第 1 章 緒論
UCN-01(7-hydroxy-staurosporine, Fig. 1)は, Ca
++及びリン脂質依存性プロテイ ンキナーゼC(PKC)の選択的阻害剤として, Streptomyces sp.
培養液から単離精製 された現在臨床試験中の抗癌剤である 1-3). UCN-01
の抗腫瘍効果を示す作用機序と してはPKC
阻害だけでなく, ヒト癌細胞においてサイクリン依存性キナーゼ(Cdk)を阻害することにより, その細胞周期を
G1
期に集積させる細胞周期特異的な作用 やアポトーシス誘導作用があることが明らかにされている 4-6). さらに , UCN-01
は
in vitro
及びin vivo
において, マイトマイシンC, シスプラチン及び 5-フルオロ
ウラシル等のいくつかの重要な既存の化学療法剤の抗癌効果を増強し, 他の抗癌剤 との併用効果も期待されている 7-9)
.
H N
N N
O H
H CO
3NHCH
3HO O
H
3C 7
Fig. 1 Chemical structure of UCN-01
非常に興味深いことに, この UCN-01 は Phase II臨床試験において, 癌患者に
72
時間定速静脈内投与を行ったところ, 動物実験の結果からは予想も出来ない異常な
体内動態を示した. 動物実験における, 分布容積(Vdss), 全身クリアランス(CLtot) 及び消失半減期(t1/2)はそれぞれ, 6000~17000 mL/kg,600~4000 mL/h/kg
及び3~12 h
という値を示した. 一方, 癌患者におけるVd
ss及びCL
totはそれぞれ79.6~158 mL /kg,
0.0407~0.102 mL/h/kg
と非常に小さな分布容積及び全身クリアランスを示した. さ
らに, 消失半減期は
253~1660 h
と動物実験に比べて約 130 倍もの長い消失半減期 が観察された10).
このような癌患者における異常な動態特性の一因として
, 極めて高い血清蛋白質
との結合性が挙げられている. 事実, 本薬物はヒトα1-酸性糖蛋白質(hAGP)に対
し10
8M
-1という結合定数が報告されている 10). この値は, 血清蛋白質に対する薬
物の結合定数としては異常とも言えるほど大きい. 例えば , ステロイドホルモン
や塩基性薬物は hAGP に強く結合することが知られているが, その結合定数は高々10
5~10
6M
-1程度である11). 加えて, 多くのリガンドが結合するヒト血清アルブミン
(HSA)についても, ビリルビン-HSA複合体の結合定数は
10
6~10
7M
-1である12). 従
って, 現在までのところ, UCN-01 とhAGP
の複合体の結合定数は, 血清蛋白質- リガンド複合体の中で最大と言える. しかしながら, UCN-01がhAGP
に対してな ぜこのような異常とも言える結合定数を示すのかについては全く不明である. また Fuse
等の研究から, 本薬物は動物種AGP
に対してはhAGP
のように強く結合しな いことが明らかにされている10).
hAGP
は, 183個のアミノ酸残基と5
本の糖鎖からなる分子量約44 kDa
の血清糖 蛋白質である13) (Fig. 2). この蛋白質の糖鎖含量は分子量の約45%と他の糖蛋白
質のなかでも極めて高いため, X 線結晶構造解析やNMR
解析等による立体構造の 解明はなされていない.N
C
164
183
54 15 38
5
85 75
72 147
Fig. 2 Structure model of hAGP
Predicted positions of secondary structure were determined from the calculation of the primary structure accoding to Chou and Fasman´s.
●:glycan, ■:-S-S-bond, ○:herical, △:ß-sheet
しかしながら, 円二色性(CD)スペクトルや分子モデルの設計によるデータか ら, 水溶液中では
ß
シート構造に富んでいることが報告されている 14, 15)(Table 1).
Table 1 Estimation of the secondary structure in hAGP
α -helix ß-structure
Contents (%) 20 60
さらに, hAGPには
2
つの異なる遺伝子(hAGP-A 及びhAGP-B/B´)産物, F1*S
及びA
体が約2:1
の割合で存在し, アミノ酸配列が22
残基異なる16-18)(Fig. 3). hAGP
の生体内での役割については十分に理解されていないが, in vivoの炎症モデルを用 いた実験により, 抗炎症作用や免疫抑制作用などが認められている19, 20). 加えて, 血
中におけるhAGP
濃度は約 20 µM であるが, 感染, 炎症及び癌などの疾患時に5~10
倍に上昇することが知られている21, 22).
YVGGQEHFAH LLILRDTKTY MLAFDVNDEK
PETTKEQLGE FYEALDCLRI PKSDVVYTDW
QHEKERKQEE
GES
10 20 30 40
50 60 70 80
90 100 110 120
130 140 150 160
170 180
QIPLCANLVP QIPLCANLVP
VPITNATLDQ VPITNATLDR
ITGKWFYIAS ITGKWFYIAS
AFRNEEYNKS AFRNEEYNKS VQEIQATFFY
VQEIQATFFY
FTPNKTEDTI FTPNKTEDTI
FLREYQTRQD FLREYQTRQN
QCIYNTTYLN QCFYNSSYLN VQRENGTISR
VQRENGTVSR YEGGREHVAH NLELRDTKTL MFGSYLDDEK
NWGLSVYADK
NWGLSFYADK PETTKEQLGE FYEALDCLCI PRSDVMYTDW
KKDKCEPLEK
KKDKCEPLEK QHEKERKQEE
GES F1*S
A
Fig. 3 Amino acid sequence of hAGP variants
Differences in the amino acid sequence between the F1*S and A variant are underlined.
血中において上昇した
hAGP
は多くの薬物と結合して, それらの分布や薬理効果 に影響を与えることが知られている. 例えば, hAGP を過剰発現させたtransgenic
mice
において, HIVプロテアーゼ阻害薬として知られているサキナビルのクリアラ ンス及び分布容積は, control miceに比べて有意な低下が観察され, その結果AUC
の増大が起こる 23). さらに, 白血病患者において, hAGP
濃度とチロシンキナー ゼ阻害剤であるイマチニブの血清中濃度は正の相関を示し, クリンダマイシン併用
により蛋白結合の競合が生じた結果, イマチニブの
AUC
の増大が起こる 24). 加え
て, hAGP の血清中濃度は乳癌, 肺癌及び卵巣癌患者において, 健常人に比べて
有意な増加が観察され, hAGP のバリアントF1*S
及びA
体のそれぞれの発現量が増加する 25)
. これらのことより, hAGP
は明らかに薬物の体内動態において重要な役割を果たしている.
hAGP
分子上の薬物結合部位の研究は, これまで, 蛍光法, 平衡透析法, CDス ペクトル法及び限外濾過法やアミノ酸残基を直接修飾する化学修飾法を用いて盛ん に行われてきた26-30). X
線結晶構造解析はリガンド-蛋白質間の相互作用様式を解析 する方法として非常に有用な方法であるが 31-33), hAGP
のX
線結晶構造解析はなさ れていない. そのためリガンド-hAGP の相互作用の詳細を検討するには, より強
力な解析法が必要となる. これを行う方法として , 光アフィニティラベル法や部
位特異的変異法が挙げられる. 光アフィニティラベル法は , 特異的化学修飾法の
代表であり, 光照射によりニトレン, カルベンと言った非常に反応性の高い化学
的活性種を発生させて, これを結合部位近傍にラベルする方法である34)(Fig. 4).hv
Protein
Photoreactive group
Ligand
Covalent bond
Fig. 4 Photoaffinity labeling techniques
一方, 部位特異的変異法は広く用いられている手法であり
, 蛋白質分子上のア
ミノ酸を他の異なるアミノ酸に置換し, 蛋白質の機能変化を見ることで薬物結合部 位を同定するもので, アミノ酸レベルで結合部位の同定ができるという点で非常に 優れている35, 36).
このような背景の下, 本研究では様々な解析法を駆使して, hAGP に対して異常 な結合性を示す
UCN-01
の結合部位の同定を試みた. まず第2
章において,UCN-01
の誘導体であるUCN-02
及びStaurosporine
を用いて, hAGP に対する結合性を検討した. さらに, hAGP のリガンド結合に重要な役割を果たすと考えられているアミ ノ酸残基を化学修飾し, これらの修飾
hAGP
に対するUCN-01
の結合性を検討した.第
3
章では, [3H]UCN-01
を光ラベル化剤として光アフィニティラベル法を用いて 検討を行った. さらに, 光アフィニティラベル法により同定したアミノ酸残基を
部位特異的変異法により置換させた変異hAGP
を作成し, UCN-01 の結合に関与す るアミノ酸残基の同定を行った. 最後に, 化学修飾法 , 光アフィニティラベル法
及び部位特異的変異法の結果をもとに, hAGP の立体構造モデルを用いてUCN-01
とのドッキングシュミレーションを行い, 結合部位を同定した . 以下に得られた
知見を詳述する.第 2 章 ヒト
1-酸性糖蛋白質( hAGP)と UCN-01 及びその誘導体 の相互作用
第
1
節 序UCN-01 は, ヒト血清蛋白質の中で
hAGP
に対してのみ非常に強く結合すること が知られている. そこで本章では, UCN-01 の C-7位の置換基が異なったUCN-02
及び
Staurosporine
を用いて, hAGP に対する結合性を検討した. さらに, UCN-01がどのような相互作用様式に基づいて
hAGP
に強固に結合しているかを明らかにす るために, UCN-01 と hAGP の結合に及ぼす塩濃度, 脂肪酸添加及びpH
の影響を 調べ, 結合様式について検討した. 最後に , hAGP
のリガンド結合に重要な役割 を果たすと考えられているアミノ酸残基を化学修飾し, これらの修飾hAGP
に対する
UCN-01
の結合性を調べ, 結合部位の同定を試みた.第
2
節hAGP
に対するUCN-01
及びその誘導体の結合特性2-1 UCN-01, UCN-02
及びStaurosporine
のhAGP
に対する結合性評価Staurosporine
は UCN-01 の C-7位の水酸基が脱離した化合物であり,UCN-02
は, こ の水酸基がα-配置したUCN-01
の立体異性体である(Fig. 5). UCN-01, UCN-02 及びStaurosporine
は, PKC及びprotein kinase A(PKA)の阻害効果が異なっており,
PKC
に対するIC
50はそれぞれ0.0041, 0.062
及び0.0027
となり, PKA に対するそ れは0.042, 0.25
及び0.082
という値を示す 37). そこで UCN-01, UCN-02
及びStaurosporine
のhAGP
に対する結合定数(Ka)を算出した(Table 2). UCN-01のKa
は288 ± 75 × 10
6M
-1と3
つの誘導体のなかで最大となり, この値はFuse
等の報 告と一致している10).
一方, C-7 位の水酸基の脱離したStaurosporine
のKa
はUCN-01
のそれと比べて1/25, C-7
位の水酸基がα-配置したUCN-02
では1/200
に低下し, 3 つの誘導体のなかで最小の値となった. UCN-01 とPKD1
(3-phosphoinositide-dependentprotein kinase 1)の X 線結晶構造解析の結果, UCN-01
の C-7 位の水酸基は, PKD1 の薬理効果に重要な役割を果たしているグルタミン(Gln)220 残基と水素結合して いることが明らかになっている. また, UCN-01 とChk1(checkpoint kinase 1)の
X 線結晶構造解析の結果, UCN-01 の C-7 位の水酸基は, Chk1 のATP(adenosine
triphosphate)結合部位においてセリン(Ser)147
残基と水素結合が観察されている38)
. UCN-01
の C-7 位の水酸基とPKD1
及びChk1
との水素結合は C-7 位の水酸基の 脱離した Staurosporine では観察されておらず, この水素結合がPKD1
及びChk1
の 阻害効果において重要な役割を演じていることが明らかにされている. このように,UCN-01
の C-7 位の置換基はその薬理効果において重要な役割を演じており, 本研究で行った
UCN-01
のhAGP
に対する結合においても重要性が示唆された. 興味深 いことに, UCN-01 の C-7 位の置換基がα -配置することにより, UCN-02
においては
1/200
もの結合定数の低下が観察され, UCN-01 のhAGP
に対する特異的な結合には C-7 位の置換基に加えてその配向性が重要な役割を演じていることが明らかに なった.
H N
N N
O
H
H CO
3NHCH
3R O
H
3C 7
R = OH (ß) : UCN-01 OH (α) : UCN-02
=
H : Staurosporine
=
Table 2 Binding parameters of UCN-01, UCN-02 and Staurosporine to hAGP at pH 7.4
Ligands
Ka (× 10
6M
-1) n
UCN-01 UCN-02 Staurosporine
288 ± 75 1.48 ± 0.11 11.33 ± 5.74 0.92 ± 0.04 0.93 ± 0.06 0.91 ± 0.11 All values are mean ± S.D. (n=3).
n:number of binding sites
2-2 UCN-01
とhAGP
の結合におけるNaCl
の影響一般に, 蛋白質と薬物の結合には静電的及び疎水性相互作用が複雑に関与してい ると言われている 39-41)
. これらの相互作用の関与を明らかにする手段としては,
塩 効果, pH の影響, リガンド結合定数, 熱力学的検討及び構造活性相関などによる 解析が知られている. 静電的相互作用の関与を確認する最も簡便な方法の1
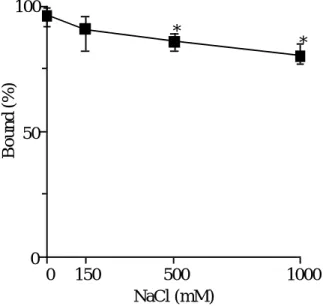
つとし て, イオン強度を変化させて結合性を調べることが知られている. そこで, 最初
に, NaCl 濃度を0
から1000 mM
まで変化させ, UCN-01のhAGP
に対する結合性 の変化について検討した. Fig. 6に示すように, NaCl濃度を変化させたところ, 変 化の程度は小さいものの, NaCl濃度依存的に結合性の低下が観察された.0 150 500 1000
0 50 100
Bound (%)
NaCl (mM)
* *
Fig. 6 Effect of NaCl on binding of UCN-01 to hAGP at pH 7.4
The concentrations are:[UCN-01]=[hAGP]=20 µM. Each point represents the mean±S.D.(n=3). *Statistically significant compared with no NaCl; p<0.01.
なお, 本実験条件下では
NaCl
添加により, hAGP 構造の有意な変化は観察され なかった(Fig. 7).200 210 220 230 240 250
0 mM NaCl 150 mM NaCl 500 mM NaCl 1000 mM NaCl
[ θ ] × 10
-3( deg ・ cm
2dmol
-1) 0
-10 10
Wavelength (nm)
Fig. 7 Far-UV CD spectra of hAGP in the presence of NaCl (0~1000 mM) The concentration is:[hAGP]=10 µM.
これらの結果は, NaCl が
hAGP
の立体構造を変化させた結果, UCN-01 結合を 低下させるのではなく, むしろ, 小さいながら hAGPに対するUCN-01
結合を阻害 していることを示唆している. すなわち, UCN-01と hAGPとの結合には, その程 度の大きさは明らかでないものの, 静電的相互作用が関与していることが推察され る.2-3 UCN-01と
hAGP
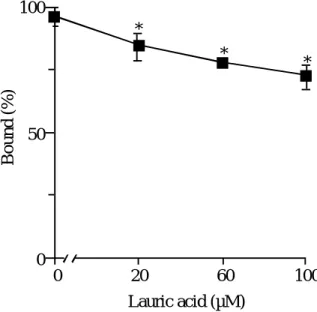
の結合における脂肪酸の影響次いで, 疎水性相互作用の関与を評価するため, 炭素数
12
の脂肪酸であるラウ リン酸を添加して結合率の測定を行った. ラウリン酸濃度は0 µM
から100 µM
ま で変化させた. Fig. 8 に示すように, ラウリン酸添加によって, UCN-01 の結合 は有意に低下した.0 20 60 100 0
50 100
Bound (%)
Lauric acid (µM)
* *
*
Fig. 8 Effect of lauric acid on binding of UCN-01 to hAGP at pH 7.4
The concentrations are:[UCN-01]=[hAGP]=20 µM. Each point represents the mean±S.D.(n=3). *Statistically significant compared with no lauric acid; p<0.01.
脂肪酸添加は, しばしば
HSA
の立体構造を変化させ, ひいてはリガンド結合性 に影響を及ぼすことが知られている 42). しかしながら, このような場合 , 結合性
を低下させるよりむしろ結合性を増加させる 43). 従って, ここで観察された脂肪
酸添加によるリガンド結合性の低下は, リガンドと脂肪酸との置換現象に基づくも のと考えられる. すなわち, これらの現象はhAGP
分子上でのUCN-01
と脂肪酸と の置換反応で説明できるかもしれない. 事実, 脂肪酸濃度依存的にUCN-01
結合が 減少することは, この考えを支持しているものと思われる.以上の結果より, UCN-01 と
hAGP
の結合には疎水性相互作用が関与しているこ とが示唆された.2-4 UCN-01
とhAGP
の結合におけるpH
の影響ところで, 薬物と
HSA
やhAGP
との相互作用は, pHに影響を受けることが知られている44-46)
. これらの変化は, リガンドと蛋白質の解離に伴う相互作用の変化, あ
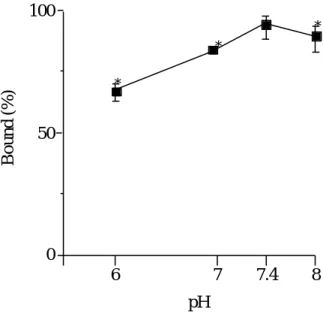
るいは蛋白質の立体構造変化に伴うものと説明されている. そこで, pH 6 から
8
における
UCN-01
とhAGP
の結合性について検討を行った.Bound (%)
pH
*
*
*
6 7 7.4 8
0 50 100
Fig. 9 Effect of pH on the binding of UCN-01 to hAGP
Concentrations are:[UCN-01]=[hAGP]=20 µM. Each point represents the mean ± S.D. (n=3). *Statistically significant compared with pH 7.4; p<0.01.
その結果, Fig. 9 のように, UCN-01 結合性は
pH
上昇に伴って増大し, pH 7.4 において最大であった. UCN-01 の構造とhAGP
の等電点(pI=2.7)から考え合わ せて, pH 7.4 での最大結合率はゲスト-ホスト間の解離現象が程よく絡み合って疎 水性相互作用と静電的相互作用が強化されたものと考えられる. ところで, hAGP
はこれらのpH
領域で立体構造変化を惹起することが考えられる. そこで, hAGP
自身の遠・近紫外領域におけるCD
スペクトルのpH
プロファイルを作成した. 二 次構造においてはいずれのpH
においても有意な構造変化は観察されなかったが(data not shown), 三次構造において
pH
上昇に伴うコットン効果の増加が観察さ れた(Fig. 10).
これらのCD
スペクトル変化は次のように説明される. すなわち,hAGP
は酸性領域では立体構造が緩み, 塩基性領域では堅固になると考えられる . 従っ
て, pH上昇に伴うUCN-01
結合の変化は, pH依存的なhAGP
コンフォメーション 変化もその一因と考えられる.[ θ ] × 10
-3( deg ・ cm
2dmol
-1) 70
0 -20
250 270 290 310 330 350
pH 8 pH 7.4 pH 7 pH 6
Wavelength (nm)
Fig. 10 Near-UV CD spectra of hAGP at various pHs The concentration is:[hAGP]=10 µM.
第
3
節UCN-01
とhAGP
の結合におけるシアル酸及びアミノ酸残基の役割3-1 UCN-01
とhAGP
の結合におけるシアル酸の役割hAGPはその分子内の
5
つのアスパラギン(Asn)残基に枝分かれ構造を有する糖 鎖が結合しており, 血清蛋白質のなかでも最も糖化されている蛋白質の1
つである13)(Fig. 11). いずれの糖鎖末端にも, 負に帯電したシアル酸が多く存在している ため, 最も小さな
pI
値をもつ血清蛋白質でもある.Sia
Sia
Gal
Gal
GlcNAC
GlcNAC
Man
Man
Man GlcNAC GlcNAC Asn
Sialidase
Asialo-hAGP hAGP
Fuc
Fig. 11 Oligosaccharide structure of asialo-hAGP and hAGP
Sia:sialic acid, Gal:garactose, GlcNAc:N-acetylglucosamine, Man:mannose, Fuc:fucose
一般に蛋白質の糖鎖は
, それ自身水溶性を高めるのみならず , 多様な生理シス
テムにおける分子認識や蛋白質のコンフォメーションの安定性に寄与していること が明らかにされている.
薬物が結合する蛋白質の部位は, 多くのアミノ酸からな
るペプチド部分と言われている. しかしながら, UCN-01 はhAGP
に対し10
8M
-1 という非常に高い結合定数を示すため, 蛋白質のペプチド以外の部分に結合する可 能性も考えられる. そこで, hAGP からシアル酸を除いたアシアロhAGP
に対するUCN-01
の結合性を検討した結果, Table 3に示すように, hAGP とアシアロhAGP
との間には
UCN-01
の結合に有意な差異は認められなかった. プロプラノロールのhAGP
に対する結合率は, シアルを除去することによりわずかに減少することが知
られているが 47), UCN-01
とhAGP
との結合にシアル酸は関与していないものと考 えられた.hAGP
Bound (%) 96.33 ± 1.26 95.59 ± 1.06 Native-hAGP Asialo-hAGP
Table 3 Effect of sialic acid on binding of UCN-01 to hAGP at pH 7.4
The concentrations are : [Native-hAGP]= [Asialo-hAGP]= [UCN-01]= 20 µM.
Each value represents the mean ± S.D. (n=3).
3-2 hAGP
バリアントに対するUCN-01
の結合性評価hAGPには,
2
つのバリアント,F1*S
体とA
体の存在が明らかになっている. F1*S 体とA
体において, 全アミノ酸残基中の22
残基が異なっている16-18). 薬物の hAGP
バリアントに対する結合特異性は, 数多く報告されている . 例えば, イミプラミ
ン, アミトリプチリン及びプロパフェノンがA
体に, F1*S 体にはジピリダモール 及びプラゾシンが特異的に結合する. しかしながら, プロゲステロン , プロプラ
ノロール及びクロルプロマジンのhAGP
バリアントに対する選択性は認められていない48)
. 興味深いことには, F1*S
体とA
体の存在比には, 個体差があることが報告されている49)
. 従って, hAGP
に特異的に結合するUCN-01
のhAGP
バリアント に対する結合性を評価することは, UCN-01 の臨床応用に際しての有用な基礎資料 になるものと思われる. そこで, hAGP の2
つのバリアント, F1*S体とA
体に対する
UCN-01
の結合性を検討した.hAGP
Native-hAGP F1*S A
Binding percentage (%) 96.33 ± 1.26 95.08 ± 0.3 95.23 ± 0.45 Table 4 Binding percentage of UCN-01 to hAGP variants at pH 7.4
The concentrations are: [Native-hAGP]=[F1*S]=[A]=[UCN-01]= 20 µM.
All values are mean ± S.D. (n=3).
Table 4 に示すように, 2 つのバリアント間において, 結合性の差異は認められ なかった. 上述したように, 2つのバリアント間において, hAGPの
183
個のアミ ノ酸残基のうち22
残基が異なっているが,UCN-01
の結合性に変化はなかったため,UCN-01
とhAGP
の結合には保存されているアミノ酸残基の関与が考えられる.3-3 化学修飾 hAGP
におけるUCN-01
の結合性評価前述したように, UCN-01 は
hAGP
分子に異常に強く結合する. UCN-01 の結合 に関わり合っているアミノ酸を明らかにすることは, ひいてはUCN-01
の強い結合 性のメカニズムを解くことにつながるものと考えられる. 蛋白質などの生体高分子 と薬物の相互作用部位解明の研究手段としては, X 線結晶構造解析などの分光学的 手法が考えられるが, hAGP 分子上のリガンド結合部位の特定がほとんどされてい ない現状では, これらの方法を使うことはできない. これに比べ化学修飾法は古
典的であるものの, 薬物結合に関与するアミノ酸残基を簡便に予測することができ る. すなわち, 特定のアミノ酸残基が化学修飾を受けたとき , 蛋白質の薬物結合
性に変化が認められれば, そのアミノ酸残基は薬物と蛋白質との結合に関与すると 見なすことができる. そこで,hAGP
のリガンド結合に関与すると言われている28, 50-52),
リジン(Lys), ヒスチジン(His), トリプトファン(Trp)及びチロシン(Tyr)の
4
つのアミノ酸残基をそれぞれ, フェニルイソシアネート, ジエチルピロカル
ボネート(DEP), 2-ヒドロキシ-5-ニトロベンジルブロマイド( HNBB)及びテト
ラニトロメタン(TNM)を用いて化学修飾した , いわゆる修飾 hAGP
を調製し,
UCN-01
との結合性を検討した. Table 5に修飾hAGP
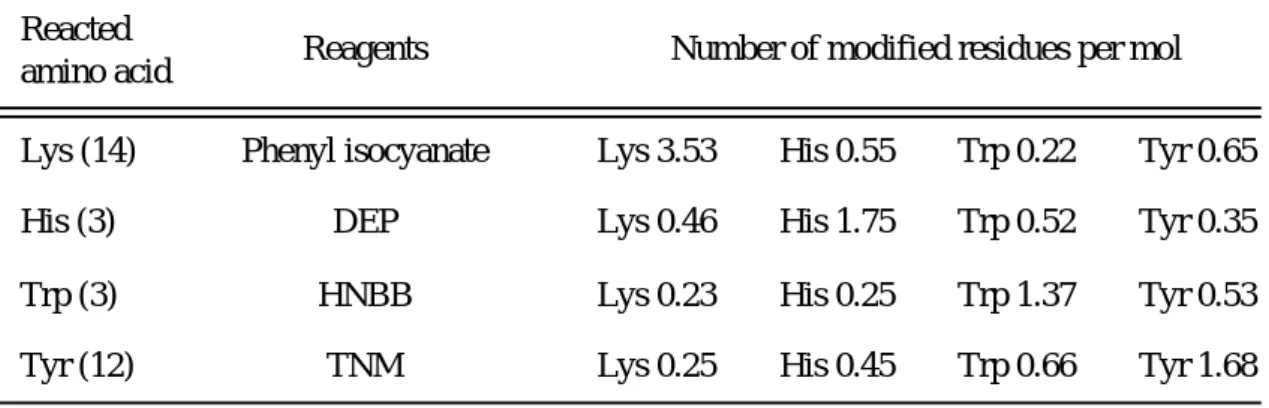
の修飾残基数を示す.Reacted
amino acid Reagents Number of modified residues per mol
Trp (3) Tyr (12) His (3) Lys (14)
HNBB TNM
DEP
Phenyl isocyanate Lys 3.53 His 0.55 Trp 0.22 Tyr 0.65 Lys 0.46 His 1.75 Trp 0.52 Tyr 0.35 Lys 0.23 His 0.25 Trp 1.37 Tyr 0.53 Lys 0.25 His 0.45 Trp 0.66 Tyr 1.68 Table 5 Number of reacted amino acid residues in hAGP
( ): total number in hAGP DEP: diethylpyrocarbonate
HNBB: 2-hydroxy-5-nitrobenzyl bromide TNM:tetranitromethane
その結果, 予測したように
, それぞれの化学修飾剤により , 4
つのアミノ酸残 基が特異的に修飾されていることが判明した. 次に, UCN-01の修飾hAGP
に対す る結合率を測定した結果, Table 6 から分かるように, 4 つのアミノ酸残基をそれ ぞれ修飾したhAGP
に対するUCN-01
の結合は, これらのアミノ酸修飾化に伴って 有意に低下した. 特に, アミノ酸修飾化に伴うリガンド結合性の低下はTrp
修飾体 で顕著であった. これらの結果より, UCN-01とhAGP
の結合には, Trp 残基が重 要な役割を演じていることに加えて, His, Lys 及びTyr
残基も関与していること が示された. さらに, hAGPに3
つ存在するTrp
残基はhAGP
のバリアント, F1*S 及びA
体においてすべて保存されているため, UCN-01のhAGP
バリアントに対す る同程度の結合性は(Table 4), Trp残基の関与を支持する結果と考えられる.Table 6 Binding percentage of UCN-01 to native and chemically modified hAGP at pH 7.4
Chemically modified hAGP
Native- hAGP His Lys Trp Tyr
Binding
percentage (%) 96.33 ± 1.26 70.44 ± 2.68 * 75.97 ± 1.56 * 56.97 ± 2.06 * 73.70 ± 1.01 * The concentrations: [UCN-01]=[Native-hAGP]=[Chemically modified hAGP]=20 µM.
All values are mean ± S.D. (n=3).
*Statistically significant compared with native-hAGP; p<0.01.
3-4 UCN-01
とhAGP
の結合におけるTrp
残基の影響hAGPは, その構造中に
3
個のTrp
残基を25, 122
及び160
位に有しており, そ れらをとりまく環境は大きく異なると考えられている. 例えば, 25
位は蛋白質内 部に,160
位は溶媒に露出,122
位はその中間に位置していると考えられている28, 53).
一般に,Trp
残基の蛍光特性はそれをとりまくミクロ環境に影響されることから, 蛋 白質の立体構造を知る上での有用なプローブとして期待されている.UCN-01
のhAGP
結合におけるTrp
残基の関与の可能性が高いことから, Trp に由来する
hAGP
の蛍光性について修飾hAGP
と比較検討した. その結果, Fig. 12 に示すように蛍光極大波長の長波長シフトと蛍光強度の減少が観察された. 修飾さ れたTrp
残基数がほぼ1
であることと, Trp 残基修飾に際し変性剤を用いていない ことより, 表面に露出している160
位のTrp
残基が修飾されたものと考えられる. す なわち, Fig. 12で観察された蛍光強度の減少には, 160位のTrp
残基が関わってい ることが推察される.300 400
Wavelength (nm)
Intensity
-900 -1400
Native-hAGP Trp modified hAGP
Fig. 12 Fluorescence spectra of Trp residues of native and Trp modified hAGP at pH 7.4
The concentrations are:[hAGP]=[Trp modified hAGP]=1 µM. Excitation was 295 nm, and emission was monitored from 300 to 400 nm.
興味深いことには, 同様な現象が
hAGP
にUCN-01
を添加することにより観察さ れた. これらの蛍光消光はUCN-01
濃度依存的であった(Fig. 13). UCN-01 添加 によるhAGP
分子の蛍光消光は, UCN-01 分子とhAGP
分子とのエネルギー移動に よるものと考えられる. 従って, UCN-01結合には160
位のTrp
残基が深く関わっ ている可能性が示唆された. しかしながら , これらのアミノ酸残基の特定につい
ては, さらに検討が必要と考えられる.Relative fluorescence intensity
[UCN-01/hAGP]
0 0.5 1 1.5 2 2.5 3
0.5 1
0
Fig. 13 Fluorescence quenching titration curve of UCN-01 binding to hAGP at pH 7.4 Concentration is:[hAGP]=1 µM. [UCN-01]=0~3 µM.
第
4
節 小括本章では, UCN-01 がどのような結合様式に基づいて
hAGP
に強固に結合してい るのかを明らかにするために, UCN-01, UCN-02及びStaurosporine
のhAGP
に対す る結合性, UCN-01 と hAGP との結合に及ぼす塩濃度, 脂肪酸添加及びpH
の影響 を検討した. 次いで, hAGP のリガンド結合に重要な役割を果たしていると考えら れるアミノ酸残基を化学修飾し, これらの修飾hAGP
に対するUCN-01
の結合性を 未処理hAGP
と比較検討した. 以下に得られた知見を要約する.1)
UCN-01
の C-7 位の水酸基の脱離した Staurosporine 及び水酸基がα-配置した
UCN-02
のhAGP
に対する結合定数を算出した結果,UCN-01
に比べて, Staurosporine及び
UCN-02
の結合性はそれぞれ1/25
及び1/200
に低下した. UCN-01の C-7 位の 置換基の重要性は, PKD1 やChk1
とUCN-01
とのX
線結晶構造解析の結果からも 報告されており, 本研究で行ったUCN-01
のhAGP
に対する結合においてもその重 要性が示唆された. 興味深いことに,UCN-01
の C-7 位の水酸基がα-配置した UCN-02 においては1/200
もの結合定数の低下が観察され, UCN-01 のhAGP
に対する特異 的な結合には C-7 位の置換基に加えてその配向性が重要な役割を演じていることが 明らかになった.2)UCN-01
のhAGP
結合に及ぼす塩濃度及び脂肪酸添加の影響について検討した結果, UCN-01 の結合性は
NaCl
及び脂肪酸添加により濃度依存的に減少したことよ り, UCN-01 とhAGP
との結合には静電的及び疎水性相互作用が関与していること が示唆された. また, UCN-01のhAGP
結合に及ぼすpH
の影響について検討した ところ,pH
の上昇に伴って結合率は増大し,pH 7.4
において最大となった.UCN-01
の構造とhAGP
の等電点(pI=2.7)から考え合わせて, pH 7.4 での最大結合率はゲ スト-ホスト間の解離現象が程よく絡み合って静電的及び疎水性相互作用が強化され たものと考えられる. さらに, hAGP 自身のCD
スペクトルのpH
プロファイルか ら, 三次構造に若干の変化が観察された. これらの結果より, pH に伴うUCN-01
結合の変化は, ホスト-ゲスト間の複雑な解離状態に伴う相互作用の強化や hAGP
のコンフォメーション変化を反映したものと推察された.3)hAGP
からシアル酸を除いたアシアロhAGP
に対するUCN-01
の結合性を検討した結果, hAGP とアシアロ
hAGP
との間には, UCN-01 の結合に有意な差異は認め られなかったことより, UCN-01 のhAGP
に対する結合にはシアル酸は関与してい ないものと考えられた. また, hAGP の2
つのバリアント, F1*S 体とA
体において
UCN-01
の結合性を検討したところ, これらの結合性に有意な差異は認められなかった. hAGPは
183
個のアミノ酸残基からなり, 2つのバリアント間において22
残基が異なっているが, UCN-01 の結合性に変化はなかったため, 保存されている アミノ酸残基がUCN-01
の結合に関与していることが推察された.4)UCN-01
のhAGP
分子上の薬物結合サイトを評価するために, Lys, His, Trp
及び
Tyr
残基をそれぞれフェニルイソシアネート, DEP, HNBB 及びTNM
を用い て化学修飾したhAGP
を調製し, UCN-01との結合性を検討した. その結果4
つのアミノ酸残基の修飾化に伴って, いずれも
UCN-01
の結合性は低下し, 特にTrp
残 基修飾により顕著に低下した. この結果より, UCN-01 とhAGP
の結合には, Trp 残基が重要な役割を演じていることに加えて, His, Lys 及びTyr
残基も関与して いることが示された. また, hAGPに存在する3
つのTrp
残基はhAGP
のバリアン ト, F1*S 及びA
体においてすべて保存されているため, UCN-01 のhAGP
バリア ントに対する同程度の結合性は, Trp 残基の関与をさらに支持する結果と考えられ る. また, 未修飾hAGP
と修飾hAGP
の蛍光スペクトルを測定した結果, 修飾hAGP
において蛍光極大波長の長波長シフトと蛍光強度の減少が観察された. 修飾されたTrp
残基数がほぼ1
であることと, Trp 残基修飾に際し変性剤を用いていないこと を考え合わせると, 表面に露出している160
位のTrp
残基が修飾されたものと考え られる. すなわち, Trp 修飾hAGP
で観察された蛍光強度の減少は, 160 位のTrp
残基が関わっていることが推察される. また, 同様な現象がhAGP
にUCN-01
を添 加することにより観察され, UCN-01 のhAGP
に対する結合におけるTrp
残基の重 要性がさらに示唆された.本章で得られた結果より, UCN-01 の
hAGP
に対する強固な結合性には, C-7 位 の置換基及びその配向性とhAGP
分子上のTrp
残基が重要な役割を演じていること が明らかになった. また, hAGP分子上の25, 122
及び160
位に存在するTrp
残基 のなかで, 溶媒に露出している160
位のTrp
残基の関与が示唆された. 今後, 修 飾された残基の特定化などの課題は残すものの, UCN-01 のhAGP
に対する特異的 な結合の鍵となるUCN-01
の C-7位の置換基と160
位のTrp
残基の重要性を明らか にすることができた.第 3 章 光アフィニティラベル法及び hAGP の立体構造モデルを用 いた hAGP 分子上の UCN-01 の結合部位の検索
第
1
節 序受容体や抗体, 酵素などの機能性蛋白質は, 特定の物質(リガンド)を認識し, 独 自の機能を発現している
. リガンドがどのような蛋白質を認識するのか , または
蛋白質構造中のリガンド結合部位がどこにあるのかを解析する方法として, 大きく 分けると分子生物学的手法と蛋白質化学的手法の2
つがある. 前者, 特に遺伝子 工学的手法を用いた部位特異的変異法は, 遺伝子の変異により , 蛋白質分子上の
アミノ酸残基を他の性質の異なるアミノ酸残基に置換し, 蛋白質の機能変化を見る ことで薬物結合部位を同定するもので, アミノ酸残基の置換により立体構造変化を 引き起こす可能性があるものの, アミノ酸残基レベルで結合部位を同定できるとい う点で非常に優れている. 一方, 後者の光アフィニティラベル法は, リガンドに
光反応基を導入し, 光照射によりニトレン, カルベンといった反応性の高い化学
種を発生させ, 薬物結合部位とその近傍を特異的に化学修飾するものである34). ま
た, リガンド結合部位が空間的な広がりを持つ部分から構成されている場合でもそ の全容を明らかにすることができる. 現在まで , 光アフィニティラベル法は , P
糖蛋白質54, 55)
, Ca
チャネル56, 57)及び GABAA(γ -aminobutyric acid A)レセプター
58, 59)の基質認識部位を同定するために威力を発揮してきた
. この 2
つの手法は, お互 いの欠点を相補うような関係にあり, 両手法で得られる結果を総合することで, X
線結晶構造解析で得られる「静的情報」に加え, 薬物結合を理解するために必要な「動的情報」を獲得できる. しかしながら, X 線結晶構造解析や
NMR
解析などに よってhAGP
の立体構造が未だ明らかにされていない現状では, これらの情報を得 ることは非常に困難である.このような背景下
, UCN-01
のhAGP
分子上の結合部位の同定を行うために,
[
3H]UCN-01(Fig. 14)を光ラベル化剤として, 光アフィニティラベル法により検討
を行った. まず,
[
3H]UCN-01
を光照射によりhAGP
分子上に光ラベル化を行った. 次 に, ラベル化されたhAGP
の糖鎖の除去を行った後, トリプシンを用いて消化を
行い, アミノ酸レベルでのUCN-01
の結合部位の検索を行った. さらに, 光アフ
ィニティラベル法により同定したアミノ酸残基を置換させた, 変異hAGP
を作成し, ラベル部位の同定を行った. 最後にこれらの結果をもとに , ドッキングシュミレ
ーションを行った.
H N
N N
O
H H CO
3NHCH
3HO O
H
3C 7
*
Fig. 14 Chemical structure of [
3H]UCN-01
*:
3H-labeled position.
第
2
節 光アフィニティラベル法による hAGP 分子上の[
3H]UCN-01
の結合部 位の同定2-1 [
3H]UCN-01
の光ラベル化剤としての有用性まず, [3
H]UCN-01
の光ラベル化剤としての有用性を検討した. 通常, 光アフィ ニティラベルを行う場合は, リガンドに光反応基を導入して光ラベル化剤として用 いるが, [3H]UCN-01
は代表的な光反応基である 34)C=O
基を有しているため, 光反 応基の導入を行わずに用いた. hAGPと[3H]UCN-01
を氷上でインキュベート後, 光 照射したときの電気泳動と放射活性の解析結果をFig. 15
に示す. 電気泳動で流し たゲルを PVDF(polyvinylidene difluoride)膜に転写し,CBB(Coomassie Brilliant Blue
R250)染色した結果, hAGP
のバンドに光照射による断片化は見られず, 単一バンドとして得られた. また, この膜をイメージングプレートに
48
時間コンタクトし て放射活性の強さを解析したところ, 光照射を行った場合のみ放射活性のバンドが
観察され, 光照射により, [3H]UCN-01 と hAGP
との間に共有結合が形成されてい ることが確認できた. また, 共有結合形成の時間依存性について検討を行ったと
ころ, 時間依存的に放射活性のバンドは強くなり,30
分でプラトーに達した(Fig. 16).
この結果より, [3
H]UCN-01
はhAGP
に共有結合しており, 光ラベル化剤として有 用であることが確認できたので, 今後の操作では, 光照射時間をプラトーに達す
る30
分間とし, 光アフィニティラベルを行った.1 2 1 2
97 66 45 20.1 14.4 kDa
97 66 45 20.1 14.4
PVDF membrane kDa Autoradiogram
Fig. 15 Photolabeling of hAGP with [
3H]UCN-01
Lane 1: Sample taken just prior to photoirradiation. Lane 2: Sample taken after 30 min
photoirradiation (>300 nm). 50 µM hAGP and 0.08 µM [
3H]UCN-01 were incubated for 60
min prior to photoirradiation.
0 5 10 15 20 25 30 0
50 100 150 200 250 300
Photoincorporation time (min)
Radioactivity, PSL (a.u.)
45 kDa
Autoradiogram
0 5 10 15 20 25 30 time (min)
Fig. 16 Time course of [
3H]UCN-01 photoincorporation
50 µM hAGP and 0.08 µM [
3H]UCN-01 were incubated for 60 min prior to photoirradiation.
The incubation mixture was irradiated for each time and separated with 10% SDS-PAGE.
2-2 光アフィニティラベル法による各種薬物の[
3H]UCN-01
結合への阻害効果超遠心分離法を用いた置換実験の結果より, hAGP 分子上の
UCN-01
の結合部位 は, ステロイドホルモン, 酸性薬物及び塩基性薬物の結合部位とオーバーラップ
していることが報告されている 60). そこで, [
3H]UCN-01
による光ラベル化の特異 性を確認するために, 光アフィニティラベル法を用いて置換実験を行った . 置換
剤として, ステロイドホルモン類のプロゲステロン, 酸性薬物のワルファリン及
び塩基性薬物のプロプラノロールを用いた. さらに, cold UCN-01, UCN-01のC-
7
位の水酸基がα -配置した UCN-02
及び水酸基の脱離したStaurosporine
も置換剤と して用いた. [3H]UCN-01
を光ラベル化する際, 置換剤を過剰量加え, hAGP のバ ンドの放射活性がどの程度阻害されるか検討した(Fig. 17).45 kDa
Autoradiogram
1 2 3 4 5 6 7
0 50 100 150 200 250
*
*
*
*
*
*
Radioactivity, PSL (a.u.)
Fig. 17 Photolabeling of hAGP with [
3H]UCN-01 in the presence of competitors 50 µM hAGP and 0.08 µM [
3H]UCN-01 were incubated for 60 min prior to photoirradiation in the presence of 250 µM competitors. The incubation mixture was irradiated for 30 min and separated with 10% SDS-PAGE.
Lane 1: no competitor, Lane 2: cold UCN-01, Lane 3: Staurosporine, Lane 4: UCN-02, Lane 5: warfarin, Lane 6: progesterone, Lane 7: propranolol.
置換実験の結果, すべての
UCN-01
誘導体により, 結合定数に依存した60%以
上の強い阻害効果が観察された. また, プロゲステロンによって 58%の阻害効果
が観察され, ワルファリン及びプロプラノロールの阻害効果はそれぞれ16%及び 26%
となった(Table 7). 各薬物により阻害効果の程度に差異が観察された
. これは,
UCN-01
の結合部位と各薬物の結合部位のオーバーラップしている領域の差異のためと考えられる. 一方, cold UCN-01, UCN-02 及び Staurosporine においては, 結 合定数に依存した阻害効果が観察されたため, これらの誘導体は
UCN-01
と同じ領 域に結合するものと考えられる.Cold UCN-01
aStaurosporine
aUCN-02
aWarfarin
bProgesterone
bPropranolol
bKa (M
-1) 2.88 × 10
81.13 × 10
71.48 × 10
61.08 × 10
61.00 × 10
52.98 × 10
5Inhibition (%) 86.38 68.54 61.52 16.43 58.71 26.26 Competitors
Table 7 Binding affinity constant and inhibition percentage by the competitors
a
Binding constant data taken from Ref. 61.
b
Binding constant data taken from Ref. 62.
2-3 hAGP
の脱糖化UCN-01
のhAGP
上の詳細な結合部位の同定を行うため, ラベル化後のhAGP
を酵素を用いて消化を行い, 最後にアミノ酸解析を行う必要がある
. しかしながら, hAGP
はその分子内に15, 38, 54, 75
及び85
位に糖鎖が結合しており(Fig. 18), その糖鎖含量は分子量の約45%と他の糖蛋白質のなかでも極めて高いため, 酵素消
化やアミノ酸解析に影響を及ぼすことが推察される. そこで, hAGP に5
本存在す る糖鎖をN-glycosidase F recombinant(PNGase F)を用いて脱糖を行った. その結果 ,
24 kDa 付近に単一バンドとして現れ, ほぼ完全に hAGP
の糖鎖を除去することができた(Fig. 19). また, 糖鎖除去後の
hAGP
は, 未処理hAGP
と同様に放射活 性を持っているため, UCN-01 の結合にhAGP
の糖鎖は関与していないものと考え られる.Fig. 18 Glycosylation position of hAGP
●: glycans
15 38 54 75 85
hAGP
0 183
43 29 18.4 14.3 6.2 kDa
3
43 29 18.4 14.3 6.2 kDa
3
PVDF membrane Autoradiogram
1 2 1 2
Fig. 19 Deglycosylation of hAGP by PNGase F
Lane 1: untreated hAGP. Lane 2: deglycosyled hAGP.
2-4 逆相 HPLC
によるラベルペプチドの粗精製hAGPの糖鎖を除去した後, Lysとアルギニン(Arg)残基の
C
末端を認識して消 化するトリプシンを用いてhAGP
の断片化を行った. 次に, 不要な試薬である界
面活性剤やトリプシンを除去するために, 逆相HPLC
によってラベルペプチドの精 製を行った. 逆相HPLC
により分離した溶離液を30
秒ごとに分取して, 各フラク ションを液体シンチレーションカウンターで放射活性を測定した結果, 10.5~11.5
分のフラクションが高い放射活性を持っており, [
3H]UCN-01
がラベル化されてい るペプチドが溶出しているものと考えられる(Fig. 20A, 20B). そこで, この2つ のフラクションを分取, 濃縮後, Capillary HPLCのサンプルとした.0 200 400 600 800
0 10 20 30 (min)
dpm
0 10 20 30
A)
B)
(min)
Fig. 20 Reverse-phase HPLC separation of tryptic peptides of hAGP photolabeled with [
3H]UCN-01
An aliquot of 20 µL of the tryptic peptides were applied on a C
18-column and eluted at 1 mL/min using an aqueous acetonitrile gradient in the presence of 0.1% TFA (from 5 to 95%
acetnitrile over the course of 40 min).
A), UV-absorption (210 nm); B), radioactivity of the reverse-phase fractions (200 µL) was determined by scintillation counting.
2-5 Capillary HPLC
によるラベルペプチドのN
末端アミノ酸解析逆相
HPLC
により精製後, さらにCapillary HPLC
を用いてラベルペプチドの分離 を行った. Capillary HPLCは, 分離した溶離液を直接PVDF
膜にブロットするため, ブロットされた膜を切り取り, アミノ酸解析を行うことができる . 溶離液をブロ
ットしたPVDF
膜の放射活性を測定した結果, 84~85 分で観察されたピークが高い 放射活性を持っていることが明らかになった(Fig. 21A, 21B). そこで, 非標識のUCN-01
を用いて同様に光アフィニティラベルを行い, 酵素消化と精製を行った. 放 射活性に対応するピーク部分を切り取り, マイクロシークエンサーによるN
末端ア ミノ酸解析を行った. その結果, N 末端からSDVVYTDXK
というペプチドが同定 された(Fig. 22). hAGP 分子は183
個のアミノ酸からなり(Fig. 23), Edman 分 解により同定されたペプチドは153
位のSer
残基から161
位のLys
残基に相当する. さ らに, この1~9
サイクルのうち8
サイクル目, つまり160
位のTrp
残基だけが全く 同定されなかった(Fig. 22). 通常, Edman 分解においてアミノ酸が同定できなか った場合は, 糖化, 酸化及びリガンドによる修飾が考えられる. 本研究において, 糖 は用いておらず, ペプチド分離後は酸化の影響を考慮して早急にアミノ酸解析を行 った. さらにアミノ酸解析は, 放射活性に対応するピークを切り取って行ったた
め, [3
H]UCN-01
による光ラベル化がおこっているため, アミノ酸が同定できなか
ったものと考えられる.
Retention time (min)
0 30 60 90 120 150 180
A)
B)
Fig. 21 Chromatogram of capillary HPLC (A) and autoradiogram of blotted PVDF membrane (B)
After purified peptides using reverse-phase HPLC, an aliquot of 10 µL of the evaporating sample was applied on a C
18-column and eluted at 5 µL/min using an aqueous acetonitrile gradient in the presence of 0.1% TFA (from 5 to 95% acetonitrile over the course of 200 min).
The blotted membrane from the capillary HPLC separation was in contact with an imaging
plate for 48 h prior to autoradiography analysis.
Number of cycle
Yield of PTH-amino acid ( pmol )
1 2 3 4 5 6 7 8 9
S
153D
154V
155V
156Y
157T
158D
159X
K
1610 0.5 1 1.5 2 2.5 3
Fig. 22 N-terminal amino acid sequence analysis by the Edman degradation method
QIPLCANLVP VPITNATLDQ ITGKWFYIAS
AFRNEEYNKS VQEIQATFFY FTPNKTEDTI
FLREYQTRQD QCIYNTTYLN VQRENGTISR
YVGGQEHFAH LLILRDTKTY MLAFDVNDEK
NWGLSVYADK PETTKEQLGE FYEALDCLRI
KKDKCEPLEK QHEKERKQEE
GES
10 20 30
40 50 60
70 80 90
100 110 120
130 140 150
160 170 180
PKSDVVYTDW
Fig. 23 Amino acid sequence of the photolabeled region of hAGP
2-6 光アフィニティラベル法による変異 hAGP
分子上の[3H]UCN-01
の結合性評価化学修飾の結果より, UCN-01 の
hAGP
に対する特異的な結合には, Trp 残基が 関与する可能性が示唆された(Table 6). しかし, hAGP 分子中には25, 122
及 び160
位にTrp
残基が存在するため, Trp 修飾により観察された結合率の顕著な減 少が, いずれのTrp
残基修飾によるものなのかは明らかではない. そこで光ラベル化における
Trp
残基の重要性を明らかにするために, 多くの組換え型蛋白質発現系 に用いられているメタノール資化酵母であるピキア酵母を用いて, recombinant
hAGP(rhAGP)を作成した. 25, 122
及び160
位の3
つのTrp
残基をそれぞれアラニン(Ala)残基に置換させた, W25A,
W122A
及びW160A
を精製後, これまで と同様に[3H]UCN-01 を加えて光照射を行った. その結果, W25A
及びW122A
の放 射活性は, Wild タイプと比べて有意な差異は認められなかったが, W160A におい て, 有意な減少が観察され,160
位のTrp
残基をAla
残基に置換させることにより, 放 射活性において約80%の減少が観察された(Fig. 24). これまで化学修飾やドッキ
ングモデルを用いた検討により, 2528)及び 122 位63)のTrp
残基の関与は示唆されて いるものの, 160 位のTrp
残基の関与は報告されていない. そのため, 本項で同定 した 160 位のTrp
残基は, hAGP に特異的に結合する UCN-01 の新規の結合部位と 考えられる.0 50 100 150 200
Wild-type W25A W122A W160A Autoradiogram
* 45 kDa
Radioactivity, PSL (a.u.)
Fig. 24 Photolabeling of wild type, W25A, W122A and W160A with [
3H]UCN-01 50 µM rhAGP and 0.08 µM [
3H]UCN-01 were incubated for 60 min prior to photoirradiation.
The incubation mixture was irradiated for 30 min and separated with 10% SDS-PAGE.
*Statistically significant compared with wild type; p<0.01.
第
3
節 hAGP と UCN-01, UCN-02及びStaurosporine
とのドッキングシュミ レーション3-1 hAGP
とUCN-01
とのドッキングモデル現在, hAGP の立体構造は明らかにされていないが, Kopecky らは, hAGP と同 じリポカリンファミリーに属する
4
つの蛋白質のX
線結晶構造解析の結果を鋳型に して,hAGP
の立体構造モデルを構築した63). そこで, このモデルを用いて, UCN-01
のドッキングシュミレーションを行った. リガンドであるUCN-01
は, 細胞の癌化 に関与するChk1
とのX
線結晶構造解析の結果を用いた 64). これまでの結果より,
UCN-01
は明らかに160
位のTrp
残基と相互作用していると考えられるため, hAGPの立体構造モデルにおいて,
160
位のTrp
残基付近の結合ポケットの検索を行った. そ の結果, hAGPの分子表面にType I
とType II
の2
つの結合ポケットの存在が明ら かになった(Fig. 25).Type I
Type II Trp160
Fig. 25 Type I and II docking models of UCN-01 and hAGP
また, Table 8 には, Type I及び
II
モデルにおける5Å
以内のdonor
とacceptor
の 相互作用様式を示す. Type I のモデルにおいて, UCN-01 は 43 位のグルタミン酸(Glu43)
やTyr50
との水素結合が観察された. 一方, Type IIモデルにおいてUCN-
01
は, Trp160との水素結合やスタッキング相互作用が観察され, さらにLys135
やLys161
との静電的相互作用, 加えてTyr157
とのスタッキング相互作用が観察された. 化学修飾の結果(Table 6)と光ラベル化における
160
位のTrp
残基の関与を考 え合わせると, Type IIにおけるアミノ酸残基のすべてが化学修飾の結果と一致して おり, 160 位のTrp
残基との相互作用も観察されている. そのため, Type II モデ ルが実験結果とよく一致したモデルと考えられる.Donors Acceptors Interactions Distance (Å)
Type I
Lys135 Trp160
Lys161 Lys161 Tyr157
UCN-01 (-O-) UCN-01 (C=O)
UCN-01 (OH) UCN-01 (C=O) UCN-01
Electrostatic Hydrogen bonding
Electrostatic Electrostatic Stacking
4.760 2.878
4.727 4.350 4.204 Table 8 Interaction and distance between donors and acceptors in the model of type I and II (UCN-01-hAGP)
Type II
UCN-01 (NH
2+) Glu43 Hydrogen bonding 2.793
Tyr50 UCN-01 (C=O) Hydrogen bonding 2.770
Trp160 UCN-01 Stacking 2.878
実験結果とよく一致した
Type II
モデルの結合ポケットは, 28 位のイソロイシン(Ile28),
131
位のプロリン(Pro131),Glu132, Lys135, 138
位のロイシン(Leu138),Tyr157, Trp160
及びLys161 残基で構成されており(Fig. 26), Trp160
残基はUCN-01
のC=O
基と水素結合しており, また, Lys161 残基はUCN-01
のC=O
基とC-7
位 のOH
基との静電的相互作用が観察された(Table 8). さらに, UCN-01の糖部分 の酸素原子は, Lys135 と静電的相互作用をしており, Type II ドッキングモデルに おいて, 160位の Trp 残基に加えて, 新たに, Lys135, Tyr157及びLys161
の重要 性が明らかになった. Lys, Trp 及びTyr
残基は化学修飾によりUCN-01
の結合率 の低下が観察された(Table 6). この結合率の低下はドッキングモデルにおいて観 察された UCN-01 と Lys, Trp 及びTyr
残基との水素結合, 静電的相互作用及びス タッキング相互作用が, 化学修飾により消失したためと考えられる . また, 化学
修飾により
His
残基の重要性も明らかになっているが, ドッキングモデルにおいて, 相互作用は観察されなかった. アミノ酸解析により同定したペプチドには,His172 が
最も近くに存在しているが, 本項で用いたhAGP
の立体構造モデルは, 170番目以 降のアミノ酸残基は正確な立体構造を構築できていないため, 相互作用が観察され なかったものと考えられる.UCN-01 Lys135
Trp160
Lys161
Pro131 Ile28 Tyr157
Glu132
Leu138
Fig. 26 Amino acid residues in a surface cleft around Trp160 that interacts with UCN-01 exhibited in type II docking model
Dotted line: electrostatic interaction. Arrow: hydrogen bonding.
3-2 hAGP
とUCN-02
及びStaurosporine
とのドッキングモデルUCN-01
に加えて, Type IIモデルにおける, UCN-02及びStaurosporine
のドッキ ングシュミレーションを行った. UCN-01, UCN-02 及びStaurosporine
は, それぞ れC-7
位の置換基が異なっているため, ドッキングモデルには C-7
位の置換基とhAGP
のアミノ酸残基との相互作用に差異が認められた. Type II モデルにおいて,UCN-01
の C-7位のß-OH 基は Lys161
と相互作用をしているのに対して(Table 8),UCN-02
のC-7
位のα
-OH 基はGlu132
との相互作用が観察された(Table 9).
一方,C-7
位に OH 基を有していないStaurosporine
では, これらの相互作用が観察されなかっ た.UCN-01, UCN-02 及び
Staurosporine
のhAGP
に対する結合定数は, UCN-01 >Staurosporine > UCN-02
の順で低下することが明らかになっている(Table 2). UCN-01
の芳香環はTrp160
とスタッキング相互作用しており, この相互作用は, Trp160 と 同じ方向にある Lys161 とUCN-01
の C-7 位のOH
基との静電的相互作用により強 められるものと考えられる. 一方, UCN-02 の C-7 位のOH
基は, Trp160 と反対側で
Glu132
と水素結合しているため(Fig. 27), Trp160 とのスタッキング相互作用が弱まり



![Fig. 7 Far-UV CD spectra of hAGP in the presence of NaCl (0~1000 mM) The concentration is:[hAGP]=10 µM.](https://thumb-ap.123doks.com/thumbv2/123deta/5728757.2025938/9.892.288.615.105.423/fig-far-spectra-hagp-presence-nacl-concentration-hagp.webp)
![Fig. 10 Near-UV CD spectra of hAGP at various pHs The concentration is:[hAGP]=10 µM.](https://thumb-ap.123doks.com/thumbv2/123deta/5728757.2025938/12.892.275.613.94.400/fig-near-uv-spectra-hagp-various-concentration-hagp.webp)