Kyushu University Institutional Repository
生物の知性の探求と知能ロボットへの応用
山口, 達也
https://doi.org/10.15017/1931719
出版情報:Kyushu University, 2017, 博士(機能数理学), 課程博士 バージョン:
権利関係:
博士学位論文
生物の知性の探求と知能ロボットへの応用
九州大学 大学院 数理学府 数理学専攻 機能数理学コース
山口 達也
目 次
第1章 はじめに 7
1.1 生物の知性が秘めた可能性 . . . . 7
1.2 脳の起源を探る . . . . 9
1.3 本論文の構成 . . . . 10
第2章 テトラヒメナの記憶メカニズムを利用した知能ロボット制御の提案 11 2.1 繊毛虫の記憶行動 . . . . 11
2.1.1 テトラヒメナの記憶行動と数理モデル . . . . 11
2.1.2 正多角形容器におけるテトラヒメナの記憶ダイナミクスの数値実験 16 2.1.3 ゾウリムシの記憶行動と数理モデル . . . . 18
2.2 対向二輪型ロボット制御への適用 . . . . 22
2.2.1 使用機材 . . . . 22
2.2.2 数理モデルの書き換え . . . . 24
2.3 iRobot Create 2 を用いた実機実験 . . . . 27
2.4 この章のおわりに . . . . 28
第3章 アメーバの位相振動子モデルに着想を得た情報処理メカニズムの提案 31 3.1 位相振動子の集団同期を用いたアメーバの学習モデルの紹介 . . . . 31
3.2 アメーバの時間記憶モデルの書き換え . . . . 33
3.2.1 数理モデル構築 . . . . 33
3.2.2 振動子の振る舞い . . . . 33
3.2.3 数値計算 . . . . 34
3.3 アメーバモデルに着想を得た入力情報を検出する位相振動子モデル . . . 35
3.3.1 問題提起 . . . . 35
3.3.2 Excitation-Inhibition(EI)モデル:二つの周期的な入力情報の位相 差を検出,学習しスループットする仕組み . . . . 36
3.3.3 T-shiftモデル:二つの周期的な入力に対して特定の時間差の組み 合わせのみ学習し,スループットする仕組み . . . . 39
3.3.4 Frequency Filter(FF)モデル:入力を周波数ごとに分類し学習する 仕組み . . . . 40
3.4 この章のおわりに . . . . 48
第4章 ダンゴムシの行動選択 49 4.1 平坦な地面の円形空間とすり鉢状の地面の円形空間 . . . . 49
4.1.1 実験装置と実験方法 . . . . 50
4.1.2 結果 . . . . 50
4.2 ワッフル状の地形での行動選択 . . . . 52
4.2.1 実験装置と実験方法 . . . . 53
4.2.2 結果 . . . . 53
4.2.3 数理モデリング . . . . 53
4.2.4 考察 . . . . 55
4.3 ダンゴムシの記憶行動を模した対向二輪車のアナログ制御機構の提案 . . 56
4.3.1 数理モデリング . . . . 57
4.3.2 数値計算 . . . . 60
4.4 集団の認識 . . . . 65
4.4.1 実験装置と実験方法 . . . . 65
4.4.2 結果 . . . . 66
4.4.3 考察 . . . . 66
4.5 この章のおわりに . . . . 67
第5章 おわりに 69
参考文献 71
図 目 次
1.1 (a)トヨタの歩行支援ロボット「ウェルウォークWW-1000」.トヨタホー
ムページより[13].(b)チェルナー錯視の例[14]. . . . . 9
1.2 アメーバの餌間ネットワーク.[16]のFig.1より. . . . . 9
2.1 テトラヒメナの記憶現象に対するKunitaの実験結果.[24]のFig.1より. 11 2.2 細胞の移動.[24]のFig.2(a)より. . . . . 12
2.3 数値実験の結果.[24]のFig.3(a)(b)より. . . . . 15
2.4 数値実験の結果.[24]のFig.3(c)より. . . . . 15
2.5 (A-a) 円形容器内で壁に沿った移動.(A-b) 広い空間で壁がない場合の移 動.容器の壁と進行方向とのなす角がϕ0のとき,壁に沿った移動の速さ はv¯′ = ¯vcosϕ0である.図中の(0,0)は円形容器の中心を表す.壁際でス リップする動きを仮定することで,微小時間では円形容器の中心,細胞の 位置(A-a),細胞の位置(A-b)は全て一直線上にある.(B-a)円形容器ない で壁に沿った移動.(B-b)広い空間で壁がない場合の円運動.円形容器の 中心と円運動の中心を重ねた. . . . . 16
2.6 一辺が800の正三角形容器に細胞を閉じ込めた場合に,テトラヒメナの記 憶ダイナミクスに従って動く細胞の軌跡(t ∈[1490,1500]). . . . . 17
2.7 正三角形容器に細胞を閉じ込めたとき,十分時間が経った後に内接円の軌 跡を描いている時間(図 2.6)での細胞が容器の壁と接触するタイミング. (a) 接触感知センサδの値.(b) (a)のδをFFTした結果.(c) (b)の結果を 拡大した図. . . . . 18
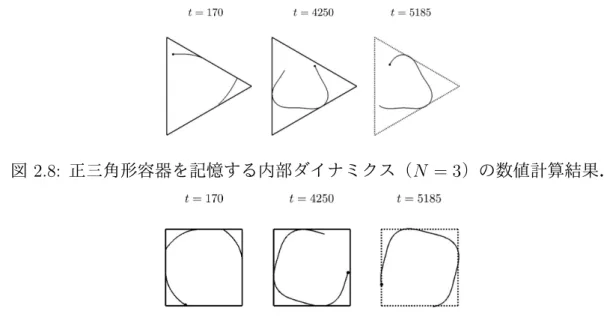
2.8 正三角形容器を記憶する内部ダイナミクス(N = 3)の数値計算結果. . 21 2.9 正方形容器を記憶する内部ダイナミクス(N = 4)の数値計算結果. . . 21
2.10 速さを一定としたときに,周期的な方向転換によって得られる移動の軌跡 の例. . . . . 21
2.11 使用した機材.(a) ルンバ.(b) Arduino.(c) Wifiカメラ. . . . . 23
2.12 ルンバのポート説明[34]. . . . . 24
2.13 式 (2.31)の相図. . . . . 26
2.14 ルンバが壁と接触している時間の制御.黒実線は壁,黒破線は移動の軌跡 を表す.(a) 壁との接触前はルンバの記憶ダイナミクスに従って移動する. (b) 壁を検出すると回避行動が発現する.(c) 回避行動とはその場でのター ンである.回避行動はセンサが壁を検出しなくなるまで継続し,回避行動 中はルンバの記憶ダイナミクスには従わない.(d) 壁を検出しなくなった ら再びルンバの記憶ダイナミクスに従って移動する. . . . . 26
2.15 Arduinoに実装したプログラムの概略. . . . . 27
2.16 ルンバの記憶ダイナミクスを用いて実機実験を行った結果.(a)実験開始 後20秒から40秒までの移動の軌跡.(b)実験開始後210秒から230秒まで の移動の軌跡.(c)軌跡から見積もられた3秒間の移動距離. . . . . 28
3.1 y= tanh(2x+ 3)のグラフ. . . . . 32
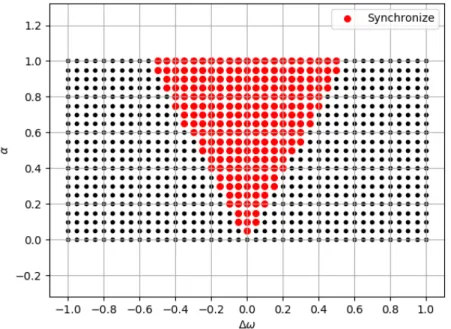
3.2 Saigusaらのアメーバモデルにおける振動子の振る舞いの模式図.図形の 違いは固有角速度の違いを表す.(a)初期状態.振動子は全てバラバラに 分布している.(b)周期的な外部刺激が繰り返された後の状態.外部刺激 のタイミングと近い固有角速度を持つ振動子集団のみが同期して位相が揃 う.(c)周期的な外部刺激がなくなった状態.同期した振動子集団は少しず つ固有角速度は異なるため,時間が経つと徐々に集団同士が離れてゆく. 33 3.3 二つの振動子に対して,ある刺激感受性α(縦軸)に対して同期できる振
動子の固有角速度の範囲.横軸は外力の角速度からみた振動子の固有角速 度のずれ∆ωを表す.赤丸:少なくとも一つの初期位相(θ0(0)̸=θ1(0))で 同期できる.黒丸:いかなる初期位相(θ0(0)̸=θ1(0))でも同期できない.
(β = 0.05とした.) . . . . 35 3.4 式(3.3)の数値計算結果. . . . . 36 3.5 EIモデルの概略図. . . . . 37 3.6 p= 0.5,ϕ= 2π/3とした場合のEIモデルの振る舞い.(a) 入力情報.(b)
各振動子集団の活性度合い.(c) t= 500における各振動子集団のローカル 同期の状態.(d) 出力. . . . . 38 3.7 様々な位相差を持つ入力情報に対するEIモデルで得られた出力の位相差. 38 3.8 T-shiftモデルの概略図. . . . . 39
3.9 Tshift =πのT-shiftモデルを数値計算した結果.Input pattern I:外部入力
をt∈[20,100]のときE1(t) = sin(0.5t), E2(t) = sin(0.5t+π),t∈[200,205]
のときE1(t) = E2(t) = 1,それ以外の時間でE1(t) = E2(t) = 0とした.
Input pattern II: 外部入力をt ∈[20,100]のときE1(t) = sin(0.5t), E2(t) = sin(0.5t),t ∈ [200,205]のときE1(t) = E2(t) = 1,それ以外の時間で E1(t) =E2(t) = 0とした. . . . . 40 3.10 (a)振動子の固有角速度の分布例.(µ, σ2) = (5,1)の正規分布に従って2万
個(J = 20000)の振動子を分布させた.振動子の固有角速度は[0.01,10]で,
dω = 0.01である.外部刺激の角速度がµに近いほどグローバル同期度が
大きくなる.(a-1) 振動子の固有角速度の分布の様子.赤い帯は振動子が 同期しているだいたいの場所である.(b)複数の周波数の重ね合わせに対 する振動子の振る舞い.I(t) = sin(3t) + sin(5t)に対しては角速度3付近と 角速度5付近の振動子が同期するが,グローバル同期に影響を与えるのは 個数がより多い角速度5の振動子の方である.比較のためにI(t) = sin(5t) に対する応答(緑実線)も載せた.(b-1)の振動子分布にある赤い帯は同期 した振動子が存在するだいたいの場所を示す.(b-2) 振動子の固有角速度 ごとのローカル同期度の時間変化の様子. . . . . 44 3.11 振動子の固有角速度の分布の例.(µ1, µ2, µ3) = (1,2,3), σ2 = 0.3とし,固
有角速度0.01, 0.02, . . ., 4の範囲に2万個の振動子を分布させた.(a)θi,j,1 の分布の振動子の固有角速度の分布.(b)θi,j,2の分布の振動子の固有角速 度の分布.(c)θi,j,3の分布の振動子の固有角速度の分布. . . . . 45 3.12 I(t) = sin(t) + sin(2t)の周波数分類.(µ1, µ2, µ3) = (1,2,3),σ = 0.3,Wi,l ∈
{0.01, 0.02, . . ., 4}, J = 20000とした.(a)グローバル同期度の時間経過.
µ1, µ2の振動子は同期できるが,µ3の振動子は同期できない.(b-1)µ1 = 1 の振動子から得られる出力F O1(t).(b-2)µ2 = 2の振動子から得られる出 力F O2(t).(b-3)µ3 = 3の振動子から得られる出力F O3(t). . . . . 46
3.13 振動子の固有角速度の分布の例.K = 2, M = 3,J = 20000, (µ1, µ2, µ3) = (1,2,3), σ = 0.3, ω ∈ [0.01,4]とした.(a-1, 2, 3) k = 1(左側)の振動子 がI1(t)を受け取る.(b-1, 2, 3) k= 2(右側)の振動子がI2(t)を受け取る. 47 3.14 I1(t) = sin(t) + sin(2t), I2(t) = sin(t) + sin(3t)の周期刺激を3種類の角速
度(µ1, µ2, µ3) = (1,2,3)に分類した結果.紫色はI1(t)の成分,緑色はI2(t) の成分である. . . . . 47 3.15 I1(t) = sin(t+1)+sin(2t−π/2)+sin(3t), I2(t) = sin(t)+sin(2t)+sin(3t+π)
を3種類の角速度(µ1, µ2, µ3) = (1,2,3)に分類した結果.紫色はI1(t)の成 分,緑色はI2(t)の成分である. . . . . 48
4.1 Moriyamaらが報告したオカダンゴムシの円形空間での行動[39][40].(a)
円形空間での移動の軌跡.[39]の図1より.(b)BALM仮説に基づいた縁を 歩く行動が出現するメカニズム. . . . . 49 4.2 観察容器に置かれた円形容器.(a)容器内部は平坦な地面である.(b)容器
内部は中心付近に向かって谷になっている. . . . . 50 4.3 平坦な円形容器で歩かせたオカダンゴムシの軌跡.(a-1, b-1, c-1)実験1個
体目.(a-2, b-2, c-2)実験2個体目. . . . . 51 4.4 すり鉢上の円形容器で歩かせたオカダンゴムシの軌跡.(a-1, b-1, c-1)実験
1個体目.(a-2, b-2, c-2)実験2個体目. . . . . 52
4.5 lower個体群の典型的な振る舞い.(a) 記録された軌跡(赤線),初期位置
(黄点),地形の標高(カラーマップ).(b) 軌跡から見積もった標高の時 間推移.(c) 各時間における二次元平面上での初期位置からの距離. . . 54 4.6 (a)勾配に対する感受性が強すぎる場合(α = 1)の数値計算結果(t ∈
[0,1000]).(b)勾配に対する感受性が弱すぎる場合(α= 0.001)の数値計
算結果(t∈[0,1000]).実線が移動の軌跡,赤丸が地形の山部分,青丸が
地形の谷部分を示す.数値計算には周期境界条件を用いた. . . . . 55 4.7 勾配に対する感受性をα = 0.3と設定した場合の数値計算結果(t ∈[0,
1000]).(a)移動の軌跡.赤丸が山,青丸が谷を示す.数値計算には周期
境界条件を用いた.(b)各時間での標高.(c)lower領域を選択した回数の割 合(左棒グラフ)とupper領域を選択した回数の割合(右棒グラフ).青 は生物実験により得られたlower個体群の結果,緑は数値実験で得られた 結果を示す. . . . . 55 4.8 Watanabeら[47]が用いた実験環境と実験結果. (a)オカダンゴムシは強制
転向点で強制的に右に方向転換する.その後T選択点に到達すると左(L) または右(R)のどちらかに方向転換する.Start点から強制転向点までの距 離は40mmに固定された.強制転向点からT選択点までの距離は40mm, 80mm,160mmのいづれかが使用された.全ての通路幅は8mmだった.(b) 強制転向点からT選択点までの距離と左右を選択する割合. . . . . 56 4.9 モデルダンゴムシの形状. . . . . 57 4.10 バネの減衰振動を用いて自然に直進するようなモデルダンゴムシの制御.
バネが自然長の位置にあるとき,可変抵抗の左右の抵抗値が一致する. 58 4.11 電磁石を用いて回避行動が発現するようなモデルダンゴムシの制御. . . 59 4.12 数値実験で設定したT字路.モデルダンゴムシは無限にT選択点に遭遇す
る. . . . . 60
4.13 数値実験で設定した円形空間. . . . . 61
4.14 (Aisle length) = 60に設定した場合のモデルダンゴムシの移動の様子. 63
4.15 T字路を用いた数値実験におけるモデルダンゴムシのTA発現頻度. . . 64
4.16 円形容器を用いた数値実験におけるモデルダンゴムシの移動の軌跡. . . 64 4.17 実験手順の概略.(a) 飼育ケースから無作為に選ばれたオカダンゴムシが
実験開始30分前にプラスチック容器に入れらた.(b) プラスチック容器か ら無作為に選ばれた1個体がピンセットで静かに裏返され,腹を突かれて 丸まらされた. . . . . 66 4.18 集団サイズごとの丸まる時間の箱ひげ図. . . . . 67 5.1 振り子による周波数検出の例. . . . . 69
第 1 章 はじめに
1.1 生物の知性が秘めた可能性
生物は長い期間をかけてそれぞれの生活環境に適応的に進化を繰り返してきた.その 結果,我々ヒトのような高度で複雑な神経系の集合体(脳)を獲得し,思考や判断,学 習などの脳機能を使って生活できている.近年の計算機処理能力の向上やデバイスの小 型化などに伴い,最近では人工知能が実用化されつつあり,多くの注目を集めている[1]. 実際スマートフォンへの人工知能技術による音声認識機能の搭載など,我々の生活で身 近なものとなってきている.元来人工知能はヒトの脳機能を人工的に造り出そうとする 取り組みである.脳の構造を理論的に理解し,それを実社会に応用することで高度な情 報処理が可能となる.逆に現象論的な仮説のもとで人工知能を造り出すことで,脳の妥 当なメカニズムを提示することも考えられる.脳のメカニズムの解明と知能ロボット,人 工知能技術は相乗的に発展しうるのである.
脳は神経細胞の集合体である.個々の神経細胞は電気的に興奮性の性質を持つことが 知られており,これは活動電位と呼ばれている[2][3].Hodgkinらは神経細胞の活動電位 が発生する仕組みを実験と理論(Hodgkin-Huxleyモデル)の両側面から研究し,1952年 にその全容が解明された[4][5][6][7][8].これらの成果により彼らはノーベル医学生理学賞 を受賞した.また,二つの神経細胞はシナプスと呼ばれる繊維で接続していることが知 られている.脳が学習を行う方法の一つとして,シナプスの可塑性を用いたやり方がよ く知られている[9].これは各シナプスに独立した結合強度があり,その結合強度を場所 ごとに変化させることで学習を行うというものである.特にシナプス結合強度が増すの は,シナプス前細胞とシナプス後細胞が同時に活動した時であると言われており,これ はHebb則[10]と呼ばれている.
このように1900年半ば頃から神経細胞の個々の活動やシナプス可塑性を用いた学習の 仕組みが報告されているにも関わらず,現時点ではヒトの脳の全てを説明できた研究は まだない.これはヒトが千億個以上の神経細胞を所持しており,さらにそれらが複雑な ネットワークを形成していることが要因の一つであると考えられる[11].一つの神経細 胞は1万程度の神経細胞と接続していると言われている.一方,人工知能技術はハイパ フォーマンスな計算機を用意すれば,ある程度複雑なネットワークであれば計算可能で ある.2013年にスーパーコンピュータ「京」を使って行われたヒトの脳のネットワーク を模した実験の結果では,おおよそ実際の脳活動と同じような結果が得られた[12].この ときスーパーコンピュータに実装されたシナプスは約10兆個であるというが,ヒトの千 億個のニューロンのそれぞれが1万個の接続を有していると考えた場合,その計算規模 はヒトの脳の1パーセント程度である.
人工知能技術の中でもディープラーニングは,その構造がヒトの神経細胞やシナプス といった脳の構造と似ており,すでに画像分析や自然言語処理に適用されている.ディー プラーニングでは複数の神経細胞が層ごとに配置されおり,フィードフォワードなネット ワークが形成されている.第一層は入力層,最終層は出力層と呼ばれ,その間の複数の
層はまとめて中間層と呼ばれている.特に中間層が1層しかないものはニューラルネッ トワークと呼ばれる.ディープラーニングの学習過程では複数の学習データが用意され,
それらは入力層に入れられる.それぞれの入力データは中間層を通って処理され,出力 層で出力される.この出力が学習データごとに適切に得られるようにネットワークを組 み替える.ディープラーニングにおける学習は神経細胞間のネットワークを適切に組み 替え,保持することなのである.このようにして得られた学習済みネットワークを用い ることで,未知のデータに対して過去の経験を生かした予測を出力できる.このように ディープラーニングは実際の脳のシナプス可塑性に基づいて構築されており,学習過程 も類似している.
ディープラーニングの学習の性質上,正確な予測を行うためには莫大な学習データが 必要であり,なおかつ実時間で学習が完了するような高速処理を行う必要がある.上述 した通り現時点で神経細胞ネットワークの計算はスーパーコンピュータを使った場合で さえ,ヒトの脳の1パーセント程度の規模である.これを考慮すると脳が非常に高度で かつ高速に情報処理を行っていることが実感出来る.
しかし,ディープラーニングがヒトの学習メカニズムを全て説明できているわけでは ない.例えば歩行機能を回復するために行うリハビリは「歩きたい」という脳からの命 令を末端に適切に伝えられるようにするフィードフォワードな学習であると同時に,「こ のようなリズムで足が動いている」という末端からの刺激がフィードバック的に脳まで 伝達することで運動を学習する.半身麻痺などで歩行が不可能な場合であっても,例え ばトヨタが開発した歩行支援ロボット「ウェルウォークWW-1000」(図 1.1(a))などが 実用されている[13]ように,歩行動作を繰り返すことで歩行能力が回復するといわれて いる.ディープラーニングでは神経細胞間のフィードフォワードネットワークを仮定し ていたが,実際には多様なメカニズムが複合的に働くことで適切な脳機能が得られるの である.
人工知能を発展させるメリットはヒューマンエラーがなくなることにある.ディープ ラーニングの学習によって獲得したネットワークは忘却することがないため,いつでも高 精度で正しい判断ができるのである.ヒトは環境や主観,疲労による判断力の低下など で聞き間違い,見間違いをしてしまう.図1.1(b)はチェルナー錯視と呼ばれ,斜めに引 かれた長い直線は全て平行であるが,それと交わる複数の短い直線の影響で見た目には 平行に見えない.実用的にはいつでも正確な判断ができる人工知能が有益であるが,生 物らしい情報処理においてはある程度のミスも容認されてしまうのである.
生物らしい情報処理とは,入力情報に対して動的に対応し,適応的に出力を変化させ ることだと考えている.ここで得られる出力はいつでも最適なものだけを得るのではな く,リスク回避やエネルギー効率まで考えられた出力である.例えばアメーバは餌間に 最短距離になるようなネットワークを作ることが知られている[15]が,そのネットワー クはいつでも最短距離の一本だけを残すのではなく,ネットワークが断裂された時のリ スクの軽減等まで考えられたネットワークである[16][17][18](図 1.2).このような生物 らしい情報処理を人工知能や知能ロボットに応用することで,計算効率の良い情報処理 を構築できる可能性がある.
果たして脳はどのようにして情報処理を行っているのだろうか.先に述べた通り,構 造の複雑さゆえ脳の全てを解明することは困難である.そこで我々はより下位の生物の 学習メカニズムを明らかすることにした.生物が進化の過程で高度な脳機能を獲得した とすれば,下位の生物たちはヒトの脳機能の起源を持っていると考えられる.
図 1.1: (a)トヨタの歩行支援ロボット「ウェルウォーク WW-1000」.トヨタホームペー ジより[13].(b)チェルナー錯視の例[14].
図 1.2: アメーバの餌間ネットワーク.[16]のFig.1より.
1.2 脳の起源を探る
脳の機能が複雑で高度なことは様々な場面で論じられてきた[11].それでは下位の生物 たちの知性,下位の部位での情報処理はどうだろうか.ヒトの脳機能を解明することの 困難さが注目されがちであるが,様々な生物において興味深い知性のメカニズムが知ら れている.例えば脳から運動器官への神経接続を絶たれた,いわゆる「除脳ネコ」をベ ルトコンベアに乗せると,脳からの命令が無いにも関わらず歩くようなスムーズな足運 びが見られた[19].このようなリズミカルな行動は脳による制御ではなく,筋肉等を形成 している個々の興奮性細胞自身が作り出すリズムによるものであるとされ,これは中枢 パターン生成器(central pattern generator, CPG)と呼ばれている[20].受動歩行ロボッ ト[21]は重力のみを駆動力として自然な足運びを実現したロボットであり,これは生物 の骨格形状自体に歩行のリズムが内在していることを示唆する.タコは8本の足を持つ 生物であるが,彼らが足を動かすときに脳が指示するのは「足を動かす」という命令に 留まり,決して各筋肉に細やかな指示を送ることは無いという[22].このようにヒトの脳 に限らず,様々な生物,様々な部位で知的な情報処理が行なわれているのである.
より生命の起源に近い生物,例えば単細胞生物の行動も興味深い.我々は繊毛虫テト ラヒメナにおいても記憶や学習といった情報処理が可能であることを2016年に報告した
[24].またKunitaらが2014年に報告した結果では,ゾウリムシが繰り返し危険な状況を 経験すると危機回避行動の発現頻度が増加するという[23].これらの結果から注目すべ きことは二つある.まず一つ目は単調な生活を送ってばかりいるように思われる単細胞 生物でさえ知性を持ったかのような振る舞いができることである.単細胞生物はその名 の通りたった一つの細胞から成るが,そこに内在する情報処理機構を馬鹿にはできない.
彼らの感覚受容器官,運動器官,生理化学的な構造は,多細胞生物と比較して非常に限 られているにも関わらずそのような知的な行動が可能なのである.二つ目は最近になっ て単細胞生物の新たな知的な行動が報告されたということである.実際,我々のテトラ ヒメナの記憶現象に関する報告は2016年,Kunitaらのゾウリムシの学習現象に関する報 告が2014年だった.特に単細胞生物の場合は単純な細胞生理学的な情報処理のみによる 行動であると期待できるが,そうは言っても未だに知られていない行動が存在するので ある.
本研究では,様々な生物が持つ知能に注目し,それらのメカニズムを数理モデリング によって解明することで生物らしい素朴な知能を持ったロボットを製作する.特に数理 モデリングによるメカニズムの理解が,生物の脳機能と知能ロボットの発展を連続的に 接続することを期待している.そのためこれまで生物実験系の研究者や制御系の研究者 らと協力して研究を遂行し,実験データや数理モデル,数値実験結果の共有を行ってき た.また実際に採集,飼育した生物を使って実験を行いデータを収集することで数理モ デルの作成を試みたり,作成した数理モデルを実社会に応用する例としてロボットの制 御に実装するなどしてきた.広い分野への知識と理解,技術を習得することで分野横断 的に研究を実施し,生物,数理,知能工学が相乗的に発展することを期待している.
1.3 本論文の構成
以下,本論文は次のような構成となっている.第2章では初めに単細胞の繊毛虫テト ラヒメナの円形容器記憶行動に関するKunitaの実験結果を紹介し,そのメカニズムを 記述した数理モデルを報告する[24].次にゾウリムシの正多角形容器記憶行動に関する
Bramstedtの実験結果を紹介し,そのメカニズムを記述した数理モデルを報告する.最後
に繊毛虫の記憶行動を記述した数理モデルをロボットに実装し,実機による動作実験を 行った結果を報告する.ここでのロボットへの実装と実機による実験は同研究室の矢野陽 大氏と共同で行った結果である.第3章では初めにアメーバの時間記憶に関するSaigusa, Teroらの実験結果及び数理モデルを紹介し,その数理モデルに基づいて記憶を持った位 相振動子モデルに改良した結果を報告する.次に,その位相振動子モデルを用いた入力 振動を周波数及び位相差といった特徴ごとに分類するモデルについて報告する.第4章 では初めにダンゴムシの行動選択について,地形の勾配に対する影響を調べた実験の結 果を報告し,その結果から思いつくダンゴムシの地形勾配に対する行動選択モデルを報 告する.ここでの数理モデルの構築及び数値実験は同研究室の吉峰瑠星氏と共同で行っ たものである.次にダンゴムシの交替性転向反応に関するMigita,Moriyamaらの結果を 紹介し,この行動に基づいて作成した対向二輪車のアナログ制御機構について報告する.
最後にダンゴムシが集団であることの意義を調べた実験の結果を報告する.ここでの実 験は福岡県立玄洋高等学校科学部と共同で行ったものである.
第 2 章 テトラヒメナの記憶メカニズムを 利用した知能ロボット制御の提案
2.1 繊毛虫の記憶行動
2.1.1 テトラヒメナの記憶行動と数理モデル
我々はテトラヒメナが周囲の空間形状を記憶できることを報告した[24].まずはこの文 献[24]の内容について紹介する.テトラヒメナは単細胞の繊毛虫で,体長0.05mm程度,水
中を1mm/sec程度で泳ぐ.一般に,自然な状態での遊泳軌道は直線である(図 2.1(a1)).
初めにテトラヒメナは直径0.3mm,0.4mm, 0.5mm, 0.6mmの円形容器に15分間閉じ込 められ,狭い空間を経験した.次に,狭い空間を経験した個体は直径35mmの十分広い ペトリ皿に移され,そこでの移動の軌跡が観察された.この実験によると,テトラヒメ ナは狭い空間では容器の壁に沿って泳ぎ(図 2.1(a2)),広い空間に移された後は円軌道 で泳いだ(図 2.1(a5)).さらにその円運動の直径は初めに経験した円形容器の直径の1.3 倍という関係があることがわかった(図 2.1(d)).
0.3 0.6 0.9
Diameter of circular trajectory (mm) Diameter of circular
trajectory (mm) Diameter of narrow space (mm)
Normalized frequency
0.4 0.3 0.6 0.5
0.4
0.3 0.5 0.6
0 0.5 1.0 1.5
Maximum distance (mm)
0.5
0 1.0
0.5
0 1.0
Time interval, τ (sec) 0.1mm
1mm
a1 a2 a3 a4
a5 a6
1
0 2 3
b2
0 1 2 3
b1
1
0 2 3
b3
c d
(22, 11, 76)
(30, 14, 375) (31, 13, 292) (34, 15, 757)
図 2.1: テトラヒメナの記憶現象に対するKunitaの実験結果.[24]のFig.1より.
テトラヒメナは空間の形状を記憶して行動できたのである.この記憶行動に対して,次の
ようなテトラヒメナの内部ダイナミクスが数理モデルによって提案された.テトラヒメナ を二次元空間上の点であるとみなし,時刻tでの細胞の位置を(x(t), y(t)),進行方向をθ(t) とする.この細胞は一定の速さ¯vで移動すると仮定すると,速度は( ˙x,y) = (¯˙ vcosθ,v¯sinθ) と書ける(図2.2(a1)).次に細胞が容器の壁と接触している場合を考える.細胞が壁と接触 した点における壁の角度をψ(t)∈[0,2π),壁の角度と進行方向がなす角をϕ(t)∈[0, π/2]
とおく.細胞は壁と衝突した場合,細胞は壁に沿ってスリップして動くと仮定する.すなわ ちその速度は壁方向の成分だけで運動するものとし,( ˙x,y) = (¯˙ vcosϕcosψ,v¯cosϕsinψ) とする.
図 2.2: 細胞の移動.[24]のFig.2(a)より.
ここから細胞の内部処理系と行動について考える.ここでは細胞内の化学物質の濃度 やイオン濃度,膜電位などを細胞の内部状態と呼び,細胞の進行方向は内部状態で変化 すると仮定する.時刻tにおける細胞の内部状態µ(t)をN + 1個の要素からなる実数値 関数と仮定してµ(t) = (µ0(t), µ2(t), . . . , µN(t))としたとき,細胞の進行方向の時間発展 は適当な関数fを用いて次式で表せる.
θ(t) =˙ f(µ(t)) (2.1)
また細胞の内部状態は細胞が障害物と接触した場合に変化すると仮定する.このとき細 胞の内部状態の時間発展は関数gを用いて次式で表せる.
˙
µ(t) =g(µ(t)) +δ(t)s (2.2)
ここでsはすべての成分が定数のベクトル(s= (s0, s1, . . . , sN))である.またδは0また は1の二値をとるセンサー関数で,壁との接触を感知した場合にδ= 1,そうでない場合 にδ = 0とする.自然な状態のテトラヒメナは直進を多くすることからθ˙= 0は安定な平 衡点であると考えられる.このことから細胞が障害物と衝突しないで自由に遊泳を続けた とき細胞の内部状態は平衡状態に落ち着くと仮定して,その平衡点をµ¯= (¯µ0,µ¯1, . . . ,µ¯N) とおく.ここでµ(t) =˜ µ(t)−µ¯とおいて,内部状態の各要素について平衡点の周りで線 形近似を施すと,次式を得る.
˙˜
µi(t) = ˜gi(˜µ(t)) +δ(t)si (2.3)
= αi,0µ˜0(t) +αi,1µ˜1(t) +· · ·+αi,Nµ˜N(t) +δ(t)si (2.4) ここでαi,j(i = 0,1,· · · , N)は実定数である.式(2.4)は[αi,j]を要素とする行列M を用 いると,内部状態の時間発展は次式で表現できる.
˙˜
µ(t) =Mµ(t) +˜ δ(t)s (2.5)
それぞれの内部状態が安定な平衡点を持つという仮定から,行列Mは負の実固有値また は実部が負の複素固有値を持つ.したがってµ˜に適当な変数変換を施して行列Mを対角
化し,変換後の新たな内部状態変数をU(t)=(u0(t),u1(t), · · · , uN(t)),定数ベクトルをS とすると次式を得る.
U˙(t) =
λ0 0 · · · 0 0 λ1 · · · 0 ... . .. ...
0 · · · 0 λN
U(t) +δ(t)S (2.6)
λi(i= 0,1,· · ·, N)は行列Mの固有値である.
また,方向転換の時間発展に対しても同様に線形近似を施すと次式を得る.
θ(t) = ˜˙ f(˜µ(t)) = α+
∑N i=0
biui(t) (2.7)
ここでα, b0, b1, . . . , bN は正定数である.ただしテトラヒメナは自然な状態において直線 軌道で遊泳することから,ここではα = 0とする.以上で細胞の方向転換と内部状態の 変化が得られた.
繊毛虫は細胞内にカルシウムイオンが流入することで繊毛打の逆転が起こり,進行方 向を変化させる[25].このことから,繊毛虫の進行方向を決定する細胞の内部状態U(t) をカルシウムイオンチャネルの状態を決定する化学物質の濃度とする.U(t)は実数値ベ クトルであり,平衡状態はU = 0,有限の値をとる.実際には複数の内部状態の相互作 用によって繊毛虫は行動しているが,ここからは式 (2.6)のうち減衰の遅いいくつかの内 部状態にのみ着目することにする.
ここではカルシウムイオンチャネルの開閉を決定する内部状態uiのうち,二種類の内 部状態u0,u1の減衰が遅いと仮定する.それ以外の内部状態はu0およびu1と比べて十 分速く平衡点に向かうと仮定し,方向転換には影響を及ぼさないとして無視する.した がって方向転換は次式で記述できる.
dθ
dt =b0u0+b1u1 (2.8)
繊毛虫が障害物と接触することでカルシウムイオンチャネルが開き,彼らは進行方向 を変化させる.繊毛虫は機械刺激受容性のチャネルが細胞前方部に分布していることが 知られており[26],細胞の側面部分が障害物と接触しても接触を感知できない.このこと を考慮して細胞の接触感知センサδ(ϕ)をsinϕ >sin 40◦の時δ = 1,そうでない時δ = 0 とする.このように接触感知センサに接触角に応じた閾値を設けることで,学習する円 軌道のサイズを調整することができる.
u0(t),u1(t)のうち,遅いタイムスケールで変化する内部状態をu0(t),速いタイムス ケールで変化する内部状態をu1(t)とする.接触感知センサδを用いて,遅いタイムス ケールで変化する内部状態u0のダイナミクスを次式で与える.
˙
u0 =−εu0+δs (2.9)
ここでε << 1は正定数であり,一度開いたカルシウムイオンチャネルがゆっくりと閉じ
てゆくことを表現している.sは機械刺激に対するカルシウムイオンチャネルの応答性を 表す正定数である.このようにゆっくりと定常状態u0 = 0(チャネルが閉じた状態)に 戻ってゆくことが,このダイナミクスで記憶を表現する重要なポイントである.
速いタイムスケールで変化する内部状態u1はFitzHugh-Nagumo型の興奮系のダイナ ミクスに従うと仮定し,次式で与える.
τuu˙1 =−u1(u1+a1)(u1+a2)−v+Iu0+ξu1(t) (2.10)
τvv˙ =a3u1−v+ξv(t) (2.11)
ここでτv >> τu >0,a1,a2,a3, Iはモデルパラメータであり,ξu1(t),ξv(t)は白色ノ イズである.このダイナミクスでは遅いタイムスケールで変化する内部状態u0の増加と ノイズ項の影響が興奮の引き金となるようになっている.I = 0のとき,u1はu0によら ず一定確率で発火する.I > 0のとき,u0がある程度大きい状態ではより高い確率でu1 が発火する.これによって動きの多様性を表現している.
以上のような細胞の内部ダイナミクスを仮定することで,次のようなプロセスで容器 形状の記憶が説明できる.はじめに細胞が円形容器に閉じ込められている場合,細胞が 壁との接触を続け,u0が徐々に増加してゆく.u0が増加してゆくに従って方向転換の程 度が大きくなり,やがて容器の壁との接触を感知できなくなる.接触を感知できなくな るとu0は徐々に減少し直線軌道に戻ろうとする.しかし移動の曲率半径が大きくなると 再び壁との接触を感知してしまい,u0が増加する.円形容器から取り出されるまでの間 この過程を繰り返す.最終的に容器から取り出さる時間には,細胞は円形容器の形状に ちょうど沿う内部状態となっており,さらにu0はゆっくりと定常状態に戻るため,円形 容器から取り出された後も容器の形状を保った円運動が実現する.
図 2.3,図 2.4は以下の数理モデルを数値計算した結果である.
( ˙x,y) =˙
{(¯vcosθ,v¯sinθ) (細胞が容器の壁と接触していない) (¯vcosϕcosψ,v¯cosϕsinψ) (細胞が壁と接触している)
θ˙=b0u0+b1u1
˙
u0 =−εu0+δs
τuu˙1 =−u1(u1+a1)(u1+a2)−v+Iu0+ξu1(t) τvv˙ =a3u1−v +ξv(t)
δ =
{0 (細胞が容器の壁と接触,かつsinϕ≤sin 40◦) 1 (細胞が容器の壁と接触,かつsinϕ >sin 40◦)
x軸,y軸の原点 (0,0)は円形容器の中心とする.ψは細胞と円形容器との接触点(¯x,y)¯ における壁の接線の角度ψ =π−arctan(¯x/¯y)である.ただしarctan(¯x/¯y)は(¯x,y)¯ がど の象限にあるか考慮して[0,2π)の値をとる.ϕは細胞の進行方向と壁とがなす角であり,
α = arctan(¯y/¯x)としたときϕ =π/2−arccos(cos(θ−α))である.設定したパラメータ は次の通りである;v¯= 0.2,ε= 0.0001,s = 0.003,τu = 0.0025,a1 =−0.3,a2 =−1, a3 = 4,b0 = 1,b2 = 5.数値計算の時間分割幅は∆t = 0.01とし,ξu1, ξv(t)は∆t毎に [−0.25,0.25]の範囲の一様乱数で与えた.
I = 0のとき,円形容器内部では容器の縁に沿って移動し(図 2.3(a1)),広い空間に 移された後は円運動を続けた(図2.3(a2)).容器のサイズを変えて数値計算したところ,
実験で得られた「円運動のサイズは円形容器のサイズの1.3倍」という特徴も再現された
(図 2.4).またI = 0.15とすると多様な円運動が観察された(図 2.4(b1), (b2)).
具体的には,学習する円軌道のサイズは,接触感知センサの閾値によって次のように 決定される.ここでは特に細胞が円形容器から広い空間に移される時間に着目して,広
図 2.3: 数値実験の結果.[24]のFig.3(a)(b)より.
図 2.4: 数値実験の結果.[24]のFig.3(c)より.
い空間に移される直前には半径R0の円形容器の縁に沿って移動し,広い空間に移され た直後に円形容器のA倍の半径AR0の円運動をする場合を考える.細胞が容器に閉じ 込められて十分時間が経過すると,進行方向と壁とのなす角ϕは一定の値ϕ¯とみなすこ とができる.容器内で細胞が壁に沿って移動する速度は図 2.2(a2)で与えられ,その速 さは¯v′ = ¯vcosϕ0である.いま容器の中心をOとする.細胞の位置を極座標に変換し,
(x(t), y(t)) = (R(t) cos Θ(t), R(t) sin Θ(t))としたときのΘ(t)について考える.円形容器 の半径はR0であるから,円形容器内部で壁に沿って移動する細胞について,
dΘ
dt = v¯cosϕ0 R0
(2.12) である.一方で細胞は広い空間に移された直後から,速さ¯vで半径AR0の円運動を行う.
その円運動の中心をO′とし,細胞の位置を極座標に変換して(x(t), y(t)) = (R′(t) cos Θ′(t), R′(t) sin Θ′(t))とする.円運動の半径はAR0であるから,
dΘ′ dt = v¯
AR0 (2.13)
である.容器内で壁際の細胞は( ˙x,y) = (¯˙ vcosθ,v¯sinθ)で移動しようとするが(図2.5(A- b)),実際には壁での移動は容器への射影(図2.5(A-a))として与えられる.細胞の微小 時間での移動を考えると,図2.5(A)において,容器の中心O,(A-a)の細胞,(A-b)の細 胞は一直線上にある.よって微小時間ではΘ˙ とΘ˙′は一致(図2.5(B))するので,
¯ vcosϕ0
R0 = v¯
AR0 (2.14)
ϕ0 = arccos 1
A (2.15)
を得る.以上のことから,細胞が容器内部で壁に対してϕ0 = arccos(1/A)という一定の 角度で接するとき,円形容器から出された後に半径AR0の円運動を実現できる.細胞は 接触を感知すると進行方向を変化させる.したがってϕ0は接触感知センサの閾値そのも のである.Machemerら[27]によると,テトラヒメナと同じ繊毛虫のゾウリムシが壁との 接触を感知できる域値は,おおよそ30◦から60◦の範囲にあるという.このことを考慮し て,数理モデルにおいては接触感知センサの閾値を40◦と設定した.ϕ0 = 40◦を式 (2.15) に代入し,Aについて解くとA≈1.305であり,円形容器のサイズに対して約1.3倍の円 軌道を学習することができる.
図 2.5: (A-a) 円形容器内で壁に沿った移動.(A-b) 広い空間で壁がない場合の移動.容
器の壁と進行方向とのなす角がϕ0のとき,壁に沿った移動の速さはv¯′ = ¯vcosϕ0である.
図中の(0,0)は円形容器の中心を表す.壁際でスリップする動きを仮定することで,微小 時間では円形容器の中心,細胞の位置(A-a),細胞の位置(A-b)は全て一直線上にある.
(B-a)円形容器ないで壁に沿った移動.(B-b)広い空間で壁がない場合の円運動.円形容
器の中心と円運動の中心を重ねた.
以上のようにテトラヒメナは空間形状に適応して行動できる能力を持っている.ただ し,実験で使われた完全な円形空間を自然界で見つけることは困難であり,テトラヒメ ナがそのような状況に陥ることは考え難い.またテトラヒメナの円形容器に閉じ込めた 時間は15分間であり,実験時間がテトラヒメナのライフサイクルと比較して十分短いこ とから遺伝的に空間形状の記憶能力を獲得したとは考え難い.そうであるにも関わらず,
テトラヒメナが円形空間に適応して行動できたことは,繊毛虫が潜在的に記憶能力を持っ ていることを示す結果である.この結果は元々の細胞自身が持っている内部ダイナミク スに記憶能力があることを示唆しており,細胞生理学的に重要な結果である.
2.1.2 正多角形容器におけるテトラヒメナの記憶ダイナミクスの数値実験
Kunitaらによって,テトラヒメナが円形容器の形状を記憶できたのは遅いタイムスケー
ルで開閉するカルシウムイオンチャネルの影響が大きことが示された[24].ここではテト ラヒメナが円形容器内で縁に沿った移動を続けることに注目したい.テトラヒメナにとっ て障害物との衝突を繰り返すことは大きなストレスである.実際,同文献[24]によると,
円形容器に30分間以上閉じ込められたテトラヒメナのうち,約7割の個体が動くのをや め,その場所に停止した.この「動かない」という行動は円形容器内で障害物との衝突
を避けるために取った行動戦略である.では容器に沿った運動はどのような意味を持つ だろうか.それはテトラヒメナが「動き続けることで行動範囲を広げたい」という行動 戦略と「障害物との衝突を可能な限り減らしたい」という行動戦略を折衷した結果であ り,この折衷案を実現できるダイナミクスが遅いタイムスケールで変化する内部状態な のである.
テトラヒメナの記憶ダイナミクスを様々な形状の容器に適応するとどのような行動が 観察されるだろうか.このような実験はテトラヒメナではなされていない.従ってここ では数値実験のみを用いて行動を観察する.テトラヒメナ記憶モデルと同様に細胞の位 置を(x(t), y(t)),進行方向をθ(t)とする.細胞は一定の速さv¯で移動すると仮定し,壁 と衝突していない場合,壁と衝突した場合の速度をそれぞれ( ˙x,y) = (¯˙ vcosθ,v¯sinθ), ( ˙x,y) = (¯˙ vcosϕcosψ,v¯cosϕsinψ)とする.ψは壁との衝突時の壁の角度,ϕは壁と進行 方向がなす角の大きさである.テトラヒメナの記憶ダイナミクスでは遅いタイムスケー ルと速いタイムスケールで変化する内部状態を仮定した.ここでは式 (2.6)のうち,記憶 を表現した遅いタイムスケールで変化する内部状態u0のみに着目し,それ以外の内部状 態はu0と比べて十分速く平衡状態に向かうと仮定する.これによって,ここでは次のよ うな細胞のダイナミクスについて考える.
dθ
dt =b0u0 (2.16)
du0
dt =−εu0+δs (2.17)
ここでb0, ε, sはモデルパラメータである.δは0または1の二値をとる接触感知センサで
ある.テトラヒメナの記憶ダイナミクスの場合,δに閾値を設けることで記憶する円軌道 のサイズを調整したが,ここでは容器の壁と接触している時間にδ= 1,そうでない時間 にδ = 0とする.
ここでは各パラメータの値をv¯= 100,b0 = 1,ε = 0.0001,s = 0.001とする.一辺 が800の正三角形容器に細胞を閉じ込めた場合の容器内での移動の軌跡(t ∈[1490,1500]) を図 2.6に示す.円形容器の場合と同様に初めの時間は容器の縁との接触が続き,u0の 値が増加する.ある程度u0が大きくなると容器の辺から離れられるが,そのあとすぐに 隣の辺と接触する.十分に時間が経った後は容器の各辺の中点付近との接触を繰り返し,
移動の軌跡は容器形状の内接円と等しくなる.
図 2.6: 一辺が800の正三角形容器に細胞を閉じ込めた場合に,テトラヒメナの記憶ダイ ナミクスに従って動く細胞の軌跡(t∈[1490,1500]).
細胞の移動の速さは一定であるから,十分に時間が経って軌跡が内接円を描くとき,細 胞は一定のタイミングで容器の壁と接触を繰り返す.図 2.7(a)は接触感知センサδの値
の変化である.これに対し高速フーリエ変換(FFT)を施し,周波数成分毎の振幅を調 べると図2.7(b)となった.さらに0Hzから1Hzの振動成分に注目すると図 2.7(c)となっ た.この結果を見るとδには約0.2Hzごとに一定間隔の周波数成分が含まれており,δは 確かに周期的なパルス波形に近いことがわかる.
図2.7: 正三角形容器に細胞を閉じ込めたとき,十分時間が経った後に内接円の軌跡を描 いている時間(図 2.6)での細胞が容器の壁と接触するタイミング.(a) 接触感知センサ δの値.(b) (a)のδをFFTした結果.(c) (b)の結果を拡大した図.
テトラヒメナの記憶ダイナミクスを用いると,他の正多角形容器に対しても内接円の 移動軌跡が得られる.先に述べた通り,テトラヒメナを正多角形容器に閉じ込めて移動 を観察した結果は報告されていないが,円形容器の場合と同様に行動範囲を広げ,かつ 障害物との衝突を減らすことを考えると正多角形容器内で内接円の軌道を描くことは合 理的な行動戦略だと考えられる.
2.1.3 ゾウリムシの記憶行動と数理モデル
ゾウリムシは単細胞の繊毛虫で,体長0.1mm程度,水中を1mm/sec程度で泳ぐ.ゾウ リムシの行動メカニズムについては細胞膜電位と繊毛打の関係についてよく研究されて いる.特にゾウリムシの細胞膜電位変化のダイナミクスが,多細胞生物の神経細胞の膜電 位変化のダイナミクスを記述したHodgkin-Huxleyモデルを用いて説明された[28]ことか ら,泳ぐ神経細胞とも呼ばれている[25].ゾウリムシにおいては1935年に空間記憶の能 力があることがBramstedtによって報告され[29],1951年に小野によって追実験が行わ
れた[30].これらの結果は以下の通りである.円形,正三角形,正方形の容器を用意し,
初めにそのいずれかの形状の容器にゾウリムシを閉じ込めた.2時間後,初めの容器の形 状とは異なる容器にゾウリムシを閉じ込め,その空間での移動の軌跡を観察した.その
結果,正方形から正三角形に移した場合,及び正三角形から正方形に移した場合には特 徴的な行動は見られなかったが,それ以外の場合では初めに泳いだ容器の形状と同じ形 状の移動軌跡が得られた(表 2.1).ただし,初めの容器のサイズと移動後の軌跡のサイ ズは相関がなかった.
表 2.1: ゾウリムシの空間記憶実験[29][30].
Shape of the 1st vessel Shape of the 2nd vessel Trajectory in the 2nd vessel
circle triangle circle
circle square circle
triangle circle triangle
triangle square (no features)
square circle square
square triangle (no features)
ゾウリムシの空間記憶現象に対して,2010年に手老,秋山によって数理モデルが提案 された[31].これに基づいて以下のような数理モデルを考える.
ゾウリムシもテトラヒメナと同様に,式(2.6)に従う行動ダイナミクスを持つと仮定す る.初めに円形容器を経験した個体は2番目の容器の形状に関わらず円軌道で移動する.
従って,テトラヒメナの記憶ダイナミクスを用いてゾウリムシの円形容器の記憶現象も 説明することが可能である.しかし,正多角形容器の形状記憶をテトラヒメナの記憶ダ イナミクスだけで説明することはできない.単細胞生物のような脳を持たない生物の知 的な行動は,生物に内在する多様なリズムに注目することが有効な手段である[32].そ こで式 (2.6)のうち,減衰の遅い内部状態をuiとする.u0を式 (2.17)に従う内部状態,
ui(i≥2)をλiが実数の内部状態とする.ui以外の内部状態は,uiと比較して減衰が十分 速いとして無視する.
さらにここではuiがu0 ̸= 0のとき振動するという性質を仮定し,その振動の角速度は u0の状態によって変化すると仮定する.uiおよびθの時間発展を次式で与える.
dui
dt =−εiui+ωiu0vi (i= 1,2, . . .) (2.18) dvi
dt =−ωiu0ui−εivi+δsi (i= 1,2, . . .) (2.19) dθ
dt =u0+b1u1 +b2u2+. . . (2.20)
εi,ωi,si,biは正定数のモデルパラメータである.上式はu0vi,u0uiの非線形項を含ん でおり,式 (2.6)の形式になっていない.平衡点ui = 0,vi = 0周りで線形化し,式(2.6) の形式に変換するとλiは全て実数であることがわかる.しかし,u0の変化が他の内部状 態と比較して十分遅いとき,uiは減衰振動とみなすことができる.このときu0 = ¯u0(定 数)とすると,uiの振動の角速度はωiu¯0である.
Kunitaらのテトラヒメナの実験において,円形容器に閉じ込めた時間は15分間だった.
これに対し,小野がゾウリムシを種々の形状の容器に閉じ込めた時間は2時間だった.こ のことを考慮すると,容器に閉じ込められたゾウリムシの内部ダイナミクスは,まず初 めにu0が内接円を描く内部状態に到達し,その後uiの内部リズムが行動に影響を及ぼす と考えられる.
ここから正N 角形の容器にゾウリムシを閉じ込めた場合を考える.ゾウリムシが内接 円周上を移動している状態でu0 = ¯u0(定数),ui = 0とすると,細胞の方向転換はθ˙ = ¯u0 である.このときuiは角速度ωiu¯0で減衰振動する内部状態である.ゾウリムシがθ˙ = ¯u0 によって容器の内接円周上を移動しているとき,ゾウリムシは壁と周期的に接触するた め,機械刺激δsiは周期的な関数となる.この周期的な機械刺激によって減衰振動する内 部状態uiのうち,機械刺激の角速度と同じ固有角速度か,その定数倍の固有角速度を持 つ内部状態だけが共振するため,共振できない内部状態は定常状態にあるとみなすこと ができる.細胞が内接円周上を移動する角速度はu¯0であり,一周する間に正N角形容器 の各辺に1回ずつ,計N回衝突することから,機械刺激の角速度はNu¯0である.よって 式(2.18)および式(2.19)のうち,ωiu¯0がNu¯0の定数倍,すなわちωiがNの定数倍であ る内部状態のみが方向転換に影響を及ぼし,それ以外は無視できると仮定する.
dui
dt =−εiui+iN u0vi (2.21)
dvi
dt =−iN u0ui−εivi+δsi (2.22) ここからさらに,i≥2の内部状態はu0およびu1と比較して十分速く平衡点に向かうと 仮定すると,次式を得る.
du1
dt =−ε1u1+N u0v1 (2.23)
dv1
dt =−N u0u1−ε1v1+δs1 (2.24) また方向転換を次式で与える.
dθ
dt =u0+u1 (2.25)
図 2.8は正三角形容器に対して,以下の数理モデルを数値計算した結果である.
˙
u0 =−εu0+δs
˙
u1 =−ε1u1+N u0v1
˙
v1 =−N u0u1−ε1v1+δs1 θ˙=u0 +u1
設定したパラメータは次の通りである;v¯= 100,ε1 = 0.000001,s1 = 0.002,N = 3.正 三角形容器に閉じ込められた直後から移動の軌跡は内接円に近づいてゆく.軌跡が内接 円に近くなると周期的に壁と衝突するようになり,それに伴って内部リズムが共振する.
内部で共振したリズムの影響を受けて移動の軌跡が正三角形に近づいてゆく.t = 5000 において容器の壁が取り除かれたが,容器内で学習した方向転換を継続した.
同様に一辺の長さが800の正方形に対して,N = 4とした場合の数値計算結果を図2.9 に示す.正三角形の場合と同様に,正方形容器を記憶できた.
ゾウリムシの正N角形記憶モデルが形状を学習できた理由について考察する.正三角 形容器(N = 3)に対して,十分時間がたった状態で内部状態はu0 = ¯u0(定数),u1 = acos 3¯u0t(aは定数)と書ける.よってこのとき,方向転換はθ˙ = ¯u0 +acos 3¯u0tであ る.θ˙が変化することは,車のハンドルを切ることに相当する.θ˙ = ¯u0はハンドルを一 定の大きさだけ切った状態で保っている状態であり,移動軌跡の曲率半径は一定である.
またθ˙=acos 3¯u0tは左右交互に同じ大きさだけハンドルを切っては戻すという状態であ
![図 1.1: (a) トヨタの歩行支援ロボット「ウェルウォーク WW-1000 」.トヨタホームペー ジより [13] . (b) チェルナー錯視の例 [14] . 図 1.2: アメーバの餌間ネットワーク. [16] の Fig.1 より. 1.2 脳の起源を探る 脳の機能が複雑で高度なことは様々な場面で論じられてきた [11] .それでは下位の生物 たちの知性,下位の部位での情報処理はどうだろうか.ヒトの脳機能を解明することの 困難さが注目されがちであるが,様々な生物において興味深い知性のメカニズムが知](https://thumb-ap.123doks.com/thumbv2/123deta/9916116.1918502/11.892.116.346.114.371/ロボットウェルウォークトヨタホームペーチェルナー.webp)
![図 2.3: 数値実験の結果. [24] の Fig.3(a)(b) より. 図 2.4: 数値実験の結果.[24] の Fig.3(c) より. い空間に移される直前には半径 R 0 の円形容器の縁に沿って移動し,広い空間に移され た直後に円形容器の A 倍の半径 AR 0 の円運動をする場合を考える.細胞が容器に閉じ 込められて十分時間が経過すると,進行方向と壁とのなす角 ϕ は一定の値 ϕ ¯ とみなすこ とができる.容器内で細胞が壁に沿って移動する速度は図 2.2(a2) で与えられ,その速 さは](https://thumb-ap.123doks.com/thumbv2/123deta/9916116.1918502/17.892.153.662.117.230/数値実験結果より図数値実験結果考える¯とみなすできる.webp)
![表 2.1: ゾウリムシの空間記憶実験 [29][30] .](https://thumb-ap.123doks.com/thumbv2/123deta/9916116.1918502/21.892.91.722.243.440/表21ゾウリムシの空間記憶実験293.webp)

![図 2.12: ルンバのポート説明 [34] . またルンバのシリアルポートに “Opcode:137” を送信することでルンバは Drive モード となり,ルンバは移動の速さと曲率半径が入力されるのを待つが,このとき 1 バイトず つのデータを送信しなければならないことに注意する.Drive モードは 4 バイトのデータ を受け付けるので,具体的には (i) ルンバの速さの上位バイト, (ii) ルンバの速さの下位 バイト, (iii) 曲率半径の上位バイト, (iv) 曲率半径の下位バイトの順に送信す](https://thumb-ap.123doks.com/thumbv2/123deta/9916116.1918502/26.892.227.745.115.447/ルンバポートルンバシリアルポートルンバモードとなりルンバ.webp)