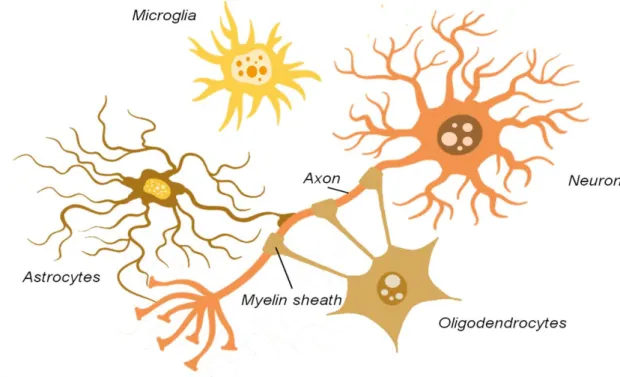
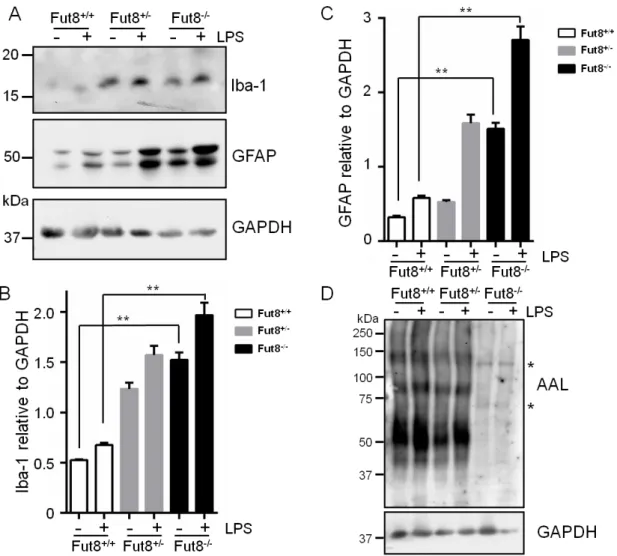
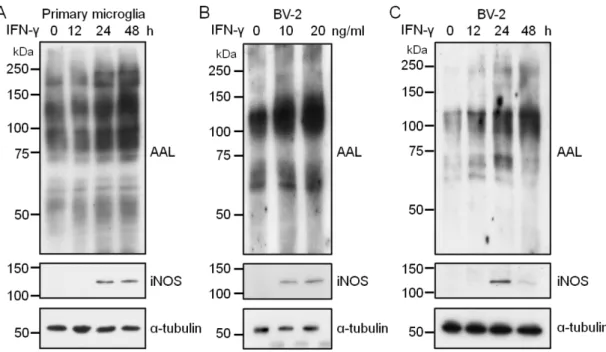
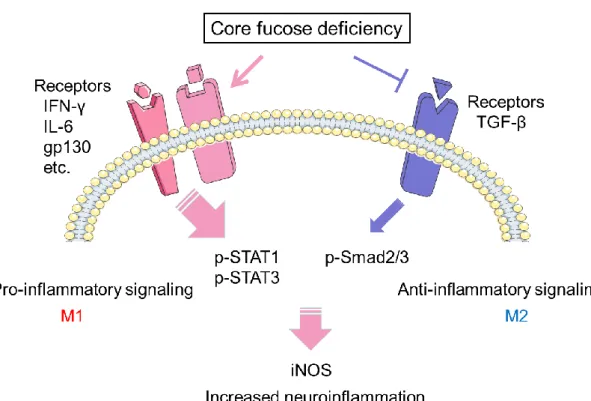
関連したドキュメント
1 Introduction and overview 1.1 Introduction 1.2 Model of the public goods game 2 Expectation of non-strategic sanctioning 2.1 Introduction 2.2 The game and experimental design
Jinxing Liang, Takahiro Matsuo, Fusao Kohsaka, Xuefeng Li, Ken Kunitomo and Toshitsugu Ueda, “Fabrication of Two-Axis Quartz MEMS-Based Capacitive Tilt Sensor”, IEEJ Transactions
第3節 チューリッヒの政治、経済、文化的背景 第4節 ウルリッヒ・ツウィングリ 第5節 ツウィングリの政治性 第6節
話教育実践を分析、検証している。このような二つの会話教育実践では、学習者の支援の
(Approximately 4,000 characters in Japanese, or 1,500 words in English. The Doctoral Thesis title, however, must be written in both Japanese and English.).. 博士論文審査委員会
クター(SMB)およびバリューファクター(HML)および投資ファクター(AGR)の動的特性を得るために、特
Cioffi, “Pilot tone selection for channel estimation in a mobile OFDM systems,” IEEE Trans.. Sunaga, “Rayleigh fading compensation for QAM in land mobile ra- dio communications,”
ている。本論文では、彼らの実践内容と方法を検討することで、これまでの生活指導を重視し