RORγt-CCR6-CCL20経路は関節リウマチ患者のTh17
細胞の炎症局所への遊走を促進する
著者
金子 駿太
発行年
2018
学位授与大学
筑波大学 (University of Tsukuba)
学位授与年度
2017
報告番号
12102甲第8697号
URL
http://doi.org/10.15068/00152477
筑波大学
The RORγt-CCR6-CCL20 axis augments Th17 cells invasion into the synovial fluid of rheumatoid arthritis patients
(RORγt-CCR6-CCL20 経路は関節リウマチ患者の
Th17 細胞の炎症局所への遊走を促進する)
2017
筑波大学大学院博士課程人間総合科学研究科
金子駿太
目次
要旨……….5 第1章 背景……….7 第2章 患者と方法……….9 2-1 患者………9 2-2 PBMC 及び SFMC 中の CD4 陽性 T 細胞の単離………..10 2-3 フローサイトメトリー………10 2-4 サイトカインとケモカインの測定………11 2-5 遊走能解析………12 2-6 統計学的解析………12 第3章 結果………..14 3-1 HC の Th 細胞の細胞表面マーカー、転写因子と サイトカインの解析……….….14 3-2 RA 患者、HC、OA 患者の Th 細胞の細胞表面マーカー、 転写因子とサイトカインの解析……….….14 3-3 RA 患者の PBMC と SFMC 中の Th 細胞の比較……….….15 3-4 Th 細胞の遊走能の解析……….….15 3-5 RA 患者の治療前後の Th 細胞の細胞表面マーカー、 転写因子の比較……….….16第4章 考察……….…….18 第5章 結論……….…….22 参考文献………..23 表……….……….32 図……….……….33 謝辞……….……….51
要旨
目的: 関節リウマチ(rheumatoid arthritis;RA)は全身の関節の炎症を主体とし た自己免疫性疾患であり、CD4 陽性 T 細胞が RA の病態に関与していることが 知られている。近年細胞表面マーカーによるヒトのCD4 陽性 T 細胞サブセット の分類が定義され、RA を含む様々な疾患において統一された分類方法を用いた CD4 陽性 T 細胞サブセットのより詳細な解析が実施されるようになった。また、 CD4 陽性 T 細胞サブセットは特定の転写因子の発現により分化、機能、増殖が 制御されることが明らかになっているが、RA において転写因子の発現の変化が CD4 陽性 T 細胞の分化や機能に影響し、病態に関与しているか否かについては 未だ不明な点が多い。本研究では細胞表面マーカーによる分類を用いて定義し たCD4 陽性 T 細胞サブセットにおける転写因子の発現と、RA の病態との関連 を明らかにする。 方法: 健常者(healthy control;HC)、変形性関節症(osteoarthritis;OA)患者、RA 患者の末梢血から末梢血単核細胞(peripheral blood mononuclear cell; PBMC)と OA 患者、RA 患者の滑液から滑液単核細胞(synovial fluid
mononuclear cell; SFMC)を磁気細胞分離(magnetic cell sorting;MACS)
によりCD4 陽性 T 細胞を単離した。CD4 陽性 T 細胞は細胞表面マーカー
(CD45RA、CXCR5、CXCR3、CCR6)によりヘルパーT(Th)細胞、さらに
の発現をフローサイトメトリーにより解析した。また、末梢血中と滑液中のケ
モカインについてLEGENDplex により濃度を測定し、末梢血中の Th 細胞の遊
走能についてchemotaxis assay により評価した。RA 患者において抗リウマチ
薬治療開始24 週時と治療前での Th 細胞サブセットの割合や転写因子の発現に ついて比較した。 結果: PBMC 中の Th 細胞サブセットの割合については、HC、OA 患者、RA 患者 で有意な差は認められなかったが、RA 患者の Th17 細胞における CCR6 と RORγt の発現が HC と OA 患者に比べ有意に亢進していた。さらには RA 患者 のTh17 細胞における CCR6 と RORγt の発現は有意な正の相関を認めた。RA 患者のSFMC 中の Th17 細胞の割合は PBMC 中に比べて有意に増加していた。 Chemotaxis assay において RA 患者の Th17 細胞の CCL20 に対する遊走能が HC、OA 患者に比べて有意に亢進していた。また、RA 患者の滑液中の CCL20 の著明な濃度上昇を認めた。RA 患者において抗リウマチ薬治療開始 24 週時に は治療前に比べ、Th17 細胞における CCR6 と RORγt 発現が有意に低下してい た。 結論: Th17 細胞の RORγt 発現の亢進と滑液中の CCL20 の上昇が CCR6 発現亢進 を介してTh17 細胞の炎症関節への遊走を惹起し、これらの事象が RA の病態に 関与していると考えられた。
第 1 章 背景 RA は全身の関節の炎症を主体とし、肺や腎臓、心血管系など全身性に症状を きたす自己免疫性疾患である。RA の病因、病態については未だ不明な点が多い が、CD4 陽性 T 細胞の関与が報告されている[1]。 CD4 陽性 T 細胞は Th1 細胞、Th2 細胞、Th17 細胞や制御性 T(regulatory T; Treg)細胞などのサブセットに分類され、それぞれ異なるサイトカイン産生パ ターンや機能を有し、細胞性もしくは液性免疫や自己免疫性疾患の発症に関与 する[2]。また、それぞれのサブセットで T-bet、GATA3、RORγt、Foxp3 など 異なる転写因子が発現し、CD4 陽性 T 細胞の分化、制御に関わっている[2]。こ
れまでRA の病態において、腫瘍壊死因子(tumor necrosis factor; TNFα)や
インターロイキン 17(interleukin-17; IL-17)などが滑膜線維芽細胞や滑膜細 胞を活性化して軟骨や骨破壊を引き起こすことからTh1 細胞や Th17 細胞が病 態に関与していることが示唆されている[1, 3-5]。特に Th17 細胞はコラーゲン 誘導関節炎モデルマウスの病態形成を担う細胞として注目されており[3]、滑液 や滑膜組織においては、健常者に比べてIL-17+CD4+細胞(Th17 細胞)が増加 しているという報告[6-8]や逆に減少していると報告[9, 10]もされている。また、
Treg 細胞については、IL-10 やトランスフォーミング増殖因子 β(transforming growth factor β; TGF β)の産生を介して末梢の effector T 細胞などの抑制によ
る自己免疫の制御に関わり[11, 12]、RA においても炎症関節に存在しているが、
TNF αによる負の制御により Treg 細胞が減少し、RA の病態が促進されている
可能性が示唆されている[1, 13-16]。このように CD4 陽性 T 細胞の RA の病態
近年の多くの研究により、CD4 陽性 T 細胞においてマスター転写因子発現や サイトカイン産生などの可塑性が報告され、一つの転写因子やサイトカインで CD4 陽性 T 細胞を定義することが困難であることが示唆されている[2]。また、 これまでの研究ではCD4 陽性 T 細胞を含めた免疫細胞の定義が研究間で異なり、 結果のばらつきが生じていた可能性が示唆されている[17]。そこでより真の生物 学的事象を捉えるために、フローサイトメトリー(flow cytometry;FCM)を 用いて細胞表面マーカーの発現によりCD4 陽性 T 細胞を分類、標準化すること
で研究間のtechnical artifact をなくすような“human immunology project”が
提唱されてきている [17, 18]。RA においても細胞表面マーカーを用いた FCM の解析が多く報告されている[19, 20]。Lurati らは無治療 RA 患者における末梢 血中のCD4+CXCR3-CCR6+の Th 細胞が HC や OA 患者に比べて割合が増加し ているという報告し[19]、Rao らは抗 CCP 抗体陽性の RA 患者の滑液中で活性 化した T 細胞である PD-1highCXCR5-CD4+細胞が抗 CCP 抗体陰性の患者に比 べて有意に増加していたという報告をしている[20]。また Treg 細胞では
CD4+CD25+CD127lowのTreg 細胞が抗 IL-6受容体抗体製剤であるトシリズマ
ブでの治療により有意に治療前に比べて増加したといった報告もされている [21]。 このようにFCM を用いた手法での Th 細胞を含めた CD4 陽性 T 細胞の解析 は多く報告されているものの、Th 細胞の RA の病態への関与については未だ不 明な点が多く、特にTh 細胞の転写因子に関する報告は少ない。マウスの関節炎 モデルでは、Th 細胞のマスター転写因子である T-bet や GATA3、RORγt をそ れぞれ過剰発現させたトランスジェニックマウスにおいて有意に関節炎が抑制
されたことが報告されているが[22-24]、ヒトにおいては無治療の RA 患者にお いて RORγt の発現が亢進しているといった報告[25, 26]しかないのが現状で、 Th 細胞の転写因子の RA の病態への関与は不明である。そこで本研究では、近 年提唱されているFCM を用いた細胞表面マーカーによる Th 細胞の分類を行い、 その転写因子の発現がRA の病態へ及ぼす影響について解析することとした。 第 2 章 患者と方法 2-1 患者 本研究では、2014 年 11 月から 2017 年 3 月の間に筑波大学附属病院において 診療を受けた RA 患者 19 人、OA 患者 12 人について、登録、解析を行った。 HC は公募により本研究への協力が得られ、かつ RA 症例および OA 症例と年齢、 性別を合わせた 16 人について、登録、解析を行った。RA 患者は 1987 年の米 国リウマチ学会(ACR)の RA の分類基準、または 2010 年の ACR/ヨーロッパ リウマチ学会(EULAR)の RA 分類基準[27]を満たしており、発症半年未満の 早期のメトトレキサート(methotrexate ; MTX)やサラゾスルファピリジン (salazosulfapyridine ; SASP ) と い っ た 抗 リ ウ マ チ 薬 、 プ レ ド ニ ゾ ロ ン (prednisolone;PSL)を使用していない無治療の患者を登録した。全ての患者 において、年齢、性別、圧痛関節、腫脹関節や患者と医療者の全般的な健康状
態に対するVAS(visual analog scale)、臨床検査所見として CRP や ESR-1h、
MMP-3、RF、抗 CCP 抗体を測定し、疾患活動性の指標である DAS(disease activity score)28-CRP の評価を行なった。本研究は、全ての患者、HC よりイ ンフォームドコンセントを得た。本研究については、筑波大学附属病院の臨床
研究倫理審査委員会において審査され、承認を受けて実施した。
2-2 PBMC 及び SFMC 中の CD4 陽性 T 細胞の単離
PBMC や SFMC は被験者より得られた末梢血や滑液より Ficoll-Paque TM
Plus (GE Healthcare, Uppsala, Sweden)による密度勾配遠心分離で採取した。
得られた PBMC と SFMC は CD4+ MicroBeads (Milteny Biotec, Bergisch
Gladbach, Germany) を用いて磁気細胞分離(magnetic cell sorting;MACS)
により単離した。CD4 陽性細胞の純度は 95%以上の精度で得られた。
2-3 フローサイトメトリー(flow cytometry;FCM)
CD4 陽性 T 細胞は下記に示す抗体を用いて 30 分間氷上もしく室温などの適
切な条件で染色を行った:
anti-CCR6-Brilliant Ultraviolet 737 (11A9, BD HorizonTM, San Diego, CA, USA)
anti-CD45RA-APC-Cy7 (HI100, BD Pharmingen; San Diego, CA, USA)
anti- CXCR3-Alexa Fluor (AF) 700 (1C6/CXCR3, BD Pharmingen; San Diego, CA, USA)
anti-CXCR5-AF488 (RF8B2, BD Pharmingen; San Diego, CA, USA)
ま た CD4 陽 性 T 細 胞 の 転 写 因 子 は regulatory T Cell Staining Kit
(eBioscience; San Diego, CA)を用いて下記のような抗体を用いて染色を行っ
anti-T-bet-AF647 (4B10, Pharmingen)
anti-GATA3-PE-Cy7 (L50-823, Pharmingen) anti-RORγt-PE (Q21-559, Pharmingen)
FCM の解析、ソーティングは FACSAriaTM 2 cell sorter (BD, New Jersey, US)
を用いて行った。 FCM のデータの解析は FACSAriaTM 2 cell sorter の
FACSDiva software を用いて行った。 本研究では、過去の報告を用いてTh 細胞サブセットを定義した[12, 13]。FCM において、MACS で単離された CD4 陽性 T 細胞にリンパ球ゲートをかけた後、 下記のように定義した: CXCR5-/CD45RA-;Th 細胞 CXCR5-/CD45RA-/CXCR3+/CCR6-;Th1 細胞 CXCR5-/CD45RA-/CXCR3-/CCR6-;Th2 細胞 CXCR5-/CD45RA-/CXCR3-/CCR6+;Th17 細胞 CXCR5-/CD45RA-/CXCR3+/CCR6+; CXCR3+/CCR6+細胞 2-4 サイトカインとケモカインの測定
Th 細胞サブセットは FACSAriaTM 2 cell sorter を用いてソーティングを行い、
それぞれ 2×104 個の細胞を抗 CD3 抗体(BioLegend)と抗 CD28 抗体
(BioLegend)存在下の培養液(RPMI 1640 溶媒 (Sigma-Aldrich, USA) 10%
FBS 含有, ペニシリン(100 U/mL) + ストレプトマイシン(100μg/mL)500μl、 2-メルカゾール 50μl)で 96 時間培養した上清を cytometric bead array (CBA)
し た 。 ま た 血 清 、 滑 液 中 の CXCL9 及 び CXCL10 、 CCL20 に つ い て は LEGENDplex (BioLegend)を用いて測定した。
2-5 遊走能解析
Th 細胞の遊走能を Transwell system(24 wells, insert pore size 5 µm、
Corning, NY)を用いて評価した。MACS により単離された PBMC 中の CD4
陽性T 細胞(1×105個)は表面マーカー(CXCR5、CD45RA、CXCR3、CCR6)
の染色を行なった後に上層の well の 100 µl の培養液(RPMI 1640 溶媒
(Sigma-Aldrich, USA) 10% FBS 含有, ペニシリン(100 U/mL) + ストレプトマ
イシン(100 µg/mL)500 µl、2-メルカゾール 50 µl)へ添加し、4時間 37℃で 培養し、下層のwell の培養液 600 µl(上層と同様の培養液を用いてリコンビナ ントのヒトケモカイン(CXCL9、CXCL10、CCL20)(BioLegend))を 100 ng/ml の濃度で調整したものとケモカインを含まないもの)へ遊走した Th 細胞を FCM によって細胞数をカウントした。遊走指数(migration index)について は、ケモカインを含んだwell 中の細胞数とケモカインを含んでいない well の細 胞数の比により算出した。 2-6 統計学的解析 データは平均±標準偏差で示した。HC 及び OA 患者、RA 患者の年齢、性別 はそれぞれKruskal-Wallis 検定、Fisher 正確確率検定で統計学的解析を行った。 平 均 の 比 較 は 3 群 間 で は Kruskal-Wallis 検 定 、 PBMC と SFMC で は Mann-Whitney U 検定、治療前後では Wilcoxon 符号順位検定を用いて統計学
的解析を行った。また、相関解析については、Spearman 順位相関係数を用い
て統計学的解析を行った。いずれも統計学的な有意性を P 値が 0.05 未満とし、
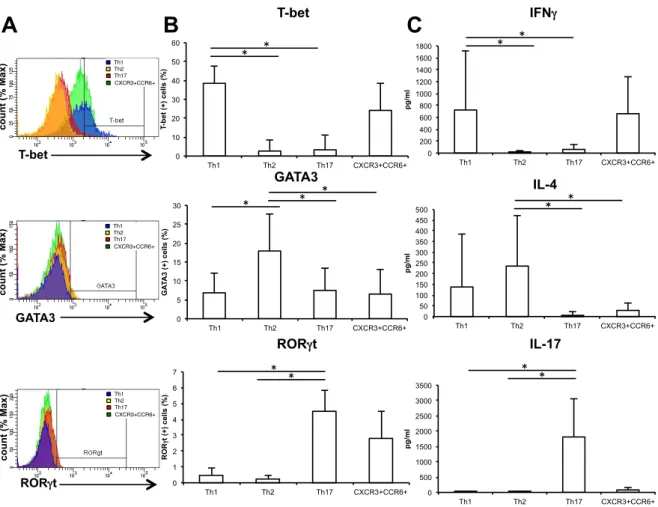
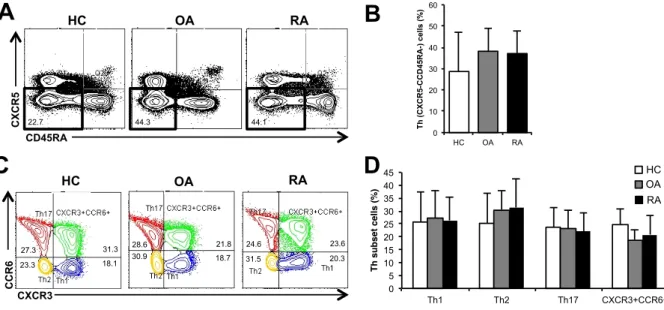
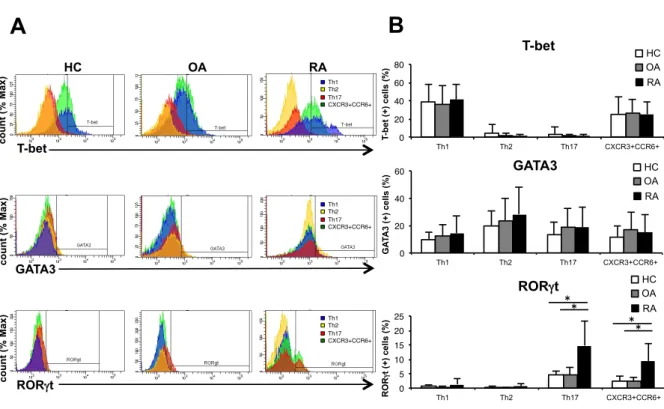
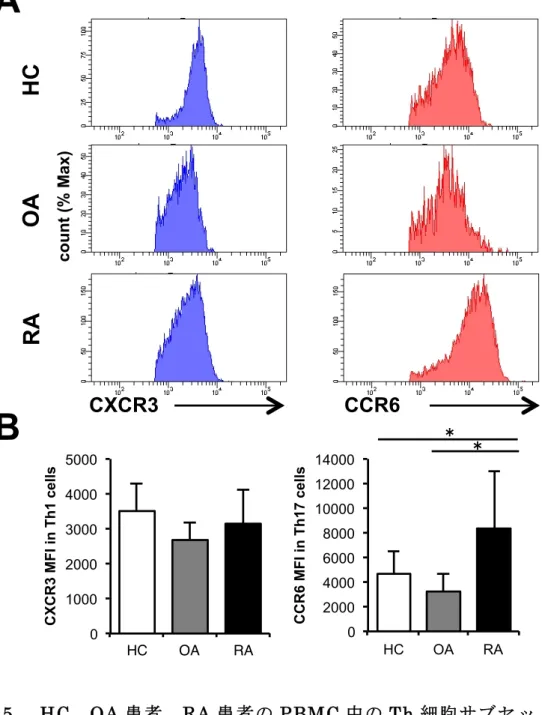
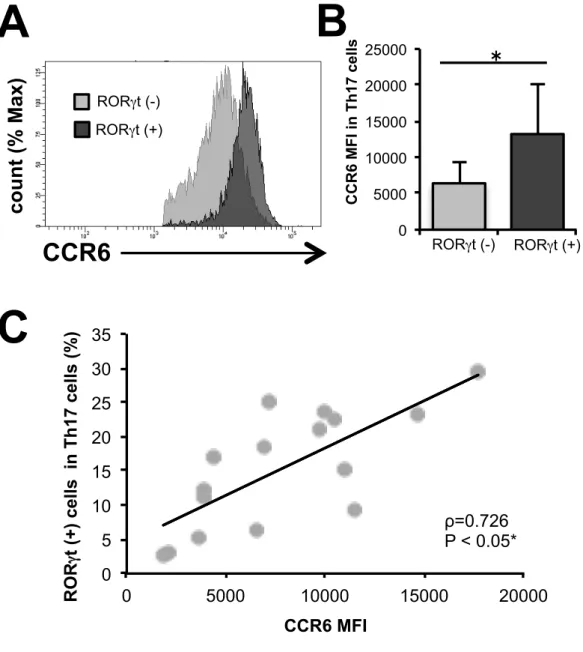
第 3 章 結果 3-1 HC の Th 細胞の細胞表面マーカー、転写因子とサイトカインの解 析 対象症例の背景や臨床データは、表1 に示す。サイトカイン測定が行えた HC (N=7)における PBMC 中の CD4 陽性細胞について、過去の報告を元に細胞 表面マーカーのCXCR5、CD45RA、CXCR3、CCR6 を用いて FCM で Th サブ セットの分類をした(図 1)。これらの分類が適切なものかを確かめるために、 転写因子のT-bet、GATA3、RORγt やサイトカインの IFNγ、IL-4、IL-17 を測 定したところ、それぞれのTh サブセットで特異的な転写因子、サイトカインの 発現が確認できた(図2)。 これらのデータから本研究の細胞表面マーカーによる分類は適切なものであ ると考えられた。 3-2 RA 患者、HC、OA 患者の Th 細胞の細胞表面マーカー、転写因子 とサイトカインの解析 3-1 と同様に RA 患者と OA 患者についても解析を行った。Th サブセットの 割合については 3 群間で有意な差は認めなかったが(図 3)、RA 患者の Th17 細胞のRORγt 発現が HC と OA 患者よりも有意に亢進していた(図 4)。さらに は細胞表面マーカーの発現については、RA 患者の Th17 細胞における CCR6 発 現が HC と OA 患者よりも有意に亢進していた(図 5)。また、Th17 細胞にお ける CCR6 と RORγt 発現の相関については、有意な正の相関を示し、RORγt 陽性細胞は陰性細胞に比べ有意にCCR6 の発現が亢進していた(図 6)。
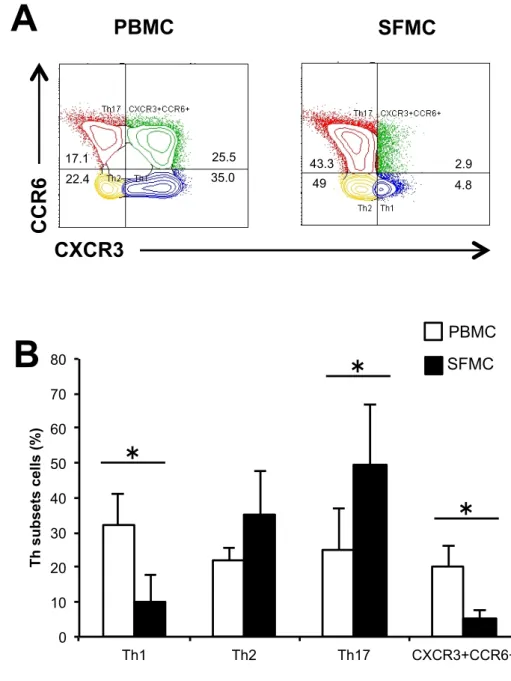
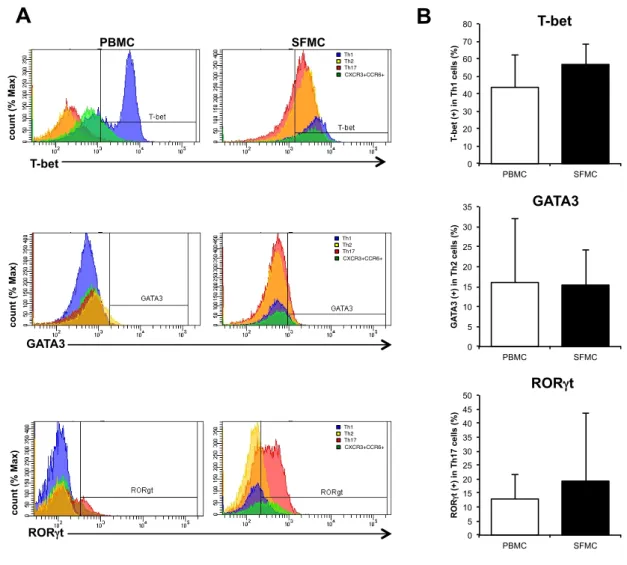
次に転写因子の発現の変化が Th 細胞のサイトカイン産生能に影響するかを 明らかにするために、分離した各Th サブセットを抗 CD3 抗体、抗 CD28 抗体 存在下で培養し、上清中のIFNγ、IL-4、IL-17 を CBA で測定した。3 群間でい ずれのTh 細胞サブセットにおいても、産生されるサイトカイン量については有 意な差は認めなかった(図7)。 これらの結果からRA 患者の PBMC における Th17 細胞では CCR6 と RORγt 発現の亢進があり、病態への関与が示唆された。 3-3 RA 患者の PBMC と SFMC 中の Th 細胞の比較 RA の病態の主座である炎症関節での Th 細胞の解析を行うために、PBMC と SFMC 中の Th 細胞について比較した。3-1、2 と同様に Th 細胞サブセットの 割合についてFCM で解析したところ、SFMC 中の Th17 細胞の割合が PBMC に比べて有意に増加し、Th1 細胞が有意に減少していることがわかった(図 8)。 また、転写因子の発現について比較したが、PBMC、SFMC 間で有意な差は認 められなかった(図9)。 これらの結果からTh17 細胞の炎症局所での増加が示唆された。 3-4 Th 細胞の遊走能の解析 RA 患者において Th17 細胞の CCR6 の発現の亢進と炎症関節での増加が見ら れることから、炎症局所への遊走能の亢進が示唆され、評価のためにtranswell
system による chemotaxis assay を行った。Th1 細胞については、特異的な細
CXCL9 や CXCL10 に対する遊走能は RA 患者、HC、OA 患者の 3 群間で有意 な差がなかった。一方で、Th17 細胞では CCR6 のリガンドである CCL20 に対 する遊走能がRA 患者で有意に HC、OA 患者よりも亢進していた(図 10)。ま た各リガンド特異的な遊走がCXCL9、CCL20 では見られたが、CXCL10 では 見られなかった(図10)。 上記リガンドの CXCL9、CXCL10、CCL20 について血清中と滑液中での濃 度を測定したところ、RA 患者の滑液中の CXCL10 と CCL20 濃度が血清中より も有意に増加し、その他HC の血清、OA 患者の血清と滑液よりも有意に増加し ていた(図11)。 これらの結果からTh17 細胞自身の遊走能の亢進と滑液中の CCL20 濃度の上 昇が炎症関節へのTh17 細胞の増加に寄与している可能性が考えられた。 3-5 RA 患者の治療前後の Th 細胞の細胞表面マーカー、転写因子の変化 MTX などの RA に対する治療による Th 細胞の細胞表面マーカーや転写因子 の発現の変化を解析するために、7 例について治療 24 週時と治療前とでの比較 を行った。治療として MTX や SASP、PSL などが用いられ、24 週時には RA の疾患活動性の指標である DAS28-CRP は有意な改善を認めた(図 12)。治療 前後での Th サブセットの割合について差が見られなかったが(図 13)、Th17 細胞におけるRORγt 発現は治療により有意に低下した(図 14)。さらには Th17 細胞のCCR6 発現についても治療により有意に低下した(図 15)。 また、1 例の報告になるが、治療前後の SFMC 中の Th 細胞について解析を行 ったところ、Th17 細胞は治療により割合の減少傾向を示し(図 16)、さらには
CCL20 の濃度の低下を認めた(図 17)。
これらの結果からTh17 細胞の CCR6 と RORγt 発現が病態に関与しているこ
第4 章 考察 近年 Th17 細胞の RA の病態への関与が多く報告されている。 Van Hamburg らは無治療RA 患者の PBMC 中の CCR6 陽性の IL-17 や TNF αを産生する Th17 細胞が増加していたと報告し[15]、Jimeno らは無治療 RA 患者において、 HC に比べて CCR6+RORC+細胞が増加し、T-bet +細胞が減少していることを 報告している[26]。さらに Arroyo らは抗 CCP 抗体陽性の無治療 RA 患者にお いて末梢血中のTh17 細胞の割合や Th17/Th1 細胞比が治療後に減少しているこ とを報告[28]するなど、末梢血中の Th17 細胞の増加を示す報告が多いが、本研 究では、RA 患者と HC、OA 患者間においては、Th 細胞サブセットの割合に有 意な差は認められなかった。この結果の違いの理由としては以下に示すような2 つの可能性が考えられた。1 つは患者背景の研究間での相違が挙げられる。多く の研究では治療を導入後や、特にRA の診断基準は満たすが抗 CCP 抗体陰性患 者を含むような不均一な集団で解析を行っているのに対し、本研究では発症半 年以内の早期の無治療かつ抗CCP 抗体陽性の患者のみ登録しており、より均質 な発症早期RA 患者群を捉えることができている。もう 1 つの理由としては、 Th 細胞の定義の違いが挙げられる。本研究は過去の報告と異なり、CD4 陽性細
胞を含めた免疫細胞の統一的な定義を図った“human immunology project“を
もとに得られたデータである。これらの患者背景、定義の違いがTh17 細胞の割
合の結果の相違に影響したと考えられた。また、本研究ではRA 患者の Th17
細胞におけるRORγt の発現の亢進が認められた。さらには RA 患者では Th17
細胞のCCR6 の発現が亢進し、RORγt と CCR6 の発現において有意な正の相関
する結果が得られた[29]。一方、IL-17 といった Th17 細胞から特徴的に産生さ れるサイトカインについては3 群間で有意な差はなかった。RA 患者のCCR6 の遺伝子多型が発症に関与している点[30]や抗 IL-17 抗体である Secukinumab や抗IL-17 受容体抗体である Brodalumab は RA には効果が乏しく開発中止 となっている点は[31, 32]、今回見出された RA 患者における Th17 細胞の RORγt の発現亢進はIL-17 産生ではなく CCR6 発現の亢進に寄与している結果と矛盾 せず、RORγt-CCR6 の経路がより強く RA の疾患機序に関与している可能性が 示唆された。 炎症の局所におけるTh 細胞については、RA の炎症関節の滑液中で末梢血に 比してTh17 細胞の割合が有意に増加していること[25, 33]や Th17 細胞の産生 するIL-17 が炎症関節において単球や好中球、滑膜線維芽細胞を活性化し、破 骨細胞の誘導、骨破壊に寄与していることなどが報告されている[34, 35]。本研 究においても、これらの報告と同様にPBMC 中よりも SFMC 中で Th17 細胞の 割合の増加を認めた。またTh1 細胞については、IFNγを介して破骨細胞を抑制 し、RA の病態に対しては抑制的な細胞であることが示唆されており[33, 34]、 本研究においても滑液中のTh1 細胞の割合が減少しており、この Th1 細胞の欠 如がRA の病態悪化の要因の一つになった可能性が考えられた。 細胞の遊走にはリガンドであるケモカインとその受容体であるケモカインレ セプターの反応が関与しており[38-40]、Th17 細胞はケモカインレセプターであ るCCR6 のリガンドである CCL20 に特異的に遊走することが報告されている [41-43]。RA の関節炎モデルマウスの SKG マウスにおいても、抗 CCR6 抗体を 投与することでTh17 細胞の遊走を抑さえられ、関節炎の発症や重症度を有意に
抑制することが報告されている[44]。また、過去の報告でもあるように[29, 45]、
本研究でもRA 患者の滑液中で著明な CCL20 の上昇を認め、近年 CCR6-CCL20
経路がRA の病態への関与が示唆されているが、RA 患者において Th17 細胞自
体の遊走能について調べた報告はない。そこで本研究では、Th 細胞の遊走能の
評価のために行ったchemotaxis assay を行ったところ、RA 患者の Th17 細胞
のCCL20 に対する遊走能の有意な亢進を認めた。これまでの結果から本研究に おけるTh17 細胞の炎症関節での集積については、RORγt の発現亢進に伴う CCR6 の増加と CCL20 の増加により惹起された可能性が示唆された。一方、Th1 細胞のケモカインレセプターであるCXCR3 のリガンドである CXCL10 も RA 患者の滑液において著明な増加を認めたが、SFMC 中の Th1 は末梢血と比較し て有意にその比率は低下していた。Th1 細胞は RA の病態改善に寄与している 可能性も報告されており[36, 37]、本研究における PBMC と比較した際の SFMC 中のTh1 の有意な比率の低下の原因には、Th1 細胞遊走を抑制する機序の存在、 もしくは炎症局所に移行したTh1 から Th17 への再分化の可能性が考慮される が、本研究では詳細を明らかにすることはできなかった。 本研究では、MTX などの治療開始前に比べ、治療 24 週時の Th17 細胞で CCR6 とRORγt 発現の有意な低下を認めた。過去に動物モデルや RA 患者における治 療によるCCR6 や RORγt 発現の変化を示した報告はないが、関節炎モデルマウ スにおけるCD4 陽性 T 細胞への影響については、T 細胞依存的に発症するプリ スタン誘導関節炎ラットにおいて、MTX 治療により Th1 や Th17 細胞を不活化 し、関節炎を抑制されたという報告[45]や Th17 細胞依存的に発症するコラーゲ ン誘導関節炎ラットにおいて、MTX 治療により CD4+CD25+の Treg 細胞の増
加が惹起され、関節炎が抑制されたといった報告がある[47]。また MTX は細胞 分化やアポトーシスを誘導すると報告されている[48, 49]。これらの報告から、 活性化されたTh17 細胞が MTX により不活化され、CCR6 や RORγt の発現が 低下した可能性が示唆された。さらに、本研究では治療前後の滑液中のケモカ インの濃度の測定を行えた1 症例において、MTX 治療 24 週後に CCL20 の低 下とCXCL10 の上昇を認め、Th 細胞の遊走を制御するケモカインの病態関与 も示唆された。 これまでにRA の病態における Th17 細胞さらには CCR6-CCL20 経路の重要 性が多く報告される中、本研究では、発症早期の無治療かつ抗CCP 抗体陽性の より純粋なRA 症例において、RORγt に伴う CCR6 の発現亢進が炎症局所への Th17 細胞の遊走を促進している可能性を示した。また、関節炎モデルマウスで のCCR6 の抑制により関節炎が改善した報告や本研究での抗リウマチ治療によ るRORγt-CCR6-CCL20 の発現低下を認める点から、RORγt-CCR6-CCL20 の 経路の制御がRA に対する新規治療戦略になる可能性を示しており、RA 以外の 炎症性関節炎を伴うような類似疾患(乾癬性関節炎など)においても今後の研 究を進めていくことが必要と考えられた。
第5 章 結論 RA においては、RORγt の発現亢進によってもたらされたと考えられる CCR6 の高発現と滑液中のCCL20 の増加が Th17 細胞の炎症関節への遊走を促進して いる可能性があり、また治療による疾患活動性の低下とともにTh17 細胞の CCR6 と RORγt の発現も有意に抑制されたことから RORγt-CCR6-CCL20 経路 が関節炎の病態に深く関与している可能性が示唆された。
参考文献
1. McInnes IB, Schett G: The pathogenesis of rheumatoid arthritis. N Engl J Med. 2011; 365:2205-19.
2. Nakayamada S, Takahashi H, Kanno Y, O'Shea JJ: Helper T cell diversity and plasticity. Curr Opin Immunol. 2012; 24: 297-302. 3. Lubberts E, Koenders MI, Oppers-Walgrenn B, van den Bersselaar L,
Coenen-de Roo CJ. Joosten LA, et al. Treatment with a neutralizing anti-murine interleukin-17 antibody after the onset of collagen-induced arthritis reduces joint inflammation, cartilage destruction, and bone erosion. Arthritis Rheum 2004;50:650-9.
4. Benedetti G, Miossec P. Interleukin 17 contributes to the chronicity of inflammatory diseases such as rheumatoid arthritis. Eur J Immunol. 2014;44:339-47.
5. Carrión M, Pérez-García S, Jimeno R, Juarranz Y, González-Álvaro I, Pablos JL, et al. Inflammatory mediators alter IL-17 receptors, IL-12 and IL-23 expression in human osteoarthritic and rheumatoid arthritis
sinovial fibroblast: immunomodulation by vasoactive intestinal peptide. Neuroimmunomodulation. 2013;20:274-84.
6. Li N, Wang JC, Liang TH, Zhu MH, Wang JY, Fu XL, et al. Pathologic finding of increased expression of interleukin-17 in the synovial tissue of rheumatoid arthritis patients. Int J Clin Exp Pathol. 2013; 6:1375-9.
7. Chabaud M, Durand JM, Buchs N, Fossiez F, Page G, Frappart L, et al. Human interleukin-17: a T cell-derived proinflammatory cytokine
produced by the rheumatoid synovium. Arthritis Rheum. 1999;42:963-70. 8. Moran EM, Mullan R, McCormick J, Connolly M, Sullivan O, Fitzgerald
O, et al. Human rheumatoid arthritis tissue production of IL-17A drives matrix and cartilage degradation: synergy with tumour necrosis
factor-alpha, Oncostatin M and response to biologic therapies. Arthritis Res Ther. 2009;11:R113.
9. Yamada H, Nakashima Y, Okazaki K, Mawatari T, Fukushi JI, Kaibara N, et al. Th1 but not Th17 cells predominate in the joints of patients with rheumatoid arthritis. Ann Rheum Dis. 2008;67:1299-1304.
10. Ito Y, Usui T, Kobayashi S, Iguchi-Hashimoto M, Ito H, Yoshitomi H, et al. Gamma/delta T cells are the predominant source of interleukin-17 in affected joints in collagen-induced arthritis, but not in rheumatoid arthritis. Arthritis Rheum. 2009;60:2294-303.
11. Sakaguchi S. Naturally arising CD4+ regulatory T cells for immunologic self-tolerance and negative control of immune responses. Annu Rev Immunol. 2004;22:531-62.
12. Shevach EM. Regulatory/suppressor T cells in health and disease. Arthritis Rheum. 2004;50:2721-4.
13. Hong Nie, Yingxia Zheng, Runsheng Li, Taylor B Guo, Dongyi He, Lei Fang, et al. Phosphorylation of FOXP3 controls regulatory T cell function and is inhibited by TNF-α in rheumatoid arthritis. Nat Med. 2013;19:322-8.
14. Walter GJ, Evans HG, Menon B, Gullick NJ, Kirkham BW, Cope AP, et al. Interaction with activated monocytes enhances cytokine expression and suppressive activity of human CD4+CD45RO+CD25+CD127low regulatory T cells. Arthritis Rheum. 2013;65:627-38.
15. Benito-Miguel M, García-Carmona Y, Balsa A, Pérez de Ayala C, Cobo-Ibáñez T, Martín-Mola E, et al. A dual action of rheumatoid arthritis synovial fibroblast IL-15 expression on the equilibrium between CD4+CD25+ regulatory T cells and CD4+CD25- responder T cells. J Immunol. 2009;183:8268-79.
16. Cao D, Malmstrom V, Baecher-Allan C, Hafler D, Klareskog L, Trollmo C.
Isolation and functional characterization of regulatory CD25brightCD4+ T
cells from the target organ of patients with rheumatoid arthritis. Eur J Immunol. 2003;33:215-223.
17. Maecker HT, McCoy JP, Nussenblatt R. Standardizing
immunophenotyping for the Human Immunology Project. Nat Rev Immunol. 2012;12:191–200.
L, Zurawski G, et al. Human blood CXCR5(+)CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity. 2011;34:108-21. 19. Lurati A, Laria A, Gatti A, Brando B, Scarpellini M, et al. Different T
cells' distribution and activation degree of Th17 CD4+ cells in peripheral blood in patients with osteoarthritis, rheumatoid arthritis, and healthy donors: preliminary results of the MAGENTA CLICAO study. Open Access Rheumatol. 2015;7:63-8.
20. Rao DA, Gurish MF, Marshall JL, Slowikowski K, Fonseka CY, Liu Y, et al. Pathologically expanded peripheral T helper cell subset drives B cells in rheumatoid arthritis. Nature. 2017;542:110-4
21. Kikuchi J, Hashizume M, Kaneko Y, Yoshimoto K, Nishina N, Takeuchi T. Peripheral blood CD4+CD25+CD127low regulatory T cells are
significantly increased by tocilizumab treatment in patients with rheumatoid arthritis: increase in regulatory T cells correlates with clinical response. Arthritis Res Ther. 2015;17:10.
22. Kondo Y, Iizuka M, Wakamatsu E, Yao Z, Tahara M, Tsuboi H, et al. Overexpression of T-bet gene regulates murine autoimmune arthritis. Arthritis Rheum. 2012;64:162-72.
23. van Hamburg JP, Mus AM, de Bruijn MJ, de Vogel L, Boon L, Cornelissen F, et al. GATA-3 protects against severe joint inflammation
and bone erosion and reduces differentiation of Th17 cells during experimental arthritis. Arthritis Rheum. 2009;60:750-9.
24. Kondo Y, Yao Z, Tahara M, Iizuka M, Yokosawa M, Kaneko S, et al. Involvement of RORγt-overexpressing T cells in the development of autoimmune arthritis in mice. Arthritis Res Ther. 2015;17:105.
25. Leipe J, Grunke M, Dechant C, Reindl C, Kerzendorf U, Schulze-Koops H, et al. Role of Th17 cells in human autoimmune arthritis. Arthritis Rheum. 2010;62:2876-85.
26. Jimeno R, Gomariz RP, Garín M, Gutiérrez-Cañas I, González-Álvaro I, Carrión M, et al. The pathogenic Th profile of human activated memory Th cells in early rheumatoid arthritis can be modulated by VIP. J Mol Med. 2015; 93:457-67.
27. Aletaha D, Neogi T, Silman AJ, Funovits J, Felson DT, Bingham 3rd CO, et al. 2010 rheumatoid arthritis classification criteria: an American College of Rheumatology/European League Against Rheumatism collaborative initiative. Ann Rheum Dis. 2010;69:1580-8.
28. Arroyo-Villa I, Bautista-Caro MB, Balsa A, Aguado-Acín P, Nuño L, Bonilla-Hernán MG, et al. Frequency of Th17 CD4+ T cells in early rheumatoid arthritis: a marker of anti-CCP seropositivity. PLoS One. 2012;7:e42189.
29. Hirota K, Yoshitomi H, Hashimoto M, Maeda S, Teradaira S, Sugimoto N, et al. Preferential recruitment of CCR6-expressing Th17 cells to in amed joints via CCL20 in rheumatoid arthritis and its animal model. J Exp Med. 2007;204:2803-12.
30. Stahl EA, Raychaudhuri S, Remmers EF, Xie G, Eyre S, Thomson BP, et al. Genome-wide association study meta-analysis identifies seven new rheumatoid arthritis risk loci. Nat Genet. 2010;42:508-14.
31. Genovese MC, Durez P, Richards HB, et al: Efficacy and safety of
secukinumab in patients with rheumatoid arthritis: a phaseⅡ,
dose-finding, double-blind, randomised, placebo controlled study. Ann Rheum Dis, 72:863-869, 2013.
32. Pavelka K, Chon Y, Newmark R, et al: A study to evaluate the safety, tolerability, and efficacy of brodalumab in subjects with rheumatoid arthritis and an inadequate response to methotrexate. J Rheumatol, 42:912-919, 2015.
33. Nistala K, Moncrieffe H, Newton KR, Varsani H, Hunter P, Wedderburn LR. Interleukin-17-producing T cells are enriched in the joints of children with arthritis, but have a reciprocal relationship to regulatory T cell numbers. Arthritis Rheum. 2008;58:875-87.
34. Brennan FM, McInnes IB. Evidence that cytokines play a role in rheumatoid arthritis. J Clin Invest. 2008;118:3537-45.
35. Choy EH, Panayi GS. Cytokine pathways and joint inflammation in rheumatoid arthritis. N Engl J Med. 2001;344:907-16.
36. H. Takayanagi, S. Kim, and T. Taniguchi. Signaling crosstalk between RANKL and interferons in osteoclast di erentiation. Arthritis Res. 2002;4:S227-32.
37. Kotake S, Nanke Y, Yago T, Kawamoto M, Yamanaka H. Human osteoclastogenic T cells and human osteoclastology. Arthritis Rheum. 2009;60:3158-63.
38. Charo IF, Ransohoff RM. The many roles of chemokines and chemokine receptors in inflammation. N Engl J Med. 2006;354:610-21.
39. Rollins BJ. Chemokines. Blood. 1997;90:909-28.
40. Sallusto F, Mackay CR, Lanzavesschi A. The role of chemokine receptors in primary, effector and memory immune responses. Annu Rev Immunol. 2000;18:593-620.
41. Baba M, Imai T, Nishimura M, Kakizaki M, Takagi S, Hieshima K, et al. Identification of CCR6, the specific receptor for a novel
lymphocyte-directed CC chemokine LARC. J Biol Chem. 1997;272:14893-8.
42. Liao F, Alderson R, Su J, Ullrich SJ, Kreider BL, Farber JM. STRL22 is a receptor for the CC chemokine MIP-3-alpha. Biochem Biophys Res
Commun. 1997;236:212-7.
43. Charbonnier AS, Kohrgruber N, Kriehuber E, Stingl G, Rot A, Maurer D. Macrophage inflammatory protein 3-alpha is involved in the constitutive trafficking of epidermal Langerhans cells. J Exp Med. 1999;190:1755-68. 44. Hirota K, Yoshitomi H, Hashimoto M, Maeda S, Teradaira S, Sugimoto N,
et al. Preferential recruitment of CCR6-expressing Th17 cells to inflamed joints via CCL20 in rheumatoid arthritis and its animal model. J Exp Med. 2007;204:2803-12.
45. Kawashiri SY, Kawakami A, Iwamoto N, Fujikawa K, Aramaki T, Tamai M, et al. Proinflammatory cytokines synergistically enhance the
production of chemokine ligand 20 (CCL20) from rheumatoid
fibroblast-like synovial cells in vitro and serum CCL20 is reduced in vivo by biologic disease-modifying antirheumatic drugs. J Rheumatol.
2009;36:2397-402.
46. Hou WK, Meng LS, Zheng F, Wen YR, Zhu WH, Jiang CS, et al.
Methotrexate ameliorates pristane-induced arthritis by decreasing IFN-γ and IL-17A expressions. J Zhejiang Univ Sci B. 2011;12:40-6.
47. Xinqiang S, Fei L, Nan L, Yuan L, Fang Y, Hong X, et al. Therapeutic efficacy of experimental rheumatoid arthritis with low-dose methotrexate by increasing partially CD4+CD25+ Treg cells and inducing Th1 to Th2 shift in both cells and cytokines. Biomed Pharmacother. 2010;64:463-71. 48. de Lathouder S, Gerards AH, Dijkmans BA, Aarden LA. Two inhibitors of
DNA-synthesis lead to inhibition of cytokine production via a different mechanism. Nucleosides Nucleotides Nucleic Acids. 2004;23:1089-100. 49. Wessels JA, Huizinga TW, Guchelaar HJ. Recent insights in the
pharmacological actions of meth- otrexate in the treatment of rheumatoid arthritis. Rheumatology (Oxford). 2007;47:249-55.
HC (N=16) OA (N=12) 無治療RA(N=19) p value 年齢 51.3 ± 10.5 55.7 ± 10.9 53.8 ± 18.5 0.61 女性, n (%) 13 (81.3) 8 (66.7) 16 (84.2) 0.61 CRP (mg/dl) - - 1.82 ± 3.03 - ESR-1h (mm) - - 26.4 ± 16.5 - MMP-3 (ng/ml) - - 187 ± 252 - RF (U/ml) - - 99.0 ± 67.8 - 抗CCP抗体, n (%) - - 19 (100) - DAS28-CRP - - 4.0 ± 1.03 - DAS28-ESR - - 4.46 ± 1.13 - CDAI - - 17.1 ± 7.71 - SDAI - - 19.4 ± 8.71 - 表 1 HC、OA 患者、RA 患者の背景と臨床データ
anti CCP antibody, anti cyclic citrullinated peptide antibody; CDAI, clinical disease activity index; CRP, c-reactive protein; ESR, erythrocyte sedimentation rate; MMP-3, matrix metalloproteinase-3; RF, rheumatoid factor; SDAI,
simplified disease activity index; (*P < 0.05、平均 ± 標準偏差、年齢: Kruskal-Wallis 検定、性別:Fisher 正確確率検定)
図 1. HC の PBMC 中の Th 細胞サブセット HC(N=7)の Th 細胞サブセットについて FCM で解析を行った。(A)(左) リンパ球ゲートをかけた後のTh 細胞群(黒枠:CD45RA-CXCR5-細胞)。(右) Th 細胞を CXCR3 と CCR6 により 4 つの細胞群に分けられた Th 細胞サブセッ ト。(B)各 Th 細胞サブセットの割合。(*P < 0.05、平均 ± 標準偏差、 Kruskal-Wallis 検定)
A
CCR6 CXCR3 CD45RA C XC R 5B
CXCR5- CD45RA- gated 27.8 27.1 22.7 22.5 22.7 Lymphocyte gated 0 5 10 15 20 25 30 35 40 Th1 Th2 Th17 CXCR3+CCR6+ T h s u b se t ce lls (% )図 2. HC の PBMC 中の Th 細胞サブセットの転写因子・サイトカイン HC(N=7)の Th 細胞サブセットの転写因子、サイトカインについて FCM で 解析を行った。(A)Th 細胞サブセットの転写因子(T-bet、GATA3、RORγt) の代表的なヒストグラム。(B)Th 細胞サブセットの転写因子(T-bet、GATA3、 RORγt)を発現する細胞の割合の平均の比較。(C)Th 細胞サブセットのサイ トカインの平均の比較。各Th 細胞サブセットを FCM により sorting 後、2× 104個の細胞を抗CD3/28 抗体存在下で 96 時間培養し、上清を CBA によりサイ トカイン(IFNγ、IL-4、IL-17)を測定した。(*P < 0.05、平均 ± 標準偏差、 Kruskal-Wallis 検定) 0 10 20 30 40 50 60 Th1 Th2 Th17 CXCR3+CCR6+ T-b et (+) ce lls (% ) 0 5 10 15 20 25 30 Th1 Th2 Th17 CXCR3+CCR6+ G A TA 3 (+) ce lls (% ) 0 1 2 3 4 5 6 7 Th1 Th2 Th17 CXCR3+CCR6+ ROR γt (+) ce lls (% ) 0 50 100 150 200 250 300 350 400 450 500 Th1 Th2 Th17 CXCR3+CCR6+ pg/ml 0 200 400 600 800 1000 1200 1400 1600 1800 Th1 Th2 Th17 CXCR3+CCR6+ pg/ml 0 500 1000 1500 2000 2500 3000 3500 Th1 Th2 Th17 CXCR3+CCR6+ pg/ml T-bet GATA3 RORγt IFNγ IL-4 IL-17
A
T-bet GATA3 RORγtB
C
co u n t (% Ma x) co u n t (% Ma x) co u n t (% Ma x) Isotype Th1 Th2 Th17 CXCR3+CCR6+ Isotype Th1 Th2 Th17 CXCR3+CCR6+ Isotype Th1 Th2 Th17 CXCR3+CCR6+図 3. HC、OA 患者、RA 患者の PBMC 中の Th 細胞サブセット HC(N=16)、OA患者(N=12)、RA患者(N=19)のTh細胞サブセットにつ いてFCMで解析を行った。(A)リンパ球ゲートをかけた後のTh細胞群(黒枠: CD45RA-CXCR5-細胞)(B)Th細胞の割合の平均の比較。(C)Th細胞をCXCR3 とCCR6により4つの細胞群に分けられたTh細胞サブセット。(D)各Th細胞サ ブセットの割合の平均の比較(*P < 0.05、平均 ± 標準偏差、Kruskal-Wallis 検定) RA
A
CD45RA C XC R 5B
C
D
HC RA HC CCR6 CXCR3 27.3 31.3 23.3 18.1 24.6 23.6 31.5 20.3 RA HC OA 22.7 44.3 44.1 28.6 21.8 30.9 18.7 OA OA 0 10 20 30 40 50 60 HC OA RA T h (C XC R 5-C C D 45 R A -) ce lls (% ) 0 5 10 15 20 25 30 35 40 45 Th1 Th2 Th17 CXCR3+CCR6+ T h s u b s e t c e ll s (% )図 4. HC、OA 患者、RA 患者の PBMC 中の Th 細胞サブセットの転写 因子 HC(N=16)、OA 患者(N=12)、RA 患者(N=19)の Th 細胞サブセットの 転写因子についてFCM で解析を行った。(A)Th 細胞サブセットの転写因子 (T-bet、GATA3、RORγt)の代表的なヒストグラム。(B)Th 細胞サブセッ トの転写因子(T-bet、GATA3、RORγt)を発現する細胞の割合の平均の比較。 (*P < 0.05、平均 ± 標準偏差、Kruskal-Wallis 検定) T-bet GATA3 RORγt HC RA
A
T-bet GATA3 RORγt OA RA HC OA RA HC OA RA HC OA co u n t (% Ma x)B
0 20 40 60 80 Th1 Th2 Th17 CXCR3+CCR6+ T-b et (+) ce lls (% ) 0 20 40 60 Th1 Th2 Th17 CXCR3+CCR6+ G A TA 3 (+) ce lls (% ) 0 5 10 15 20 25 Th1 Th2 Th17 CXCR3+CCR6+ ROR γt (+) ce lls (% ) co u n t (% Ma x) co u n t (% Ma x) Th1 Th2 Th17 CXCR3+CCR6+ Th1 Th2 Th17 CXCR3+CCR6+ Th1 Th2 Th17 CXCR3+CCR6+図 5. HC、OA 患者、RA 患者の PBMC 中の Th 細胞サブセットのケモ カインレセプター HC(N=16)、OA 患者(N=12)、RA 患者(N=19)の Th 細胞サブセットの ケモカインセレプターについてFCM で解析を行った。(A)(左)Th1 細胞に おけるCXCR3 の代表的なヒストグラム。(右)Th17 細胞における CCR6 の代 表的なヒストグラム。(B)(左)Th1 細胞における CXCR3 の平均蛍光強度(MFI) の平均の比較。(右)Th17 細胞における CCR6 の MFI の平均の比較。(*P < 0.05、平均 ± 標準偏差、Kruskal-Wallis 検定)
B
0 1000 2000 3000 4000 5000 HC OA RA C XC R 3 MF I i n T h 1 ce lls 0 2000 4000 6000 8000 10000 12000 14000 HC OA RA C C R 6 MF I i n T h 17 c el lsCXCR3
CCR6
co u n t (% Ma x)HC
OA
RA
A
図 6. RA 患者の PBMC 中の Th17 細胞における CCR6 と RORγt 発現 (A)代表的な Th17 細胞における RORγt 陰性、陽性細胞における CCR6 のヒ ストグラム。(B)Th17 細胞における RORγt 陰性、陽性細胞における CCR6 のMFI。の平均の比較。(C)RA 患者(N=19)の Th17 細胞の CCR6 の MFI とRORγt 陽性細胞の割合の相関について解析した。(*:P < 0.05、平均の比 較:Mann-Whitney U 検定、ρ:Spearman 順位相関係数) 0 5000 10000 15000 20000 25000 RORgt (-) RORgt (+) C C R 6 MF I i n T h 17 c el ls 0 5 10 15 20 25 30 35 0 5000 10000 15000 20000 ROR γt (+) ce lls i n T h 17 c el ls (% ) CCR6 MFI
C
ρ=0.726 P < 0.05*CCR6
co
u
n
t
(%
Ma
x)
A
B
RORγt (-) RORγt (+) RORγt (-) RORγt (+)図 7. HC、OA 患者、RA 患者の PBMC 中の Th 細胞サブセットのサイ トカイン HC(N=7)、OA 患者(N=5)、RA 患者(N=7)の Th 細胞サブセットのサイ トカイン(IFNγ、IL-4、IL-17)について CBA で解析を行った。各 Th 細胞サ ブセットをFCM により sorting 後、2×104個の細胞を抗CD3/28 抗体存在下で 96 時間培養し、上清を CBA によりサイトカイン(IFNγ、IL-4、IL-17)を測定 した。(*P < 0.05、平均 ± 標準偏差、Kruskal-Wallis 検定) 0 1000 2000 3000 4000 5000 Th1 Th2 Th17 CXCR3+CCR6+ pg/ml
IFNγ
RA
HC
OA
0 100 200 300 400 500 Th1 Th2 Th17 CXCR3+CCR6+ pg/mlIL-4
RA
HC
OA
0 1000 2000 3000 4000 5000 6000 7000 8000 Th1 Th2 Th17 CXCR3+CCR6+ pg/mlIL-17
RA
HC
OA
図 8. RA 患者の PBMC と SFMC の Th 細胞サブセット RA 患者(N=4)の PBMC と SFMC 中の Th 細胞サブセットについて FCM で 解析を行った。(A)Th 細胞を CXCR3 と CCR6 により 4 つの細胞群に分けら れたTh 細胞サブセット。(B)各 Th 細胞サブセットの割合の平均の比較(* P < 0.05、平均 ± 標準偏差、Mann Whitney U 検定) 0 10 20 30 40 50 60 70 80 Th1 Th2 Th17 CXCR3+CCR6+ T h s u b se ts c el ls (% )
B
SFMC PBMCCXCR3
CCR6
PBMC
SFMC
A
17.1 25.5 22.4 35.0 43.349 2.94.8図 9. RA 患者の PBMC と SFMC の Th 細胞サブセットの転写因子 RA 患者(N=4)の PBMC と SFMC 中の Th 細胞サブセットの転写因子につい てFCM で解析を行った。(A)Th 細胞サブセットの転写因子(T-bet、GATA3、 RORγt)の代表的なヒストグラム。(B)Th 細胞サブセットの転写因子(T-bet、 GATA3、RORγt)を発現する細胞の割合の平均の比較。(*P < 0.05、平均 ± 標 準偏差、Mann Whitney U 検定) 0 10 20 30 40 50 60 70 80 PBMC SFMC T-b et (+) in T h 1 ce lls (% ) 0 5 10 15 20 25 30 35 PBMC SFMC G A TA 3 (+) in T h 2 ce lls (% ) 0 5 10 15 20 25 30 35 40 45 50 PBMC SFMC ROR γt (+) in T h 17 c el ls (% ) T-bet GATA3 RORγt PBMC SFMC
A
B
T-bet GATA3 RORγt co u n t (% Ma x) co u n t (% Ma x) co u n t (% Ma x) Th1 Th2 Th17 CXCR3+CCR6+ Th1 Th2 Th17 CXCR3+CCR6+ Th1 Th2 Th17 CXCR3+CCR6+ Isotype Th1 Th2 Th17 CXCR3+CCR6+ Isotype Th1 Th2 Th17 CXCR3+CCR6+ Isotype Th1 Th2 Th17 CXCR3+CCR6+図 10. HC、OA 患者、RA 患者の PBMC 中の Th 細胞サブセットの遊 走能 0 5 10 15 20 25 30 HC OA RA m ig ra ti o n in d ex CXCL9
B
0 2 4 6 8 10 12 14 16 18 HC OA RA m ig ra ti o n in d ex CCL20 0 0.5 1 1.5 2 2.5 3 3.5 4 HC OA RA m ig ra ti o n in d ex CXCL10 0 100 200 300 ligand- ligand+ cell count 0 100 200 300 ligand- ligand+ cell count 0 100 200 300 ligand- ligand+ cell count 0 20 40 60 80 ligand- ligand+ cell count 0 20 40 60 80 ligand- ligand+ cell count 0 20 40 60 80 ligand- ligand+ cell count 0 100 200 300 ligand- ligand+ cell count 0 100 200 300 ligand- ligand+ cell count 0 100 200 300 ligand- ligand+ cell count HC OA RA CXCL9 CXCL10 CCL20A
C
0 20 40 60 80 100 before after C C R 6(+) ce lls (% ) 0 50 100 before after C C R 6(+) ce lls (% ) 0 20 40 60 80 100 before after C C R 6(+) ce lls (% ) 0 20 40 60 80 100 before after C XC R 3(+) ce lls (% ) 0 20 40 60 80 100 before after C XC R 3(+) ce lls (% ) 0 20 40 60 80 100 before after C XC R 3(+) ce lls (% ) 0 20 40 60 80 before after C XC R 3(+) ce lls (% ) 0 20 40 60 80 before after C XC R 3(+) ce lls (% ) 0 20 40 60 80 before after C XC R 3(+) ce lls (% ) HC OA RA CXCL9 CXCL10 CCL20HC(N=11)、OA 患者(N=9)、RA 患者(N=5)の Th 細胞サブセットの遊
走能ついてtranswell system を用いて chemotaxis assay を行った。CD4 陽性
T 細胞(1×105)を上層のwell へ添加し、4時間 37℃で培養し、下層の well
(ケモカイン(CXCL9、CXCL10、CCL20(100ng/ml)を含まないものと含む
もの)へ遊走したTh 細胞を FCM によって細胞数をカウントした。遊走指数
(migration index)については、ケモカインを含んだ well 中の細胞数とケモカ
インを含んでいないwell の細胞数の比により算出した。CXCL9 と CXCL10 は Th1 細胞、CCL20 は Th17 細胞数を用いて算出した。(A)Ligand を含まな いものと含むもののwell における FCM によってカウントした Th 細胞サブセ ットの細胞数。(B)(A)の結果から導き出された遊走指数の平均の比較。(C) chemotaxis assay の前後での CXCR3 陽性率及び CCR6 陽性率の平均の比較(* P < 0.05、平均 ± 標準偏差、2 群比較:Mann-Whitney U 検定、3 群比較: Kruskal-Wallis 検定)
図 11. HC、OA 患者、RA 患者の血清及び滑液中のケモカイン
HC 血清(N=8)、OA 患者血清(N=6)、RA 患者血清(N=7)、OA 患者滑 液(N=6)、RA 患者滑液(N=3)のケモカイン(CXCL9、CXCL10、CCL20) についてLEGENDplex で測定した。(*P < 0.05、平均 ± 標準偏差、 Kruskal-Wallis 検定) 0 2000 4000 6000 8000 10000 12000 HC Se rum OA Se rum RA Se rum OA SF s RA SF s C XC L 10 (p g /m l) 0 2000 4000 6000 8000 10000 12000 14000 16000 18000 HC Se rum OA Se rum RA Se rum OA SF s RA SF s C C L 20 (p g /m l) 0 200 400 600 800 1000 1200 HC Se rum OA Se rum RA Se rum OA SF s RA SF s C XC L 9 (p g /m l)
CXCL9
CXCL10
CCL20
図 12. RA 患者の治療前後の DAS28-CRP の変化 RA 患者(N=7)について治療前と抗リウマチ治療 24 週時について疾患活動性 スコアのDAS28-CRP の平均の推移を評価した。(*P < 0.05、平均 ± 標準偏 差、Wilcoxon 符号順位検定) 0 1 2 3 4 5
before treatment after treatment
D A S2 8 -C R P
4.08 ± 0.42
2.82 ± 0.35
図 13. RA 患者の治療前後の PBMC 中の Th 細胞サブセットの変化 RA 患者(N=7)について治療前と抗リウマチ治療 24 週時の Th 細胞サブセッ トについてFCM で解析を行った。(A)Th 細胞を CXCR3 と CCR6 により 4 つの細胞群に分けられたTh 細胞サブセットの代表例。(B)各 Th 細胞サブセ ットの割合の平均の比較(*P < 0.05、平均 ± 標準偏差、Wilcoxon 符号順位 検定)
CXCR3
CCR6
21.1 10.6 57.2 11 21.6 10.3 53.1 14.9after treatment
before treatment
0 10 20 30 40 50 60 Th1 Th2 Th17 CXCR3+CCR6+ T h s u b se ts (% ) after treatmet before treatmetB
図 14. RA 患者の治療前後の PBMC 中の Th 細胞サブセットの転写因子 の変化 RA 患者(N=7)について治療前と抗リウマチ治療 24 週時の Th 細胞サブセッ トについてFCM で解析を行った。(A)Th 細胞サブセットの転写因子(T-bet、 GATA3、RORγt)の代表的なヒストグラム。(B)Th 細胞サブセットの転写因 子(T-bet、GATA3、RORγt)を発現する細胞の割合の平均の比較。(*P < 0.05、 平均 ± 標準偏差、Wilcoxon 符号順位検定) 0 20 40 60
before treatment after treatment
T -b e t (+) c e ll s i n T h 1 c e ll s (% ) 0 10 20 30 40
before treatment after treatment
G A T A 3 (+) c e ll s i n T h 2 c e ll s (% ) 0 10 20 30
before treatment after treatment
ROR γt (+) c e ll s in T h 1 7 c e ll s (% ) after treatment before treatment T-bet GATA3 RORγt T-bet GATA3 RORγt
A
B
co u n t (% Ma x) co u n t (% Ma x) co u n t (% Ma x) Isotype Th1 Th2 Th17 CXCR3+CCR6+ Isotype Th1 Th2 Th17 CXCR3+CCR6+ Isotype Th1 Th2 Th17 CXCR3+CCR6+図 15. RA 患者の治療前後の PBMC 中の Th 細胞サブセットのケモカイ ンレセプターの変化 RA 患者(N=7)について治療前と抗リウマチ治療 24 週時の Th 細胞サブセッ トのケモカインレセプターについてFCM で解析を行った。(A)(左)Th1 細 胞におけるCXCR3 の代表的なヒストグラム。(右)Th17 細胞における CCR6 の代表的なヒストグラム。 (B)(左)Th1 細胞における CXCR3 の平均蛍光強度 (MFI)の平均の比較。(右)Th17 細胞における CCR6 の MFI の平均の比較。 (*P < 0.05、平均 ± 標準偏差、Wilcoxon 符号順位検定)
B
0 500 1000 1500 2000 2500 3000 3500 4000 4500 5000 before treatment after treatment C XC R 3 MF I in T h 1 c e ll s 0 2000 4000 6000 8000 10000 12000 14000 16000 before treatment after treatment C C R 6 MF I in T h 1 7 c e ll s CXCR3 CCR6 co u n t (% Ma x) b efo re tr ea tm en t afte r tr ea tm en tA
図 16. RA 患者の治療前後の SFMC 中の Th 細胞サブセットの変化 RA 患者(N=1)について治療前と抗リウマチ治療 24 週時の Th 細胞サブセッ トについてFCM で解析を行った。(A)Th 細胞を CXCR3 と CCR6 により 4 つの細胞群に分けられたTh 細胞サブセット。(B)各 Th 細胞サブセットの割 合の比較。
A
53.6 1.7 40.1 4.6 74.8 2.9 18.7 3.6CXCR3
CCR6
after treatment
before treatment
0
20
40
60
80
Th1 Th2 Th17 CXCR3+CCR6+ T h s u b se ts c el ls (% )B
after treatment before treatment図 17. RA 患者の治療前後の滑液中のケモカインの変化 RA 患者(N=1)について治療前と抗リウマチ治療 24 週時の滑液中のケモカイ ン(CXCL9、CXCL10、CCL20)を LEGENDplex で測定した。 0 1 2 3 4 5 6 7
before treatment after treatment
C XC L 9 (p g /m l)
CXCL9
0 2000 4000 6000 8000 10000 12000 14000 16000before treatment after treatment
C XC L 1 0 (p g /m l)
CXCL10
0 200 400 600 800 1000 1200 1400 1600before treatment after treatment
C C L 2 0 (p g /m l)
CCL20
謝辞 本研究遂行・論文執筆にあたり、ご指導ご鞭撻を賜りました住田孝之教授(筑 波大学医学医療系内科(膠原病・リウマチ・アレルギー)教授)、松本功准教 授(筑波大学医学医療系内科(膠原病・リウマチ・アレルギー)准教授)、近 藤裕也先生(筑波大学医学医療系内科(膠原病・リウマチ・アレルギー)講師) に心から感謝申し上げます。 また、OA 患者サンプルをご提供いただきました山崎正志教授(筑波大学医 学医療系外科(整形外科)教授)、金森章浩先生(筑波大学医学医療系外科(整 形外科)講師)に深く感謝申し上げます。 また、検体の提供にご協力を頂きました筑波大学附属病院の患者様方、なら びにスタッフの皆様に感謝致します。