ・ 6世代目)から作出された個体群であり,さらにその 1986年産親魚は, 今回用いた 1982年産の親魚 (4 ・ 5世代目)と同一の親魚群に由来する個体 群であることが分かる. しかし, 1986年産の親魚群には,それらの他に系統. O. の異なる 1981年産親魚から作出された個体群が混合していたため,そのこと が今回の 82群と 89群および90群との間で,大きな遺伝的較差が生じた一つの 要因になったものと考えられる. O. 一方,同年産の親魚群に由来する 89群と 90. A-1群および90A l群と 90B群との聞で生じた有意差については,いずれも 司. 採卵用親魚を選ぶ際の抽出誤差,あるいは繁殖に参加した親魚数もしくはその 雌雄比の違いによるものと考えられる. O. マダイでは,同一親魚群より lシーズ. ン中の数次にわたり採取した卵から別々に生産された個体群では,その対立遺 伝子頻度はそれぞれの個体群聞で有意に異なっていたことが報告されている。 6 0 ). 今回, 90A-1群と 90A-2群との間で有意差がみられなかったことは,そ. の産卵日の違いが 1日であったため,繁殖に参加した親魚に大きな違いがみら れなかったものと思われる。 通常,有効親魚数が50であるとき,その子孫集団の近交系数は毎代 1%ずつ 上昇するため, いる o. 5 1 ). 5~10 世代目には近親交配の影響が現われることが予想されて. 魚類において近親交配の蓄積による近交弱勢効果の発現は,主にサ. ケ科魚類を中心とした淡水魚においてその報告例がみられる o マス 51)やゼブラフイッシュ 50)では たことが報告されている. O. 49). 特に,ニジ. 兄妹交配により骨格異常個体が多く出現し. 今回,正常個体群と短躯個体群との間で,その遺伝. 的変異を比較したところ,本実験に用いたマダイ種苗の各ロットでは,短躯魚 の出現に,遺伝的変異の減退ならびに近親交配の影響が関与していることを示 すデータは得られなかった。また. 各対立遺伝子頻度の異質性についての検定. では,正常魚と短躯魚との聞では 2群で有意差が認められたものの,残りの 3 群にはみられなかった. O. 形態的特徴のような量的形質はポリジーン性. ( p o l y g e n i c ) 7 2 )といわれ,複数の遺伝子座からの働きを受け,影響を及ぼし合. い,また環境の影響を強く受ける形質とされている. O. 近交度が上昇することに. よって形態異常個体が出現するような現象は,そのような遺伝的要素の顕在化 を示唆している. O. 谷口ら 73)は 体形異常をもたらす要因に遺伝的要因があると. すれば,それは異常の発現を直接的に支配する遺伝子ではなく,発現に関する 後天的要因に対しての感受性に関係する遺伝子であることを推察している. O. 一. 方,短躯魚は同じ体長の正常魚に比べより多くの可食部を持つ。本実験におい ても,短躯魚の体重は 3群で正常魚よりも高い値であった。短躯魚の出現と近. 24.
(32) 親交配あるいは遺伝的変異の減少との明瞭な関係がみられなかったことは,短 躯魚がそのような優れた形質をも持ち合わせている魚として利用されうる可能 性を示唆する. O. 短躯魚の遺伝率,成長,発現機構といった形質については今後. の検討課題である. O. 今回の調査より,本実験に用いたマダイ種苗では,その長年にわたる継代交 配にも関わらず,ホモ接合型過剰,すなわち近親交配の影響は認められなかっ たものの. 1遺伝子座当たりの平均対立遺伝子数および多型的遺伝子座率の減. 退より,集団としての遺伝子保有量は天然魚に比べ著しく減少していることが 示唆された。このことは. 優良形質の獲得と固定化を目指す選抜育種の観点か. らみれば,本学のマダイ種苗が選択育種系として遺伝的に均質化されてきてい ることを示す正の効果であるとみなすこともできる. O. しかし,種々の潜在能力. を秘める本来備えているべき遺伝資源の消失という観点からみるならば,今後 選抜による正の効果とは別に. 様々な環境要因に対する適応度の低下など負の. 効果の生じる可能性のあることは否定できない。また. 継代交配を繰り返す人. 工種苗集団では兄妹交配もしくは半兄妹交配の行われる可能性は高く,さら に,親魚の雌雄比が異なる場合その確率は高くなることから, 51) 安易な親魚の 選出により今後近親交配が生じる可能性は十分考えられる. O. したがって今後. は,採卵に用いる親魚数を増加させ,その雌雄比を等しくさせること,系統上 遠縁の個体同士をかけ合わせること. 産卵日もしくは親魚群の異なる複数の. ロットより次世代の親にすることなどにより,さらなる遺伝的変異の減退,近 交度の上昇および遺伝子の機会的浮動を防ぎ. 量的形質だけでなく質的にも優. れた形質をもっ種苗を生産する必要があると考える。. 25.
(33) 第田章人工交雑による品種改良 画一. 1. 交雑魚の成長,外部形態および環境ストレス耐性. 養殖魚類の生産効率を高めるためには,より良い養殖特 性を持ワ種苗の作出 d. が必要である. O. その手段のーっとして交雑による品種改良が古くから試みられ. てきた。淡水魚については,コイ,キンギヨ,. ドジョウ,サケ・マスなどに関. して多くの知見が報告されている 20,74,7 5 )。一方,海産魚では,主としてタイ 76-78),イシダイ科 79, 80),プリ属 81-83)およびトラフグ属 24)の交配が試みら 科 25,. れている. O. しかし. それら交雑魚の養殖品種としての評価に関する報告はまだ. 著しく少ない。. m-1- 1. マダイ Xクロダイ 84) 筆者らは,これまでにマダイ,クロダイ A c a n t h o p a g r u ss c h l e g e l i ,ヘダイ Sparus s a r b α,チダイ E vynnis} α r p o n i c αを親魚としてそれらの交雑を試みてきた。. 76, 85). そこでここでは,マダイ(♀ ) Xクロダイ(♂)の交雑種(以後マクロダイと呼ぶ) の特牲を明らかにする目的で,その成長,生残率,外部形態および環境ストレ スに対する耐性などをマダイおよびクロダイと比較した。 m - l - l ー 1. 材料および方法 ( 1 ) 親魚. マダイ親魚としては人工生産種苗から田辺湾内に設置した本学水産研究所の 網いけすで 5年間養成した体重 4, 060g,尾叉長 57.6cmの雌 1尾と,体重. 4, 870g,尾叉長 63.0cmの雄 1尾を用いた。クロダイ親魚としては同じく人工生 産種苗から 7年間養成した体重790g,尾叉長 34.0cmの雌 l尾と,体重650g, 尾叉長 30.5cmの雄 l尾を用いた。 ( 2 ) 採卵および人工受精. 1990年 3月 17日に,性腺刺激ホルモン(ゴナトロビン,帝国臓器社製)を 500I .U.,クロダイ雌には 1, 200I .U .それぞれ背筋部に注射し マダイ雌には 2, た そして, 48時間後に採卵した。マダイ卵は二ヲに分けて両魚種雄それぞれ G. の精子を用い,クロダイ卵にはクロダイ精子を用いて,乾導法により人工交配 を行い 3つの組み合わせを作った。媒精に用いた卵数は,マダイとマクロダイ 用は等分になるように各 109, 000粒とした D クロダイ用のそれは 130, 000粒で. 26.
(34) あヮた。媒精した卵は一時 30e 容水槽に入れた後,沈下した未受精卵および死 卵を除去し,浮上卵を各々 500e ポリカーボネイト水槽に収容し,止水換水,. 0 " " '5 0万c e l l s /m~添加,水温18 C の条件下で受精卵 通気,ナンノクロロプシス 4 0. の管理を行ヮた。発生卵が 8" " ' 1 6細胞期に入った時点で浮上卵の一部を抽出し て万能投影器で発生卵数を計測し受精率を算出した。次に媒精約 24時間後の脹 粒ずつ無作為に抽出して 1 .5 e容ポ」ルに収容し,正常鮮化仔 体形成卵を各 100 魚の解化率を求めた。 ( 3 ) 仔稚魚の飼育. 容水槽各 l面ずつに移して解化さ 媒精後 2日目の妊体形成卵を別の 500e せ,仔魚の飼育を行った。飼育水は鮮化後 7日目から 20日目までは止水換水式 とし,以後徐々に流水式に変えた. O. 飼育水にはナンノクロロプシスを 40" " ' 5 0. 万c e l l s /m~ の密度になるように解化後20 日日まで毎日添加し, 1 5日目までは穏 やかに通気した。シオミズツボワムシを瞬化後 3日目から,アルテミアノープ. 0日目から,魚肉ミン リウスを鮮化後 18日目から給与し,市販の配合飼料は 2 5日目から給餌した。 チは 2 ( 4 ) 稚魚沖出し後の養成. 本実験では瞬化後71日目(全長 50mm前後)に稚魚を室内飼育水槽から海面 網いけすへ移すいわゆる沖出しをした. O. 実験場地先海面に垂下した縦・横・深. さ各 3mの網生賛 3面に各稚魚を収容し. マダイ用ドライベレットと冷凍マイ. ワシおよびイカナゴのミンチ肉を給餌した。マクロダイと両純粋種の成長を比 較するため 2歳魚までは約 1ヶ月日毎に. 3歳魚からは 2""'3ヶ月日毎に魚体. 測定(全長,尾叉長および体重;n =20) を行った。 ( 5 ) 外部形態. 形態的特徴を明らかにするために,マダイ,クロダイおよびマクロダイの各. 1 0尾を用いて鮮化後270日日および解化後947日目に次の各形質を測定し比較 した。計測的形質としては体長,頭長,頭高,吻長,上顎長,眼径,限窃幅, 背鰭赫長,腹鰭赫長,啓鰭赫長,胸鰭長,最長胸鰭条長,背鰭基底長,啓鰭基 底長,尾柄高,腸長および体重を測定した。また計数的形質としては背鰭,胸 鰭,腹鰭,啓鰭,尾鰭の鰭条数並びに側線上横列鱗数,側稼下横列鱗数を計数 した。なお計測,計数は岩井 86),赤崎 87)の方法に準じた。 ( 6 ) 環境ストレス耐性. 水温上昇に対する耐性試験は癖化後288日目に,水温低下に対するそれは排 化後326日目に,それぞれ前記の 3種を各 5尾ず、つ用いて行った。水温の上昇. 27.
(35) は 3~4 時間で限界水温(推定35 C) に達するようにボイラー加温海水を用いて 0. 1時間当たり 4 . 5~4.80C の上昇速度とした。水温の低下はユニットクーラーに よる冷却海水およびあらかじめ凍らせた海水を貯水槽に投入し,. 3~4 時間で. 限界水温(推定 7C ) に達するように 1時間当たり 2o Cの低下速度とした。実 0. 験開始時から 15分毎に水温, 30分毎に溶存酸素含量を測定し,供試魚の状態 を観察した。 海水比重低下耐性試験は鮮化後 285日日に各 5尾ずつ使用して行った。海水 比重の低下は水道水をチオ硫酸ナトリウムで中和して試験水槽に徐々に注入 し,約 3時間後に比重が1.0 (赤沼式比重計で測定)となるように 1時間当た り0 . 0 0 8ずつの低下速度とした。 溶存酸素低下耐性試験は,勝化後326日目に同じく各 5尾ずつを用いて行っ たO 試験水槽に流水のまま窒素ガスを通じて溶存酸素含量を徐々に低下させ た 。. なお,瞬化後 326日 Eにおける各供試魚の平均体重 ( g )はマダイ, 1 79.2. 4;クロダイ, 9 8 . 1土 2 5 . 8;マクロダイ, 1 9 0 . 3士6 5 . 5であった +24.. O. IH-1-1-2. 結 果 お よ び 考 察. ( 1 ) 受精率および瞬化率 受 精 卵 の 平 均 卵 径 (n=10)はマダイ 0.898士0.019mm,クロダイ 0.867土. 0.012mmであった. O. 受精率はマダイ. 74.0% ;クロダイ. 9 5 . 3% ;マクロダ. イ , 55.5%でありマクロダイの受精率は他の 2者より劣った( p < 0. 0 01 )。癖化率 はマダイ, 85.7%;クロダイ, 39.8%;マクロダイ, 40.0%でありマクロダイ. 0 . 0 0 1 )。 は解化率でもマダイより低かった(p< ( 2 ) 仔稚魚の成長および生残率. 解化仔魚飼育開始時の尾数はマダイ, 12.900尾;クロダイ, 37, 300尾;マ. 500尾であった。瞬化後 3日目からシオミズツボ、ワムシを給与し クロダイ, 9, n = 1 0 )は , たところいずれもよく摂餌した O 解化後数時間目の仔魚の平均全長( マダイ, 3.00土 0.06mm ,クロダイ, 2 . 7 9土 0.17mm;マクロダイ, 3.11土. 0.07mmで、あったが,鮮化後 30日日にはマダイ, 1 2 . 5士1.06mm ,クロダイ, 8 . 3 1土 0.97mm,マクロダイ, 18.8+2.06mmとなり,マクロダイの成長が最も p < O . O01 )0 一方, 30日目の生残率はマダイ, 28.1% マ ク ロ ダ イ , 良かった (. 6.3%;クロダイ, 1 .6%であり. マクロダイはマダイよりも 1 f tくクロダイより. p < O . O O l )。荒川,吉田 25)によればマダイとチダイの交雑種の沖出し も高かった ( 時の生残率はマダイに比べ著しく劣り. 特に日齢 5までの交雑種の生残率がマ. 28.
(36) ダイの約 40%に過ぎなかったのは雑種劣性の可能性もあると報告している. O. マ. ダイに比べてマクロダイの受精率,解化率および解化後 30日目までの生残率が いずれも劣るのは初期発生におけるこのような雑種劣性の影響ではないかと考 えられる o. 8 8 )しかし,解化後 71日目から. 140日目における 70日間の生残率はマ. ダイ(約 70%) に比べマクロダイとクロダイ(約 100%) が著しく高かヮた ( p< 0 . 0 0 1 )。. ( 3 ) 当歳魚の成長 2 8. 28. 2 4. 24以 。 、 r } -. 以 2 4。. ~. 205 Z g. ( E18. 20E ω E. E. ー 。 1 6己 苦 主. i. 自. 1 6 E 5 L £ S. ー1 2 l. 百 事 。. 1 2. 3 0ト. R由. --$-. R s bXB s b. 主 ー 宮 陸. l. 届. 6 -. I智 IO. 一_ Bsb. . . . . 1 2. , J J /. 制. ~. ". 』. --G- R s b. 一 線 - RsbXBsb. ー _. B s b. o. o 60 120180240300360. 0 6 01 2 0180240300360 D a y sa f t e rh a t c h i n g. F i g . I I I -1 .Comparisonofgrowthi nO y e a r o l dr e ds e abream(R s b ), b l a c k s e abream( Bs b )andt h e i rh y b r i d s(R s bXB s b ) .. 解化後 360日目までの平均体重および平均全長を比較して (n=30)F i g .I I I 1に 示した。解化後 6ヶ月日における当歳魚マクロダイとマダイの平均体重はクロ ダイの約 2倍であった. O. 勝化後 8ヶ月日においてはその差は更に大きくなっ. た。解化後 12ヶ月目ではマクロダイよりもマダイの方が僅かに大きくなった。. ( 4 ) 1' "3歳 魚 の 成 長 1---3歳魚の成長を比較して (n=10-30)F i g .1 I I 2に示した。解化後約 1年目. からマクロダイよりもマダイの方が徐々に大きくなり イはクロダイの約 2倍. マダイは更にその約 2倍となった。コイとフナの雑種. の成長はコイよりも劣るがフナよりも良く るo. 満 3年目にはマクロダ. 両親の中間であると報告されてい. 8 9 )マクロダイの成長もマダイより遅いがクロダイよりは早く,両親の中間. 29.
(37) 1600r. 28. J128. A. 24E 。 ) J E. 24LJ ). 器 』 16障. 16B 器 』 E. E. 言 語1 200. 音 ω ~色. 主 800. z. 12. 普 ・ 邑 b 宅. 900. 600. --G- Rsb. 15 1200 300 Daysa f t e rh a t c h i n g i $ -. RsbXBsb. 900. 600. 1200. ・. Bsb. 1 1 2 . Comparisonofgrowthi n1・3・year-oldredseabream(Rsb), F i g .1 blackseabream( Bsb)andt h e i rhybrids( RsbXBsb).. であった. O. ( 5 ) 外部形態の比較. 計測的形質として. マダイ. クロダイ. マクロダイそれぞれの頭長に対する. 5形質測定値の比率を比較 LTable I I I 1に示した。マクロダイに対して,マダ イは眼径比を除く頭高比 ( p < O .001 ),q 身長比ω<0.001),上顎長比 ( p < 0 . 0 5 ),眼商幅 比( p< 0 . 0 01 )の 4形質に有意差があり,クロダイは眼寓幅比 ( p< 0 . 0 5 )のみに有意差 がみられた。 T a b l eI l l l .C o m p a r i s o no f5m o r p h o l o g i c a lc h a r a c t e r sb e t w e e nr e ds e abream (R s b ),b l a c ks e abream( B s b )a n dt h e i rh y b r i d s( R s bXB s b ) C h a r a c t e r *l. Rsb. RsbXBsb. Bsb. 1 .4 3 1士0 . 0 5 1*2, a. 1 .256+0.044. 1 .218+0.039. S n o u tl e n g t h. 0. 41 2+0.041a. 0 . 3 4 9土0 . 0 2 5. 0 . 3 4 2土0 . 0 3 2. Lengthofup 戸r jaw. 0 . 3 6 0土0 . 0 4 3b. 0.323+0.024. . 0 1 6 0 . 3 2 9士0. EyeD i a m e t e r. 0.239+0.025. 0.249+0.023. 0.286+0.045. S u b o t b i t a lw i d t h. 0 . 2 1 7土0 . 0 26 a. 0. 236+0.023. b 0.250+0.022. Headdepth. * 1E x p r e s s e da st h er a t i ot oh e a dl e n g t h *2Mean+SD( n = 1 0 ) a, bS i g n i f i c a n t l yd i f f e r e n tf r o mt h ev a l u e so fh y b r i d s( a , p < O . O O I ;b,p < 0 . 0 5 ) .. 30.
図


+7





関連したドキュメント
5世紀後半以降の日本においても同様であったこ
この論文の構成は次のようになっている。第2章では銅酸化物超伝導体に対する今までの研
次に、第 2 部は、スキーマ療法による認知の修正を目指したプログラムとな
この条約において領有権が不明確 になってしまったのは、北海道の北
このたび牡蠣養殖業者の皆様がどのような想いで活動し、海の環境に関するや、アイディ
彼らの九十パーセントが日本で生まれ育った二世三世であるということである︒このように長期間にわたって外国に
単に,南北を指す磁石くらいはあったのではないかと思
(batter)又はパン粉でおおった魚の切身、加熱による調理をした魚)
