Short-chain dehydrogenase/reductase superfamily
に属する酵素の合目的改変に関する研究
山梨大学大学院
医学工学総合教育部
博士課程学位論文
平成
27
年
3
月
標 智仁
目 次
I 緒論 1
II 本論 9
第 1 章 meso-2,3-butanediol dehydrogenase 由来部位特異的変異L-2,3-butanediol
dehydrogenase の構造解析と基質結合による構造変化 9 1.1 緒言 . . . . 9 1.2 実験方法 . . . . 9 1.2.1 発現用プラスミドの構築と形質転換株の作成および培養. . . . 9 1.2.2 無細胞抽出液の調製と酵素精製 . . . 11 1.2.3 タンパク質濃度の測定 . . . 12 1.2.4 酵素活性の測定 . . . 12 1.2.5 結晶化 . . . 13 1.2.6 X線回折像の撮影と構造解析 . . . 13 1.3 結果および考察 . . . 14 1.3.1 酵素の精製 . . . 14 1.3.2 3PLBDHの触媒効率 . . . 14 1.3.3 3PLBDHおよびBsLBDHの結晶化 . . . 19 1.3.4 X線回折像の撮影と構造解析 . . . 21 1.3.5 3PLBDHの基質結合サイト . . . 26 1.3.6 BsLBDHの基質結合による構造変化 . . . 26 1.3.7 3PLBDHおよびBsLBDHにおけるAPR複合体の基質結合サイト の比較とKmに関する考察 . . . 29 1.4 総括 (1章) . . . 29
第 2 章 meso-2,3-butanediol dehydrogenase 由来部位特異的変異L-2,3-butanediol dehydrogenase の触媒能向上のための変異の検討 34 2.1 緒言 . . . 34 2.2 方法 . . . 34 2.2.1 発現用プラスミドの構築と形質転換株の作成および培養. . . 34 2.2.2 無細胞抽出液の調製と酵素精製 . . . 34 2.2.3 タンパク質濃度の測定 . . . 35 2.2.4 酵素活性の測定 . . . 35 2.2.5 結晶化 . . . 35
2.2.6 X線回折像の撮影と構造解析 . . . 35 2.3 結果および考察 . . . 36 2.3.1 酵素の精製 . . . 36 2.3.2 3PLBDH-F212Yの触媒効率 . . . 36 2.3.3 3PLBDH-F212Yの結晶化 . . . 36 2.3.4 X線回折像の撮影と構造解析 . . . 38 2.3.5 3PLBDHおよび3PLBDH-F212YのAPR複合体結晶構造の基質結 合サイトの比較 . . . 38 2.4 総括 (2章) . . . 40
第 3 章 L-2,3-butanediol dehydrogenase と meso-2,3-butanediol dehy-drogenase からのドメインキメラL-2,3-butanediol dehydrogenase の構造解析 45 3.1 緒言 . . . 45 3.2 方法 . . . 45 3.2.1 発現用プラスミドの構築と形質転換株の作成および発現. . . 45 3.2.2 無細胞抽出液の調製と酵素精製 . . . 45 3.2.3 タンパク質濃度の測定 . . . 45 3.2.4 酵素活性の測定 . . . 45 3.2.5 酵素サブユニットの生物学的単位の決定 . . . 46 3.2.6 結晶化 . . . 46 3.2.7 構造解析. . . 46 3.3 結果および考察 . . . 46 3.3.1 酵素の精製と生物学的単位 . . . 46 3.3.2 cLBDHの結晶化 . . . 48 3.3.3 X線回折像の撮影と構造解析 . . . 48 3.3.4 KpmBDH、BsLBDHおよびcLBDHの全体構造の比較 . . . 49 3.3.5 KpmBDH、BsLBDHおよびcLBDHのNAD+結合サイトの比較 . 54 3.3.6 cLBDH (NAD+複合体)結晶構造とBsLBDH (NAD+、BME複合 体)結晶構造における基質結合サイトの比較 . . . 54 3.3.7 cLBDH (NAD+複合体)と3PLBDH (APR複合体)の結晶構造の 比較 . . . 56 3.3.8 cLBDHのIle140とPhe146の側鎖配向に影響する残基の推定 . . . 56 3.3.9 cLBDHの基質認識の緩みとBsLBDHの光学異性認識機構に関する 考察 . . . 57 3.4 総括 (3章) . . . 60 第 4 章 ドメインキメラL-2,3-butanediol dehydrogenase の基質認識能向上 のための変異の検討 61 4.1 緒言 . . . 61 4.2 方法 . . . 61
4.2.1 発現用プラスミドの構築と形質転換株の作成および培養. . . 61 4.2.2 無細胞抽出液の調製と酵素精製 . . . 62 4.2.3 タンパク質濃度の測定 . . . 62 4.2.4 酵素活性の測定 . . . 62 4.2.5 結晶化 . . . 62 4.2.6 構造解析. . . 63 4.3 結果および考察 . . . 63 4.3.1 酵素の精製 . . . 63 4.3.2 各cLBDH変異体の触媒効率. . . 64 4.3.3 各cLBDH変異体の結晶化 . . . 72 4.3.4 X線回折像の撮影と構造解析 . . . 75 4.3.5 cLBDH変異体の基質結合サイト周辺とBsLBDHおよびcLBDHに おける比較 . . . 87 4.3.6 cLBDHにおけるBMEの阻害機構 . . . 94 4.3.7 BsLBDHの触媒効率および基質認識におけるQ軸対称での二量体 形成の意味 . . . 98 4.4 総括 (4章) . . . 99 第 5 章 L-2,3-butanediol dehydrogenase の安定性に関係する要因 101 5.1 緒言 . . . 101 5.2 方法 . . . 101 5.2.1 発現用形質転換株の培養 . . . 101 5.2.2 無細胞抽出液の調製と酵素精製 . . . 101 5.2.3 タンパク質濃度の測定 . . . 101 5.2.4 酵素活性の測定 . . . 101 5.2.5 立体構造の比較 . . . 102 5.3 結果および考察 . . . 102 5.3.1 BsLBDH、KpmBDHおよびcLBDHの熱安定性と保存安定性 . . . 102 5.3.2 BsLBDH、KpmBDHおよびcLBDHの安定性と立体構造について . 102 5.3.3 BsLBDHの安定性に及ぼす塩濃度の影響 . . . 104 5.3.4 cLBDH部位特異的変異体類の熱安定性と保存安定性 . . . 106 5.3.5 BsLBDHの安定性におけるQ軸対称での二量体形成の意味. . . 108 5.4 総括 (5章) . . . 110
III 総括 113
IV 謝辞 115
V 関連論文 116
I
緒論
生体触媒である酵素は無機触媒に比べ、高触媒効率、厳格な基質特異性、それに温和な 反応条件等に特徴があり、特に生体関連の有用物質生産に適している。しかし、酵素を産 業利用する上で、高い機能が求められる場合が多いものの、そのような酵素は自然界から 容易に得ることは出来ない。一方、現在は酵素機能を向上させるために、アミノ酸配列を 操り、触媒能向上や基質選択性、そして安定性などで不足する機能を合理的に補填する試みも日常的行われ、多くのタンパク質工学的知見が集積されつつある[Penning and Jez,
2001; Luetz et al., 2008; Jemli et al., 2014; Li and Cirino, 2014; Yang et al., 2014]。しか し、実際には目的にあった酵素を自在に作り出す迄には未だ多くの課題があり、限られた 改変にしか至っていない。
当研究室でBr. saccharolyticumより見出されたL-2,3-butanediol dehydrogenase
(L-BDH)も、そのような酵素の一つであり、利用に耐える安定性を付与すべく種々の試みが
なされてきた[Otagiri et al., 2010; Shimegi et al., 2011]。2,3-butanediol dehydrogenase
(BDH)はNAD+を補酵素とし、2,3-butanediol (BD)をacetoin (AC)に変換する酵素で
あるが、基質であるBDには3つの光学異性体 (D体、L体、meso体)が存在し、BDH についても種々の基質特異性を持つものが存在する (図1)。BDの異性体は、特に、医 薬品や農薬等の不斉合成におけるキラルパーツとしての利用が期待されている[Liu and Hogberg, 2001]。有機化学合成による光学異性体の生産は、一般にラセミ混合物からの光 学異性体分離用カラムを用いたクロマトグラフィーによる分離で行われるが、カラムが 高価であり、また一方のエナンチオマーを捨てることになるため、コスト的に商業生産 には適していない。そのため発酵等生物学的な光学異性体生産が注目されている。BDは 多くの微生物でその生産が知られており、3つの光学異性体のうちD-BD、meso-BDはそ
れぞれPaenib. polymyxa、Kl. pneumoniae等が発酵代謝物として大量に生産し[Stanier and Adams, 1944; Taylor and Juni, 1960]、その生成経路が明らかにされている[Ui et al.,
1986, 1984]。しかしL-BDについては、Kl. pneumoniae等の、meso-BDを生産する微生
物が副生成物として僅かに生成するのみであり、L-BDを代謝物として主要に生産する微
生物は見つかっていない。そこで、L-BDを代謝工学的に生産しようとする試みがこれま
でになされてきた。L-BDの前駆体であるL-ACについては,Kl. pneumoniae IAM1063
から単離されたKpmBDH[Ui et al., 1997]を大腸菌に導入することでdiacetyl (DA)から の生産が可能となった[Ui et al., 1999]。KpmBDHはmeso-BDのC2に結合する(R)-配
向ヒドロキシ基を厳格に認識し、C3のカルボニル基を (S)-配向のヒドロキシ基に還元す
る酵素であるが、この方法はKpmBDHはC2がカルボニル基の場合でも基質結合を許す
ため、DAをL-ACに還元できる性質を利用したものである。さらにBr. saccharolyticum
C-1012に、L-BDのC2に結合する(S)-配向ヒドロキシ基を厳格に認識し、L-ACをL-BD
に触媒する酵素L-BDHが発見されたため[Ui et al., 1998; Takusagawa et al., 2001]、こ れを用いることでL-BDの生産も可能となった[Ui et al., 2001, 2004]。しかし、KpmBDH
が安定な酵素である一方、L-BDHは安定性が低く、このことがL-BDの代謝工学的生産
における問題とされていた。
BsLBDHは安定性が低いため、精製も困難な酵素であったが、Otagiriらによって結晶
図1: BDHが触媒する反応.
複合体として構造解析された[Otagiri et al., 2010]。BsLBDHはそのアミノ酸配列から、 先にクロニーングされたKpmBDHと同じShort-chain dehydrogenase/reductase (SDR) superfamilyのsubclass ’classical’に属し[Jornvall et al., 1995]、その相同性が50%であ ることが分かっていたが[Ui et al., 1998] (図2)、構造解析の結果、先に三体複合体とし て構造解析されていたKpmBDHと同様、SDR superfamilyの’classical’に属する酵素の 典型的なトポロジーを形成し (図3)、BsLBDHとKpmBDHの立体相同性が高いことが 明らかになった (図4-A、B)。両者の基質結合サイトにおいて、構成残基の違いはIle142 (KpmBDHのGln140に対応)、Phe148 (KpmBDHのAsn146に対応)の2残基のみであっ たが、192番残基(KpmBDHのGln190番に対応)は同じTrpでありながら側鎖配向が異 なっており、結晶構造内の阻害剤BMEの結合から、KpmBDHはGln140の側鎖カルボニ ル基O原子がmeso-BDのC2(R)-配向ヒドロキシ基と、BsLBDHはTrp192の側鎖イン ドール基のN原子がL-BDのC2(S)-配向ヒドロキシ基と水素結合することでそれぞれBD のC2キラル配向を認識していると推測された。 そこでBsLBDHとKpmBDHの立体相同性の高さと基質結合サイトの違いに着目し、 安定性が高いL-BDHの創製が2つの手法によって検討された[田草川, 2004]。一つは部位 特異的変異による基質認識の改変である (図5)。先ずBsLBDHの基質結合サイトを参考
に、KpmBDHにGln140をIleに、Asn146をPheとする変異を与えたところ
(KpmBDH-Q140I/N146F、2PLBDH)、meso-BDよりもL-BDに大きな活性を示したが、BDのC2キ ラル配向の認識が緩い酵素であった。そこでさらにTrp190の配向を変えるため、Trp190 に対して基質結合サイトとは反対側にあるThr209をPheに変異させたところ (KpmBDH-Q140I/N146F/T209F、3PLBDH)、BDのC2 (S)-配向ヒドロキシ基を厳格に認識する酵 素となった。このことから、基質結合サイトに直接面してない残基が基質結合に大きな影 響を与えていることが示された。しかし、3PLBDHのL-BDに対するKmは高く、触媒 効率に関して課題が生じた[田草川, 2004]。もう一つはドメイン交換による基質認識の改 変である。BsLBDH、KpmBDHの単量体は、7つの平行βシート(βA-G)を6つのαヘ リックスが取り囲み、さらに2つの短いαヘリックス(αFG1、2)が付随している(図4)。
図2: BsLBDHとKpmBDHのアミノ酸配列のアライメント。
図 3: BsLBDHとKpmBDHに共通する典型的なSDR superfamily ’classical’に属する酵
図 4: BsLBDH (NAD+、BME複合体)結晶とKpmBDH (NAD+、BME複合体)結晶の
単量体構造と基質結合サイト。A、BはそれぞれBsLBDH、KpmBDHの単量体構造。a、
図 5: BsLBDH (NAD+、BME複合体)結晶とKpmBDH (NAD+、BME複合体)結晶の 基質結合サイトの比較。実線で囲まれた残基は基質と直接相互作用すると推測されている
残基。赤枠で囲まれた残基はKpmBDHとBsLBDHで異なる残基で、部位特異的変異の
この7つの平行β シートのうちの6本 (βA-F)と4つのαヘリックス (αB、C、E、F)
は’Rossmann fold’[Rossmnn et al., 1975]と呼ばれるジヌクレオチド結合モチーフを形成
し、単量体の中央、平行βシートのC末端側にNAD (H)が結合する。基質結合サイトは、
長い2本のαヘリックス(αヘリックスE、F)とαFG1、2で構成される基質結合ループ [Ghosh et al., 1991; Tanaka et al., 1996b; Ito et al., 2006]と呼ばれる部分によって形成さ
れている。この構造をαヘリックスとβシートで構成される剛な「基盤」と、基質結合空 間を構成する「3つの葉」と見立てた。そして3つの葉をそれぞれドメインA (KpmBDH の84-118番残基、BsLBDHの86-120番残基)、ドメインB (KpmBDHの135-161番残 基、BsLBDHの137-163番残基)、ドメインC (KpmBDHの182-236番残基、BsLBDHの 184-238番残基)、それ以外をドメインDと定義して (図6)、ドメインDをKpmBDH由 来、「3つの葉」ドメインをBsLBDH由来のものとしたキメラ酵素cLBDHが創製された (図7)。このcLBDHの安定性はKpmBDHに及ばないもののBsLBDHよりも高く、L-BD に対するKmがBsLBDHと同程度となり、ドメインDが安定性に強く影響を及ぼすこと、 そしてドメイン交換による酵素改変が有用であることが示された。しかし、BDのC2キ ラル配向の認識が緩く、ドメイン交換による基質認識の改変にも課題が生じた。 図 6: BsLBDHとKpmBDHのドメイン定義。AはBsLBDHで紫はドメインA、水色は ドメインB、黄色はドメインC。BはKpmBDHで緑はドメインD。 上記のように、3PLBDHとcLBDHはともに課題が存在していたが、3PLBDHは触媒 効率と構造の関係について、cLBDHは基質特異性および安定性と構造の関係について考 察する上で興味深い酵素である。 以上の背景から、本論文では3PLBDHとcLBDHの 構造解析を行い、その情報を元に、触媒効率、基質特異性そして安定性に優れたL-BDH の創製を試みるとともに、その過程で酵素機能と構造の関係について新たな知見を得るこ とを目的とした。 以下に、本論文を構成する5項の章について概要を記した。
図7: cLBDHの予想トポロジー。赤と青、黄色はそれぞれBsLBDH由来のドメインAと B、C。緑はKpmBDH由来のドメインD。 第1章では、KpmBDHの基質結合サイトを直接構成する残基およびその周辺残基に 変異を与えることによって創製されたL-BDH (3PLBDH)のX線結晶構造解析を行い、 3PLBDHの結晶構造を決定した。またBsLBDHと補酵素NAD+のアナログとの複合体 の構造解析の結果からBsLBDHの補酵素および基質結合時の構造変化と、3PLBDHの触 媒効率が低い原因について考察した。 第2章では、3PLBDHの構造情報に基づき高触媒効率を得るための変異を検討した。ま た、この変異によって得られたL-BDHの構造解析を行い、3PLBDHの触媒効率が低い原 因について考察した。 第3章では、KpmBDHとBsLBDHのドメイン交換によって創製されたL-BDH (cLBDH) のX線結晶構造解析を行い、cLBDHの結晶構造を決定した。この結果をBsLBDHの結 晶構造と比較することで、cLBDHの基質認識が厳格でない原因と BsLBDHの基質結合 様式を考察した。 第4章では、cLBDHの構造情報を基にBDのC2キラル配向認識への影響残基に変異 を加え、cLBDHよりBDのC2(S)-配向ヒドロキシ基を厳格に認識する酵素の創製を検討 した。この変異により創製されたL-BDHの構造解析を行い、この情報からBsLBDHの基 質結合に影響を及ぼす要因を考察した。 第5章では、cLBDHとKpmBDHおよびBsLBDHの結晶構造を比較し、BsLBDHの 安定性に影響を及ぼす要因を考察した。また、cLBDHの部位特異的変異により新たに創 製したL-BDHの安定性を評価し、その構造解析結果と照査することで、BsLBDHの安定 性に影響を及ぼす要因を考察した。 本論文は、SDR familyに属するBDHに種々の改変を加え、これらのX線結晶構造解 析を行い、得られた構造を詳細に検討することで、触媒効率、基質特異性そして安定性に
優れたL-BDHの創製に至った経緯と、そこで得られた機能-構造間に関する新たな知見に ついて報告するものである。
II
本論
第
1
章
meso-2,3-butanediol dehydrogenase
由来部位特異的変
異
L-2,3-butanediol dehydrogenase
の構造解析と基質
結合による構造変化
1.1
緒言
3PLBDHはKpmBDHにQ140I/N146F/T209Pの3点の変異を加えることで、BDの C2(S)-配向ヒドロキシ基を厳格に認識しながらも、KmがBsLBDHの400倍となり、触 媒効率に問題のある酵素であった。この3点の変異の中で最も興味深いのがT209Pの変 異である。この変異は、BsLBDH (NAD+、BME複合体)の構造解析から、基質である L-BDと直接相互作用すると推測されているTrp190 (BsLBDHのTrp192)の側鎖配向を変 更することを目的としており、基質結合サイトに対しては間接的な変異である。本章では、 3PLBDHの構造解析を行い、これを既に構造解析されているKpmBDH (NAD+、BME 複合体)と比較することで、Trp190の側鎖配向が変更されているかを検証し、BsLBDH (NAD+、BME複合体)と比較することで3PLBDHの低い触媒効率の原因を考察した。こ の過程でBsLBDHの補酵素および基質結合時の構造変化についても新たな知見が得られ たので、これについても考察した。1.2
実験方法
1.2.1 発現用プラスミドの構築と形質転換株の作成および培養3PLBDH発現用形質転換株はプラスミドベクターにpET-11a、宿主にEscherichia coli
BL21 (DE3)を用いた。3PLBDH発現用プラスミドは次に示す方法により作成した。3PLBDH
をコードする遺伝子がクローニングされているプラスミドpMQ140I/N146F/T209F[田草
川, 2004]を鋳型DNAとし、プライマーMBDH-F、MBDH-R (表1.1)を用いてPCRを
行った。プライマーはPCR増幅断片の5’末端側に制限酵素NdeIの認識サイト、3’末端
側にBamHIの認識サイトが付与されるように設計した。DNA polymeraseにはPhusion
High-Fidelity DNA polymerase (Finnzymes)を使用し、表1.2に示すサイクル (アニーリ
ング温度67◦C)でPCRを行った。
PCR増幅産物をWizard SV Gel and PCR Clean-Up System (Promega)を用いて精製 した。精製したPCR増幅産物をBamHI (TaKaRa)で30◦Cにて消化し、エタノール沈 殿後、NdeI (TaKaRa)で37◦Cにて消化した。これを1%アガロースゲルで電気泳動し、
表 1.1: 3PLBDHおよびBsLBDH遺伝子断片増幅用プライマー配列 名前 配列 MBDH-F 5’-AAAGGACATATGAAAAAAGTCGCACTTG-3’ MBDH-R 5’-TTTGGATCCTTAGTTAAATACCATCCCG-3’ LBDH-F 5’-AGTCATATGAGCAAAGTTGCAATGG-3’ LBDH-R 5’-AAAGGATCCTTAGTTGTAGAGCATGC-3’ 下 線 は MBDH-F、L-BDH-F は 制 限 酵 素 NdeI、MBDH-R、 LBDH-R は BamHI の認識サイトを示す。 表1.2: 3PLBDHおよびBsLBDH遺伝子断片増幅用PCRサイクル 初期変性 94◦C 120 sec. 変性 94◦C 15 sec. アニーリング x◦C 15 sec. 伸長 72◦C 45 sec. 25 cycle 最終伸長 72◦C 300 sec アニーリング温度xは本文中に記載した。
ターのpET-11aも同様にNdeI、BamHIによる制限酵素処理を行い、電気泳動後にゲルよ
り抽出し、精製した。精製したPCR増幅産物とベクターをLigation HIGH II (TOYOBO) を用いて連結した。
コンピテントセルは次に示すCaCl2[Cohen et al., 1972]による処理で作成した。E. coli
BL21 (DE3)をLB培地(表1.3) 5 mLに寒天培養より1 loop植菌し、30◦C、200 rpmに て一晩振とう培養した。この培養液をLB培地 200 mLに1%植菌し、30◦C、165 rpmに てOD600=0.7付近になるまで培養した。氷中で10分間冷却した後、培養液を4◦C、1,500 ×g で遠心し、上清を取り除いた。沈殿した菌体を100 mM MgCl2 100 mLに懸濁後、 4◦C、1,500 ×g で遠心し、上清を取り除いた。沈殿した菌体を10%グリセロールを含む 100 mM CaCl2 100 mLに懸濁後、4◦C、1,500×g で遠心し、上清を取り除いた。この操 作を2回行った後、氷中に10分間静置し、その後4◦C、1,500×g で遠心し、上清を取り 除き、10%グリセロールを含む100 mM CaCl2 5 mLに懸濁し、使用するまで-80◦Cで凍 結保存した。 形質転換は次に示す方法で行った。コンピテントセル懸濁液100 µLに対しライゲーショ ン溶液 5 µL を加えて氷中で30分間冷却した。その後42℃で60秒間保持し、再び氷中 にもどし2分間冷却した。これに900 µLのLB培地を加え37℃で1時間培養した。培養 液は、4℃、15,000×g で1分間遠心し、上清を捨てた後、菌体を残った培地に懸濁して、 100 µ/mL アンピシリンを含むLB寒天培地 (表1.3)に塗布し、30℃で一晩培養した。 プラスミドの構築は塩基配列の決定により確認した。シングルコロニーを100 µg/mL アンピシリンを含むLB培地で培養し、プラスミドを抽出した。抽出にはQIAprep Spin
Miniprep Kit (QIAGEN)を使用し、添付のマニュアルに従った。プラスミドの塩基配列
は、T7 promoterまたはT7 terminatorプライマーを用い、株式会社 ファスマックに依頼
して決定した。
表1.3: LB培地組成 1% (w/v)トリプトン 0.5% (w/v)酵母エキス 1% (w/v)NaCl 寒天培地には1.5%寒天を加えた。 ドpBUD119[Ui et al., 1997]を用いてPCRを行い、3PLBDHと同様の手順で作成した 2PLBDH発現用プラスミドの構築および形質転換株の作成は、鋳型DNAにプラスミド pQ140I/N146F/T209F[田草川, 2004]を用いてPCRを行い、3PLBDHと同様の手順で作 成した BsLBDH発現用プラスミドの作成は、鋳型DNAにBsLBDHをコードする遺伝子が クローニングされているプラスミドPLBD2-119[田草川, 2004]、プライマーにLBDH-F、 LBDH-R (表1.1)を用い、表1.2に示すサイクル (アニーリング温度55◦C)でPCRを行 い、その後は3PLBDH発現用プラスミドの作成と同様の手順で作成した。形質転換株の 作成も3PLBDHと同様の手順で作成した。 各発現用形質転換株は次に示す方法で培養した。前培養は、18ϕ試験管中の5 mLのLB 培地をオートクレーブ(120◦C、20分間)後、アンピシリンナトリウムを100 µg/mLとな るように添加し、この培地に寒天培地で培養された菌体を白金耳で1 loop接種後、30◦C、 300 rpmにて16時間振とう培養した。 本培養は500 mL容坂口フラスコ中の100 mLの LB培地を前培養と同様にオートクレーブ後、100 µg/mLアンピシリンナトリウムとなるよ うに添加し、これに前培養液1 mLを接種した。30◦C、150 rpmにて培養し、OD600=0.5
付近で1 mM isopropyl β-D-1-thiogaractopyranoside (IPTG)を添加後、引き続き24時間 培養した。
1.2.2 無細胞抽出液の調製と酵素精製
3PLBDHの無細胞抽出液(Cell-Free Extract、CFE)は次に示す方法で調製した。各発現
用菌体培養液を冷却遠心器にて遠心分離 (4◦C、5,000×g、10分間)で集菌し、ペレットを
1 gあたり100 mM Tris-HClバッファ(pH 8.0) 10 mLに懸濁して菌体の洗浄を行った。再
び遠心分離(4◦C、5,000×g、10分間)して集菌後、ペレット1 gあたり100 mM Tris-HCl 緩衝液 (pH8.0) 10 ml に懸濁し、0.1mM phenylmethane sulfonyl fluoride (PMSF)を添 加後、超音波破砕機より菌体破砕した。菌体破砕液を遠心分離( 4◦C、38,900×g 、30分 間、2回) し、回収した上清をCFEとした。 3PLBDHの精製は硫安分画、疎水性相互作用クロマトグラフィ、強陰イオン交換クロマ トグラフィによって行った。硫安分画は35-55%飽和硫安による塩析物を遠心分離 (4◦C、 21,500×g 、30分間) で回収した。塩析物を100 mM Tris-HCl (pH8.0) 10 mLに溶解 し、硫安分画後タンパク質溶液とした。疎水性相互作用クロマトグラフィはクロマトグラ フィシステムにAKTApurifier (GE Healthcare)、カラムにHiTrap Phenyl HP 5 mL (GE Healthcare)を使用した。結合バッファには100 mM Tris-HCl 1 M (NH4)2SO4 (pH8.0)、
で行った。カラムの平衡化は結合緩衝液5 CVで行った。硫安分画後タンパク質溶液をシ リンジを用いて0.45 µm polyvinylidene difluorid (PVDF)製メンブレンフィルターでろ 過した後、カラムに吸着させ、結合バッファ5 CVでカラムを洗浄した。溶出は50 mM (NH4)2SO4/CVのグラジエント溶出で行い、溶出液を2 mL毎に分画した。12%ポリアク リルアミドゲルを用いたSDS-PAGEにより目的タンパク質が溶出されている画分を回収 し、疎水性相互作用クロマトグラフィ後タンパク質溶液とした。強陰イオン交換クロマト グラフィはカラムにHiTrap Q HP 5 mL (GE Healthcare)を使用し、疎水性相互作用クロ
マトグラフィ後タンパク質溶液を10 mM Tris-HCl (pH 8.0)で希釈することで電気伝導度
を下げ、吸着させた。結合バッファには10 mM Tris-HCl 1 M (pH8.0)、溶出バッファに
は10 mM Tris-HCl 1M NaCl (pH8.0)を用いて疎水性相互作用クロマトグラフィと同様の
手順で行った。精製後のタンパク質溶液はAmicon Ultra 15 mL 50K NMWL (Millipore)
を使用し、添付のマニュアルにしたがって約150 µLに濃縮し、使用するまで4◦Cで保存
した。
KpmBDH、2PLBDHの精製は3PLBDHと同様に行った。
BsLBDHは硫安分画で60%飽和硫安の上清を回収し、10 mM Tris-HCl 1 M (pH8.0)に
よって透析後、弱陰イオン交換クロマトグラフィによって精製した。弱陰イオン交換クロ マトグラフィはカラムにHiTrap DEAE HP 5 mL (GE Healthcare)を使用し、結合バッ
ファには10 mM Tris-HCl 1 M NaCl (pH8.0)、溶出バッファには10 mM Tris-HCl 1M
NaCl (pH8.0)を用いた。
1.2.3 タンパク質濃度の測定
タンパク質濃度はBradford法[Bradford, 1976]によって測定した。Bradford試薬には Quick StartTM Bradford Dye Reagent、1x (Bio-Rad)を用いた。Bradford試薬1 mLに
対してタンパク質溶液5、10または20 mL を添加し、10分間反応させた後、波長595 nmの吸光度を計測した。標準試料には牛血清アルブミン (BSA)を使用し、0.2、0.4、0.8 mg/mL BSA溶液20 µLにおける吸光度から検量線を作成し、サンプルのタンパク質濃度 を算出した。 1.2.4 酵素活性の測定 酸化反応酵素活性は補酵素NAD+のNADHへの還元を測定することで算出した。酵素
活性1 unit (U)は、1分間に1 µmolのNAD+をNADHに変換する酵素量として定義し
た。反応液を50 mMリン酸ナトリウム緩衝液(pH 8.0)、5 mM基質、1 mM NAD+とし、 タンパク質溶液100 µLを加えて全量1 mLに調製して、25℃で反応させた。波長340 nm の吸光度の変化から1分間当たりの増分を求め、NADHのモル吸光係数6,220 M−1·cm−1 より、NADHの1分間当たりの増加量を算出した。Kmおよびkcatは、Hanse-Woolfプ
1.2.5 結晶化 3PLBDH、BsLBDHの結晶化はハンギングドロップおよびシティングドロップ蒸気拡 散法で行った。タンパク質溶液は10または20 mg/mL、1 mg/mL NAD+またはADP-リ ボース (APR) となるように調製した。リザーバー液には沈殿剤としてPEG4000または PEG6000を15-30%、緩衝液として50または100 mM クエン酸バッファ (pH 4.5-5.5)、 MESバッファ(pH 5.5-6.5)、HEPESバッファ(pH 6.5-7.5)を用い、抗凍結剤として15%グ リセロールを添加してスクリーニングを行った。結晶化ドロップはタンパク質溶液 1 µL とリザーバ液1 µL を混合して作成し、20◦Cで保存した。 1.2.6 X 線回折像の撮影と構造解析 3PLBDH、BsLBDH結晶のX線回折像は高エネルギー加速器機構、物質構造科学研究
所、Photon Factory (PF)またはPhoton Factory Advanced Ring (PF-AR)にて撮影し
た。撮影条件は表1.4に示した。 回折斑点の指数付け、積分、スケーリングはHKL2000 (HKL Research Inc.)またはXDS [Kabsch, 2010]を用いた。 表1.4: X線回折像の撮影条件 Enzyme 3PLBDH bound APR BsLBDH bound APR
Beamline PF-AR NW-12A PF BL-5A
Detector Quantum210r Quantum315r
Wavelength 1.0000 1.0000
Exposure time (sec) 0.50 0.75
Oscillation angle (◦) 0.5 0.25
No. of frames 360 720
Temperature (K) 95 95
位相の決定は分子置換法で行い、プログラムは molrep[Vagin and Teplyakov, 2010]
(CCP4 program suite[Winn et al., 2011]) を用いた。サーチモデルは3PLBDH結晶構
造解析にはKpmBDH (PDB code 1GEG)、BsLBDH結晶構造解析にはBsLBDH (PDB code 3A28)を用いた。構造の精密化はphenix.refine (Phenix program suite[Adams et al.,
2010])およびCoot-0.6.2[Emsley and Cowtan, 2004]を用いた。立体構造の視覚化には
Py-Mol (version 1.4.1;Schrdinger, LLC. [http://www.pymol.org/])を用いた。ラマチャンド ラン解析はMolprobity[Chen et al., 1996; Lovell et al., 2003]を用いた。
1.3
結果および考察
1.3.1 酵素の精製 3PLBDH、KpmBDH、2PLBDH、BsLBDHの精製表を表1.5から1.5に、各精製段階 におけるSDS-PAGEを図1.1から1.4に示した。3PLBDH、KpmBDH、2PLBDHはそ れぞれ硫安分画、疎水性相互作用クロマトグラフィ、強陰イオン交換クロマトグラフィの 3段階、BsLBDHは硫安分画、弱陰イオン交換クロマトグラフィの2段階の精製により、 29 KDa付近に単一バンドを確認した。アミノ酸一次配列より推定される3PLBDH単量 体の分子量が26,655 Da、KpmBDHが26,642 Da、2PLBDHが26,609 Da、BsLBDHが27108 Daであり、BDに対してそれぞれ3.0 U/mg、440 U/mg、4.7 U/mg、96 U/mgの
活性を示すことから、このバンドがそれぞれ3PLBDH、KpmBDH、2PLBDH、BsLBDH によるものと確認された。
1.3.2 3PLBDH の触媒効率
表1.9にKpmBDH、2PLBDH、3PLBDHおよびBsLBDHの動力学的パラメータを示し
た。KpmBDHのmeso-BDに対するkcat/KmはL-BDの340倍であり、meso-BDの
C2(R)-配向ヒドロキシ基を厳格に認識する酵素であること、そしてKpmBDHへのQ140I/N146F の変異がmeso-BDに対するKmを著しく上昇させ、結果としてL-BDに対するkcat/Km がmeso-BDのそれを上回ったことで、BDのC2(R)-配向ヒドロキシ基よりも (S)-配向ヒ ドロキシ基を認識する酵素になったことが確認された。また、3PLBDHのmeso-BDに対 するKmは2PLBDHよりも高く、L-BDに対するkcat/Kmも1.7倍と高いことから、さ らに厳格にBDのC2(S)-配向ヒドロキシ基を認識することが確認されたが、L-BDに対す るKmがBsLBDHに比べ840倍、kcat/Kmで700倍であることから、触媒効率の劣る酵 素であることも確認された。 表 1.9: 3PLBDH、KpmBDH、2PLBDHおよびBsLBDHの動力学的パラメータ。 kcat (sec−1) Km (mM) kcat/Km (sec−1·mM−1) Ratio of kcat/Km Enzyme Substrate KpmBDH L-BD 10 ± 1 61 ± 3 0.17± 0.01 1 meso-BD 160± 10 2.9± 0.1 57 ± 7 340 2PLBDH L-BD 37 ± 1 94 ± 5 0.39± 0.02 1 meso-BD 74 ± 10 270± 10 0.27± 0.02 1.4 3PLBDH L-BD 45 ± 1 67 ± 3 0.67± 0.01 -meso-BD - > 400 - -BsLBDH L-BD 37 ± 3 0.08± 0.01 470± 40 960 meso-BD 0.99 ± 0.02 2.0± 0.1 0.49± 0.02 1
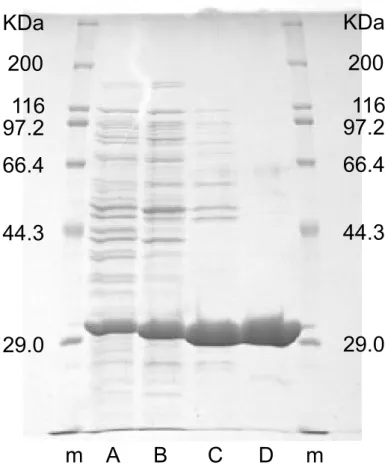
図1.1: 3PLBDHの精製段階におけるSDS-PAGE。12%ポリアクリルアミド、6×6 cmゲ ル使用。AはCFE、Bは硫安分画後、Cは疎水性相互作用クロマトグラフィ後、Dは強陰 イオン交換クロマトグラフィ後、mは分子量マーカー。各レーンタンパク質量5 µg供試。 表1.5: 3PLBDHの精製表 Process Activity (U) Protein (mg) Specific Activity (U/mg) Yield (%) Fold 無細胞抽出液 (CFE) 70 83 0.85 100 1.0 硫安分画 52 41 1.3 74 1.5 疎水性相互作用クロマトグラフィ 10 4 2.5 14 2.9 強陰イオン交換クロマトグラフィ 7.7 2.5 3 11 3.6
図1.2: KpmBDHの精製段階におけるSDS-PAGE。12%ポリアクリルアミド、6×6 cmゲ ル使用。AはCFE、Bは硫安分画後、Cは疎水性相互作用クロマトグラフィ後、Dは強陰 イオン交換クロマトグラフィ後、mは分子量マーカー。各レーンタンパク質量5 µg供試。 表1.6: KpmBDHの精製表 Process Activity (U) Protein (mg) Specific Activity (U/mg) Yield (%) Fold 無細胞抽出液 (CFE) 8300 93 90 100 1.0 硫安分画 13000 82 160 160 1.8 疎水性相互作用クロマトグラフィ 2100 5.2 410 25 4.6 強陰イオン交換クロマトグラフィ 970 2.2 440 12 4.9
図1.3: 2PLBDHの精製段階におけるSDS-PAGE。12%ポリアクリルアミド、6×6 cmゲ ル使用。AはCFE、Bは硫安分画後、Cは疎水性相互作用クロマトグラフィ後、Dは強陰 イオン交換クロマトグラフィ後、mは分子量マーカー。各レーンタンパク質量5 µg供試。 表1.7: 2PLBDHの精製表 Process Activity (U) Protein (mg) Specific Activity (U/mg) Yield (%) Fold 無細胞抽出液 (CFE) 39 85 0.46 100 1.0 硫安分画 10 36 0.28 26 0.6 疎水性相互作用クロマトグラフィ 9.6 3.1 3.1 25 6.8 強陰イオン交換クロマトグラフィ 8 1.7 4.7 21 10
図1.4: BsLBDHの精製段階におけるSDS-PAGE。12%ポリアクリルアミド、8×9 cmゲ ル使用。Aは培養液16 µL、BはCFE、Cは硫安分画後、Dは弱陰イオン交換クロマトグ ラフィ後、mは分子量マーカー。A以外、各レーンタンパク質量10 µg供試。 表1.8: BsLBDHの精製表 Process Activity (U) Protein (mg) Specific Activity (U/mg) Yield (%) Fold 無細胞抽出液 (CFE) 1900 55 34 100 1.0 硫安分画 1200 41 1.3 55 1.5 弱陰イオン交換クロマトグラフィ 750 7.9 96 41 2.8
1.3.3 3PLBDH および BsLBDH の結晶化 初期のスクリーニングでは、3PLBDHの結晶化は沈殿剤にPEG6000を、BsLBDHは 沈殿剤にPEG4000を使用した条件で、それぞれ針状の結晶や棒状の微結晶が確認された。 これらの条件を最適化することで、3PLBDHはタンパク質溶液が20 mg/mL、1 mg/mL NAD+、10 mM Tris-HCl (pH8.0)、リザーバー液が100 mMクエン酸バッファ(pH5.0)、 20%PEG6000、15%グリセロール、1% BMEの条件で、長さ約 200 µmの棒状の単結晶 が形成され (図1.5)、また、タンパク質溶液が20 mg/mL、10 mM Tris-HCl (pH8.0)、1
mg/mL APR、リザーバー液が50 mM MES酸バッファ(pH6.2)、27.5%PEG4000、15%グ
リセロールの条件では長さ約100 µmの棒状の単結晶が形成された(図1.6)。BsLBDHはタ ンパク質溶液が10 mg/mL、1 mg/mL APR、10 mM Tris-HCl (pH8.0)、350 mM NaCl、 リザーバー液が50 mM MESバッファ(pH5.0)、22.5%PEG6000、15%グリセロールの条 件で、長さ約 200 µmの棒状の単結晶が形成された (図1.7)。
図1.5: 3PLBDH結晶。タンパク質:20 mg/mL、1 mg/mL NAD+。リザーバー液:100 mM
図 1.6: 3PLBDH結晶。タンパク質:20 mg/mL、1 mg/mL APR。リザーバー液:50 mM MES酸バッファ(pH6.2)、27.5%PEG4000、15%グリセロール。
図 1.7: BsLBDH結晶。タンパク質:10 mg/mL、1 mg/mL APR。リザーバー液:50 mM
1.3.4 X 線回折像の撮影と構造解析 タンパク質溶液にNAD+とBMEを含む3PLBDH結晶は最高分解能1.92 ˚Aで回折像 が得られ、分子置換法により非対称単位に6個の単量体 (サブユニットA-F)を含んでい ることが明かになったが、サブユニットAからEではNAD+結合サイトに電子密度が確 認できたものの、ニコチンアミド環の電子密度が観測されなかった (図1.8-A)。これは結 晶化ドロップ中に存在したと考えられるNAD+の分解物APRが結合していたと推測され るが、物質を確定できないため、構造決定しなかった。また基質結合サイトにBMEと思 われる電子密度は観測されなかった。 図 1.8: A:タンパク質溶液にNAD+を含む3PLBDH結晶のNAD+結合サイトのFo− Fc 電子密度図。B:KpmBDH結晶のNAD+。 そこでタンパク質溶液にAPRを含む3PLBDHとBsLBDHの結晶を作成し、回折像の 撮影を経て構造解析を行った。表1.10に3PLBDH (APR複合体)結晶とBsLBDH (APR 複合体)結晶の回折データと精密化の統計値を示した。 3PLBDH (APR複合体)結晶のX線回折像は、X線波長1.0000 ˚Aで分解能2.55˚Aの データセットを得た。3PLBDH (APR複合体)結晶構造は空間群P 212121で決定され、非 対称単位に4個の単量体が含まれており、四量体を形成していた。3PLBDHの生物学的 単位は、加えた変異がサブユニット間の相互作用面ではないためKpmBDH同様に四量体 と考えられるが、3PLBDHは結晶中でも四量体で存在し(図1.9)、他の四量体を形成する SDR酵素と同様、222点群対称を形成していることが確認された[Tanaka et al., 2001]。 3PLBDH (APR複合体)結晶構造は、サブユニット A-Cについてはすべての残基の座 標を決定できたが、サブユニットDについては192-203番残基の座標は電子密度が不明瞭 なため決定できなかった。SDR familyの’classical’はαFG1-αFG2とその前後のループは 補酵素NAD+および基質をそれぞれの結合サイトに取り込むために開閉することが知ら
れている[Ghosh et al., 1994; Tanaka et al., 2001]。この領域は基質結合ループと呼ばれ、
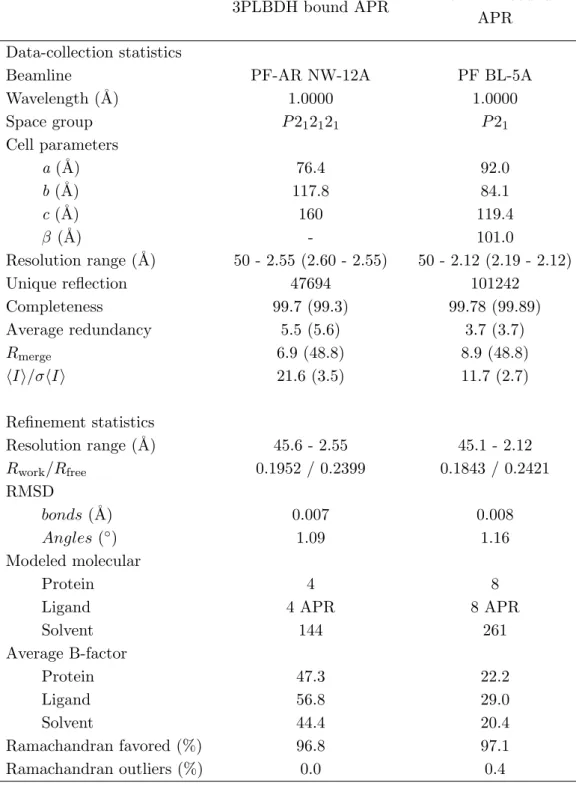
表 1.10: 回折データと精密化の統計値
3PLBDH bound APR BsLBDH bound
APR Data-collection statistics
Beamline PF-AR NW-12A PF BL-5A
Wavelength (˚A) 1.0000 1.0000 Space group P 212121 P 21 Cell parameters a (˚A) 76.4 92.0 b (˚A) 117.8 84.1 c (˚A) 160 119.4 β (˚A) - 101.0 Resolution range (˚A) 50 - 2.55 (2.60 - 2.55) 50 - 2.12 (2.19 - 2.12) Unique reflection 47694 101242 Completeness 99.7 (99.3) 99.78 (99.89) Average redundancy 5.5 (5.6) 3.7 (3.7) Rmerge 6.9 (48.8) 8.9 (48.8) ⟨I⟩/σ⟨I⟩ 21.6 (3.5) 11.7 (2.7) Refinement statistics Resolution range (˚A) 45.6 - 2.55 45.1 - 2.12 Rwork/Rfree 0.1952 / 0.2399 0.1843 / 0.2421 RMSD bonds (˚A) 0.007 0.008 Angles (◦) 1.09 1.16 Modeled molecular Protein 4 8
Ligand 4 APR 8 APR
Solvent 144 261 Average B-factor Protein 47.3 22.2 Ligand 56.8 29.0 Solvent 44.4 20.4 Ramachandran favored (%) 96.8 97.1 Ramachandran outliers (%) 0.0 0.4
図 1.9: 3PLBDH (APR複合体)結晶内の四量体構造。AはQ軸が紙面に垂直となる方向
から、BはR軸が紙面に垂直となる方向からみた四量体構造。赤がサブユニットA、青が
いる[Tanaka et al., 1996b; Alphey et al., 2005; Bakelar et al., 2013]。原子座標を決める
ことができなかったサブユニットDの192-203残基はαFG1およびそのC末端側のルー
プ、すなわち基質結合ループの一部であることから、他のSDR family酵素の構造解析と
同様に結晶中の揺らぎのために電子密度が不明瞭になったと考えられる。
各サブユニットを重ね合わせたところ、サブユニットA-Cは基質結合ループが閉じた
状態、すなわち閉構造であることが確認され[Oppermann et al., 2003; Paithankar et al.,
2007]、サブユニットDも基質結合ループのαFG2がサブユニットA-Cに重なるので閉構
造であることが推測された (図1.10)。サブユニットAに対するサブユニットB-DのCα
の平均二乗誤差 (Root mean square deviation、RMSD)がそれぞれ0.165、0.324、0.297 ˚ Aとわずかであることから、以後、サブユニットAを3PLBDH (APR複合体)結晶構造 の単量体閉構造として代表させた。 図1.10: 3PLBDH (APR複合体)結晶構造の各サブユニットの比較。赤がサブユニットA、 青がB、黄色がC、緑がD。すべてのサブユニットが閉構造を形成していたことが確認さ れた。 3PLBDH (APR複合体)の単量体構造を、部位特異的変異を与える前のKpmBDH (NAD+、 BME複合体)の単量体構造 (サブユニットA)と比較したところ、Q140I/N146F/T209F の変異はトポロジーを変化させるような大きな影響を与えなかったことが確認された (図 1.11)。 BsLBDH (APR複合体)結晶のX線回折像は、X線波長1.0000 ˚Aで分解能2.12 ˚Aの データセットを得た。3PLBDH結晶構造は空間群P 21で決定され、非対称単位に8個の単 量体が含まれており、既報のBsLBDH (NAD+、BME複合体)結晶構造とは空間群P 1が
図1.11: 3PLBDH (APR複合体)結晶とKpmBDH (NAD+、BME複合体)結晶の単量体 構造の比較。紫が3PLBDH、グレーがKpmBDH。CαのRSMD=0.769 ˚A。
異なるものの[Otagiri et al., 2010]、同様に2つの四量体を形成していた。BsLBDH (APR
複合体)結晶構造は、サブユニットCのみMet1とLys2の座標を、残る7つのサブユニッ
トはMet1の座標を、電子密度が不明瞭なため決定できなかった。ラマチャンドラン解析
で外れ値であった残基はすべてのサブユニットのIle148であり、これについては第5章で
述べる。
3PLBDH (APR複合体)結晶、BsLBDH (APR複合体)結晶ともに、NAD+結合サイト
にAPRと思われる電子密度を観測したため、APRの座標を決定した。
1.3.5 3PLBDH の基質結合サイト
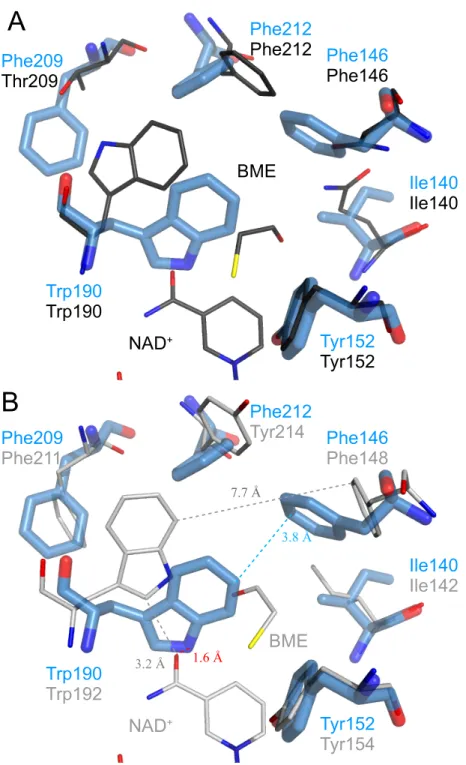
3PLBDH (APR複合体)結晶構造の基質結合サイトをKpmBDH (NAD+、BME複合
体)、BsLBDH (NAD+、BME複合体)結晶構造と比較した (図1.12)。KpmBDHとの比 較から、推察の通りTrp190の配向が変わり、インドール基のN原子が基質結合サイトに 面していることが確認された(図1.12-A)。しかし、BsLBDHとの比較から、2PLBDHお よび3PLBDHの触媒効率よりBDのC2キラル配向認識に関与することが確認されている Phe146、Ile140そしてTrp190の側鎖配向が異なっていることも確認された(図1.12-B)。 特にPhe146とTrp190の側鎖はは、それらに相当するBsLBDHのPhe148とTrp192の 側鎖よりも基質結合サイト側に大きく傾いていており、BsLBDH (NAD+、BME複合体) 構造ではPhe148とTrp192の側鎖の最短距離が7.7 ˚Aであったものが、3PLBDH (APR 複合体)結晶構造ではPhe146とTrp190の側鎖の最短距離が3.8 ˚Aであり、基質結合サイ トを狭くしていた (図1.12-B)。このことがタンパク質溶液にNAD+を含む3PLBDH結 晶で阻害剤BMEが結合されなかった原因であると推測された。また、BsLBDH (NAD+、 BME複合体)結晶構造において、Trp192のインドール環とNAD+のニコチンアミド環の 最短距離は3.2 ˚Aであり、3PLBDH (APR複合体)結晶構造を重ね合わせると、3PLBDH のTrp190側鎖とNAD+のニコチンアミド環の最短距離が1.6 ˚Aとなり、干渉することが 推測された (図1.12-B)。このことから、タンパク質溶液にNAD+を含む3PLBDH結晶 においてNAD+が結合されなかったことも、Trp190の配向によるものと推測された。 1.3.6 BsLBDH の基質結合による構造変化 3PLBDH (APR複合体)結晶構造との比較のため、BsLBDHもAPRとの複合体で結晶 化し、構造解析を行った。その結果、BsLBDHのTrp192の側鎖はNAD+およびBMEと の複合体結晶構造ではχ1 =−117、χ2 = 105であったが、APRとの複合体結晶構造では χ1=−91、χ2 = 96となり、基質結合サイトに内傾していた(図1.13)。また、その近傍の
Tyr214側鎖もNAD+およびBMEとの複合体結晶構造ではχ1 =−78、χ2 = 59、APRと
の複合体結晶構造ではχ1 =−101、χ2 = 80であった。すなわち、BsLBDHはNAD+お
よび基質を結合する時にTrp192とTyr214に構造変化を必要とすることが明かになった。
また、SDR famliy酵素は、NAD+のニコチンアミド環のNH2基とりん酸基が、基質結合
ループ上に存在するThr (BsLBDHではThr189)と水素結合を形成することで反応に必要
な閉構造が誘導されるとされているが[Oppermann et al., 2003; Paithankar et al., 2007;
Nakashima et al., 2009], 3PLBDH (APR複合体)結晶構造から、りん酸基との水素結合
図 1.12: Aは3PLBDH (APR複合体)結晶構造 (スカイブルー)とKpmBDH (NAD+、 BME複合体)結晶構造(ダークグレー)の基質結合サイトの比較。Bは3PLBDH (APR複 合体)結晶構造とBsLBDH (NAD+、BME複合体)複合体結晶構造 (ライトグレー)の基 質結合サイトの比較。
図 1.13: BsLBDH (APR複合体)結晶構造(黄色)と3PLBDH (NAD+、BME複合体)結
1.3.7 3PLBDH および BsLBDH における APR 複合体の基質結合サイトの比 較と Kmに関する考察 BsLBDH (APR複合体)結晶構造においてもTrp192は基質結合サイトに内傾していた が、Phe148とTrp192の側鎖の最短距離は6.9 ˚Aであり、3PLBDH (APR複合体)結晶構 造の3.8 ˚Aには至らなかった(図1.14)。 以上の結果からBsLBDHと3PLBDHのKmについて次のように考察した。 開構造時にリガンド(NAD+とL-BD)がそれぞれの結合サイト周辺に入り込み、NAD+ が酵素タンパク質と水素結合や疎水性相互作により徐々に閉構造を誘導していくが、閉構 造になる直前、BsLBDHのTrp192と3PLBDHのTrp190の側鎖配向はそれぞれのAPR 複合体結晶構造の配向が安定であり (図1.15-a)、この配向を平均として、熱運動のため 一定の分散を持った配向分布状態にあると考えられる (図1.16)。基質濃度が低くても、 BsLBDHはTrp192の側鎖配向の平均はリガント結合時の配向と異なるものの、そのほと んどがリガントを結合して閉構造を形成できる範囲にあるため、高い活性を示すと考えら れる。一方、3PLBDHはTrp190の側鎖配向の平均は基質結合サイトに内傾しているこ とにより、リガントを結合して閉構造を形成できるものも存在するが、多くはPhe146と の疎水性相互作用によりリガントを追い出してしまうため、活性が低いと考えられる (図 1.15-b、図1.16-a)。基質濃度が低いときは、リガンドが外れた後にほとんどの酵素分子 のTrpはAPR複合体結晶構造の配向、即ち安定な配向に戻ろうとするが、基質濃度が高 くなるとリガントが外れた直後に次のリガンドが入り込む機会が増えるため、リガントを 結合して閉構造を形成できるTrpの配向を持った酵素分子の割合が増えると考えられる。 BsLBDHは元々リガントを結合して閉構造を形成できる酵素分子が多いため、基質濃度 が高くなってもその数は大きく変わらない、即ち基質濃度が少し高くなっただけでVmax 付近に至るが (図1.16-b)、 3PLBDHは、Vmax付近に至るまでにさらに高い基質濃度が 求められると考えられる (図1.16-c)。 以上のことから、BsLBDHのKmは低く、3PLBDHは高いと考察した。
また、3PLBDH (APR複合体)結晶構造ではPhe146とPhe212の側鎖間の最短距離は
3.3 ˚Aであったことから、3PLBDHはNAD+と基質を結合しない状態で閉構造となるとき
に、Phe146とPhe212の側鎖間にも疎水性相互作用が生じることが推測された。3PLBDH
のPhe212は、BsLBDHでは疎水性尺度が低いTyr214に対応するため、Phe146とTrp190
の疎水性相互作用に寄与していることが推測された。
1.4
総括
(1
章
)
KpmBDHの基質結合サイトを直接構成する残基およびその周辺残基に変異を与えるこ とによって創製された3PLBDHはBDのC2(S)-配向ヒドロキシ基を認識する酵素であった が、Kmが高いため触媒効率が低かった。その原因を探るべく3PLBDHの結晶を、NAD+ とBMEの存在下で作成し、構造解析を行った。しかし、NAD+のニコチンアミド環と BMEによる電子密度が観測されず、L-BDと水素結合してC2ヒドロキシ基の光学異性認 識をするとされるTrp190の側鎖が基質結合サイト内側に傾き、基質結合サイトを狭くして いた。このため3PLBDHとBsLBDHをそれぞれAPRとの複合体で結晶化し、構造解析図 1.14: 3PLBDH (APR複合体)結晶構造 (スカイブルー)とBsLBDH (APR複合体)結
図 1.15: 推測されるBsLBDHと3PLBDHの開構造から閉構造に遷移するときの構造変 化。曲線の矢印はリガントまたは側鎖の動き、黒の点線は水素結合、グレーの点線は閉構 造時に生じる疎水性相互作用をそれぞれ表す。
図1.16: 推測されるBsLBDHと3PLBDHの基質結合サイトを構成する残基の開構造時の コンフォメーションの予想分布。各グラフの縦軸は分子数、横軸は開構造のコンフォメー ション、破線は最も安定な開構造のコンフォメーション、水色の領域はリガンドを結合し て閉構造をなり得る領域。
を行ったところ、BsLBDHもNAD+およびBMEが存在していないときはTrp192の側鎖
が基質結合サイトに内傾し、同時にTyr214の側鎖配向もNAD+およびBME結合時と異
なっており、BsLBDHはNAD+および基質を結合する際に構造変化が必要であることが
明らかになった。そして3PLBDHのAPRとの結晶構造をBsLBDHのAPRとの結晶構造
と比較したところ、3PLBDHのTrp190は、BsLBDHのTrp192よりも内傾しており、同
時にPhe146も内傾し疎水性相互作用を生じる距離にあった。このことから、3PLBDHの
Kmの高さは、Phe146とTrp190の疎水性相互作用による基質結合サイトの狭さによるも
のと推察された。この時、閉構造時にPhe146とTrp190の近傍にあるPhe212もPhe146
と疎水性相互作用を生じると考えられることから、Phe146とTrp190の基質結合サイトへ
第
2
章
meso-2,3-butanediol dehydrogenase
由来部位特異的変
異
L-2,3-butanediol dehydrogenase
の触媒能向上のた
めの変異の検討
2.1
緒言
第1章において、Kmが高い3PLBDHはBsLBDHのTrp192に対応するTrp190、Phe148 に対応するPhe146は、基質が結合する前の閉構造時は基質結合サイトに内傾している状態 が安定であり、Trp190とPhe146の疎水性相互作用により基質結合サイトが狭まり、NAD+ と基質が追い出されやすいことによるものと推察された。そしてPhe146とTrp190の近 傍にあり、同様に基質結合ループ上にあるPhe212も疎水性相互作用の生じる位置にある ことが明かになった。この3PLBDHのPhe212は、BsLBDHではTyr214に対応するた め、3PLBDHにF212Yの変異を加えることでTrp190とPhe146周辺の疎水性相互作用 が弱まると考えられたため、本章では変異体3PLBDH-F212Yを作成し、その触媒効率を計測した。合わせて3PLBDH-F212Y (APR複合体)結晶の構造解析も行い、F212Y変異 の構造への影響を検証した。
2.2
方法
2.2.1 発現用プラスミドの構築と形質転換株の作成および培養 3PLBDH-F212Y発現用プラスミドは、鋳型DNAに1.2.1項で作成した3PLBDH発現 用プラスミド、プライマーに3P-F212Y-Fおよび-R (表2.1)を用い、表2.2 (アニーリン グ温度:65◦C)に示すサイクルでPCRを行った後、DpnIによって鋳型DNAを消化し作成 した。発現用形質転換株は1.2.1項と同様の方法でE. coli BL21 (DE3)を形質転換により 作成した。 表2.1: 触媒能検討のための変異体作成用プライマーの配列 名前 配列 3P-F212Y-F 5’-CGAGTACGCCAAACGCATCACTCTCG-3 ’ 3P-F212Y-R 5’-TTTGGCGTACTCGGCGAAACCGTAGC-3 ’ 3PLBDH-F212Yの発現株の培養は1.2.1項に従い、同様の手順で行った。 2.2.2 無細胞抽出液の調製と酵素精製E. coli BL21 (DE3) 3PLBDH-F212Y菌体からの無細胞抽出液の調製と酵素の精製は
表2.2: 触媒能検討のための変異体作成用PCRサイクル 初期変性 94◦C 120 sec。 変性 94◦C 15 sec。 アニーリング x◦C 30 sec。 伸長 72◦C 180 sec。 25 cycle 最終伸長 72◦C 300 sec アニーリング温度xは本文中に記載した。 2.2.3 タンパク質濃度の測定 タンパク質濃度の測定は1.2.3項に従い、同様の手順で行った。 2.2.4 酵素活性の測定 3PLBDH-F212Yの酵素活性の測定は1.2.4項に従い、同様の手順で行った。 2.2.5 結晶化 3PLBDH、3PLBDH-F212Y-APR複合体のの結晶化は1.2.5項に従い、同様の手順で 行った。 2.2.6 X 線回折像の撮影と構造解析 3PLBDH 、3PLBDH-F212Y-APR複合体結晶のX線回折像の撮影と構造解析は1.2.6 項に従い、同様の手順で行った。撮影条件を表2.3に示した。 表2.3: X線回折像の撮影条件 Enzyme 3PLBDH-F212Y bound APR
Beamline PF-AR NW-12A
Detector Quantum210r
Wavelength 1.0000
Exposure time (sec) 1.0
Oscillation angle (◦) 0.5
No. of frames 360
2.3
結果および考察
2.3.1 酵素の精製 3PLBDH-F212Y の精製表を表2.4 に、各精製段階におけるSDS-PAGEを図2.1 に示 した。硫安分画、疎水性相互作用クロマトグラフィ、強陰イオン交換クロマトグラフィの 3段階の精製により、29 KDa付近に単一バンドを確認した。アミノ酸一次配列より推定 される3PLBDH-F212Y単量体の分子量が26,671 Da であり、BDに対してそれぞれ29 U/mg の活性を示すことから、このバンドが3PLBDH-F212Y によるものと確認された。 2.3.2 3PLBDH-F212Y の触媒効率 表2.5に3PLBDH、3PLBDH-F212YおよびBsLBDHの動力学的パラメータを示した。3PLBDH-F212YはL-BDに対するkcat、Kmともに3PLBDHから改善され、kcat/Kmは
3PLBDHの8.1倍となり、F212Yの変異によって3PLBDHの触媒効率が改善されたこ とが確認された。しかし、3PLBDH-F212YのL-BDに対するKmがBsLBDHの230倍、 kcat/Kmが1/87倍であるため、BsLBDHの触媒能に及ばないことも確認された。また、 3PLBDH-F212Yはmeso-BDに対する触媒効率も上昇し、基質認識が緩んだことも確認 された。 表 2.5: 3PLBDH、3PLBDH-F212YおよびBsLBDHの動力学的パラメータ kcat (sec−1) Km (mM) kcat/Km (sec−1·mM−1) Ratio of kcat/Km Enzyme Substrate 3PLBDH L-BD 45 ± 1 67 ± 3 0.67 ± 0.01 -meso-BD - > 400 - -3PLBDH-F212Y L-BD 95 ± 7 18 ± 3 5.4 ± 0.5 8.9 meso-BD 63 ± 7 100± 10 0.61 ± 0.02 1 BsLBDH L-BD 37 ± 3 0.080 ± 0.010 470± 40 960 meso-BD 0.99 ± 0.02 2.0± 0.1 0.49 ± 0.02 1 2.3.3 3PLBDH-F212Y の結晶化
3PLBDH-F212Y (APR複合体)の結晶化は、初期のスクリーニングでは沈殿剤にPEG6000
またはPEG4000を使用した条件で針状の結晶や棒状の微結晶が確認された。これらの条
件を最適化することで、3PLBDH-F212Yはタンパク質溶液が10 mg/mL、10 mM Tris-HCl (pH8.0)、1 mg/mL APR、リザーバー液が50 mM MESバッファ(pH6.2)、27.5%PEG4000、
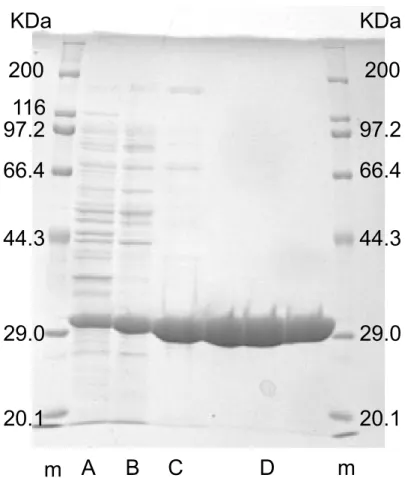
図2.1: 3PLBDH-F212Yの精製段階におけるSDS-PAGE。12%ポリアクリルアミド、6×6 cm ゲル使用。AはCFE、Bは硫安分画後、Cは疎水性相互作用クロマトグラフィ後、D は強陰イオン交換クロマトグラフィ後、mは分子量マーカー。各レーンタンパク質量5 µg 供試。 表2.4: 3PLBDH-F212Yの精製表 Process Activity (U) Protein (mg) Specific Activity (U/mg) Yield (%) Fold 無細胞抽出液 (CFE) 490 73 6.6 100 1.0 硫安分画 200 38 5.2 41 0.8 疎水性相互作用クロマトグラフィ 90 4.2 21 18 3.2 強陰イオン交換クロマトグラフィ 50 1.7 29 10 4.4
図2.2: 3PLBDH-F212Y結晶。タンパク質:10 mg/mL、1 mg/mL APR。リザーバー液:50
mM MES酸バッファ (pH6.2)、27.5%PEG4000、15%グリセロール。
2.3.4 X 線回折像の撮影と構造解析
表2.6に3PLBDH-F212Y (APR複合体)結晶の回折データと精密化の統計値を示した。
3PLBDH-F212Y (APR複合体)結晶構造も3PLBDH (APR複合体)結晶構造同様、サブ
ユニット A-Cについてはすべての残基の座標を決定できたが、サブユニットDについて
は200-204番残基の座標は電子密度が不明瞭なため決定できなかった。各サブユニットを
重ね合わせたところ、3PLBDH (APR複合体)結晶構造同様、サブユニットA-Cは閉構造
であることが確認され、サブユニットDも閉構造であることが推測された。以後、サブユ
ニットAを3PLBDH-F212Y (APR複合体)結晶構造の単量体閉構造として代表させた。
3PLBDH-F212Y (APR複合体)結晶構造はすべてのサブユニットにAPRの電子密度を
確認し、座標を決定した。
2.3.5 3PLBDH および 3PLBDH-F212Y の APR 複合体結晶構造の基質結合
サイトの比較
3PLBDH (APR複合体)結晶構造および3PLBDH-F212Y (APR複合体)結晶構造の基
質結合サイトを比較したところ、3PLBDH-F212Y (APR複合体)結晶構造では、Trp190側
鎖が3PLBDHよりもわずかではあるが、さらに基質結合サイトへ内傾いていた(図2.3-A)。
しかし、3PLBDH (APR複合体)結晶構造のTrp190側鎖とPhe146側鎖、Trp190側鎖と
Tyr212側鎖の最短距離はそれぞれ3.9 ˚A、4.1 ˚Aであったのに対して、3PLBDH-F212Y
表 2.6: 回折データと精密化の統計値
3PLBDH-F212Y bound APR
Data-collection statistics
Beamline PF-AR NW-12A
Wavelength (˚A) 1.0000 Space group P 212121 Cell parameters a (˚A) 76.7 b (˚A) 118.4 c (˚A) 158.1 Resolution range (˚A) 50 - 2.48 (2.56 - 2.48) Unique reflection 51523 Completeness 99.5 (96.9) Average redundancy 6.8 (5.2) Rmerge 4.5 (12.6) ⟨I⟩/σ⟨I⟩ 35.6 (12.6) Refinement statistics Resolution range (˚A) 47.4 - 2.48 Rwork/Rfree 0.2387 / 0.2810 RMSD bonds (˚A) 0.002 Angles (◦) 0.646 Modeled molecular Protein 4 Ligand 4 APR Solvent 56 Average B-factor Protein 27.0 Ligand 27.7 Solvent 22.6 Ramachandran favored (%) 96.9 Ramachandran outliers (%) 0.0
距離はそれぞれ4.1 ˚A、4.5 ˚Aであったため、F212Yの変異により疎水性相互作用が軽減 されたことは確認され、これによりNAD+と基質を結合した閉構造をとりうるTrp190の 配向の範囲が広がり (図2.4)、Kmが下がったものと推測された。 3PLBDH-F212YはL-BDに対する触媒効率が3PLBDHよりも向上したが、BsLBDH の1/112倍と依然小さかった。この原因を考察するため3PLBDH-F212Y (APR複合体) 結晶構造とBsLBDH (APR複合体)結晶構造の基質結合サイトを比較した結果 (図 2.3-B)、3PLBDH-F212YのTrp190のCα-Phe146のCα間が12.4 ˚Aであり、これに相当する BsLBDHでの距離が14.0 ˚Aであった。このことより、3PLBDHおよび3PLBDH-F212Y が閉構造時にTrp190-Phe146間で疎水性相互作用が生じる原因はTrp190とPhe146が含 まれている主鎖間がBsLBDHよりも近いためと推測された。 そこで、Trp190が含まれる3PLBDH (APR複合体)結晶構造の基質結合ループと、 Phe146が含まれているβE-αE間のループの主鎖骨格を重ね合わせて比較した (図2.5、 2.6)。この結果、3PLBDHのTrp190のCα自体はBsLBDHのTrp192と1.2 ˚Aの偏差で あったが、基質結合ループ内で最大3.8 ˚Aの違いがあった。3PLBDHのPhe146も同様に、 Cα自体は0.7 ˚Aの偏差であったが、Pro147では1.4 ˚Aと、βE-αE間のループ最大の偏差 があった。このことより、3PLBDHの触媒効率改善には、主鎖骨格を変えるためのさらに 広範な残基の変異を検討する必要があると推測された。
2.4
総括
(2
章
)
前章の結果から3PLBDHの変異体3PLBDH-F212Yを作成したところ、L-BDに対す るKmが約0.27倍、kcatが約2.1倍となったため、結果としてkcat/Kmが約8倍となり、 F212Yの変異が触媒効率を向上させたことが明らかになった。しかしBsLBDHのL-BD に対するkcat/Kmの約0.012倍であり、以前触媒効率に問題があった。3PLBDH-F212Y のAPR複合体結晶の構造解析を行ったところ、Tyr212による疎水性相互作用の軽減は確 認されたが、Trp190とPhe146の基質結合サイトへの内傾は改善されていなかった。改め て3PLBDHの結晶構造全体をBsLBDHと比較したところ、Trp190とPhe146のCα間は 3PLBDHではBsLBDHより近いため、閉構造に遷移するときに側鎖間に疎水性相互作用 が働きやすくなったと推測された。この原因は、3PLBDHとBsLBDHでTrp190の存在 する基質結合ループと、Phe146の存在するβE-αE間のループの主鎖骨格の違いによるも のと推測され、3PLBDHの触媒効率を向上させるためにはより広範な変異が必要と考え られた。図2.3: Aは3PLBDH (APR複合体)結晶構造(スカイブルー)と3PLBDH-F212Y (APR 複合体)結晶構造 (緑)の基質結合サイトの比較。Bは3PLBDH (APR複合体)結晶構造
図 2.4: 推測される3PLBDHと3PLBDH-F212Yの開構造時の基質結合サイトを構成す る残基のコンフォメーションの予想分布。各グラフの縦軸は分子数、横軸は開構造のコン フォメーション、破線は最も安定な開構造のコンフォメーション、水色の領域はリガンド を結合して閉構造をなり得る領域。3PLBDH-F212Yは、リガンドが結合して閉構造にな り得る領域が3PLBDHよりも広いと考えられるため、同じ基質濃度でも活性が高く、基 質濃度が上昇するに連れてリガンドが結合して閉構造になり得る酵素分子数の上昇も速い と考えられる。
図 2.5: 3PLBDH (APR複合体)結晶構造とBsLBDH (APR複合体)結晶構造の基質結合 ループの主鎖骨格の比較。
図2.6: 3PLBDH (APR複合体)結晶構造とBsLBDH (APR複合体)結晶構造のβE-αE間 ループの主鎖骨格の比較。
第
3
章
L-2,3-butanediol dehydrogenase
と
meso-2,3-butanediol dehydrogenase
からのドメインキ
メラ
L-2,3-butanediol dehydrogenase
の構造解析
3.1
緒言
前章の結果から、3PLBDHの触媒効率の低さは、基質と直接相互作用するTrp190と Phe146におけるCα間の距離がBsLBDHよりも近いため、側鎖間の距離が近くなり、側 鎖同士で疎水性相互作用が生じて基質結合サイトを狭くしていたことによるものと推測さ れた。そしてその原因は3PLBDHとBsLBDH間で、Trp190とPhe146が存在する主鎖 骨格に違いがあるためと推測され、当該残基をBsLBDH様にするためには、より広範な 残基の変異が求められた。そこで本章では、高機能L-BDH創製のための改変対象を、基 質結合サイト周辺の残基がより広域にBsLBDHと一致し、KmがBsLBDHと同程度に低 いドメインキメラ酵素cLBDHに変更した。しかし、cLBDHは基質認識が緩いことから、 本酵素を実用酵素とするための第一段階として、この緩みの原因を解明するための構造解 析を行った。またcLBDHの構造情報は、3PLBDHの触媒効率とTrp190とPhe146の側 鎖の関係についても知見が得られるものと期待された。3.2
方法
3.2.1 発現用プラスミドの構築と形質転換株の作成および発現 cLBDH発現用プラスミドの構築および形質転換株の作成は、鋳型DNAにプラスミド pLALBLCMD[田草川, 2004]を用いてPCRを行い、3PLBDH (1.2.1項)と同様の手順で 作成した。 cLBDHの発現用菌体の培養は1.2.1項に従い、同様の手順で行った。 3.2.2 無細胞抽出液の調製と酵素精製 cLBDHの無細胞抽出液の調製と酵素精製は1.2.2項に従い、同様の手順で行った。 3.2.3 タンパク質濃度の測定 タンパク質濃度の測定は1.2.3項に従い、同様の手順で行った。 3.2.4 酵素活性の測定 cLBDHの酵素活性の測定は1.2.4項に従い、同様の手順で行った。3.2.5 酵素サブユニットの生物学的単位の決定
cLBDHの精製タンパク質溶液に対してゲルろ過クロマトグラフィを行うことにより、酵
素サブユニットの生物学的単位を決定した。カラムはHiLoad 20/60 Duperdex 200 prep
grade (GE Healthcare)を使用し、展開バッファに10 mM Tris-HCl (pH 8.0)を用いた。
カラムへの送液は11.3 cm/hで行った。分子量マーカーにはGel Filtration Makers Kit for Protein Molecular Weights 29,000-700,000 Da (SIGMA-ALDRICH)を用いた。
3.2.6 結晶化 cLBDHの結晶化は1.2.5項に従い、同様の手順で行った。 3.2.7 構造解析 cLBDHの構造解析は1.2.6項に従い、同様の手順で行った。撮影条件を表3.1に示した。 表3.1: X線回折像の撮影条件 Enzyme cLBDH bound NAD+
Beamline PF-AR NE-3A
Detector Quantum270r
Wavelength 1.0000 1.8000
Exposure time (sec) 1.0 0.5
Oscillation angle (◦) 0.5 1.0 No. of frames 360 360 Temperature (K) 95 95
3.3
結果および考察
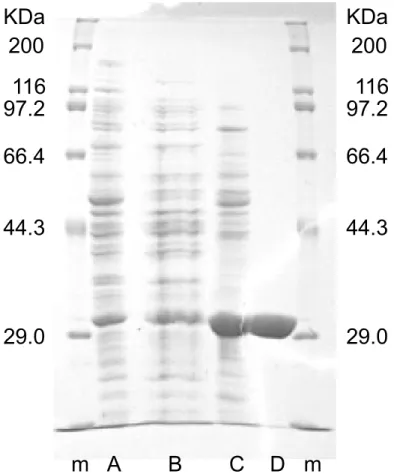
3.3.1 酵素の精製と生物学的単位 cLBDHの精製表を表3.2に、各精製段階におけるSDS-PAGEを図3.1に示した。硫安 分画、疎水性相互作用クロマトグラフィ、強陰イオン交換クロマトグラフィの3段階の精製 により、29 KDa付近に単一バンドを確認した。アミノ酸一次配列より推定されるcLBDH 単量体の分子量が26,696 Daであり、BDに対して32 U/mgの活性を示すことから、この バンドがcLBDHによるものと確認された。またcLBDHの溶液中での分子量はゲルろ過 クロマトグラフィにより、103 KDaと決定した。これより、cLBDHの生物学的単位は四量体であり、由来となるKpmBDH[Ui et al., 1997]、BsLBDH[Takusagawa et al., 2001] と同様であることが確認された。