学
位 論 文
A study of rBC2LCN as a novel cancer stem cell marker of oral squamous cell carcinoma cells
( 口 腔 扁 平 上 皮 癌 細 胞 の 新 た な 癌 幹 細 胞 マ ー カ ー と し て の 糖
鎖 認 識
レク チン rBC2LCN に 関す る研究 )
学 位 申 請 者 中 峠 洋 隆
広 島 大 学 大 学 院 医 歯 薬 保 健 学 研 究 科 医 歯 薬 学 専 攻
主 指 導 教 員 : 岡 本 哲 治 教 授
( 応 用 生 命 科 学 部 門 分 子 口 腔 医 学 顎 顔 面 外 科 学 )
平 成 28 年 度
1 第 1 章 緒 言 癌 組 織 は 一 様 な 細 胞 集 団 で は な く , 多 様 な 細 胞 集 団 か ら 構 成 さ れ , そ の 中 で も 細 胞 周 期 の 遅 い 細 胞 群 は , ご く 少 数 で も 癌 を 生 体 内 に 再 構 築 で き ,放 射 線 療 法 や 化 学 療 法 に 対 し て 抵 抗 性 を 示 す こ と か ら , 癌 に お け る 幹 細 胞( 癌 幹 細 胞 )で あ る 可 能 性 が 示 唆 さ れ て い た (Reya, et al, 2001). 近 年 , 癌 は 幹 細 胞 能 力 と 癌 形 成 能 を 併 せ 持 つ 少 数 の 癌 細 胞 群 と , 同 細 胞 に 由 来 し た 幹 細 胞 の 特 性 を 持 た な い 細 胞 群 に よ り 形 成 , 維 持 さ れ て い る こ と が 示 さ れ , 癌 幹 細 胞 は 癌 治 療 に お け る 新 た な 標 的 細 胞 と し て 注 目 さ れ て き て い る (Tan, et al, 2006; Ailles, et al, 2007; Kvinlaug, et al, 2007; Lobo, et al, 2007) .
癌 幹 細 胞 の 特 徴 と し て 正 常 幹 細 胞 と 同 様 に , ① 未 分 化 な 表 面 形 質 を 示 す (Gilbert, et al, 2009), ② 自 己 複 製 能 に よ り 癌 幹 細 胞 集 団 を 維 持 す る (Schatton, et al, 2009) , ③ 多 分 化 能 を 有 し 多 様 な 癌 細 胞 に 分 化 す る (Gupta PB, et al, 2009) , ④ 高 い 薬 剤 耐 性 を 示 す ( Hermann, et al, 2007), ⑤ 高 い 腫 瘍 形 成 能 を 示 す ( Folkins, et al, 2009), な ど が あ げ ら れ て い る . そ の た め , 癌 幹 細 胞 は 癌 の 発 生 ・ 進 展 の み な ら ず 再 発 , 転 移 お よ び 治 療 抵 抗 性 に も 深 く 関 与 し て い る と 考 え ら れ て い る (Dalerba, et al, 2007; Trumpp, et al, 2008; Visvader, et al, 2008; Jones, et al, 2009) .
癌 幹 細 胞 研 究 に お い て は , 他 の 細 胞 集 団 か ら 幹 細 胞 群 を 正 確 に 分 離 ・ 同 定 す る こ と が 重 要 で あ る . 現 在 ま で に 癌 幹 細 胞 の 特 徴 を 利 用 し た 色 々 な 分 離 方 法 が 報 告 さ れ て い る . ま ず , 正 常 幹 細 胞 の 機 能 的 特 徴 を 利 用 し た 方 法 と し て , ABC ト ラ ン ス ポ ー タ ー 分 子 ( Zhou, et al, 2001; Huls, et al, 2008) に よ る 色 素 や 薬 剤 排 出 能 力 を 利 用 し た 方 法
2
で あ る . つ ま り 幹 細 胞 で は , そ の 生 存 の た め 非 幹 細 胞 と 比 較 し て ABC ト ラ ン ス ポ ー タ ー が 高 発 現 さ れ て い る た め , DNA 結 合 蛍 光 色 素 Hoechst33342 ( Goodell, et al, 1996) で 処 理 す る と , ABC ト ラ ン ス ポ ー タ ー 高 発 現 細 胞 は Hoechst33342 を 排 出 す る と い う 染 色 抵 抗 性 を 利 用 し た も の で , UV 光 で 励 起 す る と 大 部 分 の Hoechst33342 に よ る 蛍 光 強 度 の 高 い 細 胞 集 団( main population)よ り も 蛍 光 強 度 の 低 い 細 胞 群 つ ま り side population (SP)細 胞 群 が フ ロ ー サ イ ト メ ト リ ー で 分 離 さ れ る が , こ の SP 細 胞 群 に は 高 頻 度 に 癌 幹 細 胞 が 存 在 す る と い わ れ , 乳 癌 , 肺 癌 お よ び 脳 腫 瘍 で が ん 幹 細 胞 と し て 報 告 さ れ て い る (Hirschmann-Jax, et al, 2004; Kondo, et al, 2004; Hadnagy, et al, 2006; Ho, et al, 2007; Wu, et al, 2008; Charafe -Jauffret, et al, 2009).次 に ,細 胞 表 面 マ ー カ ー を 利 用 し て 分 離 す る 方 法 が あ る .急 性 骨 髄 性 白 血 病 ( AML) の 癌 幹 細 胞 が CD34 陽 性 /CD38 陰 性 細 胞 群 に よ り 濃 縮 さ れ る こ と が 報 告 さ れ て (Lapidot, et al, 1994)以 来 , 色 々 な 分 子 が 癌 幹 細 胞 で 特 異 的 に 発 現 す る 細 胞 表 面 マ ー カ ー と し て 検 討 さ れ て い る ( Deonarain, et al, 2009). こ の よ う な 癌 幹 細 胞 表 面 マ ー カ ー の 発 現 パ タ ー ン の 同 定 は , 癌 幹 細 胞 を 検 出 す る 有 効 な 手 段 お よ び 抗 体 治 療 の 標 的 と な る と 考 え ら れ , 最 近 最 も 広 く 認 識 さ れ て い る 細 胞 表 面 マ ー カ ー と し て は ,組 織 幹 細 胞 マ ー カ ー で あ る CD44,CD133( prominin 1), CD34 等 が 有 力 で あ る ( Major et al. 2013) 1875 年 ,Cohnheim は 癌 幹 細 胞 が 組 織 幹 細 胞 か ら 発 生 す る と い う 説( コ ー ン ハ イ ム 仮 説 ) を す で に 提 唱 し て お り , 正 常 幹 細 胞 と 癌 幹 細 胞 の 関 係 の 重 要 性 を 示 唆 し た .
近 年 induced pluripotent stem (iPS)細 胞 に 代 表 さ れ る よ う な , 幹 細 胞 研 究 の 進 展 に よ り , 多 能 性 幹 細 胞 に 関 連 し た 知 見 が 集 積 さ れ て き て
3
お り , 多 能 性 幹 細 胞 と し て の 特 徴 を 備 え た 細 胞 が 癌 幹 細 胞 と な り 得 る 可 能 性 , す な わ ち 共 通 性 が 考 え ら れ て い る .( Lessard,J ら 2003 , Barker ら 2009)
未 分 化 多 能 性 幹 細 胞 マ ー カ ー と し て SSEA-3 , SSEA-4 , TRA-1-60 , TRA-1-81 が 用 い ら れ て い る が こ れ ら の エ ピ ト ー プ は 糖 鎖 で あ り ,糖 鎖 構 造 の 特 異 性 を 認 識 す る 抗 体 が 広 く 用 い ら れ , 組 織 幹 細 胞 マ ー カ ー と は 性 質 が 異 な っ て い る .( Adewumi ら 2007,Wright ら 2009)特 定 の 糖 鎖 構 造 を 認 識 結 合 す る タ ン パ ク 質 の 総 称 と し て レ ク チ ン が 存 在 し , こ れ ら , 未 分 化 多 能 性 幹 細 胞 の 表 面 上 の 特 異 的 な 糖 鎖 構 造 を 認 識 , 結 合 で き る レ ク チ ン の 存 在 が 想 定 さ れ た . 2013 年 , レ ク チ ン ア レ イ よ り Burkholderia cenocepacia (バ ー ク ホ ル デ リ ア ・ セ ノ セ パ シ ア 菌 )に 由 来 す る リ コ ン ビ ナ ン ト レ ク チ ン rBC2LCN は こ の 未 分 化 多 能 性 幹 細 胞 の 特 異 的 な 糖 鎖 構 造 を 認 識 し , か つ 分 化 し た 体 細 胞 で は 糖 鎖 構 造 の 変 化 か ら そ の 結 合 は 失 わ れ る 事 が 報 告 さ れ た .( Tateno ら 2013) さ ら に , 蛍 光 色 を 結 合 さ せ た rBC2LCN を 用 い る 事 で , 未 分 化 多 能 性 幹 細 胞 を 生 存 状 態 の ま ま で 標 識 す る こ と や , 再 生 医 療 に 腫 瘍 化 の 問 題 と な る 未 分 化 多 能 性 幹 細 胞 の 残 存 を 効 果 的 に 検 出 ・ 除 去 す る 事 へ の 有 効 性 が 示 さ れ た .( Tateno ら 2013)こ の こ と か ら ,未 分 化 多 能 性 幹 細 胞 は rBC2LCN 認 識 糖 鎖 を 発 現 し て い る が , 一 方 で 癌 細 胞 集 団 内 に お い て そ の 糖 鎖 発 現 細 胞 が 存 在 す る の か , ま た , 発 現 細 胞 が 存 在 す る 場 合 , 未 分 化 多 能 性 幹 細 胞 の 持 つ 特 徴 を 有 す る 細 胞 で あ る の か , 癌 幹 細 胞 と し て の 特 徴 を 有 す る か 否 か は 一 切 報 告 が な い . 本 研 究 で は , ヒ ト 口 腔 扁 平 上 皮 癌 (OSCC)に お け る 癌 幹 細 胞 を 標 的 と し た 新 し い 診 断 ・ 治 療 法 を 開 発 す る こ と を 目 指 し , 未 分 化 多 能 性 幹 細
4
胞 の 細 胞 表 面 上 に 発 現 す る 特 異 的 な 糖 鎖 構 造 を 認 識 す る レ ク チ ン で あ る rBC2LCN を 利 用 し , ヒ ト 口 腔 扁 平 上 皮 癌 由 来 細 胞 株 よ り 分 離 し た rBC2LCN 認 識 糖 鎖 陽 性 細 胞 群 の 細 胞 ・ 分 子 生 物 学 的 特 性 の 解 析 を 行 っ た .
5
第 2 章 材 料 と 方 法
第 1 節 細 胞 培 養 法 と 細 胞 の 調 整
1 ) 基 礎 栄 養 培 地 の 調 整
DMEM (Sigma-Aldrich) と Nutrient Mixture Ham F-12 (Sigma-Aldrich) (Ham, 1963) を 1: 1 に 混 合 し た 培 地 を Milli-Q 水 に 溶 解 し , 90mg/L ペ ニ シ リ ン G ナ ト リ ウ ム , 90mg/L カ ナ マ イ シ ン , 165mg/L ピ ル ビ ン 酸 ナ ト リ ウ ム , 20mM N-2 ヒ ド ロ キ シ エ チ ル ピ ペ ラ ジ ン -N-2-エ タ ン ス ル ホ ン 酸 (HEPES)及 び 2.0g/L 重 炭 酸 ナ ト リ ウ ム を 添 加 後 ,pH. 7.4 に 調 整 し ,PVDF メ ン ブ レ ン フ ィ ル タ ー (孔 径 0.25μ m) で 濾 過 滅 菌 し , DF 培 地 と し た (Barnes and Sato, 1980; Darmon, et al, 1981).
2 ) 血 清 添 加 培 地 の 調 整
DF 基 礎 栄 養 培 地 に 5%ウ シ 血 清 ( SH30072.03, AQC23417, Hyclone) を 添 加 し , DF5%CS 培 地 と し た .
3 ) 無 血 清 培 地 の 調 整
DF 基 礎 栄 養 培 地 に ,human transferrin (5μ g/ml) (Sigma-Aldrich), ヒ ト リ コ ン ビ ナ ン ト insulin (10 μ g/ml) (0105, KSK) , 10µM 2-mercaptoethanol, 10µM 2-aminoethanol, 10nM sodium selenite (以 上 , 片 山 化 学 , 大 阪 ), oleic acid (4μ g/ml)( oleic acid と fatty acid-free bovine serum albumin を 2:1 の モ ル 比 で 混 合 さ せ た FAF-BSA-oleic)(以 上 Sigma-Aldrich)の 6 種 類 の 因 子 (6F) を 加 え た
6
無 血 清 培 地 を , DF6F 培 地 と し た (Sato, et al, 1987; Myoken, et al, 1989).
4 ) 細 胞 株 と 培 養 方 法
OSCC 由 来 細 胞 株 と し て ,著 者 の 所 属 す る 研 究 室 で 樹 立 し , 細 胞 バ ン ク に 登 録 さ れ て い る 以 下 の 2 細 胞 株 を 用 い た . UE (HO-1-U-1) 細 胞 株 ( Miyauchi, et al, 1985) , NA (HO-1-N-1) 細 胞 株 ( Miyauchi, et al, 1988), を 用 い た . 扁 平 上 皮 細 胞 株 と し て A431 外 陰 部 由 来 扁 平 上 皮 癌 細 胞 株 (Giard DJ, et al. 1973)を 用 い た . こ れ ら の 細 胞 株 を DF6F 培 地 に て , 100mm 径 培 養 皿 ( Beckton Dickinson Falcon, USA) 上 で , 37℃ , 5%CO2気 相 下 で 培 養 し た .継 代 培 養 は 各 細 胞 の 増 殖 飽 和 前 に 以 下 の 方 法 で 行 っ た . 細 胞 を 0.05% trypsin (Sigma-Aldrich)と 0.04% エ チ レ ン ジ ア ミ ン 四 酢 酸 二 ナ ト リ ウ ム (EDTA ; 同 仁 化 学 ) を 含 む Dulbecco’ s Ca2 +, Mg2 +-free phosphate-buffer saline (PBS(-))
(Trypsin/EDTA) で 分 散 後 , trypsin inhibitor (Sigma-Aldrich) で trypsin を 中 和 後 , 継 代 培 養 を 行 っ た . 第 2 節 各 細 胞 株 に お け る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 群 の 存 在 の 有 無 の 検 討 1 )rBC2LCN-FITC に よ る フ ロ ー サ イ ト メ ト リ ー お よ び セ ル ソ ー テ ィ ン グ 各 細 胞 株 を 増 殖 飽 和 前 ま で 培 養 し , 継 代 培 養 法 に 準 じ て Trypsin/EDTA に て 細 胞 を 分 散 後 , Trypsin inhibitor で 中 和 し , 単 一 細 胞 と し た . こ れ ら の 細 胞 を Running buffer に 浮 遊 さ せ た . Running
7
buffer は PBS(-)に 0.5% BSA と EDTA(2mM)を 加 え 作 製 し た . 続 い て , 40µm 径 の フ ィ ル タ ー を 通 過 さ せ た 細 胞 懸 濁 液 に 1:100 の 濃 度 で rBC2LCN-FITC( Wako Japan) 添 加 し , 1 時 間 室 温 で イ ン キ ュ ベ ー ト し た .生 細 胞 と 死 細 胞 を 判 定 す る た め propidium iodine( PI)( Invitrogen USA)を 2µg/ml で 添 加 し 30 分 室 温 で イ ン キ ュ ベ ー ト し た .次 に , こ の 細 胞 懸 濁 液 を SH800 セ ル ソ ー タ ー ( SONY Japan) に て フ ロ ー サ イ ト メ ト リ ー 解 析 及 び , セ ル ソ ー テ ィ ン グ を 行 い , こ れ ら 癌 細 胞 株 に お い て も rBC2LCN が 認 識 す る 糖 鎖 を 発 現 し て い る か 否 か を 検 討 し た . 2 ) rBC2LCN-FITC を 用 い た ラ イ ブ セ ル イ メ ー ジ ン グ セ ル ソ ー テ ィ ン グ に て , rBC2LCN 認 識 糖 鎖 陽 性 , 及 び 陰 性 細 胞 を 分 離 後 , そ れ ぞ れ 6cm 径 細 胞 培 養 皿 ( Beckton Dickinson Falcon, USA ) に DF6F 無 血 清 培 地 を 用 い て 播 種 し , 約 60% コ ン フ ル エ ン ト 状 態 ま で 37℃ , 5%CO2 気 相 下 で 培 養 し た . 続 い て , rBC2LCN-FITC( Wako Japan)
を 1:1000 の 濃 度 で 加 え た DF6F 培 地 に 交 換 し , 1 時 間 37℃ , 5%CO2気
相 下 で 培 養 後 , 共 焦 点 レ ー ザ ー 顕 微 鏡 ( Carl Zeiss, LSM700, Jena, Germany) を 用 い て 観 察 し た .
3 ) rBC2LCN 認 識 糖 鎖 陽 性 及 び 陰 性 細 胞 の 無 血 清 単 層 培 養 系 に お け る 細 胞 増 殖 能 の 検 討
セ ル ソ ー テ ィ ン グ に よ り , rBC2LCN 認 識 糖 鎖 陽 性 及 び 陰 性 細 胞 を 分 離 し ,24 穴 マ ル チ プ レ ー ト( 16mm 径 )( Beckton Dickinson Falcon USA) に , 各 ウ ェ ル あ た り 2×103/ml の 細 胞 密 度 で DF6 F 無 血 清 培 地 を 用 い
て 播 種 し 無 血 清 単 層 培 養 を 行 い , 6 日 間 継 日 的 に コ ー ル タ ー カ ウ ン タ ー に て 細 胞 数 を 計 測 し , 細 胞 増 殖 能 を 検 討 し た .
8
4 ) rBC2LCN 認 識 糖 鎖 陽 性 及 び 陰 性 細 胞 の 無 血 清 浮 遊 培 養 系 に お け る 浮 遊 細 胞 塊 ( sphere) 形 成 能 の 比 較
セ ル ソ ー テ ィ ン グ で 分 離 し た rBC2LCN 認 識 糖 鎖 陽 性 及 び 陰 性 細 胞 を , 超 親 水 性 処 理 低 吸 着 性 細 胞 培 養 皿 Prime Surface (Sumitomo Bakelite Japan) (35mm 径 )上 に 5.0×102/ml の 細 胞 密 度 で 2 ml の DF6F を 用 い
て 播 種 し 培 養 し た . 播 種 直 後 に 全 て の 細 胞 が 単 一 で あ る こ と を 確 認 し た . 1 ,3 ,5 ,7 日 後 に 位 相 差 顕 微 鏡 ECLIPSE TE300( Nikon, Japan) 下 で sphere 数 お よ び sphere 形 態 を 計 測 ・ 評 価 し た . さ ら に 2 週 間 後 に rBC2LCN-FITC( Wako Japan) を 1:1000 の 濃 度 で 加 え た DF6F 培 地 に 交 換 し , 1 時 間 37℃ , 5%CO2 気 相 下 で 培 養 後 , 共 焦 点 レ ー ザ ー 顕 微 鏡 ( Carl Zeiss)を 用 い て sphere 細 胞 塊 中 に お け る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 の 存 在 様 態 を 観 察 し た . 第 3 節 DNA マ イ ク ロ ア レ イ に よ る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 に お け る 遺 伝 子 発 現 の 解 析 1 ) Total RNA の 抽 出 と 定 量 Ho-1-U-1 細 胞 株 よ り セ ル ソ ー タ ー を 用 い て rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 を そ れ ぞ れ 分 離 後 ,6cm 径 細 胞 培 養 皿( Beckton Dickinson Falcon) に 播 種 し DF6F 無 血 清 培 養 に て 増 殖 飽 和 前 ま で 増 殖 さ せ た . Total RNA は Trizol regent (Invitrogen USA) を 用 い て 抽 出 し , 分 光 光 度 計( NanoDrop ND-1000 Spectrophotometer,㈱ エ ル・エ ス・エ ス , 東 京 ) に て total RNA 量 を 定 量 し た . さ ら に バ イ オ ア ナ ラ イ ザ ー
9
( Agilent USA) を 用 い て total RNA integrity number( 分 解 程 度 ) を 確 認 ・ 評 価 し た .
2 )cRNA の ラ ベ ル 化 ,ハ イ ブ リ ダ イ ゼ ー シ ョ ン ,ス キ ャ ン 及 び 定 量 化 Agilent ラ ベ ル 化 キ ッ ト ( Agilent USA) を 用 い て ラ ベ ル 化 cRNA を 合 成 し , 56℃ , 17 時 間 ハ イ ブ リ ダ イ ズ さ せ た . ス キ ャ ニ ン グ と 定 量 化 は agilent ス キ ャ ナ を 用 い て 行 っ た ( Agilent USA). 得 ら れ た デ ー タ は Excel( Microsoft USA) に て 解 析 し た .
第 4 節 rBC2LCN 認 識 糖 鎖 陽 性 及 び 陰 性 細 胞 に お け る IGF-2 発 現 の 定 量 と そ の 機 能 評 価
1 ) Droplet digital PCR 法 を 用 い た rBC2LCN 認 識 糖 鎖 陽 性 細 胞 お よ び 陰 性 細 胞 に お け る IGF-2 mRNA 発 現 量 の 定 量
第 3 節 ( 1 ) に 準 じ て 精 製 し た total RNA を テ ン プ レ ー ト に , SuperScript® VILO( Invitrogen USA) を 用 い て cDNA を 合 成 し た . 得 ら れ た cDNA を 1:100 に Nuclease-Free water で 希 釈 し , QX100™ Droplet Digital™ PCR System (Bio-Rad US)を 用 い た droplet digital PCR 法 に て IGF-2 mRNA の 発 現 を 絶 対 定 量 し た . Droplet digital PCR は 以 下 の 方 法 で 行 っ た . QX100 droplet generator ( Bio-Rad USA) に よ り お よ そ 2 万 個 の droplet (21 l)に cDNA, IGF-2 特 異 配 列 プ ラ イ マ ー ( F: GCTGGCAGAGGAGTGTCC R:GATTCCCATTGGTGTCTGGA), ユ ニ バ ー サ ル プ ロ ー ブ #10( Roche Germany), 2XddPCR master mix( Bio-Rad USA) を 分 配 し た . 続 い て , C1000 Touch™ Thermal Cycler ( Bio-Rad USA)
10
に よ り ,デ ィ ネ イ チ ャ ー 94℃ , 30 秒 ,ア ニ ー リ ン グ 及 び 伸 長 反 応 54℃ , 1 分 30 秒 の サ イ ク ル を 40 サ イ ク ル 行 い ,droplet 中 で PCR 反 応 を 行 っ た . 反 応 後 の droplet は , QX100 droplet リ ー ダ ー に て ポ ジ テ ィ ブ droplet と ネ ガ テ ィ ブ droplet 数 を 計 測 す る こ と で , IGF-2 mRNA の コ ピ ー 数 を 定 量 し た . 対 照 と し て , Universal Probe Library Human GAPD Gene Assay( Roche Germany) キ ッ ト を 用 い て GAPDH mRNA コ ピ ー 数 を 測 定 し , IGF-2 mRNA コ ピ ー 数 ÷GAPDH mRNA コ ピ ー 数 に よ り , IGF-2 の relative expression を 求 め た . 2 ) ウ エ ス タ ン ブ ロ ッ ト 法 に よ る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 お よ び 陰 性 細 胞 に お け る IGF-2 発 現 の 検 討 セ ル ソ ー テ ィ ン グ に よ っ て 分 離 し た rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 を そ れ ぞ れ 60mm 径 細 胞 培 養 皿 に DF6F 無 血 清 培 地 を 用 い て 播 種 し 増 殖 飽 和 前 ま で 培 養 後 に , PBS(-)に て 3 回 洗 浄 し RIPA バ ッ フ ァ ー ( 20mM Tris-HCl,150mM NaCl,1mM EDTA,1% TritonX-100, pH7.4 ) に protease inhibitor cocktail P8340( Sigma-Aldrich USA) を 1:100 濃 度 で 添 加 し た cell lysis buffer で 細 胞 を 回 収 し た . 続 い て ,遠 心 (13,000g, 10 分 間 ) 後 上 清 を 回 収 し , Pierce BCA Protein Assay Kit(Thermo Fisher Scientific USA)に て タ ン パ ク 量 を 定 量 し た . 次 に , 10µg の 各 サ ン プ ル を 12% SDS-PAGE 法 に て 電 気 泳 動 後 ,PVDF メ ン ブ レ ン ( Bio-Rad USA) 上 に 転 写 し た . メ ン ブ レ ン を 5 % ス キ ム ミ ル ク ( 雪 印 , 群 馬 ) に て 室 温 30 分 浸 漬 し ブ ロ ッ キ ン グ を 行 っ た . 一 次 抗 体 と し て 抗 IGF-2 抗 体 0.125µg/ml( ab9574 rabbit polyclonal, abcam ), 抗 β -actin 抗 体 0.5µg/ml( 017-24551 mouse monoclonal, Wako) を
11
TBS-T wash buffer( 0.1% Tween20) に て 希 釈 し , 4℃ 下 overnight 処 理 し た . メ ン ブ レ ン を TBS-T wash buffer に て 5 分 間 , 3 回 洗 浄 後 , HRP ラ ベ ル 化 抗 ウ サ ギ IgG 抗 体 57.5ng/ml( GTX221666-01 GeneTex)及 び HRP ラ ベ ル 化 マ ウ ス IgG 抗 体 0.2µg/ml( GTX221667-01 GeneTex) を TBS-T wash buffer に て 希 釈 し 室 温 下 30 分 間 処 理 し た . メ ン ブ レ ン を TBS-T wash buffer に て 5 分 間 , 3 回 洗 浄 後 , ECL 試 薬( Bio-Rad USA) を 用 い て 化 学 発 光 さ せ , デ ジ タ ル 検 出 シ ス テ ム ChemiDoc Imaging System( Bio-Rad USA) に て 検 出 し た .
3 ) rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に お け る Akt-Pathway 分 子 の リ ン 酸 化 の 検 討
レ ク チ ン 陽 性 及 び 陰 性 細 胞 を DF6F 培 地 を 用 い て 培 養 後 ,
phosphatase 及 び protease inhibitor 含 有 RIPA cell lysis Buffer に て 細 胞 を 溶 解 し , 13,000g, 10 分 間 遠 心 後 , 上 清 を 回 収 し た . BCA Protein Assay Kit に よ り タ ン パ ク 濃 度 を 定 量 し た . 総 タ ン パ ク 20g に IRS-1 (Ser636/Ser639) , PTEN (Ser380) , Akt (Ser473) , GSK-3alpha/beta (Ser21/Ser9) ,BAD (Ser136), mTOR (Ser2448), p70 S6Kinase (Thr389) , S6Ribosome Protein (Ser235/Ser236) の 各 リ ン 酸 化 認 識 抗 体 結 合 ビ ー ズ を 加 え ,16 時 間 室 温 下 反 応 さ せ た .こ れ に ビ オ チ ン 化 標 識 検 出 抗 体 を 加 え 30 分 反 応 さ せ た 後 , さ ら に ス ト レ プ ト ア ビ ジ ン 結 合 PE( phycoerythrin)を 加 え , 10 分 間 反 応 さ せ ,BioPlex200 シ ス テ ム( Bio-Rad USA)に て Akt-Pathway 分 子 の リ ン 酸 化 を 解 析 し た .
12
4 ) IGF-2 及 び IGF-2 中 和 抗 体 の rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 の sphere 形 成 能 に 及 ぼ す 影 響
第 2 節 ( 4 ) の 方 法 に 準 じ て , rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 を 分 離 後 , 細 胞 低 吸 着 性 デ ィ ッ シ ュ (Sumitomo Bakelite Japan) (35mm 径 )へ 播 種 し , ヒ ト リ コ ン ビ ナ ン ト IGF-2(rhIGF-2)( R&D USA) を 0, 1, 5, 10, 50ng/ml の 各 濃 度 で 添 加 し , DF6F を 用 い て 無 血 清 浮 遊 培 養 を 行 い , sphere 形 成 能 を 評 価 し た . ま た , IGF-2 中 和 抗 体 ( ab9574 rabbit polyclonal, abcam UK)を 0,0.1,0.5,1.0,2.5µg/ml の 濃 度 で 添 加 後 , 同 様 に sphere 能 を 検 討 し た .
5 ) IGF-2 及 び IGF-2 中 和 抗 体 の rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 に お け る rBC2LCN 認 識 糖 鎖 の 発 現 に 及 ぼ す 影 響
各 細 胞 を rhIGF-2( R&D USA) (0, 1, 5, 10, 50ng/ml)及 び IGF-2 中 和 抗 体 ( ab9574 rabbit polyclonal, abcam UK) (0, 0.1, 0.5, 1.0, 2.5µg/ml)を そ れ ぞ れ 含 む DF6F 培 地 で , 48 時 間 37℃ , 5%CO2気 相 下 で
培 養 し た . 続 い て , 第 4 節 2 ) の 方 法 に 準 じ て タ ン パ ク 抽 出 , 定 量 を 行 い , 各 サ ン プ ル (10µg)を 10% SDS-PAGE に よ り 電 気 泳 動 後 , PVDF メ ン ブ レ ン に 転 写 し た . 次 に , 一 次 抗 体 と し て rBC2LCN-FITC(1:2000 濃 度 )( Wako Japan)及 び 抗 β -actin 抗 体 (0.5µg/ml)( 017-24551 mouse monoclonal, Wako) を TBS-T wash buffer に 希 釈 し 4 ℃ 下 overnight 処 理 し た . 続 い て , TBS-T wash buffer で 洗 浄 し , 二 次 抗 体 と し て HRP ラ ベ ル 化 抗 FITC 抗 体 ( 1:2000 濃 度 )( P5100 Dako Denmark) 及 び HRP ラ ベ ル 化 抗 マ ウ ス IgG 抗 体 ( 0.2µg/ml)( GTX221667-01 GeneTex) を 30 分 間 , 室 温 で 処 理 し た . TBS-T wash buffer で 洗 浄 後 ECL 試 薬 に よ
13
り 化 学 発 光 さ せ , デ ジ タ ル 検 出 シ ス テ ム ChemiDoc Imaging System ( Bio-Rad USA ) に て 検 出 し た . バ ン ド の 強 度 は Bio-Rad Image Lab software( Bio-Rad USA) を 用 い て 定 量 し , rBC2LCN 認 識 糖 鎖 発 現 量 を β -actin 発 現 量 で 割 っ た も の を relative expression と し て 算 出 し た .
第 5 節 rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 の 培 養 上 清 (conditioned medium:CM)か ら の exosome の 精 製 と そ の 評 価
1 ) 各 細 胞 株 の CM か ら の exosome の 精 製
A431, Ho-1-N-1, Ho-1-U-1 細 胞 株 を 100mm 径 細 胞 培 養 皿 ( Beckton Dickinson Falcon)に DF6F 培 地 を 用 い て 播 種 し ,増 殖 飽 和 状 態 の 細 胞 を PBS(-)に て 2 回 洗 浄 後 ,DF 基 礎 栄 養 培 地 に 交 換 し た .48 時 間 37℃ , 5 % CO2気 相 下 で 培 養 し , 培 養 上 清 を conditioned medium (CM)と し て
回 収 し た . CM を 1800rpm, 15 分 間 遠 心 後 , 0.45um フ ィ ル タ ー で ろ 過 し た . 続 い て , total exosome isolation® kit( invitrogen USA) を 用 い て CM 中 の exoxome を 精 製 し た . タ ン パ ク 定 量 は Qubit® Protein Assay Kit ( Themo Fisher Scientific) に て 測 定 し た .
2 ) ウ エ ス タ ン ブ ロ ッ ト 法 に よ る exosome の 品 質 評 価
Exosome( 1µg)を 10% あ る い は 12% SDS-PAGE に て 電 気 泳 動・分 離 後 , PVDF メ ン ブ レ ン に 転 写 し た . 5% ス キ ム ミ ル ク に て ブ ロ ッ キ ン グ を 行 い , 一 次 抗 体 と し て Cytochrome C( 556433 mouse monoclonal, Becton Dickinson 0.5µg/ml ), CD9 ( sc-59140 Santa Cruz 0.5µg/ml ), CD63 ( 353013 mouse monoclonal, BioLegend 5µg/ml),CD81( sc-23962 mouse monoclonal, Santa Cruz 1µg/ml) を 4℃ 下 , 一 昼 夜 処 理 し た . TBS-T
14
に て 洗 浄 後 ,2 次 抗 体 と し て HRP ラ ベ ル 化 抗 マ ウ ス IgG 抗 体( 0.2µg/ml) ( GTX221667-01 GeneTex)を 処 理 し ,室 温 下 30 分 間 イ ン キ ュ ベ ー ト し た .TBS-T に て 洗 浄 し ,ECL 試 薬 に て 化 学 発 光 さ せ ,デ ジ タ ル 検 出 シ ス テ ム ChemiDoc Imaging System( Bio-Rad USA) に て 検 出 し た .
3 ) 精 製 Exosome の 細 胞 内 再 取 り 込 み 能 の 検 討
第 5 節( 1 )で 得 ら れ た exosome 10g を , 等 量 の PKH26( mini26-1KT sigma-aldrich 4uM) と 混 和 し 5 分 間 室 温 に て 反 応 し , exosome を 標 識 し た . 限 外 ろ 過 フ ィ ル タ ー Amicon Ultra 0.5ml 100K ( Merck USA ) (14000g, 2 分 間 4 回 遠 心 し て , 未 反 応 PKH26 を 除 去 後 , PBS(-)に 懸 濁 し , PKH26 標 識 exosome と し た . PKH26 標 識 exosome(PKH26-exosome) を 0 , 250 , 500 , 1000ng/ml の 濃 度 で 培 養 細 胞 に 添 加 し . 同 時 に Hoechst33342( H3570 invitrogen USA 1:100000) に て 核 染 色 を 行 っ て 上 で . 12 時 間 37℃ , 5%CO2 気 相 下 で イ ン キ ュ ベ ー ト 後 , 共 焦 点 レ ー ザ ー 顕 微 鏡 ( Carl Zeiss, LSM700, Jena, Germany) で 観 察 し た .
4 ) 各 細 胞 由 来 exosome の sphere 形 成 に 及 ぼ す 影 響 rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 を 分 取 し , 第 5 節 ( 1 ) の 方 法 に 準 じ て CM か ら exosome を 精 製 し た .第 5 節( 2 )の 方 法 に よ り CD9 発 現 評 価 を 行 い , rBC2LCN 認 識 糖 鎖 陽 性 細 胞 に 陰 性 細 胞 由 来 exosome を ,陰 性 細 胞 に 陽 性 細 胞 由 来 exosome を ,0,250,500,1000 ng/ml 濃 度 条 件 で 添 加 し , Sphere 形 成 能 を 第 2 節 ( 4 )の 方 法 に 従 っ て 検 討 し た .
15 5 ) 各 細 胞 由 来 exosome の rBC2LCN 認 識 糖 鎖 発 現 に 及 ぼ す 影 響 rBC2LCN 認 識 糖 鎖 陽 性 細 胞 , 陰 性 細 胞 を 分 取 し , 第 5 節 ( 1 ) 及 び ( 2 ) の 方 法 に 従 い exosome を 精 製 し , さ ら に CD9 発 現 の 評 価 を 行 っ た 後 , rBC2LCN 認 識 糖 鎖 陽 性 細 胞 に 陰 性 細 胞 由 来 exosome を , ま た 陰 性 細 胞 に 陽 性 細 胞 由 来 exosome を , 0,250,500,1000 ng/ml の 濃 度 で 48 時 間 DF6F で 培 養 後 ,rBC2LCN 認 識 糖 鎖 発 現 に 及 ぼ す 各 exosome の 影 響 を 第 4 節 ( 5 ) の 方 法 に 準 じ て ウ エ ス タ ン ブ ロ ッ ト 法 で 検 討 し た .
16 第 3 章 結 果 1 . 各 細 胞 株 に お け る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 群 の 同 定 と そ の 細 胞 特 性 1 ) 各 種 癌 細 胞 株 に お け る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 の 存 在 と そ の 存 在 比
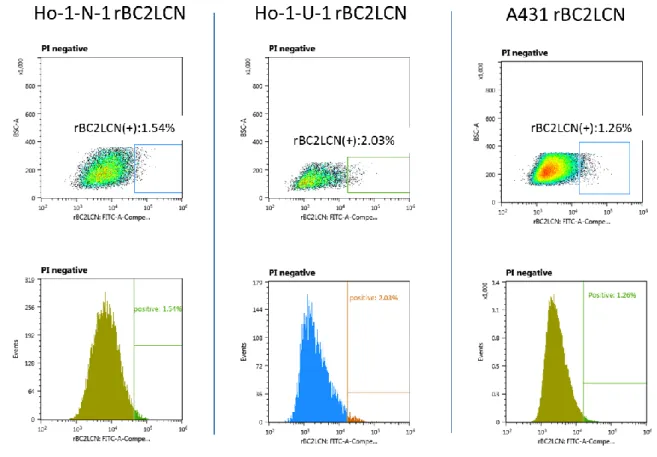
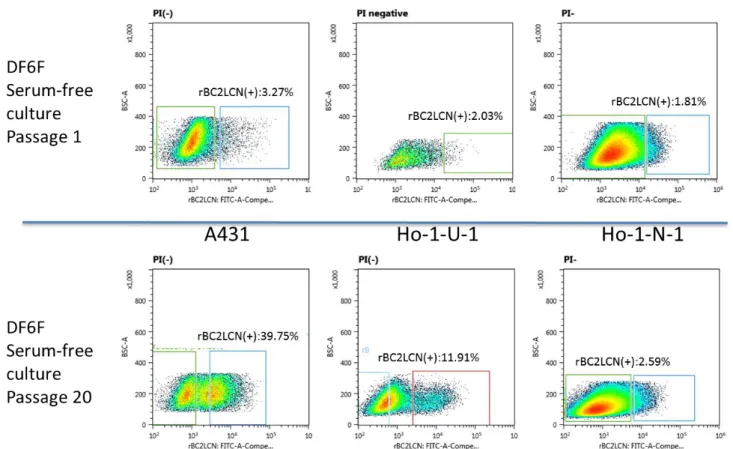
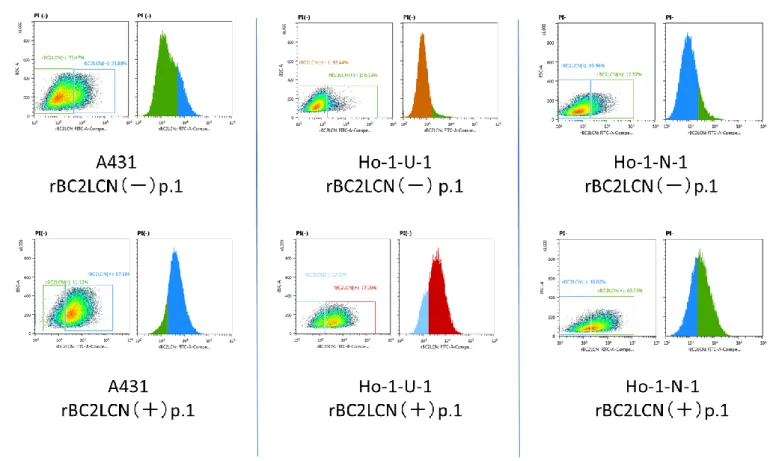
A431, Ho-1-U-1, Ho-1-N-1 細 胞 株 を DF5%CS 培 地 で の 培 養 条 件 下 で 維 持 培 養 し , フ ロ ー サ イ ト メ ト リ ー に て rBC2LCN 認 識 糖 鎖 陽 性 細 胞 の 存 在 の 有 無 を 検 討 し た . そ の 結 果 , こ れ ら の 細 胞 株 に は rBC2LCN 認 識 糖 鎖 を 発 現 す る 陽 性 細 胞 が 存 在 す る こ と が 明 ら か と な っ た . ま た そ の 全 細 胞 中 に お け る 存 在 比 は , Ho-1-N-1 細 胞 株 で は 1.5%, Ho-1-U-1 株 で は 2.0% , A431 株 で は 1.3% で あ っ た ( 図 2 ) . ま た , こ れ ら の 細 胞 株 を DF6 F 無 血 清 培 地 を 用 い た 培 養 条 件 下 で 20 継 代 後 に フ ロ ー サ イ ト メ ト リ ー に て rBC2LCN 認 識 糖 鎖 陽 性 細 胞 の 存 在 比 を 検 討 し た と こ ろ , A431 細 胞 株 ,Ho-1-U-1 細 胞 株 に お い て は 陽 性 細 胞 の 顕 著 な 増 加 を 認 め た . 一 方 , Ho-1-N-1 細 胞 株 で は ほ と ん ど 差 を 認 め な か っ た ( 図 3 ) . 2 ) ソ ー テ ィ ン グ 後 の rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 の ラ イ ブ セ ル イ メ ー ジ ン グ と 継 代 培 養 後 の 陽 性 及 び 陰 性 細 胞 率 の 変 化 の 検 討 A431,Ho-1-U-1,Ho-1-N-1 細 胞 株 よ り セ ル ソ ー タ ー に よ っ て rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 の 分 離 を 行 い , 無 血 清 単 層 培 養 状 態 で ラ イ ブ セ ル イ メ ー ジ ン グ に て rBC2LCN 認 識 糖 鎖 の 発 現 状 態 の 確 認 を 行 う と 陽 性 細 胞 は 陰 性 細 胞 と 比 較 し て 強 い 蛍 光 が 見 ら れ た .( 図 4 )
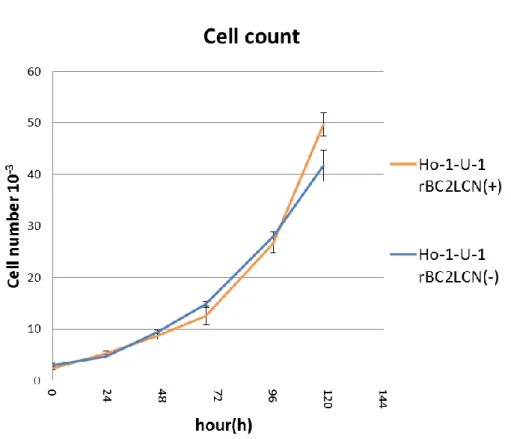
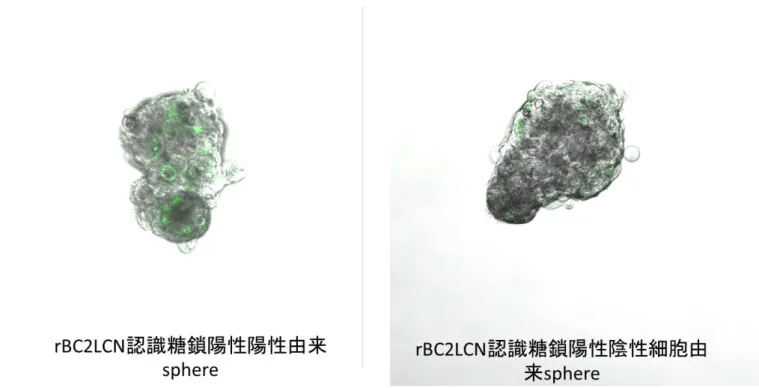
17 一 継 代 後 に , rBC2LCN 認 識 糖 鎖 陽 性 細 胞 率 を セ ル ソ ー タ ー を 用 い て 検 討 し た 結 果 , わ ず か 一 継 代 で , 陽 性 細 胞 群 か ら は 陰 性 細 胞 群 が , ま た 陰 性 細 胞 群 か ら は 陽 性 細 胞 群 が 出 現 し て い た ( 図 5 ) . 3) rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に お け る 無 血 清 単 層 培 養 系 で の 増 殖 能 Ho-1-U-1 細 胞 株 を rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に 分 離 し , DF6F を 用 い た 無 血 清 単 層 培 養 系 に て 増 殖 能 を 比 較 し た と こ ろ ,各 群 の 増 殖 能 に 有 意 な 差 は 認 め ら れ な か っ た ( 図 6 ) . 4) rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に お け る 無 血 清 浮 遊 培 養 系 で の sphere 形 成 能 rBC2LCN 認 識 糖 鎖 陽 性 細 胞 に お け る sphere 形 成 能 は 陰 性 細 胞 の そ れ と 比 較 し , 早 期 に 大 き な 多 数 の sphere を 形 成 し た ( 図 7 ). ま た , sphere 数 は 有 意 に 陽 性 細 胞 で 亢 進 し て い た ( 図 8 ) . 5) Sphere 細 胞 塊 中 に お け る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 の 存 在 動 態 rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 由 来 sphere に お け る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 を , ラ イ ブ セ ル イ メ ー ジ ン グ で 検 討 し た 結 果 ,陽 性 細 胞 由 来 sphere 及 び 陰 性 細 胞 由 来 sphere の い ず れ に お い て も , rBC2LCN 認 識 糖 鎖 陽 性 細 胞 が 存 在 し て い る 事 が 明 ら か に な っ た( 図 9 ).
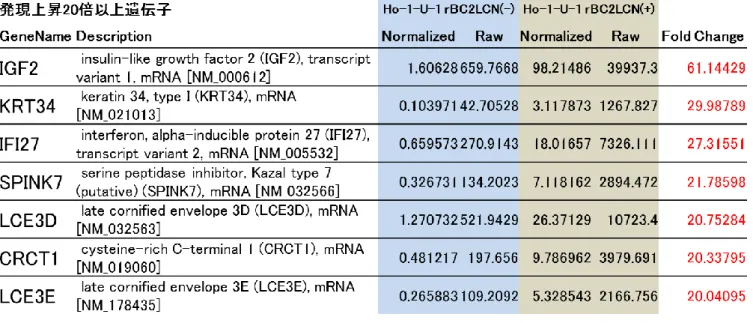
18 2 . DNA マ イ ク ロ ア レ イ に よ る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 に お け る 遺 伝 子 発 現 及 び そ の 比 較 検 討 解 析 1 ) rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に お け る DNA マ イ ク ロ ア レ イ に よ る 遺 伝 子 発 現 の 差 異 DNA マ イ ク ロ ア レ イ の 解 析 の 結 果 , rBC2LCN 認 識 糖 鎖 陽 性 細 胞 に お い て 陰 性 細 胞 と 比 較 し て 2 0 倍 以 上 の 発 現 亢 進 を 示 し た 遺 伝 子 は 7 種 類 認 め た . 中 で も IGF-2 の 発 現 は , 陽 性 細 胞 に お い て 陰 性 細 胞 と 比 較 し て 60 倍 以 上 亢 進 し て い た ( 表 1 ) . 3 . rBC2LCN 認 識 糖 鎖 陽 性 及 び 陰 性 細 胞 に お け る IGF-2 遺 伝 子 及 び タ ン パ ク 発 現 の 定 量
1 ) droplet digital PCR に よ る IGF-2 mRNA 発 現 量 の 定 量
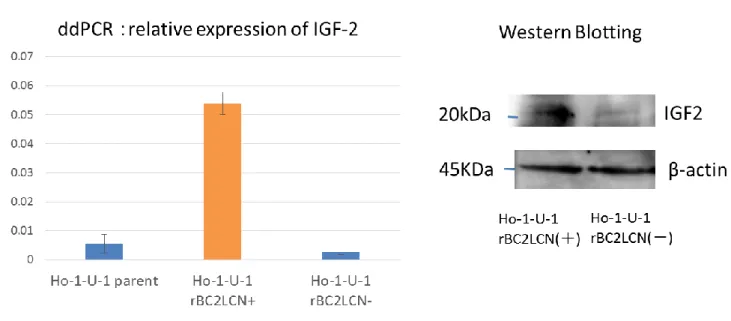
Droplet digital PCR 法 に よ る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に お け る IGF-2 mRNA 発 現 量 比 較 で は 陽 性 細 胞 は 陰 影 細 胞 に 比 較 し て 約 2 0 倍 発 現 が 亢 進 し て い た .( 図 1 0 ) 2 ) ウ エ ス タ ン ブ ロ ッ ト 法 に よ る IGF-2 発 現 量 の 比 較 ウ エ ス タ ン ブ ロ ッ ト 法 に よ る 解 析 で は rBC2LCN 認 識 糖 鎖 陽 性 細 胞 で は 陰 性 細 胞 に 比 較 し て 細 胞 内 に お け る IGF-2 タ ン パ ク 発 現 量 が 亢 進 し て い た .( 図 1 0 )
19 4 . rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に お け る Akt-Pathway 分 子 の リ ン 酸 化 1) rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に お け る Akt-Pathway リ ン 酸 化 の 検 討 結 果 rBC2LCN 認 識 糖 鎖 陽 性 細 胞 で は 陰 性 細 胞 と 比 較 し て IRS-1 (Ser636/Ser639) , PTEN (Ser380), Akt (Ser473), GSK-3alpha/beta (Ser21/Ser9) , BAD (Ser136 ), mTOR (Ser2448 ), p70 S6Kinase (Thr389) , S6Ribosome Protein (Ser235/Ser236)リ ン 酸 化 サ イ ト で の リ ン 酸 化 が 亢 進 し て い た .( 図 1 1 )
こ れ ら の 結 果 か ら rBC2LCN 認 識 糖 鎖 陽 性 細 胞 で は IGF-Akt Pathway の 機 能 亢 進 性 の リ ン 酸 化 亢 進 状 態 で あ る こ と が 示 唆 さ れ た .( 図 1 2 )
5 . IGF-2 及 び IGF-2 中 和 抗 体 の 各 細 胞 に お け る sphere 形 成 及 び rBC2LCN 認 識 糖 鎖 発 現 に 及 ぼ す 影 響
1) IGF-2 及 び IGF-2 中 和 抗 体 の sphere 形 成 に 及 ぼ す 影 響
IGF-2 お よ び ,IGF-2 中 和 抗 体 の ,rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 の sphere 形 成 能 に 及 ぼ す 影 響 を 検 討 し た . IGF-2 は 1,5,10,50ng/ml 濃 度 で 検 討 し た 結 果 , 濃 度 依 存 的 に 陰 性 細 胞 に お け る sphere 形 成 能 を 促 進 し た . 一 方 , IGF-2 中 和 抗 体 は 0.1,0.5,1.0,2.5µg/ml の 濃 度 で 検 討 し た 結 果 , で 陽 性 細 胞 の sphere 形 成 能 を 低 下 さ せ る こ と が 示 さ れ た ( 図 13) .
20 2) IGF-2 及 び IGF-2 中 和 抗 体 の rBC2LCN 認 識 糖 鎖 発 現 に 及 ぼ す 影 響 IGF-2 及 び IGF-2 中 和 抗 体 の 各 細 胞 に お け る rBC2LCN 認 識 糖 鎖 発 現 へ の 影 響 を ウ エ ス タ ン ブ ロ ッ ト 法 に て 検 討 し た . IGF-2 は 1,10,50ng/ml 濃 度 で 検 討 し た 結 果 , 濃 度 依 存 的 に rBC2LCN 認 識 糖 鎖 陰 性 細 胞 に お け る 糖 鎖 の 発 現 を 上 昇 さ せ た . 一 方 で , IGF-2 中 和 抗 体 は 0.5,1.0,2.5µg/ml の 濃 度 で , rBC2LCN 認 識 糖 鎖 陽 性 細 胞 に お け る 糖 鎖 発 現 を 低 下 さ せ た ( 図 14) . 6 . rBC2LCN 認 識 糖 鎖 陽 性 及 び 陰 性 細 胞 に お け る exosome の 細 胞 生 物 学 的 差 異 の 検 討 1 ) 各 細 胞 株 由 来 CM か ら の exosome 精 製 と そ の 評 価 A431,Ho-1-N-1,Ho-1-U-1 細 胞 株 由 来 CM か ら そ れ ぞ れ exosome を 精 製 し , 評 価 し た . 得 ら れ た exosome は , ウ エ ス タ ン ブ ロ ッ ト に て cytochrome C の 発 現 を 認 め な か っ た こ と か ら , 細 胞 成 分 の コ ン タ ミ ネ ー シ ョ ン は 無 い こ と が 示 さ れ た ( 図 15) . さ ら に , exosome マ ー カ ー と し て 知 ら れ て い る CD9, CD63,及 び CD81 の 精 製 exosome に お け る 発 現 を 検 討 し た 結 果 , こ れ ら の マ ー カ ー は 細 胞 分 画 に お い て 発 現 を 認 め た が , 精 製 exosome で は CD9 の み が 発 現 さ れ て い る 事 が 明 ら か と な っ た( 図 15). 精 製 exosome が 細 胞 に 再 取 り 込 み さ れ る か 否 か に つ い て , PKH26 に て 標 識 し た exosome を 用 い て 検 討 し た と こ ろ ,exosome の 濃 度 依 存 的 に 赤 色 の 蛍 光 小 胞 が 標 的 細 胞 内 に 確 認 さ れ た こ と か ら , 精 製 exosome は 細 胞 内 へ の 再 取 り 込 み 能 を 有 し て い る こ と が 明 ら か と な っ た ( 図 16) .
21 2 ) rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 由 来 exosome の 各 細 胞 に お け る sphere 形 成 能 に 及 ぼ す 影 響 rBC2LCN 認 識 糖 鎖 陽 性 細 胞 由 来 exosome は , 陰 性 細 胞 の sphere 形 成 能 を 促 進 し , sphere 数 の 増 加 と 大 き さ の 著 し い 亢 進 が 認 め ら れ た . 特 に , 500, 1000ng/ml の 濃 度 で 有 意 に ス フ ェ ア 形 成 能 を 促 進 し た . ま た さ ら に ,陰 性 細 胞 由 来 の exosome は , 陽 性 細 胞 の sphere 形 成 能 を 抑 制 す る 傾 向 を 示 し , 、特 に 250ng/ml の exoxome 濃 度 で は 有 意 に ス フ ェ ア 形 成 を 低 下 さ せ た .( 図 17, 18) . 3 ) rBC2LCN 認 識 糖 鎖 陽 性 細 胞 及 び 陰 性 細 胞 由 来 exosome の 各 細 胞 に お け る rBC2LCN 認 識 糖 鎖 発 現 に 及 ぼ す 影 響 ( 2 )と 同 様 に , 陽 性 及 び 陰 性 細 胞 由 来 exosome の , そ れ ぞ れ 陰 性 細 胞 及 び 陽 性 細 胞 に お け る rBC2LCN 認 識 糖 鎖 発 現 に 及 ぼ す 影 響 を 検 討 し た 結 果 , 陽 性 細 胞 由 来 exosome は 濃 度 依 存 的 に 陰 性 細 胞 に お け る rBC2LCN 認 識 糖 鎖 発 現 を 亢 進 さ せ た ( 図 19) .
22
第 4 章 考 察
悪 性 腫 瘍 が 幹 細 胞 を 起 源 と す る 仮 説 は , First formulated in 1875, Julius Cohnheim, proposed that stem cell misplaced during embryonal development were the source of tumors later in the life <embryonal-rest theory >, Bruce ら (Bruce, et al, 1963) が , 悪 性 腫 瘍 内 の 少 数 の 細 胞 の み が 腫 瘍 形 成 能 を 有 す る こ と か ら , 悪 性 腫 瘍 は 自 己 複 製 能 を 有 す る 少 数 の 特 別 な 癌 細 胞 に よ り 維 持 さ れ て い る と い う 仮 説 を 報 告 し た こ と に 端 を 発 す る . し か し な が ら , こ の 仮 説 は 広 く 受 け 入 れ ら れ ず , 悪 性 腫 瘍 は 均 一 な 癌 細 胞 集 団 と , こ れ ら 細 胞 集 団 が 巻 き 込 ん だ 正 常 細 胞 か ら 構 成 さ れ る ヘ テ ロ 細 胞 集 団 と 考 え ら れ , 悪 性 腫 瘍 の 大 部 分 を 占 め る い わ ゆ る 癌 細 胞 ( 非 癌 幹 細 胞 ) を 対 象 と し た 研 究 が 精 力 的 に 進 め ら れ て き た .し か し 1994 年 に ,急 性 骨 髄 性 白 血 病( AML) に お い て 癌 幹 細 胞 様 の 細 胞 集 団 が 同 定 さ れ (Lapidot, et al, 1994), そ れ が 組 織 幹 細 胞 の 細 胞 生 物 学 的 特 性 と 類 似 し て お り , CD34+CD38-の 表 現 型 を 持 つ 白 血 病 幹 細 胞 の み が 免 疫 不 全 マ ウ ス で の 造 腫 瘍 性 を 有 す る 事 が 示 さ れ , 造 血 幹 細 胞 と し て の 表 現 型 と 類 似 し て い る こ と が 証 明 さ れ た .以 来 ,さ ま ざ ま な 固 形 腫 瘍 に も 幹 細 胞 が 存 在 し ,腫 瘍 の 発 生 ・ 進 展 ・ 維 持 に 重 要 な 役 割 を 果 た し て い る 可 能 性 が 明 ら か に さ れ つ つ あ る . つ ま り , 癌 組 織 に は 自 己 複 製 能 を 有 し , 半 永 久 的 に 娘 細 胞 を 生 み 続 け る 少 数 の 細 胞 群 と , 高 い 増 殖 能 を 持 つ が , 最 終 的 に は 分 化 や 老 化 に よ り 増 殖 能 を 失 う 大 多 数 の 細 胞 群 か ら な る こ と が 明 ら か に な っ て き た .こ の こ と は ,正 常 の 組 織 幹 細 胞 と そ れ か ら 生 じ た 分 化 細 胞 の よ う な 関 係 が 癌 組 織 に も 存 在 し て い る こ と を 示 唆 し て い る . こ の よ う な 幹 細
23
胞 様 の 癌 細 胞 つ ま り 癌 幹 細 胞 は , 放 射 線 や 抗 癌 剤 に 対 す る 抵 抗 性 も 高 い こ と が 予 想 さ れ て い る . 事 実 , 化 学 療 法 , 放 射 線 療 法 や 手 術 療 法 後 , 臨 床 的 に CR を 示 し た 症 例 が 数 年 後 に 再 発 す る 事 は よ く 経 験 す る が ,こ れ ら 治 療 抵 抗 性 や 再 発 例 に 癌 幹 細 胞 が 関 与 し て い る と 考 え ら れ る . (Dalerba, et al, 2007; Trumpp, et al, 2008; Visvader, et al, 2008; Jones, et al, 2009) 癌 幹 細 胞 は , 正 常 組 織 幹 細 胞 そ の も の に 遺 伝 子 変 化 が 蓄 積 し た 細 胞 な の か ,あ る い は 正 常 幹 細 胞 か ら 分 化 方 向 へ ト ラ ン ジ ッ ト 中 の 細 胞 に 遺 伝 子 変 化 が 蓄 積 し 癌 化 し た 細 胞 な の か ,あ る い は 変 異 が 蓄 積 し た ト ラ ン ジ ッ ト 細 胞 に 何 ら か の リ プ ロ グ ラ ミ ン グ が 起 こ り 幹 細 胞 機 能 を 再 獲 得 し た 細 胞 な の か 不 明 で あ る が , 癌 の 起 源 と な る 細 胞 と 考 え ら れ て お り , 癌 幹 細 胞 と 組 織 幹 細 胞 と の 間 に は , そ の 自 己 複 製 能 や 分 化 能 の 制 御 機 構 に 多 く の 共 通 性 が あ る こ と が 予 想 さ れ る .( Lessard,J ら 2003, Barker ら 2009) 人 工 多 能 性 幹 ( iPS) 細 胞 は ( Takahashi ら 2007), エ ピ ジ ェ ネ テ ィ ッ ク な 変 化 が 起 こ っ て い る 分 化 し た 体 性 細 胞 に , oct3/4, sox2, klf4, c-Myc の 4 種 類 の 遺 伝 子 を 導 入 す る こ と に よ り , 自 己 複 製 能 と 多 分 化 能 を 再 獲 得 し た 細 胞 で あ る が , こ れ ら 遺 伝 子 の う ち , c-myc は 癌 遺 伝 子 で あ り klf4 は 癌 遺 伝 子 あ る い は 癌 抑 制 遺 伝 子 と し て も 機 能 す る こ と か ら , iPS 細 胞 の 誘 導 機 構 に 類 似 し た 機 構 が 癌 幹 細 胞 の 誘 導 に も 関 与 し て い る 可 能 性 が 考 え ら れ て い る . 一 方 , 組 織 幹 細 胞 な ど の 正 常 幹 細 胞 が 未 分 化 性 と 多 分 化 能 を 維 持 す る た め に は , そ れ を 可 能 に す る 微 小 環 境 (ニ ッ チ )が 必 要 と 考 え ら れ て お り , ニ ッ チ 内 で 幹 細 胞 の 数 や 細 胞 分 裂 , 分 化 が 制 御 さ れ て い る 可 能 性 が 高 い . し た が っ て , 癌 幹 細 胞 に お い て も , 正 常 組 織 幹 細 胞 と 類 似 し た 自 己 複 製 能 と 分 化 能 を 維 持 す
24 る 機 構 す な わ ち ニ ッ チ が 存 在 し て い る 可 能 性 が 高 い ( 図 1) . そ こ で 私 は ,ES/iPS 細 胞 な ど の 未 分 化 な 多 能 性 幹 細 胞 が 特 異 的 に 発 現 す る 糖 鎖 構 造 を 認 識 す る レ ク チ ン で あ る rBC2LCN の 口 腔 扁 平 上 皮 癌 細 胞 の 幹 細 胞 マ ー カ ー と し て の 可 能 性 を 検 討 し , そ の 腫 瘍 形 成 の メ カ ニ ズ ム と が ん 微 小 環 境 ( ニ ッ チ ) の 維 持 メ カ ニ ズ ム を 解 明 し , 放 射 線 感 受 性 や 腫 瘍 発 生 メ カ ニ ズ ム を 明 ら か に す る こ と を 目 指 し た . rBC2LCN 認 識 糖 鎖 は 未 分 化 多 能 性 幹 細 胞 の 細 胞 表 面 上 に 発 現 す る 糖 タ ン パ ク の ポ ド カ リ ク シ ン に 付 加 さ れ る 特 異 的 な 糖 鎖 構 造 を 認 識 し て い る と 考 え ら れ て い る ( Tateno ら 2013). こ の 糖 鎖 抗 原 は ES 細 胞 や iPS 細 胞 な ど の 未 分 化 多 能 性 幹 細 胞 に の み 発 現 し , ES 細 胞 や iPS 細 胞 か ら 分 化 し た 細 胞 や 体 性 細 胞 で は 発 現 し な い( Tateno ら 2013).糖 鎖 発 現 状 態 は 糖 転 位 酵 素 に よ り コ ン ト ロ ー ル さ れ て い る と 考 え ら れ て い る が , そ の 詳 細 な メ カ ニ ズ ム に つ い て は 未 だ 不 明 な 点 が 多 い ( Kizuka ら 2014) . rBC2LCN 認 識 糖 鎖 は 機 能 的 な 側 面 は 未 だ 不 明 で あ る が , 未 分 化 多 能 性 幹 細 胞 に お け る 機 能 維 持 に 関 連 し て 発 現 し て い る も の と 考 え ら れ て い る ( Tateno ら 2013) . rBC2LCN 認 識 糖 鎖 発 現 細 胞 は 口 腔 扁 平 上 皮 癌 細 胞 株 Ho-1-U-1 細 胞 株 及 び Ho-1-N-1 細 胞 株 で そ れ ぞ れ 2.0% 及 び 1.5% 存 在 し た .ま た , 外 陰 部 扁 平 上 皮 癌 細 胞 株 で あ る A431 細 胞 株 に お い て も 1.3%の 陽 性 細 胞 の 存 在 が 認 め ら れ た . こ れ ら 細 胞 は 無 血 清 浮 遊 培 養 系 で 陰 性 細 胞 に 比 較 し て 高 い sphere 形 成 能 を 示 し た こ と か ら , 癌 幹 細 胞 と し て の 特 徴 を 持 つ こ と が 明 ら か と な っ た . こ の こ と か ら , 未 分 化 多 能 性 幹 細 胞 に 特 異 的 な 糖 鎖 抗 原 と , 癌 幹 細 胞 に お け る 糖 鎖 抗 原 の 発 現 が 共 通 で あ る 可 能 性 が 示 唆 さ れ た . 陽 性 細 胞 と 陰 性 細 胞 の 特 性 を 明 ら か に す る た め に ,各 細 胞 を セ ル ソ
25 ー タ ー に よ り 分 離 し , 無 血 清 単 層 培 養 系 で 継 代 を す る と , わ ず か 1 継 代 後 で も 陽 性 細 胞 群 か ら 陰 性 細 胞 群 が , 陰 性 細 胞 群 か ら 陽 性 細 胞 が 出 現 し た こ と , さ ら に , 陽 性 細 胞 由 来 sphere 及 び 陰 性 由 来 sphere の い ず れ に も 陽 性 細 胞 が 含 ま れ て い た こ と か ら , 陽 性 細 胞 と 陰 性 細 胞 は 互 い に ト ラ ン ジ ッ ト す る 関 係 に あ る と 考 え ら れ た . さ ら に , 血 清 培 養 系 か ら 無 血 清 培 養 系 へ の 変 更 に よ っ て こ れ ら の 変 化 が 生 じ た こ と か ら , こ れ ら の 発 現 は , 何 ら か の 因 子 に よ っ て 制 御 さ れ て い る 事 が 強 く 示 唆 さ れ た . DNA マ イ ク ロ ア レ イ を 用 い た 解 析 で は 陽 性 細 胞 に お い て 陰 性 細 胞 と 比 較 し IGF-2 の 発 現 が 60 倍 以 上 亢 進 し て お り , droplet digital PCR 及 び ウ エ ス タ ン ブ ロ ッ ト の 結 果 か ら も , 陽 性 細 胞 に お い て IGF-2 の mRNA と タ ン パ ク レ ベ ル で の 発 現 亢 進 が 認 め ら れ る 事 が 明 ら か に な っ た . こ の こ と か ら , 陽 性 細 胞 の 機 能 維 持 に お い て IGF-2 が 重 要 な 機 能 を 担 っ て い る 可 能 性 が 考 え ら れ た . 各 細 胞 に お け る IGF-2 に 関 わ る 細 胞 シ グ ナ ル 伝 達 pathway を 明 ら か に す る た め , IGF-Akt pathway 分 子 の リ ン 酸 化 に つ い て 検 討 し た と こ ろ , 陽 性 細 胞 で は 陰 性 細 胞 に 比 較 し て , 一 様 に Akt-Pathway 分 子 の リ ン 酸 化 が 促 進 し て い る 事 が 明 ら か に な っ た . Akt-Pathway は 細 胞 の 生 存 , 増 殖 , 分 化 , 糖 代 謝 , タ ン パ ク 質 合 成 の 機 能 を 担 う 重 要 な シ グ ナ ル 伝 達 系 で あ り , 癌 幹 細 胞 と し て の 機 能 維 持 に 重 要 な 機 能 を 果 た し て い る と 考 え ら れ た . IGF-2 が 糖 鎖 発 現 に 重 要 な 機 能 を 果 た し て い る 可 能 性 は , リ コ ン ビ ナ ン ト IGF-2 が 陰 性 細 胞 の sphere 形 成 能 を 濃 度 依 存 的 に 促 進 し た こ と , ま た rBC2LCN 認 識 糖 鎖 の 発 現 を 亢 進 し た 事 , さ ら に は IGF-2 中 和 抗 体 が 陽 性 細 胞 の sphere 形 成 や rBC2LCN 認 識 糖 鎖 の 発 現 を 低 下 さ せ た こ と か ら も 強 く 裏 付 け ら れ た .
26 近 年 , IGF-2 及 び そ の シ グ ナ ル 分 子 を タ ー ゲ ッ ト と し た 分 子 標 的 治 療 薬 の い く つ か が 臨 床 試 験 段 階 に あ る こ と が 報 告 さ れ て い る ( Helen ら 2013) . こ れ ら は IGF-2 を タ ー ゲ ッ ト と し た 中 和 抗 体 , IGF-2 の レ セ プ タ ー で あ る IGF-1R に 対 す る 競 合 抗 体 , IGF-1R の チ ロ シ ン キ ナ ー ゼ 活 性 阻 害 剤 等 で あ る . こ れ ら 薬 剤 を 用 い る こ と で , 本 研 究 に お い て 明 ら か に な っ た 幹 細 胞 に お け る 機 能 維 持 に 重 要 な IGF-2 の 機 能 阻 害 を 行 う と い う , 癌 幹 細 胞 を タ ー ゲ ッ ト と し た 治 療 戦 略 の 展 開 が 期 待 さ れ る ( 図 20) . rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に お け る exosome の 生 物 活 性 の 差 異 を 検 討 す る こ と で , 各 細 胞 間 の 相 互 作 用 , ニ ッ チ 形 成 に つ い て の メ カ ニ ズ ム を 明 ら か に し た . Exosome は 直 径 40nm~ 150nm 程 度 の 細 胞 外 小 胞 で あ り ,Johnstone ら に よ り 命 名 さ れ た( Johnstone ら 1983). Exosome 小 胞 中 に は タ ン パ ク 質 ,mRNA, miRNA 等 の 情 報 伝 達 物 質 が 内 包 さ れ て い る . こ の た め , 細 胞 間 の 情 報 伝 達 に 関 わ り , 形 質 の 伝 播 や 癌 細 胞 ニ ッ チ 形 成 に 関 わ っ て い る こ と が 考 え ら れ る . 本 研 究 で は ま ず , 扁 平 上 皮 細 胞 株 A431,Ho-1-N-1,Ho-1-U-1 細 胞 株 よ り ポ リ マ ー 沈 殿 法 に て exosome の 精 製 を 行 っ た . 精 製 し た exosome に は ミ ト コ ン ド リ ア 内 膜 成 分 で あ る cytochrome C は 検 出 さ れ ず ,細 胞 成 分 の コ ン タ ミ ネ ー シ ョ ン が 無 い こ と が 確 認 さ れ た . ま た , 本 精 製 exosome は exosome マ ー カ ー と し て テ ト ラ ス パ ニ ン フ ァ ミ リ ー で あ る CD9 を 発 現 し て い る 事 が 明 ら か に な っ た . CD9 は 肺 が ん 細 胞 由 来 exosome 等 に お い て も 発 現 の 見 ら れ る exosome マ ー カ ー で あ る ( Yoshioka ら 2013 ) . ま た , exosome は 精 製 過 程 で exosome の 細 胞 再 取 り 込 み 機 能 が 低 下 す る 可 能 性 が 報 告 さ れ て い る た め , PKH26 標 識 exosome に よ る 細 胞 内 再 取 り 込 み 能 を 検 討 し た 結 果 , 濃 度 依 存 的 に exosome の 細 胞 内 取 り 込 み が 確 認
27 さ れ た こ と か ら , 本 精 製 exosome の ク オ リ テ ィ ー が 担 保 さ れ て い る こ と が 確 認 さ れ た .今 回 , 無 血 清 培 養 系 で 長 期 間 培 養 し て い る 細 胞 の CM か ら 精 製 し た 精 製 し た exoxome は CD9 の み を 発 現 し て い る こ と が 明 ら か と な っ た .腫 瘍 細 胞 由 来 exoxsome で は , CD9 に 加 え て CD63 及 び CD81 が 発 現 さ れ て い る こ と が 報 告 さ れ て い る ( Yoshioka ら 2013) . こ れ は exosome の 精 製 方 法 に も 依 存 し て い る 可 能 性 は 否 定 で き な い も の の , こ れ ら の 報 告 は 血 清 添 加 培 地 を 用 い て 培 養 し た 細 胞 か ら 精 製 し た exoxome の 解 析 で あ る こ と か ら , CD63 や CD81 発 現 exosome は 血 清 由 来 の も の で あ る 可 能 性 が 考 え ら れ た . rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 の CM か ら 精 製 し た exosome を , そ れ ぞ れ 陰 性 細 胞 及 び 陽 性 細 胞 に 0,250,500,1000ng/ml の 濃 度 で 処 理 し , rBC2LCN 認 識 糖 鎖 の 発 現 及 び sphere 形 成 能 を 検 討 し た と こ ろ , 陽 性 細 胞 由 来 の exosome は 陰 性 細 胞 に お け る rBC2LCN 認 識 糖 鎖 の 発 現 及 び sphere 形 成 能 を 濃 度 依 存 的 に 亢 進 し た . 一 方 , 陰 性 細 胞 由 来 exosome は 陽 性 細 胞 に お け る sphere 形 成 能 を 低 下 さ せ る 傾 向 を 認 め , 250ng/ml の 濃 度 で は 有 意 に 低 下 さ せ た .こ の こ と か ら ,陽 性 細 胞 由 来 の exosome に は 陰 性 細 胞 を 陽 性 細 胞 へ ト ラ ン ジ ッ ト さ せ る 活 性 を 有 し て い る 事 が 考 え ら れ た . こ の こ と は , 陽 性 細 胞 と 陰 性 細 胞 は exosome を 介 し て communicate し , 癌 細 胞 ニ ッ チ 形 成 に お い て 重 要 な 機 能 を 果 た し て い る と 考 え ら れ た . 陽 性 細 胞 由 来 exosome が , 陰 性 細 胞 か ら 陽 性 細 胞 へ の ト ラ ン ジ シ ョ ン を 引 き 起 こ す 機 構 は 不 明 で あ る が ,exosome 内 に 含 ま れ る 細 胞 増 殖 因 子 , miRNA や そ の 他 の 活 性 物 質 の 存 在 が 単 独 あ る い は 連 携 し て そ の 機 能 を 果 た し て い る も の と 考 え ら れ た . し た が っ て , rBC2LCN 認 識 糖 鎖 陽 性 細 胞 由 来 exosome は , 癌 幹 細 胞 を 含 む 扁 平 上 皮 癌 の 診 断 ・ 治 療 の タ ー ゲ ッ ト と な り 得 る 可 能 性 が 示 さ れ た ( 図
28 21) . Tateno ら , 及 び Hirabayashi ら は , 細 胞 障 害 性 毒 素 結 合 rBC2LCN を 用 い て ,ES 細 胞 や iPS 細 胞 由 来 分 化 細 胞 に よ る 再 生 医 療 時 に お い て 発 が ん 性 な ど の 問 題 と な る 残 存 し た 未 分 化 幹 細 胞 を 効 果 的 に 除 去 す る こ と が で き る こ と を 報 告 し て い る ( Tateno ら 2015 , Hirabayashi ら 2015) . 本 研 究 で は 未 分 化 多 能 性 幹 細 胞 に 発 現 し rBC2LCN に 認 識 さ れ る 糖 鎖 抗 原 が 扁 平 上 皮 癌 細 胞 株 に お い て も 発 現 し , か つ 癌 幹 細 胞 と し て の 特 徴 を 有 し て い た こ と か ら , 同 様 の 戦 略 に よ り , 選 択 的 ・ 効 果 的 に rBC2LCN 認 識 糖 鎖 発 現 癌 幹 細 胞 を 殺 滅 で き る 可 能 性 が 考 え ら れ た . 以 上 の こ と か ら ,未 分 化 な ES/iPS 細 胞 が 特 異 的 に 発 現 す る , rBC2LCN レ ク チ ン で 認 識 さ れ る 糖 鎖 構 造 を 有 す る 細 胞 が , 各 種 扁 平 上 皮 癌 細 胞 株 に お い て も 存 在 し , 同 細 胞 が 癌 幹 細 胞 と し て 機 能 し て い る 可 能 性 が 示 さ れ た こ と か ら , 糖 鎖 発 現 細 胞 は 癌 化 に 深 く 関 わ っ て い る こ と が 考 え ら れ た . ま た , IGF-2 及 び exosome は 癌 幹 細 胞 ニ ッ チ の 機 能 維 持 に 重 要 な 機 能 を 果 た し て い る こ と が 考 え ら れ , rBC2LCN 認 識 糖 鎖 発 現 細 胞 , IGF-Akt Pathway 及 び exosome を 標 的 と し た 口 腔 扁 平 上 皮 癌 細 胞 の 診 断 ・ 治 療 へ の 応 用 が 考 え ら れ た .
29 総 括 本 研 究 で は , ヒ ト 口 腔 扁 平 上 皮 癌 に お け る 癌 幹 細 胞 を 標 的 と し た 新 し い 診 断 ・ 治 療 法 を 開 発 す る こ と を 目 指 し , ヒ ト 口 腔 扁 平 上 皮 癌 細 胞 株 よ り 分 離 し た rBC2LCN 認 識 糖 鎖 陽 性 細 胞 群 の 細 胞 ・ 分 子 生 物 学 的 特 性 解 析 を , 無 血 清 培 養 系 を 用 い て 行 っ た . そ の 結 果 以 下 の 事 が 明 ら か と な っ た . 1 ) rBC2LCN が 認 識 す る , 未 分 化 な ES/iPS 細 胞 が 特 異 的 に 発 現 す る 糖 鎖 構 造 を 有 す る 細 胞 が , 各 種 扁 平 上 皮 癌 細 胞 株 に お い て も 存 在 し て い る こ と が 明 ら か と な り , そ の 存 在 比 率 は 1.5% ~ 2.0 % で あ っ た 2 ) rBC2LCN 認 識 糖 鎖 を 発 現 す る 扁 平 上 皮 癌 細 胞( レ ク チ ン 陽 性 細 胞 ) は が ん 幹 細 胞 と し て の 性 質 を 有 し て い る 可 能 性 が 示 さ れ た 3 ) レ ク チ ン 陽 性 細 胞 で は 陰 性 細 胞 と 比 較 し て , IGF-2 遺 伝 子 ・ タ ン パ ク 発 現 の 亢 進 , 及 び IGF-Akt pathway の 活 性 化 を 認 め , IGF-2 は 陰 性 細 胞 の 陽 性 細 胞 へ の ト ラ ン ジ ッ ト を 促 進 し た . 4 ) 陽 性 細 胞 由 来 exosome は 陰 性 細 胞 の 陽 性 細 胞 へ の ト ラ ン ジ ッ ト を 促 進 し た 以 上 の こ と か ら ,未 分 化 な ES/iPS 細 胞 が 特 異 的 に 発 現 す る 糖 鎖 構 造 を 有 す る 細 胞 が , 各 種 扁 平 上 皮 癌 細 胞 株 に お い て も 存 在 し , 同 細 胞 が 幹 細 胞 と し て 機 能 し て い る 可 能 性 が 示 さ れ た こ と か ら , 糖 鎖 発 現 細 胞 は 癌 化 に 深 く 関 わ っ て い る こ と が 考 え ら れ た . IGF-2 及 び exosome は が ん 幹 細 胞 ニ ッ チ の 機 能 維 持 に 重 要 な 機 能 を 果 た し て い る こ と が 考 え ら れ , rBC2LCN 認 識 糖 鎖 発 現 細 胞 , IGF-Akt
30
Pathway 及 び exosome を 標 的 と し た 口 腔 扁 平 上 皮 癌 細 胞 の 診 断 ・ 治 療 へ の 応 用 が 考 え ら れ た .
31
参 考 文 献
Adewumi O, Aflatoonian B, Ahrlund -Richter L, Amit M, Andrews PW, Beighton G, Bello PA, Benvenisty N, Berry LS, Bevan S, Blum B, Brooking J, Chen KG, Choo AB, Churchill GA, Corbel M, Damjanov I, Draper JS, Dvorak P, Emanuelsson K, Fleck RA, Ford A, Gertow K, Gertsenstein M, Gokhale PJ, Hamilton RS, Hampl A, Healy LE, Hovatta O, Hyllner J, Imreh MP, Itskovitz-Eldor J, Jackson J, Johnson JL, Jones M, Kee K, King BL, Knowles BB, Lako M, Lebrin F, Mallon BS, Manning D, Mayshar Y, McKay RD, Michalska AE, Mikkola M, Mileikovsky M, Minger SL, Moore HD, Mummery CL, Nagy A, Nakatsuji N, O'Brien CM, Oh SK, Olsson C, Otonkoski T, Park KY, Passier R, Patel H, Patel M, Pedersen R, Pera MF, Piekarczyk MS, Pera RA, Reubinoff BE, Robins AJ, Rossant J, Rugg -Gunn P, Schulz TC, Semb H, Sherrer ES, Siemen H, Stacey GN, Stojkovic M, Suemori H, Szatkiewicz J, Turetsky T, Tuuri T, van den Brink S, Vintersten K, Vuoristo S, Ward D, Weaver TA, Young LA, Zhang W. Characterization of human embryonic stem cell lines by the International Stem Cell Initiative. Nat Biotechnol. 2007 Jul;25(7):803-16. Epub 2007 Jun 17.
Ailles LE, Weissman IL, Cancer stem cells in solid tumors, Curr Opin Biotechnol, 18: 460-466, 2007.
Al-Nedawi K1, Meehan B, Micallef J, Lhotak V, May L, Guha A, Rak J. Intercellular transfer of the oncogenic receptor EGFRvIII by
32
microvesicles derived from tu mour cells. Nat Cell Biol. 2008 May;10(5):619-24. doi: 10.1038/ncb1725. Epub 2008 Apr 20.
Barker N1, Ridgway RA, van Es JH, van de Wetering M, Begthel H, van den Born M, Danenberg E, Clarke AR, Sansom OJ, Clevers H. Crypt stem cells as the cells-of-origin of intestinal cancer. Nature. 2009 Jan 29;457(7229):608-11. doi: 10.1038/nature07602. Epub 2008 Dec 17.
Barnes D, and Sato HG., Serum-free cell culture: a unifying approach, Cell, 22: 649-55, 1980.
Charafe-Jauffret E., Ginestier C., Birnbaum D., Breast cancer stem cells: tools and models to rely on, BMC Cancer,
Chen HX, Sharon E. IGF-1R as an anti-cancer target--trials and tribulations. Chin J Cancer. 2013 May;32(5):242 -52. doi: 10.5732/cjc.012.10263. Epub 2013 Apr 19.
Dalerba P, Clarke MF, Cancer stem cells and tumor metastasis: first steps into uncharted territory, Cell Stem Cell, 1: 241 -2, 2007.
Darmon M, Serrero G, Rizzino A, Sato G, Isolation of myoblastic, fibro-adipogenic, and fibroblastic clonal cell lines from a common precursor and study of their requirements for growth and
33
differentiation, Exp Cell Res, 132: 313 -27, 1981.
Deonarain MP, Kousparou CA, Epenetos AA, Antibodies targeting cancer stem cells: a new paradigm in immunotherapy?, MAbs, 1: 12 -25, 2009.
Folkins C, Shaked Y, Man S, Tang T, Lee CR, Zhu Z, Hoffman RM, Kerbel RS, Glioma tumor stem-like cells promote tumor angiogenesis and vasculogenesis via vascular endothelial growth factor and stromal-derived factor 1, Cancer Res, 69:7243 -7251, 2009.
Giard DJ, et al. In vitro cultivation of human tumors: establishment of cell lines derived from a series of solid tumors. J. Natl. Cancer Inst. 51: 1417 -1423, 1973. PubMed: 4357758
Gilbert CA, Ross AH, Cancer stem cells: cell culture, markers, and targets for new therapies, J Cell Biochem, 10 8: 1031-1038, 2009.
Goodell MA, Brose K, Paradis G, Conner AS, Mulligan RC, Isolation and functional properties of murine hematopoietic stem cells that are replicating in vivo, J Exp Med,183:1797 -1806, 1996.
Gupta PB, Chaffer CL, Weinberg RA, Cancer stem cells: mirage or reality?, Nat Med, 15: 1010-1012, 2009.
34
Hadnagy A, Gaboury L, Beaulieu R, Balicki D, SP analysis may be used to identify cancer stem cell populations, Exp Cell, 312 : 3701-3710, 2006.
Hermann PC, Huber SL, Herrler T, Aicher A, Ellwart JW, Guba M, Bruns CJ, Heeschen C, Distinct populations of cancer stem cells determine tumor growth and metastatic activity in human pancreatic cancer, Cell Stem Cell, 1: 313 -23, 2007.
Hirabayashi J, Tateno H, Onuma Y, Ito Y. A Novel Probe as Surface Glycan Marker of Pluripotent Stem Cells: Research Outcomes and Application to Regenerative Medicine. Adv Healthc Mater. 2015 Nov 18;4(16):2520-9. doi: 10.1002/adhm.201400837. Epub 2015 Apr 14.
Hirschmann-Jax C, Foster AE, Wulf GG, Nuchtern JG, Jax TW, Gobel U, Goodell MA, Brenner MK, A distinct "side population" of cells with high drug efflux capacity in human tumor cells, PNAS, 101: 14228-14233, 2004.
Ho MM, Ng AV, Lam S, Hung JY, Side population in human lung cancer cell lines and tumors is enriched with st em-like cancer cells, Cancer Res, 67: 4827-4833, 2007.
Huls M, Russel FG, Masereeuw R, The role of ATP binding cassette transporters in tissue defense and organ regeneration, J Pharmacol
35
Exp Ther, 328: 3-9, 2008.
Jones RJ, Cancer stem cells-clinical relevance, J Mol Med, 87: 1105-1110, 2009.
Julius Cohnheim, proposed that stem cell misplaced during embryonal development were the source of tumors later in the life <embryonal-rest theory> First formulated in 1875
Kondo T, Setoguchi T, Taga T, Persistence of a small subpopulation of cancer stem-like cells in the C6 glioma cell line, PNAS, 101; 781-786, 2004.
Kvinlaug BT, Huntly BJ, Targeting cancer stem cells, Expert Opin Ther Targets, 11:915-927, 2007
Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, C aceres-Cortes J, Minden M, Paterson B, Caligiuri MA, Dick JE, A cell initiating human acute myeloid leukaemia after transplantation into SCID mice, Nature, 367; 645-648, 1994.
Lessard J, Sauvageau G. Bmi-1 determines the proliferative capacity of normal and leukaemic stem cells. Nature. 2003 May 15;423(6937):255-60. Epub 2003 Apr 20.
36
Lobo NA, Shimono Y, Qian D, Clarke MF, The biology of cancer stem cells, Annu Rev Cell Dev Biol, 23: 675 -99, 2007.
Major AG, Pitty LP, Farah CS. Cancer stem cell markers in head and neck squamous cell carcinoma. Stem Cells Int. 2013;2013:319489. doi: 10.1155/2013/319489. Epub 2013 Mar 3.
Miyauchi S, Moroyama T, Sakamoto T, Okamoto T, Takada K. Establishment of human tumor cell line (Ueda -1) derived from squamous cell carcinoma of the floor of the mouth, Jpn. J. Oral. Maxillofac. Surg, 31: 1347-1351, 1985
Miyauchi S, Moroyama T, Kyoizumi S, Asakawa J, Okamoto T, Takada K, Malignant tumor cell line produced interleukin -1-like factor, In Vitro cell, 24; 753-758, 1988.
Nishii T, Yashiro M, Shinto O, Sawada T, Ohira M, Hirakawa K, Cancer stem cell-like SP cells have a high adhesion ability to the peritoneum in gastric carcinoma, Cancer Sci, 100:1397 -1402, 2009.
Okamoto T, Tani R, Yabumoto M, Sakamoto A, Tanaka K, Sato GH, Sato JD, Effect of insulin and transferin on the generation of lymphokine-activated killer cells in serum-free medium, J Immunol Methods, 195: 7-14, 1996.
37
Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor. Cell. 1983 Jul;33(3):967 -78.
Reya T, Morrison SJ, Clarke MF, Weissman IL, Stem cells, cancer, and cancer stem cells, Nature, 414: 105 -111, 2001.
Sato JD, Kawamoto T, Okamoto T, Cholesterol requirement of P3-X63-Ag8 and X63-Ag8,653 mouse myeloma cells for growth in vitro, J Exp Med, 165: 1761-6, 1987.
Schatton T, Frank NY, Frank MH, Identification and targeting of cancer stem cells, Bioessays, 31: 1038 -1049, 2009
Tan BT, Park CY, Ailles LE, Weissman IL, The can cer stem cell hypothesis: a work in progress, Lab Invest, 86:1203 -1207, 2006.
Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S, Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007 Nov 30;131(5):861-72.
Tateno H, Onuma Y, Ito Y. Live-cell imaging of human pluripotent stem cells by a novel lectin probe rBC2LCN.Methods Mol Biol. 2014;1200:313-8. doi: 10.1007/978-1-4939-1292-6_26.
38
Tateno H, Matsushima A, Hiemori K, Onuma Y, Ito Y, Hasehir a K, Nishimura K, Ohtaka M, Takayasu S, Nakanishi M, Ikehara Y, Nakanishi M, Ohnuma K, Chan T, Toyoda M, Akutsu H, Umezawa A, Asashima M, Hirabayashi J. Podocalyxin is a glycoprotein ligand of the human pluripotent stem cell -specific probe rBC2LCN. Stem Cells Transl Med. 2013 Apr;2(4):265-73. doi: 10.5966/sctm.2012-0154. Epub 2013 Mar 22.
Tateno H, Onuma Y, Ito Y, Minoshima F, Saito S, Shimizu M, Aiki Y, Asashima M, Hirabayashi J. Elimination of tumorigenic human pluripotent stem cells by a recombinant lectin-toxin fusion protein. Stem Cell Reports. 2015 May 12;4(5):811 -20. doi: 10.1016/j.stemcr.2015.02.016. Epub 2015 Apr 9.
Trumpp A, Wiestler OD, Mechanisms of Disease: cancer stem cells--targeting the evil twin, Nat Clin Pract Oncol, 2008 5 : 337-47, 2008.
Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007 Jun;9(6):654-9. Epub 2007 May 7.
Visvader JE, Lindeman GJ, Cancer stem cells in solid tumors; accumulating evidence and unresolved questions, Nat Rev Cancer,
39
8: 755-768, 2008.
Walter JD, Hunter M, Cobb M, Traeger G, Spiegel PC. Thiostrepton inhibits stable 70S ribosome binding and ribosome -dependent GTPase activation of elongation factor G and elongation factor 4. Nucleic Acids Res. 2012 Jan;40(1):360 -70. doi: 10.1093/nar/gkr623. Epub 2011 Sep 9.
Wright AJ, Andrews PW. Surface marker antigens in the characterization of human embryonic stem cells. Stem Cell Res. 2009 Jul;3(1):3-11. doi: 10.1016/j.scr.2009.04.001. Epub 2009 Apr 8.
Wu C, Alman BA, Side population cells in human cancers, Cancer Lett, 268: 1-9, 2008.
Yasuhiko Kizuka, Shinobu Kitazume, Kyohei Okahara, Alejandro Villagra, Eduardo M. Sotomayor and Naoy uki Taniguchi. “ Epigenetic regulation of a brain -specific glycosyltransferase N-acetylglucosaminyltransferase -IX (GnT-IX) by specific chromatin modifiers” . The Journal of Biological Chemistry,2014,doi: 10.1074/jbc.M114.554311
40
Comparative marker analysis of extracellular vesicles in different human cancer types. J Extracell Vesicles. 2013 Jun 18;2. doi: 10.3402/jev.v2i0.20424. eCollection 2013.
Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Samp ath J, Morris JJ, Lagutina I, Grosveld GC, Osawa M, Nakauchi H, Sorrentino BP, The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of the side -population phenotype, Nat Med, 7: 1028-34, 2001.
41
42
43
図 3 : 無 血 清 培 養 下 に お け る 各 種 癌 細 胞 株 に お け る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 存 在 率
44
図 4 : rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 ソ ー テ ィ ン グ 後 に ラ イ ブ セ ル イ メ ー ジ ン グ に て rBC2LCN 認 識 糖 鎖 の 発 現 状 態 を 検 討
45
図 5 : rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 を ソ ー テ ィ ン グ 後 に 一 継 代 し た 際 の rBC2LCN 認 識 糖 鎖 陽 性 細 胞 存 在 比 の 検 討
46
図 6 : rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 の 無 血 清 単 層 培 養 系 に お け る 増 殖 能
47
図 7:rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 細 胞 に お け る sphere 形 成 能 検 討
図 8 :
48
図 9 : rBC2LCN 認 識 糖 鎖 陽 性 及 び 陰 性 細 胞 由 来 sphere 中 に お け る rBC2LCN 認 識 糖 鎖 陽 性 細 胞 存 在 動 態
49
表 1 : DNA マ イ ク ロ ア レ イ に よ り rBC2LCN 認 識 糖 鎖 陽 性 細 胞 に お い て 陰 性 細 胞 に 比 較 し て 2 0 倍 以 上 発 現 亢 進 の 見 ら れ た 遺 伝 子 群
50
図 1 0 : rBC2LCN 認 識 糖 鎖 陽 性 細 胞 と 陰 性 に お け る IGF-2 発 現 の 検 討 ( 左 : droplet digital PCR 法 に よ る 解 析 , 右 : ウ エ ス タ ン ブ ロ ッ ト
51
図 1 1 : rBC2LCN 認 識 糖 鎖 陽 性 細 胞 お よ び 陰 性 細 胞 に お け る IGF-Akt pathway の リ ン 酸 化 状 態 評 価
52
図 1 2 : rBC2LCN 認 識 糖 鎖 陽 性 細 胞 お よ び 陰 性 細 胞 に お け る IGF-Akt pathway の リ ン 酸 化 状 態 評 価
53
図 1 3 : IGF-2 及 び IGF-2 中 和 抗 体 の rBC2LCN 認 識 糖 鎖 陽 性 及 び 陰 性 細 胞 の sphere 形 成 数 に 及 ぼ す 影 響