ペプチドグリカンによる
抗菌性タンパク質誘導の分子機構
M
o
l
e
c
u
l
a
r
mωhanisms
o
f
i
n
d
u
c
t
i
o
n
o
f
a
n
t
i
b
a
c
t
e
r
i
a
l
p
r
o
t
e
i
n
s
y
n
t
h
e
s
i
s
b
y
p
e
p
t
i
d
o
g
l
y
c
a
n
池 谷 政 男
Masao I
k
e
t
a
n
i
1
9
9
9
目次
要旨 第 I章 緒論 第 Ⅱ章 カイコ脂肪体培養系における抗菌性タンパク質の誘導 第二節結言 第2節
材料及び方法
2-1
材料 (1)カイ コ(2)バ
クテ リア13)試
薬2-2
方法 (1)カイコ幼虫か らの脂肪体組織の描出及び培養(2)抗
菌活性の測定 (3)リ ゾチーム活性の測定 (4)タ ンパク質の定量(5)脂
肪体内へのF刺―リジンの取 り込み(6)ポ
リアクリルアミ ド電気泳動 第3節 結果(1)F珂
―リジンの取 り込みによるタンパク合成の確認(2)脂
肪体培養系における抗菌性タンパク質の誘導(3)パ
クテ リア菌体による抗菌性タンパク質の誘導 ぃ)可
溶性ペプチ ドグリカンの抗菌性タンパク質誘導効果 第4節
考察´ 第Ⅲ章 抗菌性タンパク質誘導に必要なペプチ ドグリカンの最小構造 第 1節 結言 第
2節
材料及び方法(1)パ
クテリア(2)直
鎖状ペプチ ドグリカンの調製(3)細
胞壁 由来ペプチ ドグ リカンの調製(4)低
分子ペプチ ドグ リカンの調製(5)ア
ミノ酸分析(6)平
均糖鎖長の測定(7)ペ
プチ ド側鎖架橋度の測定 13 15 15 15 16 18 18 18 18 19 2。 22 28 3。 36 38 38 39 網 網 網 4︲第3節
結果
(1)直
鎖状ペプチ ドグ リカ ンによる抗菌性 タ ンパク質の誘導(2)抗
菌性タ ンパ ク質誘導 に必要なペプチ ドグ リカ ン最小構造 第4節
考察 第Ⅳ章 抗菌性 タ ンパ ク質誘導 に関す るペ プチ ドグ リカ ン認識タンパク質 第1節
緒言 第2節
材料及び方法 2‐
1
材料(1)バ
クテ リア及びペプチ ドグ リカ ン (21 試薬2-2
方法(1)PC―Afrl ty∞lumnの調製
(2)B.“
T倣
舛 "“,NCIMB 12520(印 ‐ ヵ ∫)を用 い たF呵‐PCの
調 製 (3)ド251]_BoltOn―Hunter試 薬によるPCの
標識 (4)ASD‐PGの
調製 脩 )ピ 数 ]一 ASD‐ PGの 調 製 (6)F2SI]_ASD‐PCに
よ る フ ォ トア フ ィ ニ テ ィ ラ ベ ル(7)BIACOREシ
ス テ ム 用 セ ン サ ー チ ッ プ ヘ のPGの
固 定(8)脂
肪体細胞膜の調製 (9)カイコ幼虫体液タンパクの調製 第3節
結果
(1)RI標
識PGに
よるPG結
合性タンパク質の検索 (2)『251]_ASD‐PCを
用 いたPG結
合 タ ンパ クの検 索(3)BIACORE2000に
よ るPG結
合 タ ンパ クの検 索 及び PG―ア フ ィニテ ィカ ラム によ る精製(4)PG結
合特 異性 の検 討 第4節
考察 総括 謝辞 参考文献 狼 心 5 ︲ 舅 Й Й 55 55 55 55 夕 57 ” 58 59 6。 ∝ 69 81
要旨 昆虫な どの無脊椎動物は免疫 グロプリンを持たず、生体内に侵入した異物に対 して哺乳 類 に見 られるような特異的な獲得性免疫反応を示さない。この昆虫の生体防御反応の一つ として、バクテリア感染によるセクロピン、アタシン及びリゾチームなど一群の抗菌性タ ンパク質の誘導が知 られている。カイコ幼虫にバクテ リアを注入すると、数時間後には体 液 中に抗菌性タンパク質の誘導が観察される。 この誘導はバクテ リアの細胞壁ペプチ ドグ リカ ン
eOや
グラム陰性菌外膜成分であるリポ多糖(LPS)を単独で注入す ることで起 こる ことか ら、これ ら細胞壁成分が抗菌性タンパク質の直接の誘導因子であることが明らかと なっている。抗菌性タンパク質誘導にはPG構
造の特異性力蝙悪められることか ら、この誘 導には一定のPG構
造を認識するシステムが存在する可冑酵陛が考えられる。この論文はPG
によるカイコ抗菌性タンパク質誘導の分子機構について検討 した結果をまとめたものであ る。 第 I章 では、昆虫の一般的な生体防御機構 と、 これまでに明 らかとなっている抗菌性タ ンパク質誘導機構に関する知見について述べた。 第 Ⅱ章では、抗菌性タンパク質の産生器官であるカイコ幼虫脂肪体を培養し、加 サPOで
の抗菌性タンパク質誘導系の確立を行った。あらかじめPGを
注射 し免疫化 した脂肪体をace培
地で培養すると培地中に抗菌活性が検出されたが、無処理の幼虫脂肪体では顕著 な抗菌活性は認められなかった。しかし、無処理の脂肪体にPGを
添加 して培養すると高 い抗菌活性が誘導された。電気泳動によ り同定を行ったところ、誘導された抗曲性タンパ ク質はリゾチームとセクロピンであった。また、シクロヘキシミ ドあるいはアクチノマイ シンDの
添加によ り完全に阻害されたことか ら、この誘導はPGの
刺激によるtr9 ηο乃 合 成であること力靖諷め られた。 この 加virpで の誘導系を用い、PG構
造の違いによる誘導効果を検討したところ、これ まで 加 ッル9で
得 られた結果 と一致 し】 “ ,′伽 “ 9gprcPrrT開 及 びh力9x励ヵ ω力由来の細胞壁PGに
よって高い抗菌活性が誘導されたが、M破
∝αβ ヵど筋ば 由来の細胞壁PGで
は非常 に低い活性 しか誘導されなかった。また、血球を含まない脂肪体堵養系にバクテリア菌体 を直接加えた場合にも顕著な活性の誘導が観察された。これ らの結果は、抗菌性タンパク 質誘導に血球によるバクテリアの貪食は必ず しも必要でないことを示唆 している。PGは
哺乳動物においても様々な免疫応答を引き起 こす ことが知 られているが、それに必要な構造については未だ明 らかになっていない。そ こで第Ⅲ章では誘導に必要な
PCの
最小構造の特定を行った。誘導効果に大きな差のある 】 "?♂t煎114や 二 ω力とM力
r筋1デ の細胞壁PG構
造の違いはペプチ ド架橋部分にあることか ら、まず架橋の有無と誘導効果 について検討 した。そのためにPGの
架橋形成阻害剤であるペエシリンGを
グラム陽性 菌の培地に加えることにより直鎖状PGを
調製 した。a"q♂
=T面
及びM力
協β よ り調 製 した直鎖状PGは
いずれもa
μ9gryr9rilrp細胞壁PGと
同様の強い誘導能を有 していた こ とか ら、抗菌性タンパク質の誘導にはPGの
ペプチ ド側鎖間の架橋は必要ではなく、M
rtrros細胞壁PGに
み られるようなLysを含むかさばるペプチ ド架橋は逆にPGの
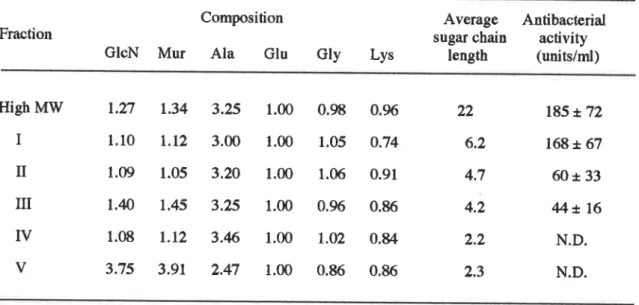
認識を 妨げるものと考えられる。次にこの直鎖状PGを
リゾチームによ り限定分解後、グルろ赴 によ り分画 した分子量の異なるPG断
片を用いて誘導に必要なPGの
最小構造 を検討 した。 その結果、ペプチ ド側鎖を持ったGhNAttMurNAcか
ら成るPGユ
ニットが2つ結合 した ものが誘導に必要な最小構造であると決定 した。 第Ⅳ章ではカイコのPG認
識 システムの検索を行った。上述のように抗菌性タンパク質 誘導効果はPG構
造に依存 していることか ら、カイコにはある特定構造のPGを
認識する システムが存在する可台9性が考えられた。また、加 vrrwの 実験結果か ら脂肪体がPGの
認 識 に関 与してお り、 この認識に直接関わるPG結
合タンパク質力Ⅶ旨肪体細胞に存在する可 能性が示唆された。また、それ以外にもPG認
識 タンパクが体液中に存在する可能性 も考 えられるためPGと
結合するタンパク質の検索を脂肪体 と体液について行った。検索方法 として、1密Iに
よ り標識 した光親和性の架橋剤(SASD)とPGを
結合 させ、 これ を用いたフ ォ トアフィニティ標識によ り検索を行った。その結果、体液中にPCと
特異的に結合する タンパク質の存在が示され、 このタンパク質を部分精製 した。しかし、脂肪体の可溶化細 胞膜 については、 この方法ではPG結
合タンパク質を検出することができなかった。そ こ で、別のアプローチとして生体分子間の相互作用を検出するBIACOItEシ
ステムを用いて 検出を行ったところ、可溶化細胞膜に,Gと
特異的な結合を示すタンパク質の存在が認め られた。 以上のように本論文では、カイコにおける抗菌性タンパク質の誘導は一定構造のPGが
認識されることで起 こることを明 らかにし、さらに誘導因子 として必要なPGの
最小構造 を哺乳動物も含め、初めて明 らかにした。また、このPGを
認識すると思われるPG結
合 タンパク質が抗曲隆タンパク質の産生器官である脂肪体細胞膜に存在することを示 した。Sunl!れ
ary
lnsects resPond tO bac俺 al infection with a se es of cemular and humoral defensive reactions,including synhesis of antibacte al proteins,such as lysozyme,cecropins and attacins.An itteCtOn Of bacteial cell wЛ l pep償doglycan(PG)or hPOpOlysaccharide
(LPS),induces an償bactedal act ity in the hemolymph.The illduction by PG is a specinc resPonse following r∝og hon of PG ofso】me deflnite stRICmre in the hemolymph.The
speciflc induction by P(〕 suggests an e stence ofPC recognition systems in insecto This
study aims to explore he mdecular mecha slxls of induction of antibacte Л pЮtein
synthetts by pepidogycaninthe silkwom,β 閉 勒 脇ば
'・
When fat bOdy fЮ
m immllnized laryae was cuimred in Grace's medium,
andbacte al acuvity was detected in the cldture medium without addition of elicitoro h the cuimre offat body fЮIn naive laⅣ ae,antibacttial activity lvas not detected duing the
irst 7h of incubation,butthe addition of PG to the mediunl,induced the activity du ng 拗 of incubation,The acidic PAGE of the cllltured medillm showed that cecЮ Pin and lysozyme were synthesized and secreted into the medium. '「 he syndleses of the
antibactenal proteins were inhibited by acinomycin D or cyciohexinude,indicaLng de
歿〃ο synthesis of the a dbactenaI PЮteins in cllltured fat bodyo PG fЮ
m
β.じο″or β.陶等″び賜 had high ability to indllce an戯b“俺五al pЮteins,butPG fЮ
m
ν町ガ?"d was very p∞r elicitor.The res』 ts suggested an e stence of PG recognidon system in fatbody celis.
PGsfЮ
m
β.′た膨η″ 服ねand ν:′′舵′∫were different each oher in their pepddelinkage structures,which might be a cause for the great difference in the inducing abihty
ofthese PGs.To PЮve he possibility,linear uncЮ ss‐linked PGs were isolated fЮ m the
bacte al culture rledia incubated with penicillin G,and tested for the elicitor activity.The
linear PGs fЮ
m肱
肋′9Иs and β.Jた膨刀ゎr胸ねWere simmary effecuve as cticitor.Theresults suggest thatthe cЮ ss―linkage between attacent pepdde Sde ch ns is not esserldal
for he elicitor activity,but a b』ky and charged penぬpeptide bidge in肱 ルrg"d PG
required for induc也 on of antibactedal pЮ tein synhesis,linear PG fra♂ments with
different md∝ular size were prepared and tested for the elicitor activity.The minimum
smcmre Of PG was deterauned to be two repeaing ttacetylgtucosamine―
洋 acetylmwamic acid u ts wih pepide side chains.As the results suggested Presence of a PC recog don molecule in the fat body cells,
detecion of PG Ыnding pЮtein was attempted using[1251]_ASD― PG,a photoafflnity
CЮSS―linking PG del aive.PG binding PЮtein was detected in the hemolymph after
suЛized the labeled bands On a gel of SDS― PAGE by autoradiography,In solubilized
membmne pЮ
teins fЮnifht body ceWs,however,no specinc binding Was detected withthis lnethod・ For more sensitive and speciflc detection of PG― binding pЮtein in
solubilized fat body mernbrane,BIACORE 2000 was usedo With this system,a binding
of Protein to irrmobilized PG on a sensor tiP was detected.The binding was sPcCiiC for PG,since the protein showed Poor binding for chitin or LPS.
第 I章 序 論
(1)昆
虫の生体 防御機構 自然界 に存在す る動物種 は現在 までに種 として記載 されているものだけで もloo万種 にも 及ぶが、そ の中で哺乳類 を含 めた脊椎動物が約38000種で あるのに対 し、無脊椎動物 に属 す る節足動物 は 930,000種 と、全動物種 の90%以
上 を占めている。 中で も昆虫は 750,OgJ 種が存在 して いる う。 この昆虫の緊栄の背景 には個体数の多さと環境への適応能力の高さ な どが あ るが、恒常性 の維持や微生物 の感染に対す る高度な生体防御 システムの存在 も、 そ の理 由の一つ と考 え られ る。一般 に生体防御反応 は異物の認識か ら始 ま り。それ に対す る無毒化 、排除を行 う一連 の反応 を示す。バ クテ リア感染な ど異物 に対す る哺乳類など脊 椎動物 の防御応答 は、非獲得性 (自然)免
疫反応 と、特異的な抗体 を介 した獲得性免疫反 応 に大 き く分 け られ る の。 ・方、昆虫な ど無脊椎動物には免疫グロブ リンに相当するもの が無 く、異物な ど非 自己に対す る免疫記憶が存在 しない。つま り、昆虫の生体防御反応は 非獲得性 免疫反応 によ り成 り立 っている。 しか し、現在 までの昆虫の生体防御機 に関する 研究結果か ら脊椎動物 の免療機構 を単 に簡略化 したモデル とは言 い難 く、系統発生的に非 獲得性免疫 を発達 させてきた と考 え られ る。 完全変 態型 の昆虫 は、幼虫や蛹 の脱皮時期 にカ ピやバ クテ リアを含 めた微生物 に感染す る頻度が成虫時 に比べ高 くなると考 え られ る。それ ら微生物の侵入 に対す る最初の防御シ ステム は、 ワックス、キチ ン、タ ンパ ク質 によ り構成された非細胞構造のクテクラ層 と呼 ばれ る物理的なバ リアー を表皮 に持つ ことで成 り立 っている。また、経 口由来の感染に対 して は消化管内が高 いアルカ リ性 を示す ことでバ クテ リアの繁殖 を抑制す る。さ らに囲食悟
僣
再
峰
彗
:稔
檀
脅
貞
】
哲
女
:絆
操
!:票
吾
景
ξ
「峰
骨
募
igョ
ほ
曇
:替
ζ
警
存
格
脅
:は
各器官が体液で満たされた同じ空間に存在することか ら、クチクラ層などの物理的なバリ アーが何 らかの形で破 られ、体腔内に微生物が侵入することは昆虫にとって致命的な出来 事である。その体14内はパクテ リアにとって栄養成分やpHな
ど繁殖するには十分な環境 ではあるが、実際には体腔内に侵入 したバクテリアに対 して、体液中に存在する様々な生 体防御システムが働きれバクテ リアの増殖を防いでいる。(2)細 胞性防御反応
昆 虫体波 内での生体防御反応 は大 き く二つ に分 け られ、血球 による細胞性防御反応 と血 漿成分 によ る体液性防御反応 によ り対抗す る。血球は体液内に侵入 したバクテ リア、カ ビ.
線虫類 、産 みつ け られた寄生昆虫の卯な どに対 し、それ ら外来性の異物 を非 自己と認識 し、
貪食作用(phagOCytosis)、 ノジュール形成(■oduに Formtion)、 包 囲化作用(cncapSulation)な ど
によ り排除す る つ。 貪 食作用 は顆粒細胞 とプラズマ細胞 によってバ クテ リアな どの微小な異物 をエ ン ドサイ トー シス によ り血球 内に取 り込み、殺菌、消化する防御反応である。また、 このパ クテ リ アな どを細胞内に取 り込んだ顆粒細胞 が集合体 をつ くった り、バクテ リアの集塊 を顆粒細 胞が取 り囲む ことによって分散 を防 ぐ反応が ノジュール形成 として知 られている。食作用 は対象 となる異物が血球 よ りも小 さな異物に対する反応であるのに対 し、包囲化 と呼ばれ る防御反応 は血球 よ りも大型 の異物 に対 し起 こる反応で、まず顆粒細胞 によ り異物 を取 り 囲んだ後 、 プ ラズマ細胞が反応 し、 この二種類 の血球層を異物の周 りに形成する ことで生 体 内で の異物隔離 を行 う反応で ある。
c)体
液性防御反応 バ クテ リアが感染 した場合 、 まず細胞性 防御反応である食作用 とノジュール形成 によ リバ クテ リアの排除が行われ、 これ に伴 い体液性防御反応が働 く。体液性防御反応 として は、1)抗菌性 タ ンパ ク質 の誘導 2)フェ ノールオキ シダーゼ系の活性化 3)レクチ ン 4)補体 様 囚子5)抗
ウイルス性 囚子 な どが ある 働。中で も、バ クテ リアに対す る主要な防御 シス テム の一 つ として、バ クテ リア感染 によって誘導 される数種の抗菌性タンパク群の存在が 挙 げ られ る。 1.リ ゾテーム 昆虫か ら単離 された最初の抗菌性タ ンパクは、Cα′′ιr,a"ιttИど′′α 由来の リゾチームで ある 6)。 鱗翅 日の 拘 α幼 "ο tt C9Cr9P'α 由来の リゾチームは120ア
ミノ酸残基、分子 景 13。8kDaで
脊椎動物である卯 白リゾテームに非常に似かよっている 7)。 ムラミシダーゼで あるリゾチームは数種のグラム陽性菌に対 して作用 し、甜1胞壁を加水分解する溶菌酵素と して働 く。誘導性の抗菌性タンパク質 とは異な り、 リゾチームは幼虫や蛹の体液中に常に 存在 し、さらにバクテ リアの感染などの刺激によって誘導され、体液中の濃度が増加する8、 9)。 バ クテ リア感染 のない健常な昆虫の体液 に含 まれ る リゾチームはバ クテ リアの感染 に対す る昆虫の初期防衛手段の一つで あると考 え られ、その後誘導 され るセ クロピン、ア タシンと相乗的に働 くことで効率 よ くバ クテ リア感染 に対抗す るもの と考 え られる Ю)。 2.セクロピン
` セ クロ ピンは
Bomanら
によ り 鳳 Gιttη,α よ り最初 に単離 された分子量約4,∝Ю の焔基 性 タ ンパ ク質で現在 まで に同族体A,B及
びマイナー成分のDタ
イ プの存在が確認 されて い る 11ヽ 盟)。 セ ク ロ ピンタイ プの抗菌性 タ ンパ クはそ の他 セ ンチニ クバエ の 肋r甲
脇gα Pιrじgr,ηαお).カイ コ 肋 脇♭Tノ “ θ′,Mヽ 15)、 タバ コスズ メガM朗
れ cα ∫ιズrn lめ な ど多 くの昆虫 か ら単離 されている。 また、昆虫のみな らず哺乳類か らの報告 もあ り、ブタ小腸か らもセ クロ ピンタイ プの抗 由性 タンパ ク質が見つか っている")。cDNAあ
るいはゲ ノム配列か ら、セクロピンは22ア
ミノ酸残基 のシグナルペプチ ドを 含 んだ 62-64残基で構成 されたプ レプロセ クロピンの形で合成 され、産 生細胞か ら分泌時 にシグナルペプチ ドが離れ る。 この後 、体液 中に存在す るジペプチダーゼ によ り、プロリ ンを含むN末
側 のジペ プチ ドが外れ matureな セクロピンとなる。その結果、セ クロピン は 35∼37ア
ミノ酸残基か ら構成 されてお り、N末
端部分 は強塩基性 を示 し、C末
端 の半 分 は疎水性 を呈 している 鰐)。 三次構造 としてN末
側 とC末
側 の2つ
の α―ヘ リックスがAla‐Gly_PЮ を介 して結合 した構造 をとる p)。 大腸菌 にセ クロ ピンを作用 させ る と
ATP量
の減 少 と共 に、細胞 内 K・の漏 出が認 め られ る 2の。 これ はセ クロピンがバ クテ リア内膜 にイオ ンチ ャンネル を形成す る ことで膜電位 を消失 させ、物質輸送、
ATP合
成 を阻害す るためで あ り、C木
端 の疎水性 の α_ヘ リックスは内膜へ侵入す るため に必要である と考 え られ る 21)。 また、 リポ ソーム を用 いてセ ク ロピンの膜へ の作用 を検 討 した結果、負 に 帯 電 した脂質で構成 された膜 の場合 に効率 よ く膜電位が消失す る ことよ り22)、N末
側 の 塩基性 が バ クテ リア内膜へ の吸着 に働 くもの と考 え られ、二つ のヘ リックス を結ぶ Ala― Gly‐Proの
嫌 ヘ リックス ア ミノ酸 を含 む トリペ プチ ドの ヒンジ構造 によ り柔 軟 に曲が る こ とで、それぞれ のヘ リックス を効率よ く働かせている21)。 セ ク ロピンの抗菌効果 は孔平板法 によ り測定 され、その抗菌スペク トル は多 くのグラム 陽性 菌、 グラム陰性菌 に対 して殺 菌効果 を示す 勾)。 このセ クロ ピンタイ プの抗菌性 タ ン パ ク質 は リゾチーム と異な り通常の体液 中には存在せず、バ クテ リアの感染 によって脂肪 体 お よび血球 で合成 され、体液 中に分泌 され る。 しか し、セ クロピンタイ プの抗苗性タ ンパ ク質で ある S.Pι′θgrttα 由来 のサル コ トキ シン
IAは
バ クテ リア感染のみな らず特定の 発 育ステー ジで発現 して いる ことが ノーザ ン分析 の結果か ら明 らか とな った 雰)。 この結 果 はセ ク ロピンが ある種 の昆虫では単 にバ クテ リアに対す る生体防御だけではな く変態時 に何 らかの役割 を果 た している可能性が考 え られ る。3.ア
タシ ン セ ク ロピンと同様 ユ cじm叩
,α よ り単離 されたアタシンは当初、Aか
らFの 6タ
イ プが 存在す る 勾)。 しか し、得 られたcDNAク
ロー ンよ りこれ らのアタシンは酸性 アタシン (aOdattacin)と塩基性 アタシン(basiC attacin)の 2つ の遺伝子産物である ことが明 らか となった%)。
ア ミノ酸配列の解析か ら酸性 アタシンは188残基 、塩基性 アタシンは 187残基か ら成 り、 それぞ れ の
C末
端側 にある、Ser‐Lys_Tyr_Phe,のテ トラペ ブチ ドの部分分解 と、酸性 アタシンの
N末
端 Glu lの 環状化 (PyЮ81utama俺 )の有無か ら6タ
イ プのアタシ ンが生 じた も の と考 え られ る2の 。 アタシンの抗菌スペク トル は狭 く、h力ιr,cttα θθ′,と2種
類 の腸 内細菌 にのみ効果が認 め られ て い る 拓)。 また、バ クテ リアの定常期 には効果が無 く、増殖期 のパ クテ リア外膜 に働 くことで、隔壁合成 を阻害す る 25、 27)。 実 際、 アタシ ンと同族 の抗菌性 タ ンパ クであ る S,Pιrg♂珈α 由来 のサル コ トキシンⅡを作用 させたバ クテ リアを電子顕微鏡 によって形 態観察す る と、菌体 の伸長化 と共 に細胞膜 のス フェロプラス ト様変化が生 じ、菌体 の破損 が認め られ る2の。4.ァ
フェ ンシ ン デ フェ ンシンは6個
のCysを
含 む29∼35ア
ミノ酸残基か らな り、多 くのグラム陽性菌 お よび陰性菌 に対 して抗菌作用 を示す抗菌性 ペプチ ドで、貪食作用 を有す る多形核 白血球 で ある ヒ トの好 中球 よ り最初 に見つか った29J。 ゥサギでは肺胞マクロファー ジに存在 し、 呼 吸経 由のバ クテ リア感染 に対抗す る 3ω 。 同 じデ フェンシンフ ァミリーである β‐デ フェ ンシ ンは ラッ ト31)、 マウス Эり、ウシ 33Jな どに存在す ることが明 らか となってお り、 この タ ンパ クは哺乳類一般 の抗バ クテ リア物質で あると考 え られた。 しか し、 これ と相 同性 を 持 つ抗菌性 タ ンパ ク質がP力伊′ “ ,α ″rrαttοソβθ 30、 ぉ ょびS.ρどκ8′ '力 α的な ど数種 の昆虫よ り 見 つか ってお り、昆虫デ フェンシン、 あるいはザーペシンとして知 られている。昆虫デ フ ェ ンシ ンはグラム陽性菌の細胞膜 に働 き.膜
透過性 に変化 を与 えATP合
成 を阻害す るなどの効果 を示すが、グラム陰性菌や真核細胞には影響が少ない3o。
5,フ
ェノールオキシダーゼ系 抗菌性タンパク質誘導 と同じく、昆虫の体液性防御に関わると考えられる防御機構にフ ェノールオキシダーゼ系がある。パクテ リアやカビの感染によって活性化されたフェノー ルオキシダーゼによ り、体液中のフェノール性物質のチロシンが酸素存在下で酸化されメ ラニ ンを形成す る。その結果、 このメラニンは体表の傷を塞 ぐと共に、侵入 したバクテリ アやカ ビなど異物の周囲を包囲化 し、増殖や分散を防ぐ37,383。 ヵィコ ∂。加οr,におけるフ ェノールオキシダーゼ系カスケー ドの誘尭は、バクテリア細胞壁 PG、 あるいはカ ビの細 胞壁成分であるβ■,3‐グルカ ンを特定のタンパク質が認識することか ら始まり、セ リンプ ロテアーゼ前駆体が活性化され、次に、このセリンプロテアーゼによ リフェノールオキシ ダーゼ前駆体(PrOPO)が活性化 される394)。 6。 その他、生体防御 に関連する体液性困子 ヘモ リン(hemol )は ユ じ¢伽T'α と 肱 ざιttα より単離された免疫 グロプ リンスーパーフ ァミリー属す る分子量50kDaの
タンパク質で、通常の体液中に低いレベルで存在 し、バ クテ リアの感染によ りOcr9P,,で
は18倍
、財加JIt側 では 30-45倍 もの濃度の増加が認め られる 41)。 このヘモ リンはパ クテ リア感染に対する防御タンパクの一つ として考えられ ているが、他の誘導性タンパク群のような抗菌性を示さない η)。 Sunら 4)は 加 ソ ,どrrpにお いて、ヘモ リンがバクテ リアヘ特異的に結合することを明 らかにし、さらにLadendor∬ ら 4)は 、ヘモ リンが血球 に結合することで血球の凝集が妨げられることを示 した。また、Sirlei ら4)はヘモ リンが LPS、 特 にその結成脂質であるリピドAに
結合することを示 したこと 等か ら、 このタンパク質がバ クテ リアの認識 に関与 し、オプソニ ンとしての働きを持つ可 能性が考 えられる。 ぃ)抗
菌性タンパク質の誘導因子及びその認識機構 抗菌性タンパク質の誘導機構の解明を目的に幾つかの研究グループによ り抗菌性タン パク質の誘導因子の特定が行われ、幼虫に様々な物質を注射することで誘導の特異性が検 討された。Powningら `)は 鱗翅 目昆虫のG.μθ′′ο附♂Jね とBo μο河の幼虫にグラム陽性苗で ある Mじ′οじじ″dゥ∫οJひ脇施 "∫ 生菌を注射 し、体液中の リゾチーム活性が無処理の幼虫体液に比べ約
10倍
程度 高 くな る ことを明 らかに した。 また、セ クロピンタイプの抗由性タン パ ク質 も リゾテーム と同様 に幼虫の体腔内にパ クテ リア菌体 を注射することで誘導される Ъ'守)。 この鱗翅 目昆虫 に認め られ る抗菌性タ ンパク質誘導は、パクテ リア感染のシグナル として グラム陰性菌外膜 の構成成分である リポ多糖(LPS)および グラム陰性 、陽性菌 に共 通 して含 まれ る細胞壁ペプチ ドグ リカ ン(PC)が誘導因子 として働 く 狙ヽり、勤)。PCは
β_1,4 結合 した N―アセチルム ラミン酸 と N‐アセチル グル コサ ミンの繰 り返 しか ら成る糖鎖 とム ラミン酸 に結合 した短 いペプチ ド鎖か ら構成 されている。 このペプチ ド鎖はリジン、ある いはジア ミノピメ リン酸 を含 み、 これ らのア ミノ酸の持つ ε―アミノ基を介 して他のペプ チ ド鎖 のC末
端 と架橋 を形成 して いる。 グラム陰性菌のPGは
ペプチ ド側鎖 同士が直接結 合 して いるが、グ ラム陽性菌の場合、架橋部分が種 によ り多様な構造 を持つ。カィコ β,μο河 で は、注射す るバ クテ リアの種類 とそれ によって誘導 される抗菌活性の間に相関性があり、 グ ラム陰性菌では種 に関係な く高 い誘導効果を示すが、肱 ′μ″′ざを含めた特定のグラム陽 性 菌では誘導効果が低 い。Moishimaら
51)は このパ クテ リアの種類 の違 いによる誘導効果 の違 いがPGの
構造 に依存す る ことを明 らかに した。 昆 虫の抗菌性タンパ ク質 の誘導因子であるLPSは
、古 くか らヒ トのバ クテ リア感染 に よる敗血症 の主要因子 として特定 されてお リマクロファー ジに作用することで腫瘍壊死因 子cNF― α)、 L-1、 L‐6な
どサィ トヵィ ンの過剰発現 を誘発することが知 られている 立ヽヨヽ 舅)。 しか し、 グラム陰性菌 の感染 によって起 こる種々の生体反応がLPSを
持 たな い グ ラ ム陽性菌 において も同様 に起 こり、その誘発因子がバクテ リア細胞壁PGで
ある ことが明 らか となった `)。 また、 Nぃアセ チルム ラミン酸 と L‐アラニ ン、Dイ
ツグル タ ミン酸の二 つのア ミノ酸か ら成 る muramyl dipepnde(MDDは 、抗体産生を誘発するアジュバ ン トペプ チ ドとして知 られているが 質)、MDPは
PCの
部分構造であ り、PG自
体 が ア ジュバ ン ト 活性 を持 ち、免疫系細胞 に働 き様々な生体反応 を引き起 こす。 これ らの結果は昆虫か ら哺 乳類 に至 るまで、PGや
LPSな
ど締胞壁成分がバクテ リア感染の共通のシグナル として働 いている ことを示唆す る。 哺乳類 にお けるLPSや
PGの
認識 システム については、直接 ヒ トの病態 と関連する研 究対象 として注 目され、多 くの研究 グループによ り解明されつつある。LPSに
関 して は、 そ の 認 識 シ ス テ ム と してcD14と
LPS―Ыnding p『Otcin(LBP)が知 られ て い る 5め 。 血 漿 中 に 存在するLBPは
生体内に侵入 したグラム陰性菌のLPSと
結合 し、LBP_LPS複
合体を形成 する。 この複合体はマクロファージ細胞表面上に存在するCD14に
よって認識されることで、細胞 内へのシグナル伝達が行われ る581。
LPSと
は対照的にPG認
識 システム につ いての知見は少ない。DziarsMら S'ヽ ωヽ61)の 報告 によれ ば リンパ球細胞 の表面上にPGに
特異的な結合部位 を検 出 し、 しか もLPSも
この部 位 に結合す る ことを示 し、LPSと
Pcは
同 じ認識 システムを共有す る と考 え られていた。 そ の後、 このPG結
合 タ ンパ クは細胞培養時 に用 い られた血清アル ブ ミン(BSA)で
ある ことが明 らか となったが、実際にBSAや
、 ヒ トアル プ ミンが免疫 システム誘発 のためのPG認
識 に関わっているのかは不明で ある。 また、最近 にな り同 じDJar飩
らのグルー プ が、LPS認
識 システムであるCD14に PGが
特異的 に結合す る ことを報告 した62、 ●、a)。 哺乳動物でのPG認
識 システムの解明が明 らかでないの と同様、昆虫において もPG認
識 システム に関す る報告 は少ない。唯一昆虫のPCの
認識 システム として知 られている も ので は前述 した Ashidaら によって報告 されているフェノールオキンダーゼ系におけるPG
認識 タンパ クが ある6め 。バ クテ リア感染 を認識す るとい う共通性か らPG認
識 系 を共有す る可能性 も考 え られ るが、 フェノールオキシダーゼ系は高分子PGに
よ り活性化 され、 リ ゾテーム処理 した低分子PGで
は活性化 されないが41)、 抗菌性 タ ンパ ク質は低分子PGに
よ り誘導 され る。5り 。 これ らの結果か らフェノール オキ ンダーゼ系 と抗菌性タンパク質誘 導系 はPGを
誘 因物質 とす るが、それぞれ異なった認識機構 によ り活性化 されるものと考 え られ る。 フェノールオキシダーゼ系でのPG認
識 タ ンパ クの解 明が進む一方で、抗菌性 タ ンパ ク質誘導系 におけるPG認
識機構 につ いては これ まで報告例がない。 前述 したよ うに、昆虫では、T細
胞 、B細
胞 な どによる特定の抗原 に対す る獲得性免疫 反応 は確認 されてお らず、記憶 を伴わない非獲得性の防御反応のみ働 くと考 え られている。 哺乳類 の免疫機構 の中で、補体系の第二経路活性化やマクロファー ジの活性化は非獲得性 免疫反応 の一 つであ り、バ クテ リア由来 の成分によ り直接活性化 される。 これ ら非獲得性 免疫 はそ の後に続 く獲得性免疫が働 くまでの ラグタイムを埋めるための初期防御反応であ る と考 え られ る。昆虫 にとって ラグタイムは致命的な出来事であ り、バ クテ リア感染に対 し、即効 的で有効的な非 自己認識 システム と急性期反応 を必要 とす る。それ故、フェノー ル オキ シダーゼ系やヘモ リンな どの非 自己認識 システムや、一度の感染によって数種類の 抗菌性 タ ンパ ク質 を誘導 させ るシステム を進化 の上で発達 させてきた と考 え られる。その 一方で、セ クロピンやデ フェンシンな どの抗菌性タンパク質が哺乳類にも存在するなど. 昆虫特有 と考 え られていた抗菌性 タンパ ク質が生物種 を越え、共通 して存在することか ら、 昆虫で主要な働 きを担 う抗菌性 タ ンパ クは系統発生的に保存 された防御 システムの一つでr はないか と考えられる。故に昆虫の抗菌性タンパク賀誘導機構の解明は昆虫の枠 に留まら ず、哺乳類 における初期防御反応の解明や、哺乳類の進化の過程で洵汰され休眠 している 新たな免疫システムの発見につながる可能性が考え られる。 本研究は昆虫において、パクテリア感染によ り生 じる抗菌性タンパク質誘導の経路解明 を目的とし、特に誘導因子 としてバ クテ リア細胞壁
PGの
必要な最小構造の決定、及び昆 虫のみな らず哺乳類 において も明 らかとな っていないPG認
識 システムについて検討 した。第Ⅱ章
脂肪体培養系を用いた加ッF施 での抗菌性タンパク質誘導 第 1節 緒言 抗体を介 した抗原特異的な生体防御システム持つ脊椎動物 とは対照的に、免疫記憶を伴 わない昆虫の生体防御システムを構成する主要な因子の一つである抗菌性タンパク質は、 バクテ リア感染に対するエフェクター因子 として、セクロピンやアタシン、デフェンシン な ど様々な種類の抗曲性タンパク質が多 くの昆虫か ら見つかっている。 これまで抗菌性タ ンパク質は、将来的な有効利用 を目的としてタンパク質あるいは遺伝子 レベルから多くの 情報が得 られた。 しかし、バクテリア感染から抗菌性タンパク質の誘導に至るまでの経路 に関しては今だ完全に解明されていない。 抗菌性タンパク質は、バクテ リア菌体を直接昆虫の体腔内に注射することで体液中に誘 導される 守ヽ48Do Dunnら 到は麟翅目昆虫である
M肋
肋留 陀陶 を用いリゾチームや他の抗 菌性タンパク質の誘導が、M′
"rfLT∫ 由来の細胞壁可溶性ペプチ ドグリカン(SPG)の刺激に よつて起 こることを示 した。また、彼 らは、昆虫体腔内に侵入 したバクテリアが体液中の リゾチームによ り分解を受け、その結果生 じたPG断
片が抗菌性タンパク質合成のための 誘導因子であると考えた。Morish:Ⅲ ら う。はある特定構造のバクテリア細胞壁PCが
抗菌 性タンパク質誘導に必要であることをβο用夢 PPTOri幼虫を用いて明 らかにした。β.融ガ に おいて、ペプチ ド側鎖同士が直接架橋 した構造を持つ 二ω “ やB.“駕tJr9‐iw′Иの細胞壁SPG
を注射 した場合、抗菌性タンパク質の高い誘導効果が得 られたが、ベプチ ド側鎖間にリジ ンを含むベンタペプチ ド架橋を有する 肱 肋例∫由来のSPGに
よる誘導効果は低いもので あった。 この結果は、バクテ リア細胞壁PG構
造 を識別するシステムが宿主であるカイコ に存在することを示唆している。 抗菌性タンパク質の合成は、主に脂肪体 と呼ばれる哺乳類の肝臓に相当する代謝機能を 持 った器官 によって行われる。Fayeら ⑮は予めバ クテ リアを注射 して免疫化 した 打 cισψ力 蛹の脂肪体組織を 加 ッF肋 で培養 した場合でも抗菌性タンパク質を合成すること を示 した。また、Dunn6DらはM∫
α協幼虫の脂肪体をPG存
在下で培養すると培地中に抗 曲 性タンパク質が誘導されることを明らかにした。誘導経路を解明する上で昆虫の生体そ のままを用いる 加 めり の条件では体液中の血球の関与を除外できず、明確な答えが得 ら れ難いため、Dunnらのように 加 ッ,rroの条件下で行 う実験系は、バクテリア感染から抗苗 性タンパク質誘導までの経路を分子 レベルで解明する上で、有効な手法であると考えられ 13「
る。
本章では、カィコ生体内のバクテリアに対する複雑な生体防御システムを簡略化 し、そ の解明のための知見を得るためカイコ幼虫脂肪体の 加 ッJ胞 での抗菌性タンパク質誘導系
第
2節
材邪l及び方法 2‐1本オ米半 (1)カイコ カイコ (段 嚇 胴9ri C108)は 人工飼料 (ヤクル トクロレラ配合飼料原種稚蚕用)で
努℃、短 日条件下にて無菌的に飼育 し、5令
3日 目又は4日 目の幼虫を用いた。 孵化直前の卵を70%エ
タノール (1分間)、3%ホ
ルマ リン (15分 問)100%エ
タノール (1分間)の
順で浸す ことで無菌処理 した後、オー トクレーブで滅菌 した濾紙で水分を除 去 し、風乾 した。通気性のシリコン栓を施 したメリクロン培養用フラスコ ●CXIHu容)に
人工飼料を入れ、オー トクレーブしたものを用意 し、この中で無菌処理した卵を孵化させ、 無照明で飼育 を継続 した。無菌飼育 したカイコ幼虫を転が した栄養寒天培地プレー ト上で 転が し、 これを3‐
で2日
間イ ンキュベー トし、パクテリア生育の有無を調べ幼虫体表 面が無菌状態であることを確認 した。また、以上の無菌操作はすべてクリーンベンチ内に て行った。(2)バ
クテ リア aπ湖】∫′たル勁協成,IF0 12195〕 B∝れ,′肋∫脇q″茄 "“IF0 3970,晦
∝∝cDJd JJJ″州IF0 3333= E記″ガcカカ じο″K121F03301,は
大阪発酵研究所より入手 した。セクロピン感受性菌であ る こ ω″D22●rr)は、ス ッ トクホルム大学 (スウェーデン)の
H.Stther博 士よ り譲 り受 けた。その他のバクテリアは研究室保存苗を用いた。13)試
薬 [3刺_リジンはアマシャム・ジャパ ン社より、二 じο′,0111:B4由 来 リポ多糖、E G9ri F588 由来 リピドA、S“
cttOttceS側り 'Й9由
来ザイモサ ン、 勘 加励 兜 'お ″ι由来ラミ ナ リン、アジュバ ン トペプチ ド、説ψり 鋸ο “β 例厖"dのベプテ ドグリカンを構成するペン タ ベ プ チ ド (A協―D‐iso」umminyI_Lys―D‐A協ぃD工la)、 シ ク ロ ヘ キ シ ミ ド、 ア ク チ ノ マ イ
シンD、 卵白リゾチーム及びM IncMs凍結乾燥菌体は 罰
gn4社
より購入した。 また、 キチンオリゴ糖 14糖:5糖 :6糖 :7糖 :8糖 :9糖
)=6.8%:46.9%:16.8%:12.6%:
10.3%:6.6%=M%)、 キ トサンオリゴ糖 “ 糖:5糖 :6糖=19,6%:48.2%:32.2%)は
、 鳥取大学農学部 平野茂博 名誉教授より譲り受けた。 152-2 方法
(1)カイコ幼虫か らの脂肪体組織の描出及び培養
脂肪体摘出に用いるカイコは解音J直前に
70%エ
タノール、滅菌水の順で体表面を洗浄 し、解剖皿内に虫ピンで固定化 した後、氷冷した鱗翅目昆虫用Ringer常猟110mM KCt,4mMNaCl,15mM MgCL,4mM CaCL.5mM K―
phosphatc pH6.5)を 満 た した 。腹 部 を縦 に切 開 し、 切 り口を広げピンで固定化した後、脂肪体以外の組織を除去した。脂肪体の付着したまま の体皮を解剖皿に固定化したまま、体液の除去を目的とし氷冷Ringer液で2回洗浄した。 クチクラ層を傷つけない程度で、スパテルを用いて脂肪体を掻き取った。描出した脂肪体 はGmce培
地に移 し、"℃
、1時 間放置し、残存する血球を除いた。組織培養 に用 いた
Gmce培
地 ①BCO社
製)は
ベニシリンGを
10,OCXI U 偽′耐 、ス ト レプ トマイ シン硫酸 を 100 μ g/1dとなるように力日えた後、0.22μmの
メンプ レンフィルターでろ過滅菌を行ったものを用いた。湿重量で約 鯨 g程度の脂肪体を200 μIの
Gmcc培
地を含むガラス試験管 (15×105団■
)に
移 し、27℃で振とう培養 した。一定時間培養後の培地は抗菌活性測定に用い、脂肪体組織は遠心して集め、タンパク定量を行った。また、 解剖及び培養に用いた器具類はすべてオー トクレーブあるいは乾熱殺菌したものを用いた。
鱗翅目昆虫組織培養用
Gmce培
地 ωCompoment
lnorganic salt CaC12M8C12
M82S04
KCI
Na2HP04
Ari員■O aClds β‐Alaninc L‐Alanine LぃArgininc,HcIL‐Aspartic acid,free acid L‐Asparagine,anhydrous L‐Cystine,dihydroch10ride L―GIutamic acid Glycine,free base L―Histidinc,free base L‐Isoleucine L‐Leuclne L‐Lysinc,HCI L‐Methioninc ( L―Phcnylalaninc L‐
ProHne
DL―Scrine L―Thrconine LぃTryptophan L―Tyrosine,disodium L―Valine mg/1 1.000 1,068 1。357
2.240 0,876 0.200 0.225 0,700 0,350 0,350 0.025 0.600 0,650 2.500 0.050 0.075 0。625
0.050 0,150 0.350 1.100 0。175 0,100 0.072 0.100Component
Vittmlns p―Anunobenzolc acld Biotin Chohne chioridc Folic acid myO_Inositol Niacin D―Pantohenic acid,hemicalcium Pyridoxinc HCI Ribonavin Thiaminc Other D(‐)―FructoscFumaric acid,Free acid
D(+)―Glucose
αぃKeto81utaric acid
L(ぃ)―MaliC acid,frcc acid
Sucrose mg/1 0,000020 0.000010 0.000200 0.000磁0 0.000020 0.000020 0.000020 0.000020 0.000020 0,m1020 0。400 0.055 0.700 0,370 0.670 23.680
Sodium B,carbOna鯰
を
Ot35 m邸となるように
カロえ た 後 、'lN HCIある い は
lN NaOHを
用 いpH6.3と した。(後のフィルターろ過によ り
pH
が0.1刊
B上
昇することを見越 してpHを
6.5とした。実際は最終pH6.5)
(2)抗
菌活性の測定 セクロピン活性の測定には、二G9JiD226仔)を検出菌とした孔平板法6oにより行った。 まず、40℃に保った1%ブ
ィョン培地 (1%低 温グル化寒天、1%肉エキス、1%ペ
プ トン、 0.5%NaCI、 PW・5)Ю
耐 に 対数増殖期のだ。ω,D22、 2×107 cclに を加え、シャーレに広 げ、検出菌を含んだ寒天プレー トを作成した。このプレー トに直径2皿■ の孔をあけ、抗 菌活性の測定のための試料を2 μl注入し、37℃で一晩インキュベー トした後、孔の周 り に形成されるバクテリア生育阻止円の直径を測定し、カイコセクロピン 19に より作成し た標準曲線と比較して活性を求めた。lμ8の
カイコ精製セクロピン相当量をlunitとし、 各試料の抗菌活性をセクロピン相当値のu t数で示した。 (3)リ ゾチーム活性の測定 リゾチーム活性の測定はM加
セ】∫乾燥菌体の濁度減少を測定することで行った。 0.05 M Phosphatc bllfcr,PH 6.5に 肱 JItrf郎乾燥 菌体を0.2m8綱 となるように懸濁 した。 リ ゾチームを含む各試料溶液 50 μlを 950 μIの乾燥菌体懸濁液に加え、Ъ∽Inに おける濁 度減少を30℃で継統測定した。1分
間に吸光度が 1,0減少 したときのリゾチーム活性を 1 unitとして表した。また、l unitはカイコリゾチーム約2μ8の活性に相当する。 14)タンパク質の測定 培養後の脂肪体に1耐
の0.lN NaOHを加え、超音波破砕装置によってソニケーション(10W,3随
)し、可溶化 した。ウシ血清アルブミンを標準試料 とし、BrtldFord法 OJにより可 溶化 した脂肪体タンパク量の測定を行い、各試料中に含まれるセクロピン活性あるいはリ ゾチーム活性を脂肪体タンパクlmg当た りのu t数で示 した。 (引 脂肪体内への[3H]_リ ジンの取 り込み 脂肪体培養系へ[3H]_リ ジンを添加 しタンパクヘの取 り込みを測定する場合、用いた培 養液は Trenczckら 701の用いたGmce培
地改良型を参考 にし、 リジンを除いた培地組成で 調製 した。通常の脂肪体の培養 と同様、カイコ5令
3日 日の幼虫より描出した脂肪体を10 μCiの [3H]_リ ジン及び0.5mMの
非標識 リジンを含む培養液5∞μl中 、27℃で振 とう培 養 した。 また、[3H]_リジンを加えた直後にTCA処
理 した場合をネガティブコントロール とした。 一定時間イ ンキュベー トしたサ ンプルを 10001pmで
5分
問遠心 し、組織 と培養液に分 けた。脂肪林組織は lCIIMリ ジンを含む5%TCAを
lml ttΠえた後ガラスホモジナイザー を用いてホモジナイズし、3000rpmで、lo分間遠心 した。沈殿物を lndの 5%TCA、 lml のエタノーブ膠エーテル(1:1)で それぞれ2回
づつ遠心洗浄 し、脂質を除去 した。残った脂 肪体 由来のタンパクの沈殿物に lndの O.lN NaOHを加え可溶化 し、これに トライ トン‐キ シレン系のシンテ レーション液 を 6ndカロえ脂肪体への[3H]…リジンの取 り込み量を測定 し た。 インキュベー ト後の培養液中へ分泌された可溶性タンパク質の場合、遠心して得 られた 培養液 にBSAを
最終濃度0。lm8/mu、 リジンを10mM、TCAを
1079になるように添加 し、可溶性タンパク質を沈殿 させた。 これを3000rpm、
10分
間遠心 し、得 られた沈殿に組織 の場合 と同様に5%TCA、 エタノーブレエーテル (1:1)の順で2回遠心洗浄 し、0。lN NaOH で溶解後、液体シンテ レーションカウンターによ り取 り込まれた[3H]_リ ジン量 を測定 し た。(6)ポ
リアクリルアミド電気泳動 タンパクの電気泳動は Hdtmarkら 1つ の方法に従い、PH4.3の 酸性電気泳動を行った。[8H]_ リジンの取 り込まれたタンパクを電気泳動する場合、イ ンキュベー ト後の培養液に8Cl% 飽和 となるように硫酸アンモニウムを加え、得 られた沈殿を30 μIの O.lMリ ン酸ナ トリ ウム緩衝液、pH6.5に 再溶解 したものを泳動用試料とした。laskeヅリの方法に従い、フルオログラフィcDMSO―
PPO系
)によって泳動後の[3H]標識 されたタンパク質の検出を行った。また、
X線
フィルムは前露出し、増感板を用いた。第 節 結果
(1)培
養脂肪体によるタンパク質合成の確認 脂肪体組織による 加 ッ,肋 での抗菌性タンパク質の誘導系を確立するために、摘出した 脂肪体組織を用いて 加 ッ,胞 で正常なタンパク合成が行われることを確認した。セクロピ ンのアミノ酸配列よリリジン含量が高いことに着日し、抗菌性タンパク質の合成を示す指 標として培養した組織への[3H]_リ ジンの取 り込みを調べた。培地には鱗翅目昆虫の制胞 培養に一般的に用いられているGmce培
地より、リジンを除いたものを用いた。 脂肪体組織への[3H]_リジンの取 り込みは少なくとも培養開始48時
間まで継続して認め られた 仰8・ Ⅱ■)oまた、組織培養液をTCA沈
殿して得 られる可溶性画分も脂肪体細胞 タンパクの場合と同様に、培養時間の経過に従い[3H]_リ ジンの取 り込み量が上昇した。 この結果は少なくとも48時
間は脂肪体組織の培養が可能であり、可溶性タンパクの分泌 を含め、生合成系が維持されていることを示唆している。「 巧 ど 、 角 o 畠 拍 o ● ・ 日 盈 口 D 冨 ﹂ 佃
含
︻ o
ぢ
角 働
m
日
が
ち
=
x
日
働
e
60
20
20 30 40
1ncubation tilme(hr.)
Hg.Ⅱ
‐l PrOtein synhesis in fat bOdy cultured,ηッ′″ο
Fat body isOlated from 5h instariarvae was cultured in GHlce's Hlediu】
n wtth
習胤∬
鰍 躍篤毛
思鞘詮賞鰍船ξ
織絡星
鶏伊∝
TCA precipitated ttac償 Ons were meisured・
(2)脂
肪体培養系における抗曲陛タンパク質の誘導 細胞内及び分泌性タンパクの合成が 加 ッ,胸 での培養脂肪体でほぼ正常に行われている と考え られたことか ら、,ヵ ソ,拗 での抗菌性タンパク質誘導を試みた。カイコ抗菌性タン パク質は、PGを
注射することで誘導されることか ら、あらか じめPGに
より免疫化 した 幼虫の脂肪体は培養条件さえ整っていれば 加 朔乃 においても抗菌性タンパク質の合成を 行 うものと推測される。そ こでEω
れ可溶性 PG 10μ8を幼虫に注射 し、体液中に抗菌活 性が認め られる6時
間経過後の幼虫脂肪体を培養に用いた。この免疫化 した脂肪体を培養 す ると、培養開始直後か ら抗菌活性が培地中に検出され、その後24時
間まで分泌されて いること力湘召め られた Oig.Ⅱ-2)。 この脂肪体培養液に E・ coli SPGを 50μ3/ndとなるよ うに添加 した場合、培養開始5時
間までは無添加で培養 した場合 とほぼ同じ抗菌活性を示 すが、雰 時間後には数倍高い活性が認められた。 無処理の幼虫か ら描出した脂肪体を培養すると、培養開始か ら、7時
間までは培地中に 抗菌活性は検出されなかった。さらに培養を続けると24時
間後 にわずかに抗菌活性が検 出された CFig.Ⅱ_2)。 この無処理の幼虫脂肪体に 舒℃ を添加 して培養すると、10時間後 よ り顕著な抗菌活性が認められれ免疫化 した幼虫脂肪体 と同様に培養時間に伴う抗菌活性 の増加が認め られた。 この 滋 ッ,物 の培養では抗菌活性 と共にリゾチーム活性も誘導された仰 8・ Ⅱ-3A)。 培養 に伴 うリゾチーム活性の分泌は抗菌活性の場合 とは異な り、SPG無
添加の場合で も培養 開始30分
か ら認められた仰8・ Ⅱ-3B)。 また、培養開始から5時
間経過までではsPGの
添加の有無による違いは認め られず、同程度のリゾテーム活性力島検出されたが、SPG添
加 した場合は12時
間後か らリゾチーム活性の顕著な増加が認められた。その結果、培養 か ら48時
間経過後の培地中のリゾチーム活性はSPG無
添加にくらべ約3∼4倍
程度高 く なった。 この抗菌活性 とリゾチーム活性を持つタンパク質の同定を目的とし、SI℃ 添加培地 と 無添加培地を用いて脂肪体 を24時
間培養後、それぞれの培地を試料 として酸性電気泳動 を行ったo8。 Ⅱつo'カ ツ 'rrDの培養系において脂肪体か ら分泌したタンパクの染色パター ンcane3,oは、比較対照として用いたカイコ幼虫体液の場合canel,2)とほぼ同じであった。 また.SPGを
注射し抗申性タンパク群を誘導させたカイコ免疫化体液仰ne2)との比較から、SPG添
加培地を用いて培養した脂肪体からセクロピンとリゾチームが分泌されていることが示された tlane 4)。 また、HB.Ⅱ
4の
フルオログラフィの結果は培地に添加した3H_ リジンが新たに合成されたタンパク質に取 り込まれていることを示しており、培地中に分 泌されたセクロピンとリゾチームは、培養脂肪体において 虎 胞"合
成されることが明ら かとなった。さらに、予めシクロヘキシミド、あるいはアクチノマインンDを
培地に加 え、前培養 した脂肪体ではセクロピンとリゾチームの合成が阻害されたことも、これら抗 菌性タンパク質の活性発現が細胞内プールから放出されたものではなく、詑 “θ刀 合成で あることを支持しているTable`Ⅱ ‐1).r 省 ︻E 2 磁 m 日 ヽ ∽ ︺ , H目 目 ︶ b F F 常 ﹃ ■ o 一o s p ︻H く I■1lnunized F.B.
+PG
Non― treated F.B.+PG
Non―treated F.B。Time(h)
Fig.Ⅱ -2 Bactericidal activity in fat bOdy cuhures.Silkwon11larvae were
itteCted with 10μ g Ofだ
。
cο′
,sPG,then kept at 28℃ fOr 6h,Fat bodies fЮm
itteCted(●,O)Or non_treated(▲ ,△)larvae were dissected out and incubated inGrace's medium with additiOn of 50μ g/mi Of β.ω″SPG(solid Symbols)Or saline(open symbols).Aliquots of medium were withdrawn atintervals and
assayed for bacteicidial activity against E.じ οど, D22.Values are the means of
three deteニュ▲linations.
r
含
増9
9
儀
∞
日
\望
召
β
︶
掛
F
F
o
く
30
20
Cecropin
Added PG
None
Lysozyコ
te0 10 20 30 40 50
Time(hr)
Fig.Ⅱ -3 1nducdon of bactedcidЛ and lysozyme activities by Stt in cultlllred fat body.Fat body fЮm aseptically reared untreated lawae was incubated with(o)Or without(o)β.Cο″
SPG(拗
μg/ml)。 AliquOts of medium were withdrawn atinteⅣalsand assayed for bac俺 cidal?り and lysozyme(B)act ities,Values and bars are the
mealls± sD ofive deteminaions.
10
2
I II I
Coonlassie Blue R-250
Fluorograpy
五Юソ,チ″οI I
=カッ
J7ο 肋 ッJ″οFig.Ⅱ 望 Acidic PAGE of he larval hemolymph protein and protein secreted from fat
body culture.Left Paneli protein was stained wih Coomassie Blue R「 250.Lane l and 2,
hemolymph(10 μ
l)fЮm ulltteated ttd ttω′
,SPG ttecttd lttae,respecivdμ lane 3and 4,fat body culture medillln(20
μ
l)inCubattd for 24 h wittoutand wih 50μ
g/d
ofE.じ οJ,SPG,respecivelyo Righ Panel:■uorograpy of FH]… labeled PЮteinsynhesized加 ガ″ο culture wihout(lane 5)md with(lane 6)50μ
g/m ofsPG for24
h.A,B:cecropin A and B;L:lysozyHle.r
Table l. Effect of actinomycin D and cyciohexilnide on inducdon of pЮteinst Fat bodies were preincubated with actinomycin D(50
cyciohe
mide(lmtt at 27℃
for 30min in Gmce's medium and incubated for"lhr wih Or wihOllt50μ g/mIPG.antibacterial
μ
g/ml)or
then funerInhibitor dose
Act ity(uniⅣm8 PЮtein)
CecЮPin
Lysozyme
Control ttPG Control 十PG
None Cycioheximidelm M
16,4 N.D, N.D. 38,1 N.D。 N.D. 52,2 2.4 8.7 96.033
6.8Acinomycin D 50μ
8/mlNoDoi not det∝tled
(3)バ
クテ リア菌体による抗曲性タンパク質誘導 抗菌性タンパク質の誘導実験の多 くは、カィコ生体に直接バクテリアやPGな
ど誘導因 子 を注入することで行われた。このような 加 "ο での実験系ではバクテ リア感染からそ のシグナルが脂肪体に伝わるまでに血球細胞を含め多 くの体液成分などが存在 し、抗菌性 タンパク質の誘導経路を解明する上で血球を含めた体液の関与を除外することが出来ない。 そ こで抗菌性タンパク質誘導における血球の必要性について検討するため 加 胡ヵ の系に バクテ リア菌体を直接加えて これまでと同様に脂肪体の培養を行った。また、バクテリア イまLPSの
影響を避けるため、LPSを
構成成分に持たないグラム陽性苗のB.施
施 “っ栃減♂を 実験に用いた。β.施施"紳
述Sの細胞壁PGは
E.coliのPCと
同じくペプチ ド側鎖が直接 架橋 した構造 を持ち、高い誘導効果を示す ことが明 らかとなっている。UV照
射 し殺菌 し た β.施ル卵 を5×108 cc17mlと なるように培地に加え、これまでと同様に脂肪体の 培養を行った仰gⅡ_ぅ。Pc添
加の場合8時
間経過後にセクロピン活性が検出されたが、 バクテリア菌体を加えた場合10時
間以降よ り検出された。 しか し、その後PGと
同様に 培養時間に従いセクロピンとリゾテームの顕著な誘導が観察された。この結果はカイコ生 体内に侵入 したバクテリアによる抗菌性タンパク質誘導に血球は必要でないことを示唆し ている。30
Cecropin
PG
Bacte al censNone
Lysozy]me
PG
Bacte al censNone
0 10 20 30 40 50
、
Time(hr)
Fig.IIⅡも inducior ofibacteicidЛ and lysozyme acuvities by SPG orげ V―killed
bacte a in cul red fat body,Fat body fЮ m aseptically reared lawae was culttred with
SPG from】
.胸鋸″Tttμ(拗
μg/ml,o),げ
V―killed B.′,ご乃?“ゎ 翻 ね (5× 108cells/nu,▲)。「no addition(日), Aliquots ofmedium were withdmwn ati eⅣals and
assayed for cecЮ
∬
n(llpper panЫ)and lysozyme(10Wer panel)acti ties・ Vdues andbars are the means 量三 SE)offour deterlmnadons.
40
20
︵
。
g
日
9
日
∞
日
\望
狙
こ
う
T
F
o
く
10
10
6
4
2
29(4)可
溶性ペプチ ドグリカンによる抗曲陛タンパク質誘導効果 この 加 ッヵ聰 での抗菌性タンパク質誘導について、誘導因子であるSPGの
構造 と誘導効 果 との相関性を検討した。SPGを
カイコ幼虫へ注射 して検討した 加 ッ肋 の実験結果によ るとaω
ヵのSPG注
射では高い抗菌活性の誘導が認められるが、舷 肋鉗∫由来SPGの
誘 導効果は低いことが明 らか となっている 丸)。 そ こで、それぞれのパクテ リアよ り調製 し たSPGを
培地中に0,C1350μ B/Hdと なるように加え、脂肪体を%時
間培養 し、培地中に 誘導された抗菌活性を調べた 仰8・ Ⅱつo SPG無
添加の場合に比べ、二clo,SPGを加え た ものは、0.o5μ8/ndの 濃度よ り抗菌活性の誘導が認め られ、測定 した 50μ 8/ndの 濃度 まで添加量に従い抗菌活性の上昇が認められた。一方、M「
Irr91rd sPGの場合、5μ 8/11uの 添加まで、顕著な抗菌活性の誘導は認め られず、ほぼコン トロールと同じであった。実験 で用いた最大添加量である 側 μg/ndに おいて抗菌活性が表れたが、その抗菌活性は 14士 2 units/mg pntehで、 この値はa c9ヵsPGに
換算 した場合、0.lμ 8/Euの S〕℃ 濃度で得 ら れる活性に相当する。 このことはM脇
♂ "∫sPGは
、ユ じθ″sPcに
比べ 1,期 程度の誘導効 果 しかない ことを示 している。さらに誘導因子の構造特異性を調べるため、その他のグル カンやペプチ ドを用いて同様に,■ ガ施 での誘導を検討 したTable.Ⅱ_2)。 先の実験 と同様、 培地中に各種グルカンなどを50μ8/耐 となるように加え、24時
間培養後の培地中に誘導 された抗菌活性を測定 した。昆虫の外骨格を構成するβ■,牛グルカンのキチンやその脱ア セチル化物のキ トサ ンは哺乳類で免疫賦活作用があることが報告されている 729。 バクテ リア細胞壁PCの
構成糖の一つである N‐アセテル グルコサミン (GluNAc)のポ リマーで あ り、PG糖
鎖 と同様 β‐1,4結合であることか らこの糖鎖の誘導効果を検討 したが、コン トロール と同程度の活性 しか得 られなかった。また、単独で 50μ8/m!添加 しても効果が認め られなかった
PGの
ペプチ ド部分であるPGド
plidcぃ Й―DlsoGlu―Lys― D―Ala―D‐Aめ
をキチ ンあるいはキ トサ ンと組み合わせ 同じ培地中に添加 したが、 この場合 も顕著な誘導は 認められなかった。
PGと
共にフェノールオキシダーゼ系を活性化する誘発因子として知 られるβ■,3-グルカンを構成糖に持つラミナリンや酵母細胞壁成分であるザイモサンによ っても抗菌性タンパク質の誘導は得 られなかった。さらに、PGの
部分構造であり、アジ ュバント活性を持つアジュバン トベプチ ドにおいても効果は認められなかった。これらい ずれのPG類
似物質とも抗菌性タンパク質の誘導因子としての効果は無く、この結果は 加 れりで得られた結果と一致したTable Ⅱ_3)。︵
・
9
日
9
缶
押
要
望
召
こ
う
F
F
o硝
瑠
唱
g
驚
つ
F
目
く
! 1Soluble PG(μ
g/ml)
Fig. Ⅱ_6 Dose dependentinduction of bacteicidal activity in fat bOdy by
SFDG from E.じο′,and J躍″じ′οじοじじИ♂′
"チ
?′∫.Fat body froni aseptically reared
untreated laⅣae was incubated with the indicated cOncentrations of SF)(3 from
だ。じο″
(o)Or財
'σ
οじοcじ
"s'"彪′∫(o)fOr 24 h,Valued and bars are the means +SD of ive dete.二 二二inations.
β
.じο′
,Table Ⅱ-2 Effect Of vaiOus glucans and peptides on indllction of bactericidal
activity in cuitured fat body.Fat body from aseptically reared laⅣ ae was cultllred with
the addi償Ons Of various materiЛ s at a concentmtion of 50μ
ゴ
ml,and incubtted for%h. Values are he nleans tt SD ofive dete∬ 皿natiOns.
Matenals added
(50 μ g/ml) Bactericidal activity (unitS/mg pЮtein) ExP.1 NoneChitin oLgomer
C
tosan oligomerPG peptide
Chitin oligorner+PG pepdde
Chitosan ongotrler+PG pcptide
E.じοJli PG
ExP,2
None
hminarin
Zymosan
Attwant pepide
E.じο
" PG
14.2±2.8 18。9±
4.7 14.0±1.6 1867ΞL 10.0 18.3±4.1 18.1」L2.6
46.5三上
0。6*
9.OΞL2.0
10.OΞL3.8
7.7±3.0 5.9EL l.631.6EL7.0*
*Sig ncalldy greater than control,P<0,01
Table
Ⅱ
_3 SpedicinducdOn ofcecЮ
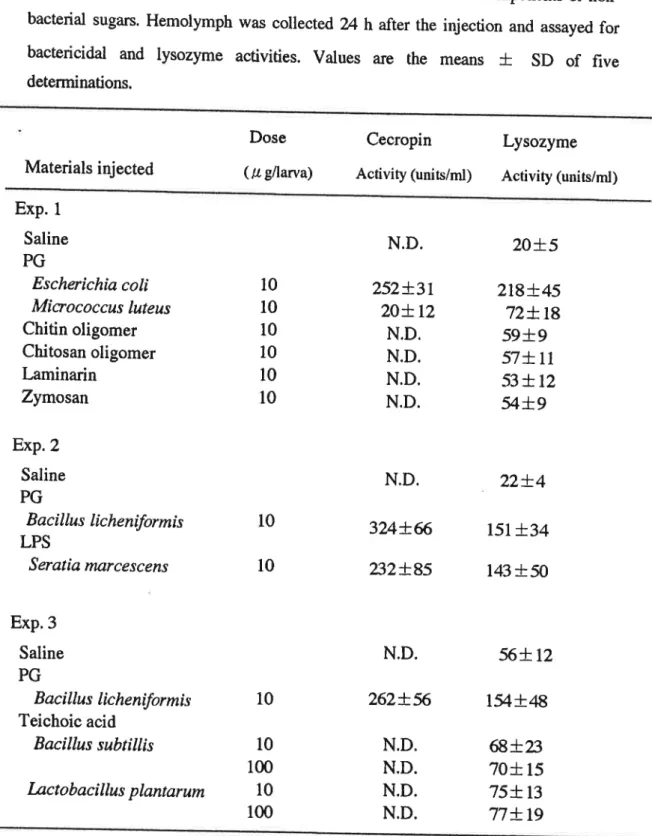
ttn andlysOzyme、by SPG and LPS,SilkwoH二二▲laⅣae were inJected witt variOlls bacteial cell wali cOmPonents Or non― bac俺五d sugars,Hemolymph was cdlected 24 h after tte ittecdOn and assayed fOr
bacte cidal and lysOzyme aciviues, Values are the means ±
sD of flve
detemillatiOnsむ Mateinis inJected Dose (μ gttarva) CecroPin Ac償 ty(unitS/mu)
LysOzyme
Acnvity OnitS/nd)ExP,1
SalinePG
Es》T,》
滋 じο′ れイ,Tοcοcc"∫ ′ "′ ?′s Chitin oligomer Chitosan oligomer Laminan
Zymosan
ExP.2
Sahne
PG
βαO′JBI♂ Jた力?刀:″′閉 ねLPS
STα′,α ′″,″じ?Sc?力∫ExP,3
SalinePG
Bα o,Jル♂ι
ich?ヵ!″′
“
ね
Teichoic acid Bαc,JJpJ∫ "♭ ガJJね 盈zcゎうαO′肋dP肋
刀勉r"加 10 10 10 10 10 10N.D.
252± 31 20± 12 N.D. NoD. N.D. NtD. N.D. 262」L56
N,D. N.D。 N.D. N,D. 20」L5
218士心 72± 18 59」L9
57±11 53± 12 54±9
10 10 N.D. 324±66
232」L85
22±4
151±34
1弔±50
56± 12 154±48
68± 23 70±15 75±13 77±19 10 10 100 10 100N.D.I not detec団