東北薬科大学
審査学位論文(博士)
氏名(本籍) クドウ アツシ
工藤 敦(宮城県)
学位の種類 博士(薬科学)
学位記番号 博薬科第
1
号学位授与の日付 平成
27
年3
月17
日学位授与の要件 学位規則第4条1項該当
学位論文題名
Aspergillus
属菌の細胞壁ガラクトマンナンの構造及 び増殖条件によるその変化の解析論文審査委員
主査 教 授 大久保 恭 仁
副査 教 授 山 下 幸 和
副査 教 授 柴 田 信 之
Aspergillus 属菌の細胞壁ガラクトマンナンの構造 及び増殖条件によるその変化の解析
東北薬科大学大学院薬学研究科 感染生体防御学教室
工 藤 敦
目
次
序
章………
1
第一章
異なる培地で培養した
Aspergillus fumigatus
のO-
結合型オリゴ糖の構造解析 第一節 緒論……… 5第二節 実験材料及び実験方法……… 7
第三節 結果………12
第四節 考察………20
第二章 異なる培地で培養した
Aspergillus fumigatus
のN-
結合型ガラクトマンナンの構造解析 第一節 緒論………22第二節 実験材料及び実験方法………24
第三節 結果………29
第四節 考察………44
第三章
Aspergillus niger
及びAspergillus terreus
のN-
結合型ガラクトマンナンの構造解析 第一節 緒論………48第二節 実験材料及び実験方法………50
第三節 結果………52
第四節 考察………60
総 括………62
論文目録………65
謝 辞………66
引用文献………67
序
章
我 々 人 類 は 細 菌 や 真 菌 と 密 接 に 関 わ っ て 生 活 し て い る . 人 間 の 体 に は
100
兆 個 以 上 の 常 在 菌 が 生 息 し て お り , 特 に 腸 内 細 菌 は 人 間 の 免 疫 の 調 節 に 関 与 し て い る と 言 わ れ て い る 1 ). ま た , 人 間 は 嗜 好 品 の 製 造 や 環 境 保 全 の た め に , さ ら に バ イ オ テ ク ノ ロ ジ ー の 分 野 で も 細 菌 や 真 菌 を 利 用 し て き た .一 方 で ,人 類 は 細 菌 類 に よ る 感 染 症 に 苦 し め ら れ , 死 亡 原 因 の 大 半 が 感 染 症 と な っ て い た 時 代 も あ っ た .こ の 感 染 症 の 治 療 に 大 き く 貢 献 し た の が 抗 菌 薬 で あ る . ペ ニ シ リ ン は 世 界 初 の 抗 菌 薬 と し て
1929
年 に 発 見 さ れ ,特 に 第 二 次 世 界 大 戦 の 際 に は 負 傷 後 の 破 傷 風 に よ る 感 染 症 に 広 く 使 わ れ た . そ の 後 セ フ ェ ム 系 抗 菌 薬 の 発 見 や 全 合 成 に よ っ て 作 ら れ た ニ ュ ー キ ノ ロ ン 系 抗 菌 薬 な ど 多 く の 抗 菌 薬 が 作 ら れ , 感 染 症 の 治 療 は 急 速 に 進 歩 し た . 例 え ば , 放 線 菌 か ら 発 見 さ れ た 抗 菌 薬 で あ る ス ト レ プ ト マ イ シ ン や 結 核 に 効 果 が あ る イ ソ ニ ア ジ ド , ピ ラ ジ ナ ミ ド な ど の 抗 結 核 薬 の 発 見 は , か つ て1
位 で あ っ た 結 核 の 死 亡 率 を 現 在 で は26
位 ま で 低 下 さ せ た .し か し 抗 菌 薬 の 開 発 の 一 方 で , 細 菌 類 の 抗 菌 薬 に 対 す る 耐 性 化 が 問 題 と な っ て い る . 例 え ば ペ ニ シ リ ン に 対 す る 耐 性 菌 は 実 用 化 か ら 間 も な い
1940
年 台 に 発 見 さ れ て い る .ペ ニ シ リ ン の 作 用 機 序 は 細 菌 類 が 有 す る ペ ニ シ リ ン 結 合 タ ン パ ク 質 に 結 合 す る こ と に よ る 細 胞 壁 合 成 阻 害 だ が , 耐 性 化 は そ の タ ン パ ク 質 に 変 異 が 起 こ り ペ ニ シ リ ン の 結 合 能 が 低 下 し た た め に 生 じ た . 人 類 は こ の 耐 性 菌 に 対 抗 す る た め の 抗 菌 薬 と し て メ チ シ リ ン を 開 発 し た が , 更 に メ チ シ リ ン に 耐 性 を 獲 得 し たM e t h i c i l l i n - r e s i s t a n t S t a p h y l o c o c c u s a u re u s (MRSA)が 発 生 し た .こ の MRSA
は 医 療 施 設 内 で の 院 内 感 染 だ け で は な くcommunity-acquired MRSA (CA-MRSA)と 言 わ れ る 市 中 感 染 型 の MRSA
の 増 加 ,治 療 薬 で あ る バ ン コ マ イ シ ン の 最 少 発 育 阻 止 濃 度 (MIC
) が2 µg/mL
の 株 の 出 現 に よ り 治 療 難 渋 例 が 増 え て い る 2 )と い う 問 題 が あ り , よ り 注 意 が 必 要 な 菌 種 で あ る .一 方 真 菌 症 に 着 目 す る と , 近 年 増 加 傾 向 に あ る の は
Aspergillus
属 菌1
が 原 因 で 生 じ る ア ス ペ ル ギ ル ス 症 で あ り , 剖 検 例 中 の 内 臓 真 菌 症 ( 深 在 性 真 菌 症 ) と し て 検 出 さ れ る 頻 度 が 最 も 多 い 菌 種 で あ る 3 )
(Fig. 1).
以 前 は
Candida
属 菌 の 検 出 が 最 も 多 か っ た が , 新 規 抗 真 菌 薬 の 登 場 に よ り カ ン ジ ダ 症 の 治 療 効 果 が 上 昇 し た こ と 及 び 易 感 染 性 の 病 態 で 患 者 が 長 期 生 存 可 能 に な り ,Aspergillus
属 菌 の 感 染 を 受 け る 機 会 が 増 加 し た た めAspergillus
属 菌 の 検 出 頻 度 が 上 昇 し た と 考 察 さ れ て い る 4 ). ア ス ペ ル ギ ル ス 症 の 代 表 的 な も の と し て は ,Aspergillus
属 菌 が ア レ ル ギ ー 源 と な り 発 症 す る ア レ ル ギ ー 性 気 管 支 肺 ア ス ペ ル ギ ル ス 症,
肺 あ る い は 気 管 支 に 器 質 的 な 病 変 が 存 在 す る 場 合 に 発 症 す る 肺 ア ス ペ ル ギ ロ ー マ,
好 中 球 減 少 , 強 力 な 免 疫 抑 制 薬 及 び 抗 が ん 剤 投 与 時 に 発 症 頻 度 が 高 い 侵 襲 性 ア ス ペ ル ギ ル ス 症 が あ る . こ の 中 で も 侵 襲 性 ア ス ペ ル ギ ル ス 症 は 致 死 率 が58
% と 高 い 予 後 不 良 な 疾 患 で あ る 5 ).Fi g. 1 . わ が 国 の 病 理 剖 検 例 に お け る 内 臓 真 菌 症 の 年 次 別 発 生 頻 度 と 抗 真 菌 薬 の 上 市 年 6 ).
こ の
Aspergillus
属 菌 の 感 染 に は 胞 子 が 重 要 と さ れ て い る が , 空 気 中 に も 多 く 存 在 し 無 意 識 に 一 日 数 百 個 を 吸 い こ ん で い る と 言 わ れ て い る7 ).特 に 建 物 の 建 築 や 改 築 が 近 く で 行 わ れ て い た 場 合 に ア ス ペ ル ギ ル ス 症 の 発 症 率 が 上 が る た め 8 , 9 ),病 院 内 の 空 調 設 備 の 清 掃 や
HEPA
フ ィ ル タ ー に よ る 管 理 が 重 要 と さ れ て い る 1 0 , 11 ).こ の 胞 子 は 健 常 人 で あ れ ば 肺 胞 マ ク ロ フ ァ ー ジ や 好 中 球 な ど の 生 体 の 防 御 機 構 に よ り 排 除 す る こ2
と が で き る が 1 2 , 1 3 ),免 疫 能 の 低 下 し た 患 者 で は 排 除 で き ず に 感 染 し て し ま い 1 4 ),特 に 骨 髄 移 植 患 者 な ど の 好 中 球 減 少 患 者 は 発 症 の リ ス ク が 高 い と 言 わ れ て い る 1 5 -1 8 ).
こ れ ら の 真 菌 症 に 対 す る 治 療 薬 で あ る 抗 真 菌 薬 と し て は ,
cytochrome P450
を 阻 害 し ,細 胞 膜 成 分 の エ ル ゴ ス テ ロ ー ル 合 成 を 阻 害 す る ア ゾ ー ル 系 , エ ル ゴ ス テ ロ ー ル に 結 合 し 細 胞 膜 を 破 壊 す る ポ リ エ ン 系 が あ る が ,近 年 新 し く キ ャ ン デ ィ ン 系 抗 真 菌 薬 が 発 売 さ れ て い る . こ れ は 新 規 作 用 部 位 と し て 細 胞 壁 に 存 在 す るβ-1,3-
グ ル カ ン 合 成 酵 素 阻 害 作 用 を 有 す る が , 真 菌 特 異 的 な 作 用 部 位 の た め 副 作 用 が 少 な い と い う 利 点 が あ る . 一 方 で 細 菌 と 同 様 に 真 菌 の 耐 性 化 の 問 題 が 危 惧 さ れ て お り , イ ト ラ コ ナ ゾ ー ル や ボ リ コ ナ ゾ ー ル 耐 性 のA. fumigatus
1 9 -2 2 ), ア ム ホ テ リ シ ンB
耐 性 のAspergillus terreus
2 3 )な ど が 出 現 し て お り , 今 後 の 耐 性 化 の 進 行 に 注 意 が 必 要 で あ る . ま たAspergillus
属 菌 で は 未 だ 認 め ら れ て い な い が ,Candida
属 菌 で は キ ャ ン デ ィ ン 系 抗 真 菌 薬 低 感 受 性 菌 の 存 在 が 報 告 さ れ て い る こ と か ら 2 4 -2 6 ), 更 な る 耐 性 化 の 防 止 , 新 規 抗 真 菌 薬 の 開 発 が 求 め ら れ て い る .こ れ ら の ア ス ペ ル ギ ル ス 症 の 治 療 に は 早 期 診 断 が 重 要 で あ り , 診 断 法 と し て は 微 生 物 学 的 検 査 , 病 理 組 織 学 的 検 査 , 画 像 検 査 , 真 菌 抗 原 検 査 , 分 子 生 物 学 的 検 査 の 有 用 性 が 明 ら か と な っ て い る . こ の う ち 確 定 診 断 は 培 養 検 査 , 鏡 検 , 病 理 組 織 学 的 検 査 で 行 え る が , 状 態 不 良 の 患 者 に 侵 襲 的 検 査 は 行 え ず 明 確 な 結 果 が 得 ら れ な い 場 合 が あ り , 補 助 診 断 法 と し て こ れ ら 以 外 の 診 断 法 も 用 い ら れ て い る . そ の 中 で も 真 菌 抗 原 検 査 と し て ,菌 体 が 産 生 す る ガ ラ ク ト マ ン ナ ン 抗 原 を
ELISA
法 で 検 出 す る キ ッ ト(Plateria Aspergillu s EIA)は 発 症 早 期 に 短 時 間 で 感 染 の
有 無 の 確 認 が で き る た め 頻 用 さ れ て い る . こ れ は ガ ラ ク ト マ ン ナ ン に 含 ま れ る オ リ ゴ ガ ラ ク ト フ ラ ノ ー ス(Galf )鎖 が 特 異 的 な 抗 原 性 を 有 す
る た め , こ の 抗 原 に 対 す る モ ノ ク ロ ー ナ ル 抗 体 を 診 断 に 応 用 し た も の で あ る 2 7 ).こ れ ま で に 真 菌 の 細 胞 壁 抗 原 多 糖 の 研 究 は 広 く 行 わ れ て い る . 特 に
Candida
属 菌 の 細 胞 壁 の 最 外 層 に 存 在 す る マ ン ナ ン タ ン パ ク 質 複 合 体3
2 8 )は ,宿 主 細 胞 へ の 付 着 2 9 , 3 0 ),マ ク ロ フ ァ ー ジ と の 相 互 作 用 3 1 , 3 2 ),細 胞 壁 の 構 築 3 3 , 3 4 ),病 原 性 へ の 関 与 3 5 )等 に 関 す る 解 析 が 進 め ら れ て い る . 一 方 で 細 胞 壁 ガ ラ ク ト マ ン ナ ン に つ い て は ,こ れ ま で に
Penicillium
3 6 ), Malassezia
3 7 ), Trichophyton
3 8 )で 構 造 解 析 の 報 告 が あ り , ま たGal f
を 有 す る 菌 の 細 胞 壁 糖 鎖 構 造 と し てCladosporium
3 9 )やLactobacillus
4 0 , 4 1 ),Fusarium
4 2 )が 近 年 報 告 さ れ て お り , 自 然 界 で のGalf
の 存 在 が 徐 々 に 明 ら か と な っ て い る .こ のAspergillus
属 菌 が 有 す るGalf
に つ い て は ,A.
fumigatus
に お い て 病 原 性 へ の 関 与 4 3 , 4 4 )やAspergillus nidulans
に お い て 抗 真 菌 薬 に 対 す る 感 受 性 へ の 影 響 4 5 )と い っ た 報 告 が あ り ,ヒ ト が 有 し て い な い 糖 鎖 構 造 と い う 特 徴 を 利 用 し たGalf
の 機 能 に 関 す る 研 究 の 進 展 が 期 待 さ れ て い る .一 方 で ,近 年 遺 伝 子 欠 損 株 の 作 製 に よ るGalf
の 機 能 解 析 は 盛 ん に 行 わ れ て い る も の の ,Gal f
鎖 の 構 造 に 着 目 し た 研 究 は ほ と ん ど 行 わ れ て い な か っ た .現 在 臓 器 移 植 な ど の 医 療 技 術 の 進 歩 に 伴 う 真 菌 症 の 増 加 が 危 惧 さ れ て お り 4 6 ),特 に 内 臓 真 菌 症 の 中 で 検 出 割 合 の 大 き い
Aspergillus
属 菌 は 今 後 更 に 増 加 す る 可 能 性 が あ る . ま た 抗 真 菌 薬 は 細 菌 感 染 症 と 異 な り 治 療 薬 が 限 ら れ て い る こ と や 耐 性 化 の 懸 念 も あ る こ と か ら ,Aspergillus
属 菌 の 性 質 及 び 病 原 性 に 関 す る 研 究 を 更 に 進 め る 必 要 が あ る . そ こ で 本 研 究 で は ,ア ス ペ ル ギ ル ス 症 の 新 た な 診 断 法 や 治 療 薬 開 発 の た め に , そ の 基 礎 と な るAspergillus
属 菌 の 細 胞 壁 構 造 の 理 解 を 目 的 と し , 細 胞 壁 に 特 徴 的 に 存 在 す る ガ ラ ク ト マ ン ナ ン に 着 目 し 構 造 解 析 を 行 っ た . そ の 結 果 ,培 養 条 件 ,菌 種 に よ りGalf
側 鎖 の 長 さ が 大 き く 変 化 す る こ と , 更 に 新 規 糖 鎖 構 造 の 存 在 す る こ と を 明 ら か に し た .第 一 章 で は 異 な る 培 地 で 培 養 し た
A. fumigatus
のO-
結 合 型 オ リ ゴ 糖 の 構 造 の 変 化 , 第 二 章 で は 異 な る 培 地 で 培 養 し たA. fumigatus
のN-
結 合 型 ガ ラ ク ト マ ン ナ ン の 構 造 の 変 化 , 第 三 章 で はAspergillus niger
及 びA. terreus
のN-
結 合 型 ガ ラ ク ト マ ン ナ ン の 詳 細 な 構 造 に つ い て 解 析 し た 結 果 を 報 告 す る .4
第 一 章
異 な る 培 地 で 培 養 し た
Aspergillus fumigatus
のO-
結 合 型 オ リ ゴ 糖 の 構 造 解 析第 一 節 緒 論
糖 タ ン パ ク 質 と は タ ン パ ク 質 に 糖 鎖 が 結 合 し た も の で あ る が , そ の 中 で も
O-結 合 型 糖 鎖 は セ リ ン ま た は ス レ オ ニ ン 残 基 に 結 合 し た も の を
指 す . 人 類 が 有 す るO-結 合 型 糖 鎖 と し て は 免 疫 グ ロ ブ リ ン
4 7 )や ヒ ト 絨 毛 性 ゴ ナ ド ト ロ ピ ン4 8 )の 構 造 が 解 析 さ れ て お り 、 特 にα -ジ ス ト ロ グ リ
カ ン に つ い て はSiaα2→3Galβ1→4GlcNAcβ1→2Man
と い う シ ア ル 酸 を 含 む 糖 鎖 を 初 め4 9 ), 様 々 な 構 造 を 持 つ 一 連 のO-
結 合 型 糖 鎖 構 造 が 報 告 さ れ て い る5 0 -5 3 ).一 方 ,哺 乳 類 と は 異 な りSaccharomyces cerevisiae
な ど の 真 菌 が 持 つO-
結 合 型 糖 鎖 は 全 てO-mannose
型 糖 鎖 で あ る5 4 ).パ ン 酵 母 はα-1,2
結 合 及 びα-1,3
結 合 のmannoseが 1-5残 基 繋 が る 糖 鎖 を 有 し て お
り ,O-
結 合 型 糖 鎖 は 酵 母 の 生 存 や 細 胞 壁 の 生 合 成 に 関 与 し て い る と 報 告 さ れ て い る5 5 ).ま たCandida albicansの 構 造 は S. cerevisiaeの 糖 鎖 と 類
似 し て お りα-1,2
結 合 のmannoseが 2-3残 基 結 合 し て い る が
5 6 ),Cryptococcus neoformans
で はmannose
以 外 にgalactose
やx ylose
を 有 す る 特 徴 的 な 構 造 で あ り5 7 ), 真 菌 のO-
結 合 型 糖 鎖 は 菌 種 に よ っ て 様 々 な 構 造 の 糖 鎖 を 有 す る こ と が 知 ら れ て い る .一 方Aspergillus属 菌 に つ い て の
構 造 解 析 で は ,A. fumigatusの O-
結 合 型 糖 鎖 はGlcp α1→6Man
,Gal f β1→
6Manp α1→6Man
,Galf β1→5Gal f β1→6Man p α1→6Man
,Galf β1→5Gal f β1→
5Galf β1→5Gal f β1→6Man
を 有 す る こ と5 8 )や ,Aspergillus awamoriの glucoamylase I
はmannose, α-1,2-mannotriose
を 有 す る と い う 報 告 が ある5 9 ).ま た 近 年 タ ン パ ク 質 の セ リ ン あ る い は ス レ オ ニ ン 残 基 に
mannose
を 転 移 す る 酵 素 で あ る
protein O-mannosyltransferase
に 関 す る 研 究 も 行 わ れ て お り ,A. fumigatusの AfPmt1p, AfPmt2p, AfPmt4p
6 0 , 6 1 )や ,A.
nidulansの PmtA
6 2 )の 欠 損 に よ り 細 胞 壁 構 築 に 影 響 が 生 じ る こ と か ら ,5
Aspergillus属 菌 が 有 す る O-
結 合 型 糖 鎖 も 重 要 な 役 割 を 有 し て い る と 考 え ら れ て い る . し か し , 様 々 な 構 造 の 糖 鎖 の 存 在 理 由 や 遺 伝 子 欠 損 状 態 で の 糖 鎖 構 造 は 明 ら か に な っ て い な い .こ れ ま で に
C. albicans
をyeast nitrogen base
培 地 で 培 養 し た 際 にyeast extract-peptone-dextrose
培 地 で 培 養 し た 場 合 よ り も 細 胞 壁 表 面 の 疎 水 性 の 増 加 す る こ と が 報 告 さ れ て い る 6 3 ).ま た 同 培 地 で 培 養 し た 際 の マ ン ナ ン 構 造 や 抗 原 性 に つ い て , リ ン 酸 基 やβ-1,2
結 合mannose
が 消 失 し , 非 還 元 末 端α-1,3
結 合 が 増 加 す る こ と や 抗 原 性 の 変 化 す る こ と が 報 告 さ れ て い る 6 4 ).こ れ ら の 結 果 はAspergillus
属 菌 に お い て も 細 胞 壁 糖 鎖 の 構 造 が 変 化 す る 可 能 性 を 示 唆 し て い た . そ こ で , 本 章 で はA. fumigatus
をyeast nitrogen base
培 地 及 びyeast extract-peptone-dextrose
培 地 で 培 養 を 行 い ,得 ら れ たO-結 合 型 オ リ ゴ 糖 鎖 の 構 造 の 違 い に つ い
て 検 討 を 行 っ た .6
第 二 節 実 験 材 料 及 び 実 験 方 法
1. 使 用 菌 株
Aspergillus fumigatus var. fumigatus NBRC 33022
を 用 い た . 菌 体 は 独 立 行 政 法 人 製 品 評 価 技 術 基 盤 機 構 バ イ オ テ ク ノ ロ ジ ー 本 部 よ り 入 手 し た .2. 菌 の 培 養
培 地 は yeast extract -peptone-dextrose 培 地
(0.5% yeast extract (透 析 後
の 透 析 外 液 を 使 用)
,1% peptone, 2% glucose) (以 下 YPD
培 地 と 略 す)
は500 mL
三 角 フ ラ ス コ に 分 注 し オ ー ト ク レ ー ブ 滅 菌 を 行 い 使 用 し た .yeast nitrogen base-galactose
培 地(9% galactose
,0.67% yeast nitrogen base (0.5% ammonium sulfate
, 少 量 の ア ミ ノ 酸 , ミ ネ ラ ル , ビ タ ミ ンを 含 む) )
( 以 下YNB
培 地 と 略 す ) は , ま ずgalactose
を 三 角 フ ラ ス コ に 分 注 し オ ー ト ク レ ー ブ を 行 い ,YNB
は0.22 µm
メ ン ブ ラ ン フ ィ ル タ ー を 用 い て 分 注 す る こ と に よ り 調 製 し た .本 菌 は2% agar
を 含 むYPD
斜 面 寒 天 培 地 で 前 培 養 の 後 , 上 記 の 培 地 に 適 量 移 植 し ,28℃ ,14 日 間 静 地 培 養 し た .3
. 培 養 濾 液 粗 画 分 の 調 製培 養 後
formaldehyde
を 最 終 濃 度 が1%に な る よ う 加 え 一 晩 静 置 し た
後 ,ろ 紙 を 用 い て 培 養 濾 液 と 菌 体 を 分 離 し た .培 養 濾 液 は30-40℃ で 減
圧 濃 縮 し ,2-3
日 流 水 透 析 を 行 い , 再 度 減 圧 濃 縮 し 凍 結 乾 燥 を 行 っ た .4. ガ ラ ク ト マ ン ナ ン の 分 離
培 養 濾 液 粗 画 分 を 精 製 水 で 溶 解 後
3000 rpm,10
分 間 遠 心 分 離 し ,上 清 をDEAE-Sepharose Fast Flow column (5×20 cm)
に か け 水 ,0.1 M NaCl
,1 M NaCl
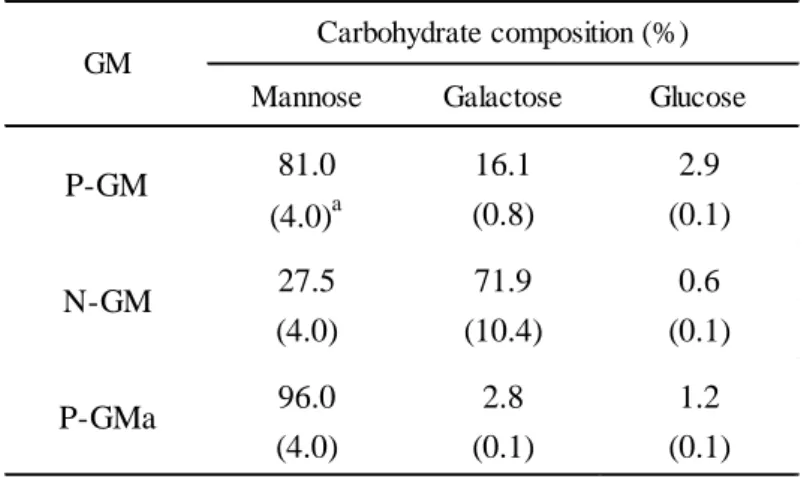
で 溶 出 し た . 溶 出 液 の 糖 含 量 を フ ェ ノ ー ル ・ 硫 酸 法 で 測 定 後 回 収 し ,減 圧 濃 縮 ,2-3 日 流 水 透 析 を 行 っ た 後 ,再 び 減 圧 濃 縮 し て 凍 結 乾 燥 し た .YPD
培 地 及 びYNB
培 地 で 培 養 し て 得 ら れ た ガ ラ ク ト マ ン7
ナ ン を 以 下 そ れ ぞ れ
P-GM
及 びN-GM
と 略 す .5. 菌 体 か ら の オ ー ト ク レ ー ブ 粗 抽 出 物 の 調 製
菌 体 は ろ 紙 を 用 い て 分 離 し た 後 ,生 理 食 塩 水 で 洗 浄 し ,acetone を 加 え て 沈 積 し
4
℃ で 保 存 し た .acetone
脱 脂 し た 菌 体 を 濾 紙 濾 過 し , 再 度acetone
に 沈 積 し 濾 過 し た .こ れ を3
回 繰 り 返 し た 後 ,菌 体 を 室 温 で 乾 燥 さ せ 乳 鉢 中 で 粉 砕 し た . 粉 砕 し た 菌 体 に 精 製 水 を 加 え ,121
℃ ,3
時 間 , オ ー ト ク レ ー ブ に か け 熱 水 抽 出 を 行 っ た . 冷 後 ,3000 rpm, 20
分 間 遠 心 分 離 し 上 清 を2-3
日 流 水 透 析 し た 後 , 凍 結 乾 燥 を 行 っ た .6. Cetavlon
法 に よ る 粗 ガ ラ ク ト マ ン ナ ン の 分 離オ ー ト ク レ ー ブ 粗 抽 出 物 を
Cetavlon
法 に よ り 精 製 し た 6 5 ).粗 抽 出 物2 g
を 精 製 水40 mL
に 溶 か し ,9% Cetavlon (hexadecyl-trimethyl-ammo- nium bromide)
溶 液20 mL
を 加 え 室 温 で1
時 間 攪 拌 後 ,3000 rpm
,25
℃ で30
分 間 遠 心 分 離 を 行 っ た .得 ら れ た 上 清 に1% boric acid 40 mL
を 加 え 撹 拌 し な が ら1 M NaOH
溶 液 を 滴 下 しpH 9.5
に 調 整 し た . 生 じ た 沈 殿 を3000 rpm, 10
分 間 遠 心 分 離 で 回 収 し ,0.5% sodium acetate
溶 液(pH 9.5) 100 mL
で 洗 浄 後 ,2% acetic acid 2 mL
で 溶 か し ,0.8% sodium acetate/ethanol
溶 液 に 少 量 ず つ 加 え た .生 じ た 沈 殿 を3000 rpm
で10
分 間 遠 心 分 離 し , 沈 殿 を2% acetic acid/ethanol
溶 液 で 洗 浄 し 乾 燥 後 , 適 量 の 水 に 溶 か し2-3
日 流 水 透 析 し , 回 収 後 凍 結 乾 燥 を 行 っ た .7. O-結 合 型 糖 鎖 の 分 離
O-
結 合 型 糖 鎖 は2
種 の 条 件 でβ-elimination
に よ り 分 離 し た . 還 元 条 件 下 で の オ リ ゴ 糖 調 製 は , 粗 ガ ラ ク ト マ ン ナ ン300 mg
に0.5 M NaBH
4/0.1 M NaOH 30 mL
を 加 え , 室 温 で24
時 間 反 応 さ せ る こ と に よ り 遊 離 さ せ た 6 6 ).そ の 後acetic acid
で 中 和 し ,Amberlite IR-120 (H
+型)
で 脱 イ オ ン し , 生 成 物 をBio-Gel P-6 column( 2.5×100 cm) に か け た .
非 還 元 条 件 下 で の オ リ ゴ 糖 調 製 は , 粗 ガ ラ ク ト マ ン ナ ン300 mg
に0.1 M NaOH 30 mL
を 加 え 室 温 で24
時 間 反 応 さ せ る こ と に よ り 遊 離 さ せ た8
6 7 ). そ の 後
1 M HCl 3 mL
で 中 和 し ,Bio-Gel P-6 column( 2.5×100 cm)
に か け た . 両 溶 出 液 の 糖 含 量 を フ ェ ノ ー ル ・ 硫 酸 法 で 測 定 後 各 オ リ ゴ 糖 別 に 回 収 し ,更 に
Bio-Gel P-2 column( 2.5×100 cm)又 は Bio-Gel P-4 column( 2.5×200 cm) に か け , 同 様 に 発 色 後 そ れ ぞ れ の オ リ ゴ 糖 を 回
収 し 凍 結 乾 燥 を 行 う こ と に よ り 精 製 し た .溶 出 位 置 はCandida
マ ン ナ ン の ア セ ト リ シ ス に よ り 得 ら れ た2
糖 か ら6
糖 ま で の オ リ ゴ 糖 6 8 )を 標 準 品 と し て 用 い た .8. メ チ ル 化 分 析
メ チ ル 化 分 析 は 多 糖 , オ リ ゴ 糖 共 に
Ciucanu
ら の 方 法 6 9 )に 準 じ , 次 の よ う に 行 っ た . 試 料1 mg
にDMSO 1 mL
を 加 え ,1
時 間 攪 拌 し 試 料 を 溶 解 し た .次 に 多 糖 の 場 合 は 乳 鉢 で す り つ ぶ し た 粉 状 のNaOH 50 mg
を 添 加 し ,3
時 間 攪 拌 後CH
3I
を500 µL
加 え , 更 に3
時 間 撹 拌 し た . オ リ ゴ 糖 の 場 合 は 同 様 に 粉 状 のNaOH
を50 mg
添 加 し ,30
分 攪 拌 後CH
3I
を500 µL
加 え , 更 に30
分 撹 拌 し た . 反 応 液 に , 水1 mL
とchloroform 1 mL
を 加 え 激 し く 振 り 混 ぜ て 抽 出 後 ,1500 rpm,3
分 間 遠 心 分 離 を 行 いchloroform
層 を 回 収 し , 水 で3
回 洗 浄 後chloroform
層 に 無 水 硫 酸 ナ ト リ ウ ム を 適 量 加 え 脱 水 し メ チ ル 化 誘 導 体 を 得 た .続 い て ,得 ら れ た メ チ ル 化 誘 導 体 を
GC/MS
で 解 析 を 行 う た め に ,部 分 メ チ ル 化 ア ル ジ ト ー ル ア セ テ ー ト を 以 下 の よ う に 調 製 し た . ま ず2 M trifluoroacetic acid (TFA) 3 mL (TFA 450 µL
と 水2.55 mL
を 混 和 し て 調 製 し た)
を 加 え110
℃ で3
時 間 加 水 分 解 し , 濃 縮 乾 固 し た . そ の 後2-propanol
を2 mL
加 え40
℃ で 濃 縮 乾 固 を3
回 繰 り 返 し た .次 にNaBD
415 mg, 水 1 mL
を 加 え 一 晩 放 置 し 還 元 し た . そ の 後acetic acid 50 µL
で 中 和 し 濃 縮 乾 固 の 後 ,methanol
を2 mL
加 え40
℃ で 濃 縮 乾 固 を3
回 繰 り 返 し た .更 にacetic anhydride
とp yridine
の 混 合 溶 液(1
:1, v/v) 4 mL
を 加 え100
℃ で3
時 間 反 応 さ せ ア セ チ ル 化 を 行 っ た . そ の 後 減 圧 乾 固 し ,更 にtoluene
を2 mL
加 え50℃ で 濃 縮 乾 固 を 3
回 繰 り 返 し た .そ の 後chloroform 1 mL
で 抽 出 し ,水 で3
回 洗 浄 しchloroform
層 を 濃 縮 乾 固 後 ,acetone 5 mLに 溶 解 しGC/MS
で 解 析 し た .GC/MS の 条 件 は 以 下 の9
通 り で あ る .
カ ラ ム 名 :
DB-5
カ ラ ム サ イ ズ :
0.32 mm × 30 m He
ガ ス :20.4 mL/min
カ ラ ム 充 填 剤 : (5%-Phenyl)-methylpolysiloxane
昇 温 プ ロ グ ラ ム :100
℃(10 min), 20
℃/min (2.5 min)→5
℃/min (18 min)→20
℃/min (3 min)→300
℃9. HPLC
に よ るGalf
オ リ ゴ 糖 異 性 体 の 分 離得 ら れ た オ リ ゴ 糖 を 精 製 水
100 µL
に 溶 解 し ,50 µL
ず つTOSOH
製 高 速 液 体 ク ロ マ ト グ ラ フ 装 置(PX-8010)
を 用 い 分 離 し エ ル マ 光 学 製 示 差 屈 折 計 で ピ ー ク を 検 出 し , 各 ピ ー ク を 回 収 し た . 回 収 し た ピ ー ク を 再 度HPLC
で 分 離 し , 異 性 体 の ピ ー ク を 除 去 し て オ リ ゴ 糖 を 精 製 し た . 分 離 し た オ リ ゴ 糖 は 減 圧 濃 縮 後 凍 結 乾 燥 し た .HPLC
の 条 件 は 以 下 の 通 り で あ る .カ ラ ム 名 称 :
YMC-PACK PA-25
カ ラ ム サ イ ズ :10×500 mm
移 動 相 :
acetonitrile
:H
2O ( 63
:37 )
10
. 核 磁 気 共 鳴 ス ペ ク ト ル 分 析1
H-NMR
,1 3C-NMR
,total correlation spectroscopy (TOCSY), 2D nuclear Overhauser enhancement spectroscopy (NOESY)の 測 定 は 日 本 電 子 核 磁 気
共 鳴 装 置(JEOL JNM-LA600)
を 用 い て 行 っ た .1H-NMR
の 測 定 は 試 料1-10 mg
を 重 水(D
2O) 0.7 mL
に 溶 解 し ,acetone
を 内 部 標 準 物 質(
1H-NMR
は2.225 ppm
,1 3C-NMR
は31.07 ppm)
と し て 室 温 ,45
℃ 又 は70℃ で 行 っ た .
10
11
. 糖 量 測 定糖 量 は フ ェ ノ ー ル ・ 硫 酸 法 7 0 )に よ り 次 の よ う に 測 定 し た . 試 料 を 試 験 管 に
20
,50
又 は100 µL
ず つ 取 り ,5% phenol 100 µL, sulfuric acid 500 µL
を 加 え ,37℃ , 20
分 イ ン キ ュ ベ ー ト し ,TOSOH
製 マ イ ク ロ プ レ ー ト リ ー ダ ー(MPR-A4)
で492 nm
の 吸 光 度 を 測 定 し た .12
.α-mannosidase
処 理還 元 条 件 下 で 得 た 糖 ア ル コ ー ル 型 の 各 オ リ ゴ 糖
20 mg
を50 mM borate buffer (pH 4.5) 1 mL
に 溶 解 し ,Jack bean
由 来α-mannosidase
を5 unit
加 え ,37
℃ で24
時 間 イ ン キ ュ ベ ー ト し た 7 1 ). 次 に こ のα-mannosidase
処 理 成 績 体 混 合 物 をBio-Gel P-2 column (2.5×100 cm)
に か け て 分 画 し , そ れ ぞ れ の オ リ ゴ 糖 を 分 取 し た .11
第 三 節 結 果
O-
結 合 型 糖 鎖 に 及 ぼ す 培 地 の 影 響2
種 の 異 な る 培 地 で 培 養 し た 菌 体 か ら 培 地 中 に 遊 離 し た 細 胞 壁 ガ ラ ク ト マ ン ナ ン のO-
結 合 型 糖 鎖 に つ い て 差 異 を 検 討 す る た め に ,β-elimination
で 遊 離 し た オ リ ゴ 糖 をBio-Gel P-6 column
に か け て 多 糖 画 分 と オ リ ゴ 糖 画 分 に 大 き く 分 け て 分 取 し , そ の 後 オ リ ゴ 糖 画 分 を 再 度Bio-Gel P-2 column
も し く はBio-Gel P-4 column
に か け て 分 離 し た(Fig.
1-1).そ の 結 果 P-GM
は4
糖 ま で の オ リ ゴ 糖 し か 得 ら れ な か っ た の に 対 し ,N-GM
で は10
糖 前 後 ま で の オ リ ゴ 糖 が 得 ら れ た . こ の こ と か ら , 培 地 の 変 化 はO-結 合 型 糖 鎖 の Gal f
オ リ ゴ 糖 鎖 の 長 さ に 影 響 す る こ と が 明 ら か と な っ た .Fi g. 1 - 1. Co mp ar i s on o f t h e ch ain l en gt h o f O - li n ked ol i go s acch ar id es ob t ain ed fr o m A .
fu m ig at u s gr o wn u nd er d i s ti n ct cu l tu r e co nd i ti on s . C ul t ur es wer e gr o wn i n YP D o r Y N B med iu m;
gal act o man n an s wer e i s o l at ed b y D E AE - S eph ar o s e co l u mn ch ro mat o gr aph y; an d t h e r es ul t in g P - GM an d N - GM ( r es p ecti v el y) wer e s ub j ect ed to β-elimination under non-reducing conditions ( s ee M at er i al s and met h od s fo r fu l l d es cr ip t ion . ) Th e O- li n ked ol i go s acch ari d es r el eas ed fr o m P - GM wer e s ep ar at ed b y a col u mn (2 .5 × 10 0 c m) o f B io - Gel P -2 ( A) and fr o m N - GM b y a col u mn (2 .5 × 20 0 c m) o f B io - Gel P -4 ( B) . Th e carbo h yd r at e con t en t i n each fr act ion was d et er mi n ed b y t h e p h en ol / s ul fu r i c acid met ho d. Th e n u mb er s fr o m 1 to 6 ind i cat e th e elu t ion po s it io n o f mann o s e and manno ol i go s acch ari d es fr o m bio s e to h exao s e ob t ain ed f ro m C a nd id a man n an b y acet o l ys i s .
12
新 規
O-
結 合 型 糖 鎖 の 構 造 解 析こ れ ら
O-結 合 型 糖 鎖 の 構 造 を NMR
及 び メ チ ル 化 分 析 に よ り 解 析 し た .菌 体 抽 出 物 か ら 得 た ガ ラ ク ト マ ン ナ ン のO-
結 合 型 糖 鎖 は 培 養 上 清 か ら 回 収 し た ガ ラ ク ト マ ン ナ ン のO-
結 合 型 糖 鎖 と 同 じ 構 造 で あ っ た た . オ リ ゴ 糖 は 非 還 元 条 件 下 でβ-elimination
す る こ と に よ り 得 ら れ た 還 元 末 端 が ア ノ マ ー 異 性 体(α
体 及 びβ
体)
の 平 衡 状 態 の も の と ,還 元 条 件 下 でβ-elimination
す る こ と に よ り 得 ら れ た 還 元 末 端 糖 残 基 が 糖 ア ル コ ー ル 型 の も の の2
種 類 を 用 い て 解 析 し た . メ チ ル 化 分 析 の 結 果(
Table 1-1), 2
糖 はMan p α1→2Man p
で あ る こ と が 明 ら か と な り , ま た3
糖 以 上 で は 糖 ア ル コ ー ル 型 の オ リ ゴ 糖 の 還 元 末 端 が2,6-di-O-substituted mannitol (2,6-Man-ol)
と6-O -substituted mannitol (6-Man-ol)
の2
種 類 存 在 す る こ と が 明 ら か と な っ た . こ の2,6-Man-ol
残 基 の メ チ ル 化 で 得 ら れ た2,6-di- O-acetyl-1,3,4,5-tetra- O-methyl mannitol (2,6-Man-ol) (Fig. 1-2B)
は 非 還 元 末 端 のmannose
残 基 由 来 の1,5-di-O-acetyl-2,3,4,6-tetra- O-methyl mannitol (t-Manp) (Fig. 1-2A)
の180
° 回 転 対 称 体 で あ る た め ガ ス ク ロ マ ト グ ラ フ ィ ー で は 同 じ ピ ー ク と し て 検 出 さ れ 区 別 で き な か っ た . そ こ で 還 元 条 件 下 , 非 還 元 条 件 下 そ れ ぞ れ でβ-elimination
を 行 い 両 者 を 比 較 し た . す な わ ち 非 還 元 条 件 下 でβ-elimination
を 行 っ た 際 に 得 ら れ た オ リ ゴ 糖 に つ い てGC-MS
分 析 を 行 っ た と こ ろ , 非 還 元 末 端mannose
残 基(t-Man p)
か ら はm/z 102
,m/z 118,m/z 162
の フ ラ グ メ ン ト イ オ ン が 現 れ る(Fig. 1-2C)
の に 対 し , 還 元 条 件 下 で 得 た オ リ ゴ 糖 を 同 様 に メ チ ル 化 分 析 す る とm/z 101
とm/z 102, m/z 117
とm/z 118, m/z 161
の フ ラ グ メ ン ト イ オ ン が 強 く 現 れ(Fig.
1-2D), 2
種 の 部 分 メ チ ル 化 ア ル ジ ト ー ル ア セ テ ー ト が 確 認 で き た . 従 っ て 還 元 条 件 下 で 得 た オ リ ゴ 糖 の メ チ ル 化 物 中 に1,5-di-O-acetyl-2,3,4,6-tetra- O-methyl mannitol (t-Man p)
及 び2,6-di-O- acetyl-1,3,4,5-tetra-O-methyl mannitol (2,6-Man-ol)
が 含 ま れ て い る こ と , す な わ ち2,6-分 岐 マ ン ノ ー ス の 存 在 が 明 ら か と な っ た . 以 上 の 結 果 は
結 合 様 式 が 異 な る2
種 類 の オ リ ゴ 糖 の 存 在 を 示 し て お り , そ の 構 造 は3
糖 で は 分 岐 オ リ ゴ 糖 で あ るGal f β1→6(Man p α1→2)Man p
と 直 鎖 オ リ ゴ13
糖 の
Galf β1→5Gal f β1→6Man p
で あ る こ と が 明 ら か と な っ た .ま た4
糖 以 上 の 構 造 は3
糖 と 同 様 の2
種 類 の 基 本 構 造 の 末 端 にGalf
がβ-1,5
結 合 で 伸 長 し て お り , 長 い オ リ ゴ 糖 で はβ-1,6
結 合 の 存 在 も 示 し て い た(Table 1-1).
T abl e 1 -1. GC – M S an al ys i s o f O- met h yl al d i to l acet at es d er i ved fr o m met h yl at i o n an al ys es o f O - li n ked ol i go s acch ari d es
R el eas ed o l i go s acch ar id es b y β-elimination in the presence (+) or the absence (−) of NaBH4 wer e s ep ar at ed u s in g B io - Gel P - 2 col u mn an d el u at es co r r es pond i n g t o bi o s e fr act io n to p ent ao s e fr act i on wer e an al yzed b y met h yl at io n an al ys i s .
aR et en t io n t i me r el at i ve to th at o f 1, 5 -d i -O- acet yl - 2 ,3 , 4,6 - t et r a-O- met h yl man ni t o l.
O-Methylalditol acetate Sugar linkage RRTa
- + - + - + - +
2,3,4,6-Me4-Man t-Manp 1.00 1.00 1.00 2.60 - 3.00 - 1.40 -
3,4,6-Me3-Man 2-Manp 1.08 0.80 - - - -
1,3,4,5,6-Me5-Man 2-Man-ol 0.88 - 0.80 - - - -
2,3,4,6-Me4-Man t-Manp
+ + 1.00 - - - 6.00 - 4.10 - 2.50
1,3,4,5-Me4-Man 2,6-Man-ol
3,4-Me2-Man 2,6-Manp 1.20 - - 2.10 - 3.40 - 1.40 -
2,3,4-Me3-Man 6-Manp 1.11 - - 1.00 - 1.00 - 1.00 -
1,2,3,4,5-Me5-Man 6-Man-ol 0.90 - - - 1.00 - 1.00 - 1.00
2,3,5,6-Me4-Gal t-Galf 1.01 - - 3.40 2.60 3.20 1.70 2.40 1.50
2,3,6-Me3-Gal 5-Galf 1.09 - - 1.60 1.50 7.00 4.30 8.00 5.70
2,3,5-Me3-Gal 6-Galf 1.14 - - - - 0.10 0.10 0.50 0.30
Pentaose fraction
NaBH4 NaBH4 NaBH4
NaBH4
Biose fraction Triose fraction Tetraose fraction
14
Fi g. 1 - 2. M as s s p ect r a o f p ar ti al l y met h yl at ed al di t ol acet at e d er i ved fr o m O- l in ked ol i go s acch ar i d es. ( A) F r agmen t at io n s ch eme o f d eu t er iu m l ab el ed 1, 5- di -O- acet yl -2 ,3 ,4 ,6 - t etr a-O- met h yl man ni to l , w hi ch co rr es po nd s to a non - r edu cin g t er mi n al man no s e ( t -M anp).
( B ) F r agmen t at i on s ch e me o f r ever s ed 2, 6 -d i -O- acet yl -1 ,3 ,4 ,5 - t etr a-O- met h yl ma nni t ol , wh i ch cor r es po nd s t o 2, 6 -d i-O- s u bs t i tu t ed man n it ol (2 ,6 - M an- ol ) . ( C ) M as s fr ag men t io ns o f
d eu t eri u m- l ab el ed 1 ,5 - di -O- acet yl - 2 ,3 ,4 ,6 - t et r a-O- met h yl man n i to l ( t -M anp) as t h e s t and ar d.
( D ) M as s fr ag men t io n s o f a mi xt u r e o f d eut er iu m- l ab el ed 1 ,5 - di -O- acet yl - 2, 3, 4, 6- t et r a-O- met h yl mann i to l ( t -M anp) and 2, 6 -d i -O- acet yl - 1, 3, 4, 5 -t et r a-O- met h yl man n it ol (2 , 6- M an - ol ) obt ai n ed fr o m o l i go s acch ar id es lon ger th an b io s e fr act ion r el eas ed b y β-elimination under r ed u cin g cond i ti on s .
15
こ の 構 造 の 確 認 の た め に 1
H-NMR
解 析 を 行 っ た 結 果 , 還 元 末 端mannose (5.35-5.37 ppm),非 還 元 末 端 及 び 中 間 Gal f (5.19-5.22 ppm)
及 びmannose
に 結 合 す るGalf (5.02-5.05 ppm) の シ グ ナ ル が 得 ら れ た (Fig.
1-3A).還 元 条 件 下 の β-elimination
で 得 た オ リ ゴ 糖(2
糖-5
糖 を そ れ ぞ れO2-O5
と 略 す)
は 還 元 末 端 のmannose (Man-A)
の シ グ ナ ル が 消 失 し ,こ れ に 隣 接 す るα-1,2-
結 合mannose (Man-B)
の シ グ ナ ル の 高 磁 場 シ フ ト が 観 測 さ れ た(Fig. 1-3B)
. そ こ で 分 岐 オ リ ゴ 糖 のα-1,2
結 合mannose
を 除 去 し 構 造 解 析 を 進 め る た め に , こ れ ら の オ リ ゴ 糖 に つ い てα-mannosidase
処 理 を 行 い , 再 度Bio-Gel P-2 column
に か けα-mannosidase
抵 抗 性 の 直 鎖 オ リ ゴ 糖(n)
と 分 岐 オ リ ゴ 糖 由 来 のα-mannosidase
分 解 成 績 体 で あ る 直 鎖 オ リ ゴ 糖(n-1)
を 分 離 し(Fig.
1-3E), そ れ ぞ れ に つ い て
1H-NMR
解 析 を 行 っ た(Fig. 1-3C,1-3D). そ
の 結 果 , オ リ ゴ 糖 異 性 体 混 合 物 中 の 直 鎖 オ リ ゴ 糖 と 分 岐 オ リ ゴ 糖 由 来 の オ リ ゴ 糖 が 区 別 で き ,主 鎖 のmannose
に 結 合 す る 非 還 元 末 端Gal f
は5.045 ppm, mannose
に 結 合 す るGalf
は5.022 ppm, 1,5
結 合 中 間Galf
は5.195 ppm,非 還 元 末 端 Gal f
は5.220 ppm
で あ る こ と が 明 ら か と な っ た . ま た ,mannose
にGalf
が 直 鎖 状 に 繋 が る も の と ,mannose
にGalf
とmannose
が 分 岐 状 に 繋 が る も の の 存 在 は3
糖 のα-mannosidase
抵 抗 性 直 鎖 オ リ ゴ 糖(O3-3)と 4
糖 の 分 岐 オ リ ゴ 糖 由 来 のα-mannosidase
分 解 成 績 体 で あ る 直 鎖 オ リ ゴ 糖 の3
糖(O4-3)
の 1H-NMR
シ グ ナ ル が 一 致 す る こ と ,さ ら に4
糖 のα-mannosidase
抵 抗 性 直 鎖 オ リ ゴ 糖(O4-4)と 5
糖 の 分 岐 オ リ ゴ 糖 由 来 のα-mannosidase
分 解 成 績 体 で あ る 直 鎖 オ リ ゴ 糖 の4
糖(O5-4)の
1H-NMR
シ グ ナ ル が 一 致 す る こ と か ら 確 認 で き た . ま た こ れ ら のO-結 合 型 糖 鎖 の
1H-NMR
の 化 学 シ フ ト 値 をTable 1-2
に 示 し た .16
Fi g. 1 - 3. 1H N M R s p ect r a o f O - l in ked o li go s acch ar id es . ( A) O l i go s acch ari d es ob t ain ed b y β-elimination under non-reducing conditions. (B) Oligosaccharides obtained by β-elimination und er r edu ci n g co nd it io n s . Th e b io s e fr act i on to p ent aos e fr act i on wer e d es i gn at ed as O2 to O5 , r esp ect i vel y. O3 to O5 wer e t r eat ed wi th α-mannosidase and downsized (n-1) oligosaccharides ( C ) and α-mannosidase resistant (n) oligosaccharides (D) were separated by Bio-Gel P-2 column chr o mat o gr aph y. (E ) Th e s ep ar at io n s ch eme o f α-mannosidase degradation products. For the s t ru ctu r e in t hi s fi gu r e, s ymb o l s
○
and□
in di cat e gal acto fu r ano s e ( Galf) and man no p yr an o s e (M an ) r es i du es , r es p ect i vel y. O 3 -3 an d O 3 -2 i nd i cat e α-mannosidase resistant triose and its d egr ad at io n pr od u ct , b io s e, r es p ect i vel y. O 4 -3 an d O 5 - 4 ind i cat e t ri o s e and t etr ao s e o bt ai n ed fr o m O 4 an d O 5, r es p ecti ve l y, b y α-mannosidase degradation.17
T abl e 1 -2 . 1H NM R ch e mi c al s h i ft s (δ) of O-linked oligosaccharides Sugar residue and structure
F E D C A F E D C A
B B
Biose fraction Manpα - 5.366
↑2
Manpα1 5.046
Mannitol + -
↑2
Manpα1 5.003
Triose fraction Galfβ1→6Manpα - 5.046 5.352
↑2
Manpα1 5.056
Galfβ1→6Mannitol + 5.05 -
↑2
Manpα1 5.003
Galfβ1→5Galfβ1→6Manpα - 5.192 5.028 5.056
Galfβ1→5Galfβ1→6Mannitol + 5.22 5.023 -
Tetraose fraction Galfβ1→5Galfβ1→6Manpα - 5.222 5.019 5.352
↑2
Manpα1 5.053
Galfβ1→5Galfβ1→6Mannitol + 5.222 5.025 -
↑2
Manpα1 5.002
Galfβ1→5Galfβ1→5Galfβ1→6Manpα - 5.222 5.194 5.019 5.053
Galfβ1→5Galfβ1→5Galfβ1→6Mannitol + 5.222 5.195 5.025 -
Pentaose fraction Galfβ1→5Galfβ1→5Galfβ1→6Manpα - 5.221 5.195 5.024 5.352
↑2
Manpα1 5.048
Galfβ1→5Galfβ1→5Galfβ1→6Mannitol + 5.221 5.197 5.025 -
↑2
Manpα1 5.003
Galfβ1→5Galfβ1→5Galfβ1→5Galfβ1→6Manpα - 5.221 5.195 5.195 5.024 5.048 Galfβ1→5Galfβ1→5Galfβ1→5Galfβ1→6Mannitol + 5.221 5.197 5.197 5.025 -
Galfβ1→6Galfβ1→5Galfβ1→5Galfβ1→6Manpα - 5.048 5.23 5.195 5.024 5.048 Galfβ1→6Galfβ1→5Galfβ1→5Galfβ1→6Mannitol + 5.048 5.232 5.197 5.025 -
Oligosaccharides Reduction
by NaBH4
H-1 Chemical shift, δ (ppm)
18
O-
結 合 型 糖 鎖 の 全 体 構 造こ れ ら の 結 果 か ら
2
種 の 異 な る 培 地 で 培 養 し 産 生 さ れ る ガ ラ ク ト マ ン ナ ン のO-結 合 型 糖 鎖 の 構 造 を Fig. 1-4
に 示 す .P-GM
か ら 得 たO-
結 合 型 糖 鎖 は 直 鎖 状 及 び 分 岐 状 の も の が4
糖 ま で し か 存 在 し て い な い の に 対 し ,N-GM か ら 得 た も の で はGal f
鎖 が 長 く ,10 糖 前 後 ま で の オ リ ゴ 糖 の 存 在 し て い る こ と が 明 ら か と な っ た .Fi g. 1 - 4. P r opo s ed st r u ct u r es fo r P - GM ( A) and N- GM ( B ) . E ach o f th e s t ru ctu r es i s o n e o f th e po s s ib il i ti es ou t o f th e s t ati s ti cal en s emb l e. M an and G alf d eno t e D - man no p yr an o s e an d D - gal act o fu r ano s e r es id u es , r es p ect i vel y. Th e s i d e ch ai n s equ en ce i s n ot sp eci fi ed .
19
第 四 節
考 察
通 常 ,
O-
結 合 型 糖 鎖 の 解 析 の 際 にβ-elimination
を 行 う 場 合 に は ,NaBH
4下 で の 還 元 を 行 い な が ら オ リ ゴ 糖 を 得 る .こ れ は ,β-elimination
の 際 に 遊 離 す る オ リ ゴ 糖 の 還 元 末 端 が 異 性 化 も し く は ピ ー リ ン グ 反 応 を 起 こ す 恐 れ が あ る た め で あ る . ま た 還 元 末 端 を 糖 ア ル コ ー ル 型 と す る こ と で 異 性 体 の 存 在 が な く な る た め ,1H-NMR
シ グ ナ ル が 単 純 と な り 解 析 し や す い と い う 利 点 も あ る . 一 方 で , フ ェ ノ ー ル ・ 硫 酸 法 に よ る 発 色 の 際 に 通 常 単 糖 と し て 現 れ る 糖 が 糖 ア ル コ ー ル と な る た め 検 出 で き な い こ と や , メ チ ル 化 分 析 を 行 っ た 際 に 還 元 末 端 の 糖 の ガ ス ク ロ マ ト グ ラ フ の ピ ー ク が 糖 ア ル コ ー ル の 場 合 異 な る 位 置 に 現 れ て し ま い 解 析 が 複 雑 に な る 場 合 が あ る . そ こ で 今 回 は 還 元 条 件 下 及 び 非 還 元 条 件 下 でβ-elimination
を 行 い , ガ ス ク ロ マ ト グ ラ フ ィ ー でretention time
が 同 じ に な る2
種 類 の オ リ ゴ 糖 を 区 別 し ,そ れ ぞ れ メ チ ル 化 分 析 ,NMR
解 析 を 行 う こ と で 詳 細 な 解 析 を 行 っ た .こ れ ま で に も
A. fumigatus
のO-
結 合 型 糖 鎖 の 構 造 は 報 告 さ れ て い るが 5 8 ), 今 回 同 様 の 構 造 が 確 認 で き た の は
5
糖 の み で あ り , 新 た に 分 岐状 の
Gal f β1→6(Man p α1→2)Man
と 直 鎖 状 のGal f β1→5Gal f β1→6Man
及 び 長 いGal f
鎖 の 中 にβ-1,6
結 合 を 有 し て い る 糖 鎖 の 存 在 を 明 ら か に し た . 特 にmannose
とGal f
の 分 岐 オ リ ゴ 糖 は 今 ま で 報 告 さ れ て い るO-
結 合 型 糖 鎖 の 中 で も 珍 し く ,こ れ ま で の 報 告 で はFusarium
属 菌 に 関 す る 報 告5 4 )の み で あ る . 今 回
A. fumigatus
に 新 た な 構 造 のO-
結 合 型 糖 鎖 を 発 見し た が ,
A. niger
で は 糖 の 種 類 や 結 合 様 式 が 異 な る5
種 類 のO-
結 合 型 糖 鎖 の 存 在 が 報 告 さ れ て い る た め 7 2 ), 菌 種 , 菌 株 に よ り 存 在 す る オ リ ゴ 糖 の 構 造 が 僅 か に 異 な っ て い る 可 能 性 が 考 え ら れ る .ま たO-
結 合 型 糖 鎖 の 役 割 と し て 細 胞 壁 の 形 成 や 維 持 へ の 関 与 が 報 告 さ れ て い る が 5 4 ,7 3 ), こ れ ら の 構 造 の 違 い が 与 え る 影 響 に つ い て は 未 解 明 で あ り 今 後 の
研 究 課 題 と な っ て い る .
本 研 究 に よ り 培 地 成 分 の 違 い が ,O-結 合 型 糖 鎖 の
Galf
鎖 の 長 さ に 影 響 を 及 ぼ す こ と が 明 ら か と な っ た .Galf
鎖 の 生 合 成 に 関 与 す る 遺 伝 子20
と し て は ,近 年
galactofuranosyltransferase (GfsA)
が 報 告 さ れ て い る7 4 ). 今 回 解 析 し たGal f
鎖 はβ-1,5
結 合 及 びβ-1,6
結 合 が 共 存 し た 構 造 と な っ て い た が ,GfsA
に よ り 合 成 さ れ るGalf
鎖 の 結 合 様 式 に つ い て は 未 だ 明 ら か と な っ て い な い .Mycobacterium tuberculosis
が 有 す るUDP-galacto- furanosyl transferase (UGT)
はβ-1,5
結 合Galf
及 びβ-1,6
結 合Galf
両 転 移 酵 素 活 性 を 有 す る た め 7 5 ),GfsA
も 同 様 の 活 性 を 有 す る 可 能 性 が あ り 今 後 検 討 す る 必 要 が あ る .Leitao
ら 5 8 )はA. fumigatus
のO-
結 合 型 オ リ ゴ 糖 に 抗 原 活 性 が あ る こ と を 報 告 し て い る が ,今 回 解 析 し た 構 造 に もGal f
鎖 が 存 在 し て い る こ と か ら 同 様 に 抗 原 活 性 を 有 す る と 考 え ら れ る .し か しN-
結 合 型 ガ ラ ク ト マ ン ナ ン と 比 較 す る と 糖 鎖 の 長 さ が 短 い こ と か ら ,O-
結 合 型 糖 鎖 はN-結 合 型 ガ ラ ク ト マ ン ナ ン に 覆 わ れ て 存 在 し て い る 可 能 性 が あ る が そ
の 立 体 的 位 置 関 係 は 未 だ 明 ら か に な っ て い な い . そ の た め ア ス ペ ル ギ ル ス 症 の 診 断 用 モ ノ ク ロ ー ナ ル 抗 体 のO-
結 合 型 オ リ ゴ 糖 と の 反 応 性 の 程 度 は 不 明 で あ り , 今 後 更 な る 解 析 が 求 め ら れ る .21
第 二 章
異 な る 培 地 で 培 養 し た
Aspergillus fumigatus
のN-
結 合 型 ガ ラ ク ト マ ン ナ ン の 構 造 解 析第 一 節 緒 論
真 菌 類 や 細 菌 類 は 細 胞 壁 を 有 し て い る が , 特 に 酵 母 は 細 胞 壁 に 特 徴 的 な
β-
グ ル カ ン と マ ン ナ ン の 存 在 が よ く 知 ら れ て い る . そ の 構 造 は 属 や 種 に よ っ て 異 な っ て お り , 当 研 究 室 で はC. albicans
,Candida tropicalis, Candida glabrata
な ど の 細 胞 壁 マ ン ナ ン 構 造 の 解 析 が 進 め ら れ て き た 6 8 , 7 6 - 8 3 ). こ れ ら の 構 造 は 代 表 的 な 酵 母 で あ るS. cerevisiae
と 同 様 の 櫛 型 構 造 で あ る . ヒ ト は こ れ ら の 糖 鎖 を 有 し て い な い た め , 抗 体 を 用 い た 感 染 症 診 断 の た め の 抗 原 と し て 利 用 し て き た . ま た グ ル カ ン に 対 し て はβ-1,3-
グ ル カ ン 合 成 酵 素 を タ ー ゲ ッ ト と し , 副 作 用 の 少 な い 薬 が 開 発 さ れ 用 い ら れ て い る .一 方 で
A. fumigatus
の ガ ラ ク ト マ ン ナ ン の 構 造 は こ れ ま で に も 解 析 さ れ て い る が ,部 分 的 に 異 な っ た 解 析 結 果 が 報 告 さ れ て い る 8 4 -8 8 ).そ の 原 因 と し て , 解 析 に 用 い た 菌 種 が 異 な る こ と や , 異 な る 培 養 条 件 で 培 養 を 行 っ て い る こ と が 考 え ら れ る .Latgé
ら 8 8 )は ガ ラ ク ト マ ン ナ ン の 構 造 をα-1,2
結 合 のmannotetraose
がα-1,6
結 合 で 繋 が っ た 直 鎖 状 の 繰 り 返 し 構 造 を し て い る コ ア マ ン ナ ン に , 側 鎖 と し て4-5
残 基 か ら な るβ-1,5
結 合Galf
オ リ ゴ 糖 側 鎖 がmannose
のC-3
位 又 はC-6
位 に 結 合 し た 構 造 で あ る と 報 告 し て い る(Fig. 2-1).一 方 Van Bruggen-Van Der
Lugt
ら 8 9 )は ガ ラ ク ト マ ン ナ ン の 構 造 の 中 にβ-1,6
結 合 が 存 在 し て い るこ と を 示 し て お り , こ れ は
Latgé
ら 8 8 )の 報 告 に は 含 ま れ て い な い 結 合 様 式 で あ っ た .Latgé
ら 8 8 )はA. fumigatus
の 培 養 の 際 にYPD
培 地 を 用 い て い た が ,Van Bruggen-Van Der Lugt
ら 8 9 )はYNB
培 地 を 使 用 し て い た .YNB 培 地 は 酵 母 や 真 菌 を 培 養 す る た め の 代 表 的 な 培 地 の1
つ で あ り , 窒 素 源 と し て(NH
4)
2SO
4を 含 み ,pH
は 酵 母 や 真 菌 の 成 長 の た め22