熊本大学学術リポジトリ
Kumamoto University Repository System
Title 口腔扁平上皮癌における癌幹細胞関連分子Nucleostemin,
Notch1の発現と機能解析
Author(s) 吉田, 遼司
Citation
Issue date 2012-03-23
Type Thesis or Dissertation
URL http://hdl.handle.net/2298/25190
学位論文
Doctoral Thesis
口腔扁平上皮癌における癌幹細胞関連分子
Nucleostemin, Notch1
の発現と機能解析
(Investigation of the expression of cancer stem cell related molecules, Nucleostemin and Notch1, and their roles in oral squamous cell carcinoma)
吉 田 遼 司
Ryoji Yoshida
熊本大学大学院医学教育部博士課程医学専攻顎口腔病態学
指導教員
篠原 正徳 教授
熊本大学大学院医学教育部博士課程医学専攻顎口腔病態学伊藤 隆明 教授
熊本大学大学院医学教育部博士課程医学専攻機能病理学学
位
論
文
Doctoral Thesis
論文題名 : 口腔扁平上皮癌における癌幹細胞関連分子 Nucleostemin, Notch1 の発現と機能解析 (Investigation of the expression of cancer stem cell related molecules, Nucleostemin
and Notch1, and their roles in oral squamous cell carcinoma)
著 者 名 : 吉 田 遼 司 Ryoji Yoshida 指導教員名 : 熊本大学大学院医学教育部博士課程医学専攻顎口腔病態学 篠原 正徳 教授 熊本大学大学院医学教育部博士課程医学専攻機能病理学 伊藤 隆明 教授 審査委員名 : 頭頸部感覚病態学担当教授 湯本 英二 脳神経外科学担当教授 倉津 純一 呼吸器外科学担当教授 鈴木 実 損傷修復学担当講師 立石 智 2012年3月
1. 要旨
【目的】近年、様々な腫瘍で癌幹細胞 (cancer stem cell; CSC) の存在やそのマー カー、および制御に関わる多くの CSC 関連分子が報告されつつある。しかし、 口腔扁平上皮癌 (OSCC)において、病理組織標本を用いてそれらの発現様式の変 化を観察、あるいは機能解析を行なった報告は少ない。本研究では OSCC の発 生過程における癌幹細胞関連分子 Nucleostemin および Notch1 の発現様式を明ら かにするとともに、in vitro の実験からこれらの分子の OSCC における機能につ いて検討した。
【方法】種々の口腔扁平上皮組織標本を用いて Nucleostemin と Ki-67 の免疫染色 を行った。次に OSCC 培養細胞株における Nucleostemin の発現を確認し、OSCC
培養細胞株 SAS 細胞に siRNA を導入して機能解析を行なった。また、Ca2+依存
的な培養細胞の角化誘導実験を行い、OSCC 培養細胞の分化と Nucleostemin の関 係を検討した。Notch1 については、同様な組織標本を用いて Notch1 とそのリガ ンドの Jagged1(JAG1)の免疫染色を行った。また、ラット舌発癌モデルを用い、 Notch1 の発現変化を観察した。OSCC 培養細胞株における Notch1 の発現を確認 し、SAS 細胞に Notch signaling 活性化の阻害剤を添加、あるいは siRNA を導入 して機能解析を行なった。さらに、Notch1 と Laminin5 2 chain (L52)の免疫染色 を行い、OSCC 培養細胞の遊走能、浸潤能と Notch signaling との関連について組 織および in vitro で検討した。
【結果】Nucleostemin は組織標本上で様々な分化段階の細胞に発現していたが、 その陽性率は病変の進行とともに増加し、Ki-67 陽性率と正の相関関係を認めた。 OSCC 培養細胞では全ての細胞で Nucleostemin の発現が認められ、siRNA 導入 によって癌細胞の増殖活性は顕著に抑制された。また、角化誘導実験では OSCC 培養細胞の分化と Nucleostemin の発現量に相関関係は認めなかった。Notch1 と
JAG1 は扁平上皮癌で高頻度に発現し、OSCC 培養細胞でも Notch1 やその関連分
子が発現していた。Notch signaling を阻害すると細胞増殖活性が抑制された。 L52 と Notch1 には相関関係があり、in vitro における機能解析で TNF-依存的 な癌細胞の浸潤能は Notch signaling の阻害によって減弱した。
【考察】Nucleostemin は癌細胞の増殖活性維持に重要な役割を果たしていること が示唆された。しかし、角化誘導実験や正常組織における発現プロファイルか
向にあった。in vitro の実験結果から、Notch1 は腫瘍細胞の増殖活性維持や遊走、 浸潤能の制御を介して OSCC の維持に関与する可能性が考えられた。 【結論】癌の成り立ちを考えると、単一分子の制御のみで CSC を含め癌細胞の コントロールが可能であるとは考えにくい。種々の CSC 関連分子の研究が進ん でいる現在、これらの関連分子への様々な角度からの知見が統合されることで OSCC の研究が進展し、治療へのアプローチが発展すると考えられた。
2. 発表論文リスト ①関連論文
Ryoji Yoshida, Tetsuhiro Fujimoto, Shinji Kudoh, Masashi Nagata, Hideki Nakayama, Masanori Shinohara and Takaaki Ito.
Nucleostemin affects the proliferation but not differentiation of oral squamous cell carcinoma cells.
Cancer Sci. 102 (7): 1418-23, 2011.
②その他の論文
Nagata M, Nakayama H, Tanaka T, Yoshida R, Yoshitake Y, Fukuma D, Kawahara K, Nakagawa Y, Ota K, Hiraki A, Shinohara M.
Overexpression of cIAP2 contributes to 5-FU resistance and a poor prognosis in oral squamous cell carcinoma.
Br J Cancer. 105(9):1322-30, 2011.
Tanaka, Takuya; Nakayama, Hideki; Yoshitake, Yoshihiro; Irie, Atsushi; Nagata, Masashi; Kawahara, Kenta; Takamune, Yasuo; Yoshida, Ryoji; Nakagawa, Yoshihiro; Ogi, Hidenao; Shinriki, Satoru; Ota, Kazutoshi; Hiraki, Akimitsu; Ikebe, Tetsuro; Nishimura, Yasuharu; Shinohara, Masanori Selective inhibition of NF-κB by NBD peptide suppresses the metastasis of highly metastatic oral squamous cell carcinoma
Cancer Sci. (in press)
吉田遼司, 中山秀樹,永田将士,吉武義泰,手島慶子,篠原正徳 歯性感染症が原因で生じたと考えられた敗血症性肺塞栓症の 1 例 日本口腔外科学会雑誌, 56 巻 1 号: 49-53, 2010.
3. 謝辞 本研究は熊本大学大学院生命科学研究部 総合医薬科学部門 感覚・運動医 学講座 顎口腔病態学分野、篠原 正徳 教授、および同生体情報分析医学講 座 機能病理学分野、伊藤 隆明 教授の御指導のもとに行いました。研究に おいては勿論、多面にわたり御指導をいただき、深く感謝いたします。 研究手法や研究に対する姿勢を御教授いただきました、機能病理学分野助教、 新森 加納子 先生、同前助教、藤本 哲広 先生ならびに宇高 直子 先生 に厚く感謝の意を表します。 本研究で用いました OSCC 細胞株 HSC-2 細胞、HSC-3 細胞、SAS 細胞、KB 細胞、 Ca9-22 細胞、SCC-KN 細胞は九州大学大学院 歯学研究院 口腔顎顔面病態学分 野より、OSC-20 細胞、OSC-19 細胞、HOC-313 細胞は金沢大学大学院 がん医科 学専攻 がん細胞学講座 細胞浸潤学分野より、また、ラットの口腔癌モデル については、東海細胞研究所の田中 卓二 先生より御供与頂いたものであり、 深く感謝申し上げます。 さらに、顎口腔病態学分野および機能病理学分野の研究室の皆様には常日頃 より有形無形の多大なる御助言、御協力頂き誠にありがとうございました。皆 様に心より感謝いたします。
4. 略語一覧 CK19:cytokeratin19 CSL:CBF1/Su(H)/Lag-1 CSC:癌幹細胞 DAPI:4',6-diamidino-2-phenylindole DLL:Delta like
DMENM:Dulbecco’s modified Eagles medium DMSO:Dimethyl sulfoxide
EMT:上皮間葉転換epithelial-to-mesenchymal transition
FBS:Fetal bovine serum HE:hematoxylin & eosin Invo:Involucrin JAG1:Jagged1 LI:labelling index L52:Laminin5 2 chain MTS assay: 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophen yl)-2H-tetrazolium, inner salt assay
NECD:細胞外ドメイン
NICD:Notch intracellular domain p-H3:phospho histone H3
RT-PCR:reverse transcriptase-polymerase chain reaction
siRNA:small interfering RNA
TACE:ADAM メタロプロテアーゼ TNF-α変換酵素 TM:膜貫通ドメイン
TSC:Tissue stem cell TSCC:舌 OSCC
WHO:World Health Organization 4NQO:4-nitroquinoline 1-oxide
5. 研究の背景と目的 5-1. 口腔扁平上皮癌(OSCC) 5-1-1. OSCC の概要 口腔癌は種々の悪性腫瘍の中でも代表的な疾患のひとつで、舌や頬粘膜など の口腔領域に発生する(1)。なかでも OSCC は口腔の被覆粘膜である重層扁平上皮 にその起源を持ち、口腔癌の約 80%以上を占めている。発生部位としては舌が最 も多く、約 45%を占め、ついで歯肉 30%、口底 10%、頬粘膜 8%、口蓋 3%と続く(2)。 治癒率は発生部位によりわずかに異なるが、全 OSCC 患者の 5 年累積生存率は概 ね 60~70%である。しかし、病期別に見ると、StageⅠ85~95%、StageⅡ80~95%、 StageⅢ60~75%、StageⅣ40~60%となり癌の進行に伴って予後は急速に悪化す る。進行癌においては、救済された患者でも程度の差こそあれ、摂食、嚥下、 および構音など日常生活の根本に関わる口腔機能および審美に影響を残す。そ のため、生命予後、QOL の両面から、早期発見・早期治療の重要性が示唆される。 近年、OSCC の診断は MRI や PET-CT などの新たな画像診断機器の開発や技術の向 上により、確実に進歩してきた。治療においても、再建技術の進歩に伴う手術 範囲の拡大や新規抗癌剤の認可により治療法の選択肢は多岐にわたるようにな った。しかし、これらの進歩にもかかわらず、OSCC を含む頭頸部癌患者の生存 率の劇的な向上には至っていない(3)。その要因として難治性癌の存在、すなわち 抗癌剤耐性、放射線耐性、あるいは高転移・浸潤能を獲得した癌細胞の存在が 想定されている。 これらの問題を解決するためには、上述のような悪性度の高い癌細胞の形成 に関わる機構の解明が重要で、そのことが新たな治療法の開発や予後予測因子 の発見に繋がっていくと期待される。 5-1-2. OSCC の発生過程 OSCC にはde novoに発生するものと、多段階的に発生し前癌病変としての上 皮異形成、上皮内癌を経て浸潤癌へと進展するものがある(4)。上皮異形成につい ては、世界保健機構(WHO)によって細胞異型および組織異型の程度より上皮異 形成の病理学的基準が定義されており(5)、上皮異形成の発生には細胞増殖と上皮 組織の成熟に関わる機構の異常が関連しているとされる(6, 7)。また、その異常は 浸潤癌へと移行した OSCC の増殖・分化にも深く関与していると考えられる。 以上より、OSCC 発生過程における増殖・分化機構破綻の分子機構を解明しそ
の理解を深めることは、新たな治療方法の開発や予後予測に寄与するものと考 えられる。
5-1-3.癌幹細胞(CSC)と研究の現状
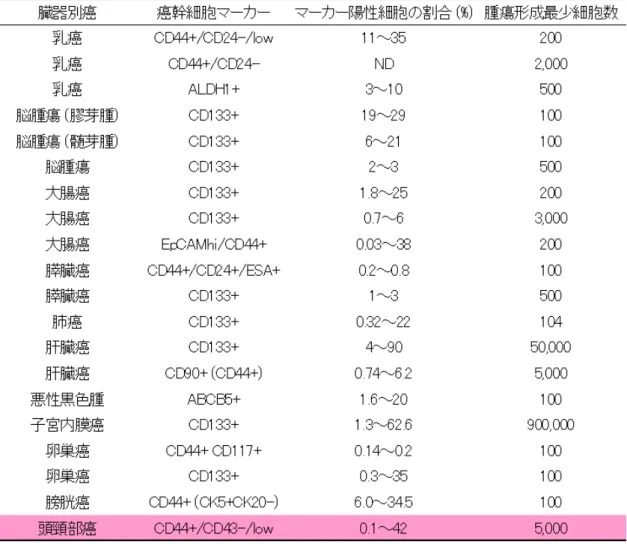
正常組織中には組織幹細胞(Tissue stem cell;TSC)が存在し、それらを頂 点とする階層的な細胞社会が構成され、それらに基づいて器官形成・組織修復・ 恒常性維持が行われている。1960 年代の Till、McCulloch、Becker らによる血 液幹細胞の同定などを皮切りに(8)、各種組織で活発な増殖能、多分化能(複数の 異なった機能を持つ成熟細胞へ分化する能力)、自己複製能(不均等分裂により 自己と同じ幹細胞を維持する能力)を兼ね備えた組織幹細胞の存在が明らかに なりつつある。 CSC の概念は、これら正常幹細胞に関する知見に加え、①癌組織が遺伝子に変 異を来たした異常増殖をする単一の細胞集団であるという概念では再発や転移 を説明することに限界があるのではないか、②癌組織の一部には癌組織を再構 築する能力に非常に長けた性質を持つ細胞集団が存在するのではないか、とい う考え方に端を発する概念で、癌研究の比較的初期から提示されてきた(9)。Dick らによって細胞表面マーカーによる分離技術と動物移植手法の確立によって白 血病の CSC 集団の特定が報告されると、「CSC」は癌根治のためのパラダイムシ フトをもたらす有益な研究対象であるとして認識されはじめ、その後の固形癌 における CSC の発見も相まって急速な研究展開がみられるようになった。 固形腫瘍における CSC の特定は乳癌、脳腫瘍、大腸癌、膵臓癌などをはじめ 頭頸部癌でも進んでおり、種々のマーカーを応用して CSC 分画の同定が進んで いる(表 1)。また、現在はその性状解析へと研究の対象は移行しているが、OSCC の発癌過程における CSC 関連分子の発現の変化やその分子が果たす役割につい ての検討はあまり多くない。
表 1. 各癌腫における代表的 CSC マーカー 以上より、OSCC において CSC 関連分子の発現解析や機能解析を行なうことは、 OSCC の CSC ひいてはその分化・増殖機構の理解に繋がり治療や予後予測にも寄 与しうると考えられる。そこで、本研究においては近年新たに同定された幹細 胞関連分子 Nucleostmein と従来から幹細胞に関わりが深いとされている Notch signaling に焦点をあてた。各々の分子の病理組織標本や培養細胞での発現解析 を行なうと共に、それらの分子が OSCC において果たす役割についての検討を行 なった。
5-2. Nucleostemin
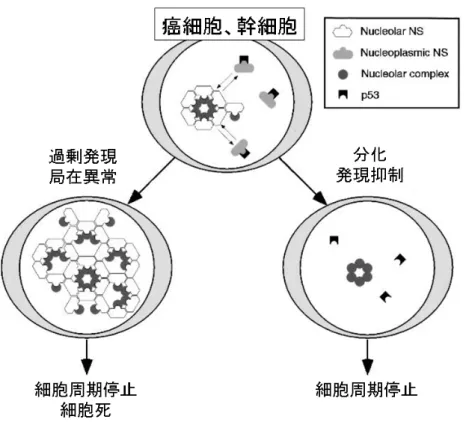
Nucleostemin は腫瘍抑制タンパク質である p53 と結合し、細胞増殖を調節す る新規の核小体蛋白質として Tsai 博士らによって同定された分子である。NS は 最初に成獣ラットの脳幹細胞で特定され、続いて骨髄や胚由来の幹細胞や数種 の変異細胞株、腫瘍細胞で発現していることが見出された。Caenorhabditis elegans,、Drosophila melanogaster、Xenopus laevis、zebrafish、mouse、rat、 human などの遺伝子にコードされており、アミノ酸配列は種間で良く保存されて いる。核小体局在には GTP との結合が必要であり、GTP binding domain がこの 分子の局在に重要である。また種々の Nucleostemin 結合タンパク質が同定され ており、なかでも p53、MDM2、Nucleophosmin、TRF1 などの分子は細胞周期調節 に関わっている分子であり注目されている(図 1、図 2)(10)。その後いくつかのグ ループから Nucleostemin が種々の幹細胞や癌細胞の細胞株でも発現しているこ とが報告されている(11-13)。 図 1 Nucleostemin の基本的構造
図 2 癌細胞、幹細胞における Nucleostemin の機能 Nucleostemin は一般的に増殖能の高い前駆細胞が細胞周期を脱し、ある分化 段階に進入する際に その発現が抑制される(10, 14, 15)。 また、Nucleostemin の発 現を抑制することで幹細胞や癌細胞の増殖が著明に阻害されることがわかって いる(12, 16-21)。これら初期の知見は Nucleostemin が幹細胞の多分化能や癌あるい は幹細胞の増殖の調節に関与している可能性を示唆しうるものであった。 CSC においても Nucleostemin は重要な機能を果たしていることが知られてい る。多形神経膠芽腫においては、Nucleostemin の存在下で特異的に応答するプ ロモーターを搭載した GFP 発現ベクターを応用したマウスモデルで Nucleostemin 強発現分画にある腫瘍細胞で腫瘍形成能や浸潤能が高いことが分 かっており、CSC 分画と非常に近似している細胞集団と Nucleostemin 発現量と の密接な関係が示唆されている(22)。乳癌においても悪性度が高い組織型で Nucleostemin が高発現しており、MMTV-Wnt1 や MMTV-PyMT マウス乳癌モデルで は悪性度と Nucleostemin の発現に相関が認められ、かつ Nucleostemin が高発 現している乳癌細胞は非常に高い造腫瘍能や幹細胞関連分子の発現を認め、そ の形質は Nucleostemin 依存的であったとする報告がある(23)。また Hela 細胞を 用いた研究では、Nucleostemin やその類縁分子である GNL3L を過剰発現させた
場合に多能性幹細胞マーカーや腫瘍形成能が上昇し、放射線感耐性を獲得する ことが報告され、その形質変化から Nucleostemin の高発現腫瘍細胞と CSC との 強い関連が示唆されている(24)。 しかしながら、最近の研究で、Nucleostemin が実際には正常な増殖能を有す る細胞にも腫瘍細胞と同じレベルで広範囲に発現していることが分かってきて いる(25-27)。一方で、Nucleostemin が口腔扁平上皮およびそれに由来する OSCC に おいてどのような発現パターンを示し、どのように OSCC の発生に関わるのかに ついてはあまりよく分かっていない。 そこで本研究において、OSCC 発生における Nucleostemin の重要性を明らかに するために、舌の正常上皮、前癌病変(上皮異形成)そして浸潤癌の組織切片 上での Nucleostemin の発現様式を検討した。続いて Nucleostemin 特異的な small interfering RNA(siRNA)を用いて Nucleostemin の発現を抑制し、腫瘍 増殖と Nucleostemin の関係を明らかにした。最後に、角化促進実験を行い Nucleostemin 発現が上皮分化マーカーの発現に及ぼす影響について検討を行っ た。
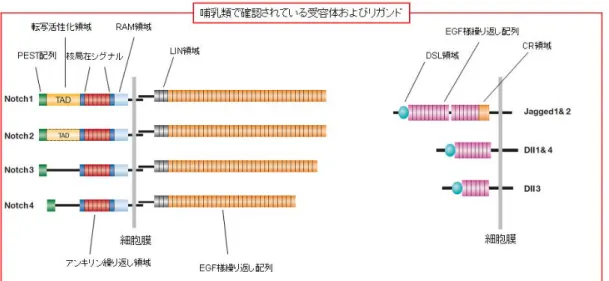
5-2. Notch signaling Notch signaling 伝達経路は、多細胞生物において進化的に保存された経路で、 発生の過程及び幹細胞において細胞の運命の決定を調節する。神経、心臓、免 疫及び内分泌系の発生においては多様に整列した細胞の運命が制御されている が、その際に隣接する細胞間の近傍分泌型 (Juxtacrine) シグナル伝達を示す ことが知られている(28-31)。Notch 受容体は、1 回膜貫通型タンパク質で、機能的 な細胞外ドメイン(NECD)、膜貫通ドメイン(TM)、及び細胞内ドメイン(Notch intracellular domain;NICD)からなる。受容体の種類は全部で 4 種類あり、 Notch1,2,3,4 の名称が付けられている。哺乳類では、シグナルを送る側の細胞 に存在する Delta like(DLL1、DLL3、DLL4)及び Jagged ファミリー(JAG1、JAG2) が、Notch シグナル受容体に対するリガンドとして働く(図 3)。
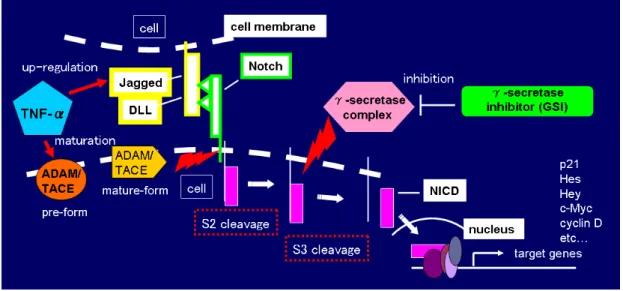
図 3. 哺乳類における Notch signaling 関連蛋白質の種類と構造 リガンドが結合すると直ちに、NECD は TACE (ADAM メタロプロテアーゼ TNF-α変換酵素) によって TM-NICD ドメインから切り離される(S2 切断)。この NECD はリガンドに結合した状態を維持し、この複合体はシグナル送信側の細胞内で エンドサイト―シスとリサイクリング/分解を受ける。シグナル受信側の細胞内
では、-secretase が TM から NICD を遊離する (S3 切断) が、これによって NICD
は核内へ移行し、そこで CSL (CBF1/Su(H)/Lag-1) ファミリー転写因子複合体と 会合するが、その結果、基本的な Notch の標的遺伝子である Myc、p21 及び HES ファミリーメンバーの活性化が起こる(図 4)。
図 4. Notch signaling 伝達経路の概要 Notch signal は、種々の疾患と関連があるため治療標的としての有用性が検 討されてきた(32, 33)。機能欠失型の Notch 受容体及びリガンドの変異は、JAG1 過 剰発現や Notch2 変異により発症するアラジール(Allagille) 症候群や、常染 色体優性型の大脳動脈症であり、Notch3 の変異によりは生じる CADASIL などの 数々の疾患への関与が示唆されている。さらに、NICD の核内への蓄積に結び付 くような Notch 受容体の活性化型変異は、成人 T 細胞急性リンパ芽球性白血病 及びリンパ腫に共通しており(34, 35)、その他の悪性腫瘍でも Notch signaling の 異常が癌の発症や進展に関与しているという報告がある(36-42)。しかし、Notch signaling が腫瘍に促進的に作用するのか、抑制的に作用するのかは細胞系譜や 同じ系譜の細胞でもそれを取り巻く環境によって変化するとされている。 元来 Notch シグナルは種々の細胞の発生・分化・増殖など細胞の運命決定を 司ることが知られているが(43, 44)、これは組織幹細胞の運命決定において Notch siganling が重要な役割を果たしているということである。したがって、最近で は悪性腫瘍における CSC の制御にも深く関与している可能性が示唆されている (45-52)。CSC と Noctch signaling の関わりを示す重要な根拠は乳癌(53-56)、胚性脳 腫瘍(57)、膠芽腫(58, 59)で最も良く示されている。-secretase inhibitor によっ て種々の乳癌細胞株や患者から採取した癌細胞の mammosphere 形成が著しく阻 害され(54)、非浸潤性乳管癌においても様々な系統のスフェロイドを形成する能
力が-secretase inhibitor、抗 Notch4 モノクローナル抗体あるいはゲフィニチ
signaling の feedback loop が関与しているという根拠が示されている(61, 62)。
Sansone らはヒト乳癌患者から作製した mammosphere では、IL-6 によって JAG1 の発現が誘導され、Notch3 signaling が活性化し、CSC の性質の一つである低 酸素抵抗性を示すようになると報告している(63)。Fan らは CD133 高発現分画の変 化と色素排出能を指標とした実験系を用いて、Notch signaling の阻害によって 髄芽腫や膠芽腫の CSC が選択的に除去されることを示している(57, 58)。また、膠 芽腫に発現している Notch は CSC の放射線耐性にも関わっている可能性も示唆 されている(59)。この研究では、-secretase inhibitor 処理により、膠芽腫の CSC 分画のみが選択的に放射線性細胞死を増加させることが分かっている。その 効果は、Notch1 あるいは Notch2 の発現抑制によっても惹起され、Mcl-1 の発現 上昇や AKT signaling の抑制あるいは阻害が関与していることが示されている。 他の癌腫についても積極的に研究が進められており、例えば肝細胞癌の CSC 維 持には、Notch や STAT3、TGF-が関わっている可能性が示唆されている(64)。ジ
ェムシタビン耐性膵臓癌細胞株では、Notch signaling を介して Epithelial to Mesenchymal Transition (EMT)が起こっており、EMT は Weinberg らの提唱する モデルでは幹細胞維持に関与しているとされていることから、Notch signaling
は間接的に転移能が高い CSC の維持に関わると考えられる(65)。また、-secretase
inhibitor(58)や抗 DLL4 抗体(66)を用いて Notch signaling を阻害すると CSC の数
が減少し、造腫瘍能も抑制されることが臨床前モデルで示されている。 頭頸部扁平上皮癌においては、網羅的遺伝子解析などにより Notch signaling が腫瘍発生に関わっている可能性が示される(67-69)。 一方、同じ扁平上皮癌であ る皮膚癌においては腫瘍抑制的に機能することが報告されている(37, 70)。OSCC に おいては Notch signaling 関連分子の発現やその意義についての検討がなされ てきているが(71-76)、未解明な部分が多い。また、口腔扁平上皮およびそれに由 来する OSCC において Notch ファミリーがどのような発現パターンを示し、どの ように OSCC の発生や進展に関わるのかについては現在も研究が進められている 状況である。
本研究においては、OSCC における Notch signaling の機能を明らかにするた めに Notch1 および代表的なリガンドの一つである JAG1 に着目し、舌の正常上 皮、前癌病変(上皮性異形成)そして浸潤癌の組織切片上での Notch1 の発現様 式を検討した。また、発癌過程における同分子の発現パターンを確認する目的 でラット発癌モデルにおいてもその発現を検討した。Notch1 特異的 small interfering RNA(siRNA)および Notch signaling の代表的な阻害剤である secretase inhibitor 用いて Notch1 の発現を抑制し、腫瘍増殖能と Notch signaling の関係を検討した。最後に、腫瘍の浸潤能と Notch1 発現との関係を
との発現様式の相関を Notch1 の免疫組織化学染色で用いたのと同じ検体上で検
討し、そこから得られた結果をもとに更にin vitroにて細胞の遊走能、浸潤能
6. 対象と実験方法 6-1. Nucleostemin に関する実験 6-1-1. 患者検体 患者検体は熊本大学医学部附属病院歯科口腔外科を受診した患者の生検材料 を対象とした。内訳は治療前の舌扁平上皮癌(TSCC)10 例、舌上皮異形成 (dysplasia)25 例、 および異形上皮と連続する形態学的に正常な舌上皮 (normal tongue)10 例とした。 組織標本は全て 10%ホルマリン固定後にパラフ ィン包埋されたものを用いた。上皮異形成の組織標本抽出にあたっては、高分 化型の上皮異形成が存在することを念頭に置き(77)、まず組織標本より薄切した
切片に hematoxylin & eosin (HE)染色を施して病理診断を行ったのち、World
Health Organization (WHO)分類(5)に従って上皮異形成と診断できた検体のみを
使用した。
6-1-2. 免疫組織化学染色と評価法
組織はホルマリン固定後にパラフィン包埋して、厚さ 4m に薄切し、MAS-GP
コートスライド (Matunami Glass Ind., LTD., Osaka, Japan)に接着・固定し た。切片はキシレンで脱パラフィン後、エタノールにて再水和を行い、0.01 mol/L クエン酸緩衝液 (pH 7.0) を用いオートクレーブ (121℃、15 分間)で抗原の賦
活化を行った。その後、0.3% H2O2 含有メタノールにて 30 分間処理し、内因性
ペルオキシターゼ活性の除去を行った。非特異的反応のブロッキングには Protein Block Serum Free Reagent (Dako, Glostrup, Denmark)を用い、15 分 間、湿潤状態で室温にて反応させた。1 次抗体には各々、Nucleostemin (希釈倍 率 1:200,R&D Systems,Abingdon,UK)、Ki-67 (希釈倍率 1:100,MIB-1,Dako)
を用い、湿潤状態で 4℃、一晩反応させた。なお、希釈液には Dako RealTM Antibody
Diluent (Dako)を使用した。続いて 2 次抗体として EnVision+ System-HRP Labelled Polymer (Dako)を標識させ、発色は Liquid DAB+ Substrate Choromogen System (Dako)を用いて行った。脱水・封入に先立って、hematoxylin にて核染 色を行った。 組織染色の評価として、Ki-67 は核が茶褐色に染色されたものを、 Nucleostemin は核小体に一致して茶褐色の染色を認めるものを陽性と判断した。 評価には 2 人の研究者があたり、まず 200 倍視野で染色性の比較的強い部位を 5 カ所選定し、400 倍視野の顕微鏡下で視野内の全陽性細胞数をカウントした。標 識率 (labelling index,以下 LI)を視野内の全細胞数で全陽性細胞数を除し、
その数値に 100 をかけて算出した。5 視野の平均 LI をその組織の LI (%)とした。 6-1-3. 培養細胞 本研究で使用した培養細胞株は HSC-2 細胞、HSC-3 細胞、SAS 細胞、KB 細胞、 Ca9-22 細胞である。ヒト OSCC 由来の培養細胞株であるこれらの細胞は九州大学 大学院歯学研究院口腔顎顔面病態学分野より御供与頂いた。 培養に用いた全ての器具類は滅菌済みのものを使用し、全ての操作はクリー ンベンチ内で無菌的に行った。また、全ての細胞は 10%ウシ胎児血清 (Fetal bovine serum; FBS)を加えた Dulbecco’s modified Eagles medium (DMEM; GIBCO, Gland Island, NY, USA) で 37℃、5% CO2下で静置培養した。
6-1-4. Western blotting 法.
細胞をプロテアーゼ阻害剤カクテル (Roche Diagnostics, Basel,
Switzerland)を含んだ細胞溶解液 (150 mM NaCl, 1%triton X-100, 0.5% sodium deoxycholate, 0.1% SDS, 1mM EDTA-2Na, pH8.0, 1mM EGTA, pH7.5 2.5mM sodium pyrophosphate 1mM β-glycerophosphate, 1mM Na2VO4, 1mM PMSF and 20 mM Tris-HCl, pH 7.5)で溶解した。細胞をボルテックス等で十分に溶解した後、 15000rpm で 30 分間遠心器にかけ不要物を除去した。蛋白質の定量は BCA assay kit (Bio-Rad Laboratories, Hercules, CA)を用いて行い、SDS サンプルバッフ ァーで調製して 95℃で 5 分間加熱処理した。
等量のタンパク質を 0.1%SDS を含むポリアクリルアミドゲルを用いて電気泳 動し (SDS-PAGE)、 PVDF 膜 (Millipore, Hamburg, Germany)に 116mA、1 時間で 転写した。転写後の膜は 0.1%Tween 加 5%スキムミルクトリス緩衝生理食塩水 (TBST)溶液で 1 時間ブロッキングを行った後 1 次抗体で 4℃、一晩反応させた。 TBST で洗浄後に HRP 標識 2 次抗体を室温で 1 時間反応させ、再度 TBST で洗浄し た。発色は ECL Western Blotting Detection System (Amersham Pharmacia Biotech, Buckinghamshire, UK) を用いて行った。用いた 1 次抗体は表 2 の通りである(表 2)。
表 2. 抗体リスト
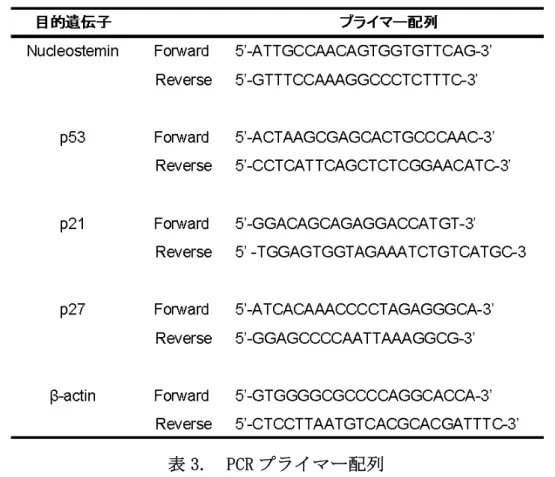
6-1-5. RNA 抽出とreverse transcriptase-polymerase chain reaction (RT-PCR) RNA は TRIZOL Kit (Invitrogen, Carlsbad, CA)を用いて培養細胞株から抽出 した。Oligo (dT)プライマーによる SuperScript Ⅲ First-Strand Synthesis kit (Invitrogen)を用いて 2μg の RNA から逆転写反応によって一本鎖 cDNA を作成 し、ExTaq DNA ポリメラーゼ (Takara)を用い、RT-PCR を行った。RT-PCR 反応条 件は denature、annealing、elongation をそれぞれ 30 秒ずつ行い、denature は 94℃、elongation は 72℃で行った。NS、p53、p21、p27、β-actinのプライマ
ー配列は表 3 に示す通りである(表 3)。PCR 産物は 1.5%アガロースゲルで電気
表 3. PCR プライマー配列 6-1-6. 蛍光免疫染色
培養細胞を蛍光免疫染色に使用する際には、まず培養ディッシュの底面に滅 菌処理した 24mm×24mm のカバーガラスを敷き、その上で細胞培養を行った後に
染色工程を進めた。細胞を 4%パラフォルムアルデヒドで 30 分間固定した後、0.1%
Tween 加リン酸緩衝生理食塩水(PBST)で 3 回洗浄した。non-Specific Staining Blocking Reagent (Dako)で 15 分間ブロッキングした後、カバーガラスを
Nucleostemin 特異的 1 次抗体希釈溶液 (1:200)50 μl の上に注意深く被せた。
4℃、一晩反応させた後に PBST で 3 回洗浄し、Alexa Fluor®
568 donkey anti-goat IgG (Molecular Probes, Eugene, OR)に 1 時間反応させた。PBST で 3 回洗浄後、 2000 倍希釈した 4',6-diamidino-2-phenylindole (DAPI, Sigma Aldrich)に 5 分間反応させ、PBS で十分に洗浄した後、Gel Mount Aqueous Mounting Medium (Sigma Aldrich)で封入し、蛍光顕微鏡で観察した。
している Ca9-22 細胞 (79)を用いた。いずれの実験も対数増殖期にある細胞を用
いた。Transfection 開始の 24 時間前に細胞を回収して新鮮な培地に希釈し、6 well プレートに播種した。通常と同様の条件下で培養を行い、播種より 24 時間 後に Nucleostemin 特異的 small interfering RNA (siRNA)および Stealth™ RNAi negative contorol を Lipofectamine RNAi MAX (Invitrogen)を用いて
Transfection した。Tarnsfection 時の細胞は、約 50%のコンフルエント状態で、
終濃度は 40nM になるように siRNA を調整した。なお、Nucleostemin 特異的 siRNA
の配列は以下の通りである。
Sense 5’- AAG AAC UAA AAC AGC AGC AGA dTdT -3’ Antisense 5’-UUC UUG AUU UUG UCG UCG UCU dTdT -3’
細胞は Transfection から 48 時間後に回収し、Western blotting 法、蛍光染 色、RT-PCR による解析に用いた。
6-1-8. 細胞増殖活性の測定
対数増殖期にある SAS 細胞を 2000 個/100l になるように希釈し、96 穴マイ
クロプレートに 100 μl ずつ播種した。播種から 24 時間後に Nucleostemin 特 異的 small interfering RNA (siRNA)および Stealth™ RNAi Negative Contorol を Transfection した。炭酸ガスインキュベーター内で任意の時間培養し、Cell Counting Kit-8 溶液 (Dojindo, Kumamoto, Japan)を各ウェルに 10 μl ずつ添 加した。炭酸ガスインキュベーター内で 2 時間呈色反応を行った後、マイクロ プレートリーダーを用いて 450 nm の吸光度を測定した。
測定は Transfection を行ってから 24 時間後、48 時間後に行った。
6-1-9. SAS 細胞のin vitroにおける角化誘導
角化を誘導する為に、約 80%コンフルエントの状態にある SAS 細胞を 0.5 mM の
Ca2+を含有する keratinocyte serum-free free-Ca2+ medium (KSFM, GIBCO)で培
養した。コントロールとして、Ca2+濃度が 0.05 mM となるように調製した
serum-free free-Ca2+ KSFM.でも SAS 細胞を培養した。いずれの細胞も 37℃、5%
CO2 下で 5 日間培養し、その間 2 日おきに培地交換を行った。
角化の評価を行うために、Rheinwald and Green (80, 81)の方法に従って
Rhodanile Blue 染色を行った。また、 角化誘導における Nucleostemin 発現と 扁平上皮分化マーカーの変化を観察するために細胞を回収し、Western blotting 法による解析を行った。mouse anti-human cytokeratin19 (CK19; Ks 19.1, PROGEN, Heidelberg, Germany)を未分化の、Involucrin (Invo; mouse anti-human
involucrin, Ab-1, Thermo Fisher Scientific, Rockford, IL)を角化への分化
マーカーとして用いた(82-85)。
6-1-10. 統計学的解析
Nucleostemin と Ki-67 の分布は Chi square test による適合度検定では正規 分布していなかった。したがって、Kruskal-Wallis test と Bonferroni/Dunn test を用いて舌 OSCC (TSCC)10 例、上皮異形成 (dysplasia)25 例、 および正 常舌(normal tongue)10 例の各グループの Nucleostemin と Ki67 の LI の差を検 討した。
Nucleostemin と Ki-67 の発現量の相関関係は Pearson’s correlation coefficient test を用いて行った。Nucleostemin 特異的 siRNA 導入細胞と Stealth™ RNAi Negative Contorol 導入細胞の増殖活性の差は Student’s t-test を用いて行った。
全ての解析には統計ソフト Statcel2 (OMS publisher, Saitama, Japan)を使 用した。統計学的有意差は P 値<0.05 とした。
6-2. Notch signaling に関する実験 6-2-1. 患者検体
患者検体は熊本大学医学部附属病院歯科口腔外科を受診した患者の組織標本 を用いた。内訳は治療前の OSCC (OSCC)12 例、上皮異形成 (dysplasia)36 例、 お よび異形上皮と連続する形態学的に正常な正常口腔上皮 (normal epithelium)12 例。 固定方法や試料の取り扱いに関しては Nuclestemin に関する実験に準じた。 6-2-2. ラット舌検体 発癌化合物 4-nitroquinoline 1-oxide(4NQO)を用いて誘導した(86, 87)ラットの 舌癌、異形成、および発癌誘導していない正常舌の組織検体を東海細胞研究所 の田中卓二先生より御供与頂き、実験に用いた。 6-2-3. 免疫組織化学染色と評価法 1 次抗体には Notch1 (D1E11) XP®
Rabbit mAb (Cell signaling Technology)、 L52 (clone 4G1, Dako)および JAG1 Rabbit monoclonal antibody (Epitomics,
CA, USA)を用いた。Notch1、L52 免疫染色の抗原賦活化には 1mM-EDTA Tris/HCl
Buffe (pH 9.0) を、0.01 mol/L クエン酸緩衝液 (pH 7.0)を用いた以外は Nucleostemin、Ki-67 の免疫組織化学染色と同様に行った。 組織染色の評価として、Notch1、JAG1 は細胞膜が、L52 は細胞質が茶褐色に 染色されたものを陽性と判断した。評価には 2 人の研究者があたり、染色強度 を陰性:1 ポイント、弱陽性:2 ポイント、中等度陽性:3 ポイント、強陽性:4 ポイント、陽性細胞占有率を 5%未満:1 ポイント、6%以上 35%未満:2 ポイント、 36%以上 70%未満:3 ポイント、71%以上:4 ポイントとしてスコア化し、合計ス コアを算出した。合計スコアの平均が 4 以上であったものを陽性と判断した。
Notch1 と L52 の 2 重染色を行なう際には、Notch1 の発色基質には DAB を、L52
の発色基質には HistoGreen ペルオキシダーゼ基質キット (AbCys S.A., Paris, France)を用いた。
6-2-4. 培養細胞
Nucleostemin に関する研究で用いた培養細胞株に加えて、SCC-KN 細胞、OSC-20
細胞、OSC-19 細胞、HOC-313 細胞を使用した。ヒト OSCC 由来の OSCC 細胞株で あるこれらの細胞のうち、SCC-KN 細胞は九州大学大学院歯学研究院口腔顎顔面 病態学分野より、OSC-20 細胞、OSC-19 細胞、HOC-313 細胞は金沢大学大学院が ん医科学専攻がん細胞学講座細胞浸潤学分野より御供与頂いた。
培養に用いた全ての器具類は滅菌済みのものを使用し、全ての操作はクリー ンベンチ内で無菌的に行った。また、全ての細胞は FBS を加えた DMEM(GIBCO, Gland Island, NY, USA) で 37℃、5% CO2下で静置培養した。
6-2-5. Western blotting 法.
基本的に Nucleostemin に関する研究と同様の手技および方法にて行った。 Nucleostemin に関する研究で用いた抗体以外に、Notch signaling に関する研 究で新たに用いた 1 次抗体は以下に示す通りである(表 4)。 表 4. 抗体リスト 6-2-6. RNA 抽出と RT-PCR Nucleostemin に関する研究と同様に行った。各標的遺伝子のプライマー配列 を表 5 に示す(表 5)。PCR 産物は 1.5%アガロースゲルで電気泳動し、エチジウム ブロマイド染色法にて可視化した。
表 5. PCR プライマーリスト 6-2-7. - secretase inhibitor 処理
- secretase inhibitor による細胞処理は SAS 細胞を用いて行なった。- secretase inhibitor には InSolutionTM -secretase inhibitor X (CALBIOCHEM,
Darmstadt, Germany)を使用した。
処理 24 時間前に細胞を播種し、終濃度が 5mM、10mM となるように InSolutionTM
-secretase inhibitor X を添加した。コントロールとして、溶媒である Dimethyl sulfoxide (DMSO; WAKO, Osaka, Japan)を培地に添加した InSolutionTM
-secretase inhibitor X と等量加えた。添加より 48 時間後に細胞を回収し、 Western blotting 法に使用した。
また、増殖活性の検討においては、添加より 24 時間後、48 時間後にの増殖活 性の測定を行った。
6-2-7. RNA 干渉(siRNA)
干渉実験にあたっては SAS 細胞を用いた。基本的に Nucleostemin に関する研 究と同様の手技にて Notch1 特異的 small interfering RNA (siRNA)および Stealth™ RNAi negative contorol を Lipofectamine RNAi MAX (Invitrogen)を 用いて Transfection した。Notch1 特異的 siRNA の配列は以下の通りである。 Sense 5’- UCG CAU UGA CCA UUC AAA CUG GUG G -3’
Antisense 5’-CCA CCA GUU UGA AUG GUC AAU GCG A -3’
細胞は Transfection から 48 時間後に回収し、Western blotting 法に用いた。
6-2-8. 細胞増殖活性の測定
対数増殖期にある SAS 細胞を 2000 個/100l になるように計数・希釈し、96
穴マイクロプレートに 100 μl ずつ播種した。播種から 24 時間後に
InSolutionTM-secretase inhibitor X を前述のように添加、または Notch1 特
異的 small interfering RNA (siRNA)および Stealth™ RNAi negative contorol を Transfection した。炭酸ガスインキュベーター内で任意の時間培養し、Cell Counting Kit-8 溶液 (Dojindo, Kumamoto, Japan)を各ウェルに 10 μl ずつ添 加した。炭酸ガスインキュベーター内で 2 時間呈色反応を行った後、マイクロ プレートリーダーを用いて 450 nm の吸光度を測定した。
測定は Transfection を行ってから 24 時間後、48 時間後に行った。
6-2-9. 傷付けアッセイ
SAS 細胞を 6 ウェルプレートに播種し、100%コンフルエントになるまで培養し、
200l ピペット(Molecular BioProducts, SAN, USA) にて可及的に直線となるよ
うに傷付けを行なった。PBS にて 2 回洗浄を行なって新鮮培地に交換後、コント ロール群には DMSO を、実験群には InSolutionTM -secretase inhibitor X を加
えて 24 時間通常条件化で培養を行なった。傷付け直後と 24 時間後の傷の幅を 測定し、Wound healing area (%) = (24 時間後の傷の幅 / 傷つけ直後の傷の幅) ×100 で算出した。尚、InSolutionTM -secretase inhibitor X および DMSO の添
加量は-secretase inhibitor 処理の量に従った。
6-2-10. マトリゲル細胞浸潤試験
浸潤試験は、マトリゲルでコートされた 8m ポアサイズの PET メンブレン(直
径7mm)からなるセルカルチャーインサート(BD BioCoatTM cell culture
insert, BD Biosciences)を使って行った。カルチャーインサートを直径16
mmのウェル内に置き、血清無添加の DMEM 500l 中に 2×105個のSAS細胞を
調整して、カルチャーインサート上に播種した。下部ウェルには 750l の DMEM
(5% FBS)を入れておいた。SAS 細胞を TNF-(25g/ml)で処理したものをコント
ロールとし、更に DMSO、InSolutionTM -secretase inhibitor X を加えた。
また、Notch1 発現を抑制した細胞を用いる際には播種する 24 時間前に Notch1
特異的 siRNA および Stealth™ RNAi negative contorol の SAS 細胞への導入を
60mm ディッシュ上で行い、同数の細胞をカルチャーインサートに播種して TNF-
処理を行なった。
24 時間後、インサートメンブレンの上面に付着した非浸潤細胞とマトリゲル を取り除いた。浸潤細胞はマトリゲルとインサートメンプレンを通過し、メン ブレン下面に付着しているため、メンプレンを Diff-Quick stain (Systemex, Hyogo, Japan)で固定、染色した。顕微鏡下で 4 か所の視野で浸潤した細胞数を 計測し、平均値によって評価した。
6-2-11. 統計学的解析
Notch1、JAG1 および L52 の各病変におけるスコアおよび傷つけアッセイに関
する多重比較検定は、Bonferroni/Dunn test を用いて行なった。
Notch1 と L52 の発現量の相関関係は Pearson’s correlation coefficient test および Mann・Whitney’s U test を用いて行った。
Notch1 特異的 siRNA 導入細胞と Stealth™ RNAi negative contorol 導入細胞 および DMSO 添加群と InSolutionTM -secretase inhibitor X 群の細胞との増殖
活性の差は Student’s t-test を用いて行った。
全ての解析には統計ソフト Statcel2 (OMS publisher, Saitama, Japan)を使 用した。統計学的有意差は P 値<0.05 とした。
7. 結果 7-1 Nucleostemin
7-1-1. 組織標本における Nucleostemin と Ki-67 の免疫染色の結果
OSCC における Nucleostemin 発現と発癌の関係を調べるために、免疫組織化学 染色を用いて正常舌 (normal tongue)、上皮異形成 (dysplasia)、舌扁平上皮 癌 (TSCC)の各組織における Nucleostemin の発現パターンを検討した。また、 Nucleostemin は細胞の増殖能と関連するという報告があることから、同一の組 織にて増殖マーカーの一つである Ki-67 の免疫染色も同時に行った。 Nucleostemin、Ki-67 ともに核に陽性像が認められた。Nucleostemin は正常 舌上皮から浸潤癌に到る全ての病理組織学的段階で発現が認められた。また、 全ての組織において Nucleostemin の方が Ki-67 よりも LI が高い傾向にあった (図 5)。 図 5. 各病理組織における Ki-67 および Nucleostemin の発現パターン
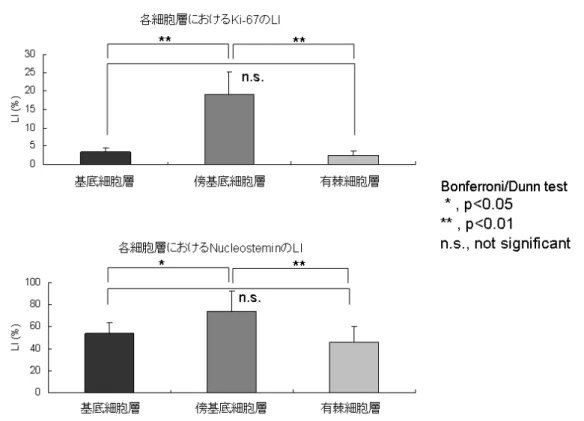
正常舌では Nucleostemin、 Ki-67 ともに最も低い LI を示した。いずれの蛋 白質も傍基底細胞層に最も多く発現していたが、Ki-67 が同層に非常に特異的に 発現しているのとは対照的に、Nucleostemin は基底細胞層だけでなく有棘細胞 層にもその発現を認めた。 その詳細は以下の通りとなる。すなわち、正常舌において Ki-67 は傍基底細 胞層に数は少ないが散在しており、基底細胞層や有棘細胞層にはほとんど認め られなかった。一方で、Nucleostemin の LI は Ki-67 に比べると基底細胞層、傍 基底細胞層、有棘細胞層、全ての層で高かったが、Nucleostemin の発現自体は Ki-67 と同様に傍基底細胞層で最も高かった (図 6)。
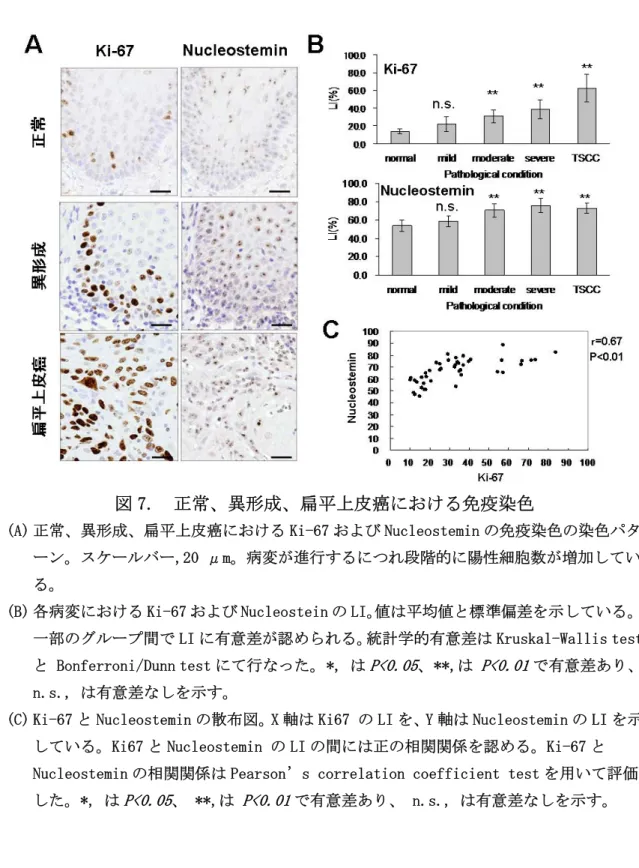
図 6. 各細胞層における Ki67 および Nucleostemin の陽性率(LI) Nucleostemin、Ki-67 の LI はともに病変が浸潤癌へ進行していくにつれて高く なる傾向にあった。特に、中等度異形成 (moderate dysplasia)から舌扁平上皮 癌(TSCC)にかけては正常舌 (normal tongue)と比較すると有意に LI が高かった (mild dysplasia; n.s., moderate, severe dysplasia, TSCC; P<0.01) (図 7A お よび B)。
散布図上で Nucleostemin と Ki-67 の LI の間には相関関係があると考えられ たので、統計学的に解析したところ、P<0.01 で有意に両者は正の相関関係にあ った (r = 0.67) (図 7C)。
と腫瘍性細胞、両者の増殖活性と関係していることが示唆された。
図 7. 正常、異形成、扁平上皮癌における免疫染色
(A) 正常、異形成、扁平上皮癌における Ki-67 および Nucleostemin の免疫染色の染色パタ ーン。スケールバー,20 μm。病変が進行するにつれ段階的に陽性細胞数が増加してい る。
(B) 各病変における Ki-67 および Nucleostein の LI。値は平均値と標準偏差を示している。 一部のグループ間で LI に有意差が認められる。統計学的有意差は Kruskal-Wallis test と Bonferroni/Dunn test にて行なった。 *, はP<0.05、 **,は P<0.01で有意差あり、 n.s., は有意差なしを示す。
(C) Ki-67 と Nucleostemin の散布図。X 軸は Ki67 の LI を、Y 軸は Nucleostemin の LI を示 している。Ki67 と Nucleostemin の LI の間には正の相関関係を認める。Ki-67 と Nucleostemin の相関関係は Pearson’s correlation coefficient test を用いて評価 した。*, はP<0.05、 **,は P<0.01で有意差あり、 n.s., は有意差なしを示す。
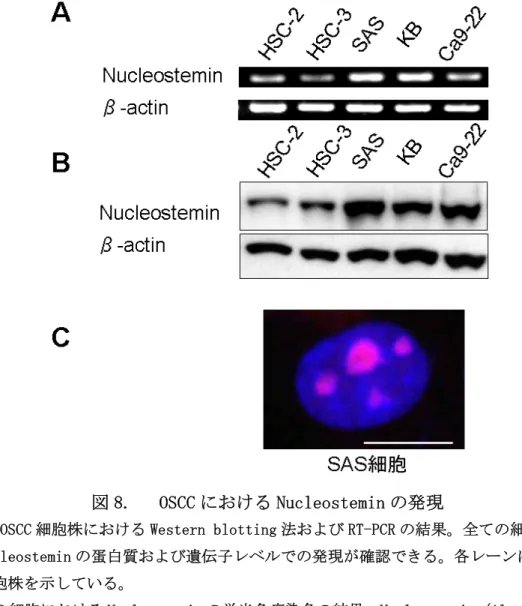
OSCC における Nucleostemin の発現を確認するため、OSCC 細胞株 HSC-2, HSC-3, SAS, KB, Ca9-22 を用いて遺伝子レベル、蛋白質レベルでの解析を行った。
RT-PCR で全ての細胞株に mRNA レベルでの Nucleostemin の発現を認め(図 8A)、 Western blotting 法でも同様に全ての株で Nucleostemin の蛋白質発現を認め た(図 8B)。また、蛍光染色でも核内に明らかな NS の局在を認めた (図 8C)。 これらの結果から他の癌腫と同様に、Nucleostemin は OSCC 由来の細胞におい ても遺伝子、蛋白質レベルで発現していることが明らかとなった。
図 8. OSCC における Nucleostemin の発現
(A) (B)OSCC 細胞株における Western blotting 法および RT-PCR の結果。全ての細胞株で Nucleostemin の蛋白質および遺伝子レベルでの発現が確認できる。各レーンは以下の 細胞株を示している。
(C) SAS 細胞における Nucleostemin の蛍光免疫染色の結果。Nucleostemin (Alexa Fluor®
568, 赤) が SAS 細胞の核 (DAPI, 青)中心部に明瞭に観察できる。スケールバー, 10 μ m。
7-1-3. siRNA による Nucleostemin の発現抑制が細胞増殖活性に及ぼす影響と 細胞周期調節分子の発現の変化
細胞増殖における Nucleostemin の役割を明らかにするために siRNA を用いて Nucleostemin の発現抑制を行い、Western blotting 法にて細胞分裂関連マーカ ーの変化を観察した。加えて、Nucleostemin 特異的 siRNA 導入後の細胞増殖活 性を MTT assay 法を用いて観察した。また、過去の報告では Nucleostemin は p53 と結合しその機能を阻害することで細胞増殖能を促進すると考えられている(10)。 このことから本実験においては wild-type TP53 を保有しているとされる SAS お よび, mutant TP53 を有しているとされる Ca9-22 について p53 をはじめとした 細胞周期調節分子の変化を観察した。
Western blotting 法にて、Nucleostemin 特異的 siRNA 導入後の SAS で明らか な Nucleostemin および細胞分裂マーカーである phospho histone H3 (p-H3)の 減少を認めた (図 9A)。 蛍光免疫染色でも Nucleostemin の明らかな発現減少を 認めた (図 9B)。MTS assay 法による細胞増殖活性の観察でも Nucleostemin 特 異的 siRNA を導入した SAS は contorol siRNA を導入した SAS に比べて明らかな 増殖活性の低下を認めた (図 9C, P<0.05)。また、図 9D に示すようにアポトー シスの代表的マーカーである cleaved caspase-3 (C-casp3)の発現は
Nucleostemin 特異的 siRNA 導入後 48 時間で増加していた。これらの結果から、 Nucleostemin は培養下にある OSCC 細胞株の増殖能にも重要な役割を担っている と結論付けた。
加えて、Nuclestemin 発現抑制下での p53、p21、 p27 などの細胞周期調節分 子の発現変化を、p53 が wild type である SAS、mutant type である Ca9-22 両方 の細胞を用いて観察したところ、図 9D、および図 9E に示すように Western blotting 法、RT-PCR いずれにおいても p53 の発現レベルに明らかな変化は認め なかった。一方で、Nuclestemin 特異的 siRNA を導入した細胞において遺伝子レ ベル、蛋白質レベルの両方で contorol siRNA 導入細胞と比較して p21、p27 の 発現上昇を認めた。 これらの結果から、Nucleostemin の発現抑制によって生じる OSCC 細胞株の増 殖活性の低下および細胞周期停止、アポトーシスは p53 非依存的であるが、p21、 p27 は関わっている可能性があることが示唆された。
図 9. Nucleostemin が細胞増殖と細胞周期調節関連分子の発現に及ぼす影響
(A) Stealth™ RNAi negative contorol (si-cont.)
(B) および Nucleostemin 特異的 siRNA (si-NS)導入から 48 時間後の SAS 細胞における Nucleostemin および mitosis マーカー:phospho-Histone H3(p-H3)の発現変化。 Western blotting 法にて Nucleostemin 特異的 siRNA 導入細胞で明らかな
Nucleostemin および p-H3 の発現低下を認める。NT は未処理の SAS 細胞を示す。 (C) Stealth™ RNAi negative contorol (si-cont.) および Nucleostemin 特異的 siRNA
(si-Nucleostemin)導入 SAS 細胞の蛍光免疫染色。Nucleostemin 特異的 siRNA 導入細 胞で明らかな Nucleostemin の発現の消失を認める。スケールバー, 10 μm。 (D) MTS 法を用いた Stealth™ RNAi negative contorol および Nucleostemin 特異的 siRNA
導入 SAS 細胞の経時的増殖活性測定の結果。導入後 48 時間後、Nucleostemin 特異的 siRNA 導入 SAS 細胞において Stealth™ RNAi negative contorol 導入細胞と比較して 著明な増殖活性低下を認める。統計学的有意差は Student’s t -test を用いて検討し た。 *, はP<0.05、 **,は P<0.01で有意差あり、 n.s., は有意差なしを示す。 (E) Stealth™ RNAi negative contorol および Nucleostemin 特異的 siRNA 導入後の SAS
細胞および Ca9-22 細胞における細胞周期関連蛋白質の発現変化(Western blotting 法)。分子によっては Stealth™ RNAi negative contorol および Nucleostemin 特異的 siRNA 導入細胞間で発現に変化を認める。
(F) Stealth™ RNAi negative contorol および Nucleostemin 特異的 siRNA 導入後の SAS 細胞における Nucleostemin, p53, p21, および p27 遺伝子の発現変化(RT-PCR 法)。 遺伝子によっては Stealth™ RNAi negative contorol および Nucleostemin 特異的 siRNA 導入細胞間で発現に変化を認める。
7-1-4. SAS 細胞株におけるin vitro 角化誘導実験と Nucleostemin、表皮角 化マーカーの発現パターン 培養下での SAS 細胞の分化に伴って Nucleostemin の発現が変化するのかを確 認するために、in vitroでの角化誘導実験とその際の Nucleostemin、上皮分化 マーカーである cytokeratin19、Involucrin の発現量の変化を観察した。 Involucrin は舌を含む口腔粘膜、皮膚の有棘細胞層の上方から発現し始める 分子で、角化へと分化方向を定めた上皮細胞の有用なマーカーである(88-90)。し たがって、舌を含む OSCC の分化マーカーとしても用いられてきた(82, 83, 85)。 cytokeratin19 は重層上皮の基底細胞層マーカーである(84)。 Ca2+ は上皮の分化を司る調節因子の一つとしてよく知られている。細胞内の
Ca2+レベルの上昇は培養下の OSCC 細胞株の最終分化 (角化)を惹起する。OSCC
細胞株における分化能の欠落は適正なレベルの Ca2+ に到達するための機構に欠
陥があることに起因するとされている(91-93)。これらの報告を元に、Ca2+ free で
ある KSFM を用いて細胞外の Ca2+ 濃度を調節することとした。
Rhodanile blue 染色による確認の結果、SAS 細胞の角化は 0.5mM の高 Ca2+濃
度で観察されたが 0.05mM の低 Ca2+濃度では観察されなかった (図 10A)。 次に、
分化した SAS 細胞と未分化の SAS 細胞での Nucleostemin と上皮分化マーカーの 蛋白質レベルでの発現を比較した。その結果、Involucrin と cytokeratin19 は 両者の間で顕著な変化を示したが、Nucleostemin の発現レベルには大きな変化
は無かった (図 10B)。これらの結果から、Nucleostemin の発現レベルはin vitro
図 10. SAS 細胞を用いた in vitro における角化誘導実験と、Nucleostemin および上皮分化マーカーの発現パターンの変化
(A) 高濃度および低濃度 Ca2+存在下で 5 日間培養した SAS 細胞の Rhodanile Blue 染色の結 果。 赤く染色される部分は角化が亢進している部分を示す。0.5 mM Ca2+ 存在下で角化 が増強されている。スケールバー, 200 μm。
(B)角化誘導後の Nucleostemin および上皮分化マーカーの発現変化。Involucrin および Cytokeratin19 は上皮分化マーカーとして用いている。分化マーカーの変動は認めるが、 Nucleostemin の発現に明らかな変化は認めない。
7-2. Notch signaling
7-2-1. 組織標本における Notch1、JAG1 の発現様式
Notch signaling の代表的なレセプターである Notch1 と代表的リガンドであ る JAG1 の正常口腔上皮(normal epithelium)、異形成 (dysplasia)、扁平上皮 癌 (OSCC)の各段階における発現様式を明らかにするために、組織標本を用いて 免疫染色を行なった。図 11 に示すように、Notch1 は正常から異形成、扁平上皮 癌になるにつれて染色性が強くなる傾向があった。また JAG1 についても同様の 結果が得られた(図 12)。 図 11. 正常、異形成、扁平上皮癌における Notch1 の染色結果 正常から異形成、扁平上皮癌になるにつれて染色性が強くなる傾向がある。
図 12. Notch1 および JAG1 の代表的な染色パターン
Notch1 および JAG1 の代表的な染色像を示す。免疫染色結果のスコア化に際し、図中の代表 的な染色像を参考に陰性:1 ポイント、弱陽性:2 ポイント、中等度陽性:3 ポイント、強 陽性:4 ポイントとスコア化して染色強度の判定を行った。
そこで Notch1 および JAG1 の免疫染色結果をスコア化して再評価したところ、
Notch1、JAG1 ともに正常口腔上皮 (normal epithelium)から異形成 (dysplasia)、
扁平上皮癌 (OSCC)と病理組織学的な悪性度が高くなるにつれて免疫染色スコ アは段階的に高くなる傾向がみられ、正常口腔上皮 (normal epithelium)にお けるスコアと比較した場合に扁平上皮癌 (OSCC)において有意にスコアが高か った。また、陽性と判定した検体の割合もスコアと同様に段階的に増加してい た(図 13)。
図 13. Notch1 および JAG1 の免疫染色スコアの結果
Notch1、JAG1 ともに正常口腔上皮 (normal epithelium)から異形成 (dysplasia)、扁平上 皮癌 (OSCC)と病理組織学的な悪性度が高くなるにつれて免疫染色スコアは段階的に高く なる傾向がみられ、正常口腔上皮 (normal epithelium)におけるスコアと比較した場合に 扁平上皮癌 (OSCC)において有意にスコアが高い。また、陽性と判定した検体の割合もスコ アと同様に段階的に増加している。統計学的有意差は Kruskal-Wallis test と Bonferroni/Dunn test にて行なった。 *, はP<0.05、 **,は P<0.01で有意差あり、 n.s., は有意差なしを示す。
7-2-2.ラットモデルにおける正常口腔上皮、4-nitroquinoline 1-oxide(4-NQO) 誘導性異形成、扁平上皮癌での Notch1 の発現様式
ヒト口腔粘膜組織で得られた結果の妥当性を検討する目的で、ラットに発癌 誘導物質 4-nitroquinoline 1-oxide(4NQO)を長期間投与することで誘発した舌
の上皮異形成(dysplasia)、扁平上皮癌(OSCC)、および 4-NQO 未投与のラット
正常舌上皮(normal epithelium)おける Notch1 の発現変化を免疫染色にて観 察した。
図 14A,B に示すように、染色強度、陽性細胞の割合はともに正常舌、上皮異 形成、扁平上皮癌になるにつれて高くなり、陽性と判定した検体数の割合は扁
平上皮癌のグループで明らかに高かった(図 14C)。尚、検体の陽性・陰性の判
図 14. ラット発癌モデルにおける正常口腔上皮、異形成、扁平上皮癌での Notch1 の発現様式 (A) 正常口腔上皮、異形成、扁平上皮癌における Notch1 の代表的染色像(弱 拡大)。 (B) 正常口腔上皮、異形成、扁平上皮癌における Notch1 の代表的染色像(強 拡大)。扁平上皮癌組織で腫瘍細胞に強く Notch1 が発現している。 (C) 各病変における Notch1 陽性・陰性標本数。扁平上皮癌において Notch1 陽性と判定し た標本が多く存在する。
7-2-3. OSCC 細胞株における Notch1 蛋白質および Notch signaling 関連遺伝 子の発現
組織標本上で Notch1、JAG1 の発現上昇が認められたことから、細胞株におい ても Notch signaling 関連分子が発現しているかを確認するために、Western blotting 法で Notch1 および Notch signaling の活性化によって生じる NICD 蛋 白質の発現を、RT-PCR で Notch signaling 関連分子の遺伝子発現を観察した。 Western blotting 法で大部分の OSCC 細胞株に Notch1 および NICD の発現が認
められ、SAS 細胞および Ca9-22 細胞では特にその発現が高かった(図 15)。RT-PCR
では Notch1 や JAG1 以外に Notch signaling の代表的標的遺伝子である Hes1、
Hey1 の発現も確認できた(図 15)。
図 15. OSCC 細胞株における Notch1 蛋白質および Notch signaling 関連遺伝 子の発現
RT-PCR では大部分の OSCC 細胞株で Notch1 や JAG1 の発現や Notch signaling の代表的ター ゲットである Hes1、Hey1 の発現が確認できる。また、Western blotting 法(図中 WB)でも Notch1 の蛋白質レベルでの発現と Notch siganling の活性化に伴って生じる NICD の発現を 認める。
7-2-4. Notch signaling が細胞増殖能に及ぼす影響
OSCC 細胞株における Notch1 の機能を検討するために、Notch signaling の阻
害が腫瘍細胞の増殖能に及ぼす影響をsecretase inhibitor および Notch1 特
異的 siRNA 導入を行なって観察した。なお、実験には Notch1 の発現が最も高い 細胞株の一つであった SAS 細胞を用いた。
図 16A に示すように SAS 細胞にsecretase inhibitor を添加すると顕微鏡下
で浮遊した死細胞の割合が明らかに増加し、Weatern blotting 法で、secretase
inhibitor を投与した場合に Notch receptor の活性化で生じる NICD と細胞増殖
マーカーである p-H3 の発現低下を認めた。MTS アッセイにおいてもsecretase inhibitor 処理した細胞でコントロールにもちいた DMSO よりも有意に細胞増殖 活性が低下していた(図 16B)。Notch1 の発現抑制を行なった場合にも、Notch1 特異的 siRNA 導入より 24 時間後での SAS 細胞はコントロール群に比較して細胞 数が少なく、MTS アッセイでも著しい増殖活性の低下が認められた。また p-H3 の発現低下も認めた(図 17A、B)。
図 16. Notch signaling の阻害が細胞増殖能に及ぼす影響
(A) DMSO、GSI 添加時の SAS 細胞の顕微鏡像と Weatern blotting 法の結果。SAS 細胞に secretase inhibitor(GSI)を添加したものでは浮遊した死細胞の割合が明らかに 増加している。Weatern blotting 法ではsecretase inhibitor を投与した細胞で NICD と細胞増殖マーカーである p-H3 の発現低下を認める。
図 17. Notch 発現抑制が細胞増殖能に及ぼす影響
(A) Notch1 特異的 siRNA 導入時の SAS 細胞の顕微鏡像と Weatern blotting 法の結果。 Notch1 特異的 siRNA を導入した SAS 細胞はコントロール群と比較して細胞数が明らか に少ない。Weatern blotting 法では Notch1 特異的 siRNA 導入細胞で Notch1 の発現抑 制が確認できる。また、細胞増殖マーカーである p-H3 の発現低下を認める。 (B) Notch1 特異的 siRNA 導入時の SAS 細胞の増殖活性の変化。Notch1 特異的 siRNA 導入
細胞(si-Notch1)で、コントロール群(si-control)と比較して有意な増殖活性の低下を 認める。統計学的有意差は Student’s t-test を用いて検討した。 *, はP<0.05、 **,
7-2-5. 腫瘍組織における Notch1 陽性細胞の特徴的な局在と浸潤マーカーL52 との関係 腫瘍組織検体において、図 18A に示すように Notch1 陽性細胞が腫瘍の浸潤先 端部に多く見られる傾向があった。このことから、Notch1 の発現と腫瘍細胞の 浸潤能との関連を検討するために浸潤マーカーとして用いられる L52 の免染を Notch1 で用いた検体で行い、両者の相関関係と局在を検討した。 結果、L52 は Notch1 陽性部位と近似した部位に陽性像が存在し、免疫染色ス コアは Notch1 のスコアと正の相関関係(r = 0.617)を認めた(図 18B、C)。
図 18. 腫瘍組織における Notch1 陽性細胞の特徴的な局在と浸潤マーカー
L52 との関係
(A) 扁平上皮癌組織における Notch1 の局在。腫瘍浸潤部を中心に Notch1 の染色像を認め る。
(B) Notch1 と L52 の 2 重染色像。Notch1 は茶色で、L52 は緑色で可視化されている。こ の標本上では、両者は腫瘍胞巣の辺縁部に集中して発現しており、その局在は近似し ている。
(C) Notch1 の発現と L52 の免疫染色スコアの関係。Notch1 陽性標本では有意に Notch1 陰性標本よりもスコアが高い。統計学的有意差は Mann・Whitney’s U test を用いて 検討した。 *, はP<0.05、 **,は P<0.01で有意差あり、 n.s., は有意差なしを示す。
Notch signaling が SAS 細胞の遊走能や浸潤能に影響を与えるかを調べるた
めに、in vitroにおいて検討を行なった。
図 19A、B に示すように、secretase inhibitor 処理を行なうと傷付けアッ
セイでは細胞遊走能の低下が有意にみられた。また、マトリゲル細胞浸潤試験
では TNF-依存的な SAS 細胞の浸潤はsecretase inhibitor によって明らかに
抑制された(図 19C、D)。さらに、Notch1 発現抑制を行なった SAS 細胞を用いた
場合には、TNF-依存的細胞浸潤はコントロール群に比較して有意に低下してい
図 19. Notch signaling の遮断が細胞増殖能に及ぼす影響
(A) 傷つけアッセイ開始後 12 時間目の培養ディッシュ底面。secretase inhibitor(GSI) 処理したものでは傷つけた部位への細胞遊走が明らかに阻害されている。
(B) 傷つけアッセイ開始 12 時間後に SAS 細胞が遊走した面積の比較。実験は別日に計 3 回行い、統計学的有意差は Student’s t-test を用いて検討した。 *, はP<0.05、 **,
は P<0.01で有意差あり、 n.s., は有意差なしを示す。
(C) GSI 処理時のマトリゲル細胞浸潤試験の結果。未処理(control)の SAS 細胞はほとんど 浸潤を起こさないが、TNF-処理(TNF-)によって著明に浸潤が惹起される。また、そ の細胞浸潤はsecretase inhibitor(GSI)の溶媒である DMSO 処理(TNF-+DMSO)では あまり変化しないが、secretase inhibitor 処理(TNF-+GSI)によって著明に抑制 される。
(D) 各処理における 1 視野当たりの浸潤細胞数。実験は日時を異にして計 3 回行った。統 計学的有意差は Kruskal-Wallis test と Bonferroni/Dunn test にて行なった。 *, は
negative contorol を導入した SAS 細胞(si-control)では、TNF-処理(si-control+ TNF-)による浸潤能の上昇が見られるが、Notch1 特異的 siRNA を導入した SAS 細胞 (si-Notch1+TNF-)では TNF-依存的な細胞の浸潤が減少している。
(F) 各処理における 1 視野当たりの浸潤細胞数。実験は日時を異にして計 3 回行った。統 計学的有意差は Kruskal-Wallis test と Bonferroni/Dunn test にて行なった。 *, は
P<0.05、 **,は P<0.01で有意差あり、 n.s., は有意差なしを示す。