大阪府立大学博士(獣医学)学位論文
マウスにおける Toxoplasma 特異的ヘルパーT 細胞 type-1 型
免疫応答誘導に関わる Toxoplasma 溶解抗原に含有する
タンパク質成分に関する研究
加 藤 美 穂 子
2 0 1 1 年
目次 緒言 1 Ⅰ章 Toxoplasma 溶解抗原(TLA)に対するマウス脾臓 T 細胞の特異的増殖 反応における p24 (GRA1) および p30 (SAG1) の関与 序論 5 材料及び方法 7 結果 18 考察 20 小活 22 Ⅱ章 p24 (GRA1) および p30 (SAG1) の Th1 細胞型免疫応答誘導能の比較 序論 23 材料及び方法 25 結果 29 考察 32 小活 35 Ⅲ章 Th1 細胞型免疫応答を誘導する p30 (SAG1) ペプチド領域と構造的特徴 序論 36 材料及び方法 38 結果 43 考察 46 小活 50 総括 51 図表 54 引用文献 69
緒言
Toxoplasma gondii (Tp) は 1908 年に Nicolle 及び Manceaux によりげっ歯類の一 種 Ctenodactylus gundi から発見された偏性細胞内寄生原虫で、アピコンプレッ ク ス 門 (Apicomplexa) 、 胞 子 虫 綱 (Sporozoasida) 、 真 コ ク シ ジ ウ ム 目 (Eucoccidiorida)、アイメリア亜目(Eimeriorina)に属し、ヒトを始めとした多 くのほ乳類や鳥類などの温血動物を中間宿主として感染する。ヒトは Tp 感染に 対して、健康な状態であれば臨床的には無症状で経過する。しかし、免疫能が 低下している場合は、初感染時に肺炎、心筋炎や脳炎などの急性症状を発症し て死に至る。また、妊婦においては、母体から胎盤を経由して胎児への感染が 成立し、流産、死産あるいは水頭症などの新生児機能不全症を引き起こすこと が報告されている (20)。さらに、Tp 初感染に耐過した場合でも、ヒト免疫不全 ウイルス (Human Immuno-Deficiency Virus; HIV) 感染に伴う後天性免疫不全症 候群 (Acquired Immuno-Deficiency Syndrom; AIDS) の発症により、脳などの組織 内でシストを形成していた Tp が再増殖して致死的なトキソプラズマ脳炎を発 症することが報告されている (29)。Tp のヒトへの感染は、一般的に Tp のシス トが存在する動物の肉、あるいは終宿主である猫などが排出するオーシストの 摂取により引き起こされる。したがって、生の食肉を摂取する機会が多い欧州 や、ペットとして猫を飼育する家庭が増えている日本において、ヒトの Tp 感染
症は特に予防が必要な原虫感染症である。 Tp がヒトの体内で致死的な活動をする妊娠期や HIV 感染症における AIDS 期 の共通点は、生体の細胞性免疫応答が抑制された状態にあるということである。 この様な事実から、Tp に対するヒトの感染防御における細胞性免疫応答の重要 性が指摘され、マウスを用いた様々な研究で報告されている (2, 50, 51)。特にイ ンターフェロンγ (Interferon-γ; IFN-γ) によりマクロファージ内における感染 Tp 虫体の増殖を抑制することが報告されている (60, 69)。また、細胞性免疫応答を 担う NK 細胞や細胞傷害性 T 細胞の活性化も、Tp に対する感染防御機構におい て重要な役割を担っている (9)。細胞性免疫応答の活性化には、ヘルパーT 細胞、 特にヘルパーT 細胞 type-1 (Th1 細胞) の活性化が強く関係しており、Tp に対す る感染防御において、Th1 細胞型免疫応答の誘導の重要性が示されている (9, 45)。したがって、Th1 細胞型免疫応答の誘導に関わる Tp の抗原領域の特徴を明 らかにすることは、Tp 感染に対する予防法を確立する上でも重要な課題である。 Toxoplasma 溶解抗原 (Toxoplasma gondii lysate antigen; TLA) はタキゾイト虫 体から抽出した成分であり、鈴木らのグループが TLA 投与マウスにおいて誘導 される免疫応答について多くの研究を報告している (22, 42, 43, 49, 53, 59)。小俣 ら は TLA 投 与 マ ウ ス の 血 清 中 に IFN-γ や マ ク ロ フ ァ ー ジ 遊 走 阻 止 因 子 (Macrophage migration inhibitory factor; MIF) 等のサイトカインの産生が上昇す
ることを報告している (50, 51)。また、TLA 投与マウスは、感染防御において 細胞性免疫応答が重要であるマウスマラリア原虫 (Plasmodium berghei) やマウ スバベシア原虫 (Babesia rodhainii) の感染に対して抵抗性が高くなると報告し ている (49)。さらに、TLA 投与マウスでは、メチルコラントレン誘発腫瘍や、 様々な移植腫瘍細胞の増殖を細胞傷害性活性の上昇により抑制する (42, 44)。こ れらの報告から、TLA には、Tp 特異的 Th1 細胞型免疫応答の誘導に関わる Tp 由来抗原が存在するものと考えられる。しかしながら、TLA 中の Tp 特異的 Th1 細胞型免疫応答誘導に関与する抗原の検討はほとんど行われていないのが現状 である。 本研究では、Tp 特異的 Th1 細胞型免疫応答誘導に密接に関係する TLA 抗原 成分を明らかにするため、現在までに報告のある Tp 抗原成分の中で、マウスに おいて Tp 特異的 Th1 細胞型免疫応答を誘導する可能性が考えられる p24 (GRA1) ならびに p30 (SAG1) に注目した。Tp の分泌型抗原成分である p24 によ り誘導された T 細胞は IFN-γ の産生が亢進していると報告されている (25, 54)。 また、p30 は Tp の主要膜抗原成分であり、p30 の蛋白質や遺伝子導入プラスミ ドを投与したマウスでは、Tp 特異的細胞性免疫応答が誘導されることが報告さ れている (19, 31, 32, 38, 55, 56, 65)。そこでまずⅠ章では、TLA 中における p24 および p30 の存在を明らかにすると共に、TLA に対するマウス脾臓 T 細胞の特
異的増殖反応におけるそれらの関与を検討した。次に、Ⅱ章では p24 ならびに p30 のマウスに対する Th1 細胞型免疫応答誘導能を検討した。さらに、Ⅲ章で はⅡ章で高い Th1 細胞型免疫応答誘導能を示した抗原の合成ペプチドを 20 種類 作成し、マウスに Th1 細胞型免疫応答を誘導するペプチドの二次構造の特徴を 明らかにした。マウスにおいて Th1 型の免疫応答誘導に関わる TLA 中の抗原領 域が明らかになることにより、Tp 感染防御のための生体内免疫賦活効果の増強 やワクチンとしての抗体医薬の開発に有効な結果が得られると考え、検討を行 った。
I 章 Toxoplasma 溶解抗原(TLA)に対するマウス脾臓 T 細胞の特異的増殖反応 における p24 (GRA1) および p30 (SAG1) の関与 序論 Toxoplasma 溶解抗原は、マウスに投与することにより IFN-γ の産生や細胞傷 害性活性を上昇させることから (43, 51, 68)、Th1 細胞型免疫応答を誘導する抗 原物質が存在するものと推測されている。 近年 Tp の抗原成分の研究が進み、それぞれの抗原の機能や分布から排出/ 分泌性、細胞内局在および表面局在などの種類が報告されている (12, 28, 41)。 その中でマウスに対して IFN-γ 産生を誘導することが報告されている抗原は、 p24 (GRA1) と p30 (SAG1) である (1, 28, 36, 37)。p24 は分泌型抗原であり Tp の 液胞や膜内に存在し、宿主侵入に関係するタキゾイト、ブラディゾイトに由来 する可溶性タンパク質である (6, 10, 54, 58)。また、p30 は glycophosphatidylinositol (GPI)アンカーにより結合している表面抗原の一つであり、宿主の原形質膜へ の接触に関係し (1)、タキゾイトにおける全タンパク質の 3.0 から 5.0% を占め ている (27)。TLA は凍結融解と超音波処理したタキゾイト虫体の可溶性抗原で あることから、分泌型抗原である p24 のみならず膜タンパク質である p30 も抽
出される可能性は高いため、これら抗原が TLA の Th1 細胞型免疫応答誘導能に 密接に関与していることも推測される。しかしながら,TLA 中における p24 と p30 の存在を検討した報告や、TLA に対する T 細胞の特異的反応における p24 や p30 の抗原としての関与について検討した報告は確認されない。 そこで本章では、TLA 中における p24 と p30 の存在をウエスタン-ブロッティ ングで確認した上で、マウス脾臓 T 細胞の TLA に対する特異的増殖反応におけ る p24 および p30 の関与について検討した。
材料及び方法 1. マウス 本研究では、BALB/c 系の 6‐8 週齢雌マウス(日本クレア、東京)を実験に 供した。 2. Toxoplasma gondii (以下 Tp) 虫体 国立大学法人帯広畜産大学原虫病研究センターで継代維持されている Tp RH 株を実験に用いた。感染は、1×103 個のタキゾイト虫体を含んだ 1 ml のハンク ス氏リン酸緩衝生理食塩水液(Hank’s balanced salt solution; ヘパリン 5 U/ml、硫 酸カナマイシン 60 μg/ml 含有; 以下 HBSS)をマウスの腹腔内に接種して行った。 感染 2 日後のマウスの腹腔に 5 ml の HBSS を注入し、回収した懸濁液中のタキ ゾイト虫体を実験に供した。
3. HeLa 細胞単層培養法による Tp 虫体の培養
Toxoplasma 溶解抗原(Toxoplasma lysate antigen; 以下 TLA)の調製のために、 Tp RH 株タキゾイト虫体を Valkoun らの方法 (64) に準じた in vitro 培養法によ り増殖させた。HeLa 細胞 1×107 個を 10 ml の 10% 熱非働化(56℃、30 分間) 胎仔牛血清(Fetal bovine serum, Flow laboratories, Australia; 以下 FBS)含有ダル
ベッコ改変 MEM 培地(以下 10% FBS-D-MEM)に懸濁後 75cm2細胞培養用フラ スコ(旭テクノグラス、東京)に播種し、37℃、5% CO2下で単層となるまで培 養した。5×107 個/ml のタキゾイト虫体 HBSS 懸濁液を、単層化した HeLa 細胞 の培養液に添加して 37℃、5% CO2下で 3‐4 日培養した。培養後虫体を HeLa 細胞と共に回収し、遠心分離(3,000×g 、10 分間、4℃)した。遠心上清を捨 て、沈渣にリン酸緩衝生理食塩水(Ca、Mg 不含、pH 7.2; 以下 PBS)10 ml を添 加して懸濁液を調製した。ディスポーザブルシリンジを用いて懸濁液を数回 27 G 注射針に通過させ、虫体と HeLa 細胞を分散させた。分散後の懸濁溶液 5 ml をあらかじめ調整した Percoll-Sucrose 密度勾配液に静かに重層し、15,000×g、 20 分間、4℃で遠心分離した。Percoll-Sucrose 密度勾配液は、Percoll (GE Healthcare,
USA) 9 ml に 2.5 M Sucrose 溶液 1 ml を重層し、15,000×g、20 分間、4℃で遠心 分離して調整したものを用いた。遠心分離後、虫体の層を回収して 0.25 M Sucrose 溶液を 3 倍程度添加、攪拌し、3,000×g、15 分間、4℃で遠心分離した。 遠心上清を捨て、沈渣に PBS を添加、撹拌して再度遠心分離した(3,000×g、 15 分間、4℃)。上清を取り除いた沈渣を使用時まで-80 ℃で保存した。 4. TLA の調整 TLA の調整は Saito らの方法 (53) を元に行った。前述の項目 3 で得られた
タキゾイト虫体の沈渣に重量と同量の滅菌蒸留水を加えて懸濁した。懸濁後、 -80 ℃による急速冷凍と室温における緩慢解凍による凍結融解を 3 回繰り返し た。次に融解した懸濁液に超音波処理(40 W、1 分間、5 回、30 秒間隔; Sonicator; model w220 F, Heat-systems-Ultrasonics Inc., Plainview, New York,USA)を行い、超 遠心分離(144,000×g、120 分間、4℃)した。遠心分離後、得られた上清を TLA 溶液として凍結乾燥した。凍結乾燥後の TLA は-20℃で保存し、用途に応じ適 当な溶媒で溶解して実験に用いた。TLA の総蛋白質量は牛血清アルブミン (Bovine Serum Albumin; 以下 BSA)を標準に用いた Bradford 法 (4) にて定量し た。マウスに投与する TLA は PBS にて 100 μg/ml となるように調整し、0.45 μm ディスポーザブルシリンジフイルターユニット(旭テクノグラス,東京)にて 濾過滅菌後、-80 ℃にて使用時まで保存した。
5. p24 および p30 組み換えタンパク質 (rp24 および rp30) の作製
本論文では、図 1 の p24(Accession No. M26007)と図 2 の p30(Accession No. X14080)の遺伝子配列を元にして以下の方法で作製して実験に供した (34)。ま ず、in vitro 培養で増殖させた Tp 虫体 1×107個を Trizol 試薬(Invitrogen, USA)
1 ml と共にホモジナイズし、試薬の添付書類で推奨されている方法を元に total
ml 添加し、さらによくボルテックスミキサーにより攪拌し、15,000×g、4℃、
15 分間遠心分離した。遠心分離後、回収した上清に等量のイソプロパノールを 添加して室温で 5 分静置後 15,000×g、4℃、15 分間遠心分離した。得られた total RNA の沈渣を 70% エタノールにて洗浄して風乾させた後、0.5% SDS 溶液にて 溶解して 260 nm における吸光度を測定し濃度を算出した。total RNA のうち 40 μg を 使 用 し 、 一 本 鎖 cDNA を 合 成 し た 。 逆 転 写 酵 素 (Superscript II, Invitrogen,USA) にキット付属の 5×RT 溶液 (4 μl) 、0.1 M DTT 溶液 (2 μl) 、
10 mM dNTPs (1 μl) 、Oligo dT25 primer (Promega, USA , 0.5 μl) を添加し、65℃で 2 分間加熱した。その後、氷冷して逆転写酵素及び RNase inhibior を各 1 μl ず つ添加し、42℃で 60 分間転写を行った。転写反応後、合成した cDNA を鋳型と し、TAKARA EX Taq (タカラバイオ、東京) を用いた PCR 法により p24、p30 の open reading flame (以下 ORF) 領域 (p24, 613-1185: 573bp; p30, 311-1321: 1011bp) を増幅した。PCR は、各 ORF 領域の PCR 産物の 5’及び 3’末端にそれ ぞれ EcoRI、SalI の制限酵素認識配列を付加するように実施した。PCR 後、各産 物を pGEM-T ベクター (Promega, USA) へライゲーションし、大腸菌 DH5α へ トランスフォーム (42℃、40 秒間) した後、氷上で 2 分間静置し、SOC 培地を 加え、37℃で 1 時間振盪培養した。その後アンピシリン (50 μg/ml) を含む LB 寒天培地 (以下 LB/Amp) に播き、37℃で一晩培養した。得られたコロニーを釣
菌し、ORF 増幅に用いた primer にて colony direct PCR (1 cycle; 94℃ 3 分間、30 cycle; 94℃ 20 秒間、55℃ 20 秒間、72℃ 45 秒間)を実施し、ORF 領域の挿入 を確認した。挿入を確認したコロニーを LB/Amp 液体培地にて培養し、Wizard plasmid purification system (Promega, USA) によりプラスミドを抽出した。抽出さ れたプラスミドに挿入されている p24 ならびに p30 遺伝子は DNA シーケンサー (CEQ2000, Beckman, USA) により配列を確認した。配列が確認された遺伝子は、
pGEM-T ベクターより制限酵素 EcoRI、BamHI で切り出し、p30 は pGEX4T-2 vector
(GE Healthcare, USA) 、p24 は StrepTag II IBA-2 vector (Sigma Genosys, USA) に、 それぞれライゲーションし、DH5α にトランスフォームさせた後、LB/Amp 寒天 培地に播いた。37℃で一晩培養後、得られたコロニーを釣菌し、LB/Amp 液体培 地 (500 ml) において振盪培養を行い (37℃) 、600 nm の吸光度が 0.6 程度に到 達した時点で、タンパク質発現誘導試薬として p30 には 0.1 mM IPTG、p24 には 200 ng/ml 無水テトラサイクリン (いずれも終濃度) を用いて 30℃で 3 時間、発 現誘導を行った。誘導後、p24 においては培養液を遠心分離 (3,000×g、4℃、10 分) し、得られた菌体に、緩衝液 W (100 mM Tris-HCl, pH 8.0, 1 mM EDTA, 4℃) 2.5 ml を加え懸濁し、氷上で超音波破砕機 (Micro-Ultrasonic Cell Disruper, Kontes,
USA) により菌体の破砕 (30 秒間、3 回)を行い、遠心分離 (3,000×g、4℃、10 分間) した。遠心分離後の上清を 0.8 μm の濾過滅菌フィルターを通過させて、5
ml の緩衝液 W にて平衡化を行った Strep-Tactin レジン 1 ml に反応させた。上 清反応後のレジンを 10 ml の緩衝液 W で洗浄し、緩衝液 E (緩衝液 W に 2.5 mM Desthiobiotin (Sigma, USA) を含む) を 0.5 ml ずつレジンに添加して精製画分を 回収した。一方、p30 においては培養液 500 ml を遠心分離 (3,000×g、4℃、10 分間) して集めた菌体に、冷 PBS 5 ml を加え懸濁し、p24 と同様に超音波破砕 を行い遠心分離 (3,000×g、4℃、10 分間) した。遠心分離後の上清を 0.8 μm 濾 過滅菌フィルターを通過させて、Glutathione Sepharose (GE Healthcare, USA, 1 ml) に反応させた。反応後の Glutathione Sepharose を PBS にて洗浄した後、
GST Tag を切断するために Thrombin による酵素消化処理を行った。PBS 1 ml で レジンを懸濁させ、そこに 0.2 U の Thrombin (GE Healthcare, USA) を添加し、4℃ で一晩静置して消化処理を行った。切断された p30 画分はレジンから分離回収 した。得られた組換えタンパク質をそれぞれ rp24 、rp30 とした。SDS-PAGE による純度確認の後、フェニルチオヒダントインアミノ酸誘導体を標準物質と したエドマン分解法 (11) によるアミノ酸分析により N 末端から 5 アミノ酸の 配列確認を行った。配列確認後、BSA を標準に用いた CBB Protein Assay Reagent (PIERCE, USA) によるタンパク質濃度測定を行った。上述したように調整され た p24 および p30 の組換えタンパク質 (rp24 および rp30) は、虫体由来の同蛋白 質と同様な抗原性を有することが報告されていることから (52, 61)、以下の実験
にそれぞれの抗原として供した。
6. マウス抗 rp24 抗体および抗 rp30 抗体の作製
rp24 および rp30 を抗原としてマウスを用いて抗 rp24 抗体ならびに抗 rp30 抗 体を作製した。各抗原タンパク質の投与は、3 頭のマウスに行った。マウス 1 頭当たり 50 μg (200 μl の PBS に溶解) のそれぞれのタンパク質を 2 週間間隔 で計 4 回腹腔内に投与した。初回投与は Freund’s Complete Adjuvant (DIFCO, USA) 、2 回目以降は Freund’s Incomplete Adjuvant (DIFCO, USA) とそれぞれ の抗原タンパク質を 1:1 で混和したエマルジョンを用いた。最終抗原投与日から 一週間後、各マウスの血清を採取し、抗 rp24 抗血清ならびに抗 rp30 抗血清とし て実験に供した。
7. ウエスタン-ブロッティング
TLA 1 μg (100 μg/ml 滅菌蒸留水) をタンパク質試料緩衝液 (125 mM Tris-HCl, 2% SDS, 10% Glycerol, 0.1% Bromophenol blue, 10% 2-Mercaptoethanol, pH 6.8) と
1:1 の割合で混合し 95℃、 5 分間の熱処理を行った。SDS-PAGE には 12.5 %の スラブゲルを用い、1つのコーム穴にはプレステインド分子量スタンダードマ ーカー (アプロサイエンス、徳島) を 5 μl、他のコーム穴には上述のように処理
し た TLA 20 μl を ア プ ラ イ し た 。 泳 動 装 置 は Mini-Protean Ⅱ (Bio-Rad Laboratories, USA) を、パワーサプライには RealPower (バイオクラフト、東京) を 用い、40 mA で 90 分間泳動行った。 泳動後、ゲルをクマシーブリリアントブ ルー(CBB)による染色あるいは Towbin らの方法 (62) に従いセミドライ型ブロ ッティング装置 (バイオクラフト、東京) によって PVDF 膜 (Fluorotrans, PALL, USA) に転写した。PVDF 膜は、転写前に 20 秒間メタノールに浸漬した後、ト ランスバッファー (0.1 M Tris, 20% Methanol, 0.192 M Glycine) に湿らせた。ゲル と PVDF 膜をトランスバッファーに湿らせた濾紙の間に挿み、0.8 mA/cm2でブ ロッティングを行った。転写後、膜を 0.5% スキムミルク-PBS にて室温で 2 時 間ブロッキング操作を行った。ブロッキング終了後、膜を PBS-T にて振盪洗浄 (5 分間、3 回) し、5% スキムミルク-PBS で 3,000 倍に希釈した抗 rp24 および rp30 抗血清に浸して 37℃、1 時間振盪して反応させた。反応後の膜から未反応の抗 血清を除去するため、PBS-T にて振盪洗浄 (5 分間、3 回) し、5% スキムミル ク-PBS で 3,000 倍希釈したアルカリフォスファターゼ標識抗マウス IgG (Promega, USA) を 37℃、1 時間振盪して反応させた。反応後、膜を PBS-T にて 振盪洗浄後(5 分間、3 回) アルカリフォスファターゼ用発色液 (nitro blue teteazolium (NBT)/5-bromo-4-chloro-3-indolyl-phosphate (BCIP) color substrate
ン溶液 (pH 9.8) 10ml にて溶解) で発色させた。 8. TLA 感作マウスの作製 マウスの腹腔内に TLA 50 μg (100 μg/ml 滅菌蒸留水) を投与し、投与 2 週間後 に同量の抗原をさらに投与して 2 週間を経過したものを TLA 感作マウスとして 実験に供した。 9. 脾臓内 T 細胞ならびに付着性細胞の調整 脾臓内 T 細胞ならびに付着性細胞は、Inaba らの方法 (23) により調製した。 脱頚椎死させたマウスから脾臓を無菌的に摘出し、脾臓内に PBS で溶解した 100 U/ml コラゲナーゼ (和光純薬、東京) 溶液 100 μl を注入し、37℃、30 分間シャ ーレ内で静置した。コラゲナーゼ処理した脾臓をハサミで細切し、ステンレス 製メッシュ上でガスケットを用いて臓器内の細胞を押し出した。メッシュを冷 RPMI1640 (以下 RPMI) で洗浄し、通過した脾臓内細胞の懸濁液を 50 ml の遠心 管に回収して遠心分離した (800×g、10 分間、4℃)。上清を吸引除去し、混入 赤血球を溶血するため 37℃の 0.83% 塩化アンモニウム溶液を沈渣に添加、撹拌 して遠心分離した (800×g、10 分間、4℃)。上清を吸引除去して沈渣に RPMI を添加撹拌後遠心した (800×g、10 分間、4℃)。細胞数を算出後、同様に洗浄
した。洗浄後の脾臓内細胞を 10% FBS 加 RPMI で 3~5×106 個/ml に調整し、こ の浮遊液 10 ml を 9 cm の滅菌プラスチックディッシュに分注して 37℃、5% CO2下で1晩培養した。培養後、非付着性細胞を回収するためにプラスチック ディッシュを振盪して上清を回収し、更に 37℃に温めた RPMI をプラスチック ディッシュに再度加え、再度軽く振盪して回収したものを遠心管に移して遠心 分離した (800×g、10 分間、4℃) 。遠心分離後、細胞数を算出し、脾臓内非付 着細胞を再度冷 RPMI で洗浄した。RPMI で 1×107 個/ml に調整した 1 ml を、 Julius ら (26) の方法で用いられる T 細胞分離用ナイロンウールカラム (和光純 薬、東京) に通過させて T 細胞を回収した。この方法ではカラム通過後の細胞 の T 細胞含有比率は 90% 以上であることから、回収した細胞を脾臓内 T 細胞と して RPMI で洗浄後、実験に供した。一方、脾臓内非付着性細胞を除去したデ イッシュは、0.02% EDTA-PBS で数回洗浄して上清を吸引除去した。適量の PBS をデイッシュに添加し、ラバーポリスマンを用いて付着性細胞をデイッシュか ら剥離、回収し、遠心分離した。遠心上清を捨て、さらに RPMI で遠心洗浄を 行ったものを脾臓付着性細胞として実験に供した。 10. 脾臓内 T 細胞増殖性の検討 細胞内 ATP 活性は、生細胞の増殖性を示していることから (8)、以下の方法
で細胞の増殖活性の測定を行った。脾臓内 T 細胞ならびに付着性細胞は 10% FBS 加 RPMI で各々2×106 個/ml、1×105 個/ml に調整し、96 穴マイクロウェルの各 well に 50 μl ずつ分注した。対照群ならびに各抗原添加群を設け、それぞれ 3 well に対照群では培養液のみ、抗原添加群では抗原最終濃度が 1.0 μg/ml 及び 0.1 μg/ml となるように 50 μl ずつ添加し、37℃、5% CO2で 5 日間培養を行った。 Yamashita ら(67) は抗原特異的 T 細胞の活性化の増殖性は、抗原添加培養後 4-5 日目に最大となると報告していることから、培養 5 日目に活性の評価を行った。 培養後、各 well に Celltiter GLO 試薬 (Promega, USA) を 50 μl ずつ添加し、ル ミノメーター (Orion-Ⅱ, Roche, Germany)で ATP 活性を測定した。増殖活性の単 位は相対的発光量の指標として Luminescence (RLU) で表示した。 増殖活性は 各群 3 well の平均値ならびに標準偏差 (Standard Deviation; SD) を算出して比較 した。
11. 統計処理
測定データは平均値と標準偏差 (SD) とを求め、対照群との有意差は Student-t 検定により算出した。
結果 1. TLA における p24 および p30 抗原の検出 TLA 中における p24 および p30 抗原成分の存在をウエスタン-ブロッティング 法で検出した。TLA の SDS 電気泳動により、TLA には様々な分子量の異なるタ ンパク質の存在が確認された (図 3 Lane 3)。また、ウエスタン-ブロッティング では、抗 rp24 血清により 24 kDa の位置に単一のバンドが検出され(図 3 Lane 1)、 同様に抗 rp30 血清を用いた検討では、30 kDa の位置に単一のバンドが検出され た(図 3 Lane 2)。TLA に含まれる全タンパク質中の p24 および p30 の含有量を、 SDS 電気泳動のクマシーブリリアントブルー (CBB) 染色像から Scion Image
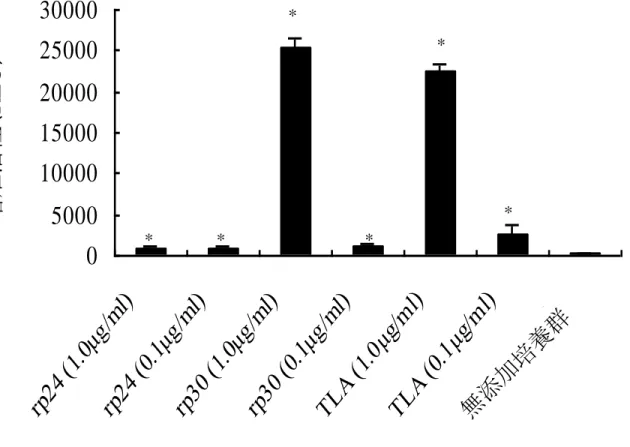
(Scion Corporation, USA) を用いて解析したところ、p24 および p30 は各々約 3.0 および 1.2%の含有量だった。 2. TLA 感作マウス脾臓内 T 細胞の TLA、p24 および p30 に対する特異的増殖 反応 結果 1 から TLA には p24 および p30 の存在が確認されたが、マウスの TLA 特異的な T 細胞の反応に p24 と p30 が抗原として関与しているかどうかは不明 である。そこで TLA 感作マウスの脾臓内 T 細胞の TLA、rp24 および rp30 に対 する増殖反応を検討した。
TLA : 0.1 μg/ml 添加群は、顕著ではないが無添加培養群と比較して有意な増 殖活性値を示した (TLA : 0.1 μg/ml 添加群, 2,535 ± 1,138 RLU; 無添加群, 287.5 ± 99.7 RLU; p < 0.01)。また、TLA : 1.0 μg/ml 添加群は、無添加培養群と 比較して有意な活性値を示し、TLA : 0.1 μg/ml 添加群よりも顕著に高値であっ た(TLA : 1.0 μg/ml 添加群, 22,450.0 ± 1,000.5 RLU; 無添加群, 287.5 ± 99.7 RLU; p < 0.01)。 rp30 : 0.1 μg/ml 添加培養群は、顕著ではないが無添加培養群と比較して有意な 増殖活性値を示し、その値は TLA : 0.1 μg/ml 添加群とほぼ同等であった (rp30 : 0.1 μg/ml 添加群, 1,298.0 ± 219.0 RLU; p < 0.01)。また、rp30 : 1.0 μg/ml 添加群 は、無添加培養群と比較して有意な顕著に高い増殖活性値を示し (rp30 : 1.0 μg/ml 添加群, 25,353.0 ± 1,163.0 RLU; p < 0.01)、その値は TLA : 1.0 μg/ml 添加 群とほぼ同等であった。一方、rp24 : 0.1 μg/ml 添加培養群は、無添加培養群と比 較して比較して有意な増殖活性値を示したが、その値は TLA : 0.1 μg/ml 添加群 よりも低い傾向であった (rp24 : 0.1 μg/ml 添加群, 750.8 ± 284.3 RLU; p < 0.01)。 また、rp24 : 1.0 μg/ml 添加培養群は rp24 : 0.1 μg/ml 添加培養群と同様に無添加培 養群と比較して有意ではあるものの高い活性値は示さなかった (rp24 : 1.0 μg/ml 添加群, 885.2 ± 125.0 RLU; p < 0.01)(図 4)。
考察
マラリア原虫 (Plasmodium berghei) やバベシア原虫 (Babesia rodhainii) 感染 マウス (49)、メチルコラントレン誘発腫瘍などの腫瘍を移植したマウス (42, 44) などに対し、TLA はマウスに IFN-γ 産生や細胞傷害性活性を誘導することが報 告されている。Kato らは、これら報告に基づき TLA から IFN-γ 産生や細胞傷害 性活性を誘導する成分の解析を行ったが、TLA と同様の活性を示す特定のタン パク質の一次構造を同定するには至らなかった (35)。そこで本研究では、Tp の 抗原として既に報告があり、IFN-γ 産生誘導能をもった抗原である p24 と p30 に 注目した。本研究を開始する前は、p24 は分泌抗原であることから TLA 中に存 在すると考えられたが、p30 は GPI アンカーにより結合している表面抗原であ るため、可溶性タンパク質成分として回収された TLA では含有量としては尐な いものと予測した。しかしながら、抗 rp24 血清と抗 rp30 血清を用いたウエスタ ン-ブロッティングでは、それぞれ 24 kDa と 30 kDa の位置に単一のバンドが検 出され、TLA 中に p24 と p30 の存在が確認された。p30 は Tp の主要膜タンパク 質の一つであり、通常 Tp 虫体から回収する場合、TritonX-114 などの界面活性剤 を用いて糖脂質成分として抽出される (30)。 p30 が TLA 中に溶出された理由 としては、調製方法である凍結融解および超音波処理を繰り返すことにより細 胞膜の脆弱化が生じ、可溶化成分と共に抽出されたものと考えられた。
TLA の電気泳動像から TLA 中における p24 および p30 のタンパク質含有率を 解析したところ、主要成分では無いものの各々3.0 および 1.2%程度含まれている ことが判明した。そこで、TLA 感作マウス脾臓内 T 細胞において rp24 および rp30 と TLA を同一濃度で刺激して増殖活性を比較した場合、TLA で刺激した群より も高い増殖活性を示すものと予測し、検討を行った。TLA 中の p24 および p30 の含有量から、p24 の方が p30 と比較して増殖活性を高く誘導するものと予測し た。rp24 あるいは rp30 刺激群のいずれも無添加培養群に対しては有意差のある 高い増殖活性を示したが、TLA で刺激した群との比較では有意差はなかった。 しかし、rp30 刺激群では TLA で刺激した群と同等の増殖活性を示した。p24 は TLA 中の他の抗原成分の存在がなくては増殖活性を相乗的に誘導出来ないが、 p30 は TLA 感作マウス脾臓内 T 細胞の TLA に対する特異的増殖反応に密接に関 与していると推察された。本章では、p24 と p30 の TLA 感作マウス脾臓内 T 細 胞の抗原特異的増殖反応への関与を明らかにしたが、p24 と p30 のマウスにおけ る Th1 細胞型免疫応答誘導能については明らかではない。そこで、p24 と p30 の Th1 細胞型免疫応答誘導能をさらに検討することとした。
小活 TLA における分泌型抗原 p24 と膜結合型抗原 p30 の存在を抗 rp24 と抗 rp30 血清を用いたウエスタン-ブロッティングで検討した。その結果、抗 rp24 と抗 rp30 血清によりそれぞれ 24 kDa と 30 kDa の位置に単一のバンドが検出された。 次に、TLA 感作マウス脾臓内 T 細胞の TLA、rp30 および rp24 に対する特異的 増殖反応を検討した。その結果、脾臓内 T 細胞は rp30 に対して TLA と同等の 高い増殖活性を示したが、rp24 に対しては低い活性を示した。以上の結果から、 TLA には p24 および p30 が存在し、TLA 感作マウス脾臓内 T 細胞の TLA に対 する特異的増殖反応には、いずれの抗原も増殖活性を示すものの p24 に比較し て p30 が抗原として密接に関係している可能性が推測された。
Ⅱ章 p24 (GRA1) および p30 (SAG1) の Th1 細胞型免疫応答誘導能の比較 序論 Ⅰ章において、TLA には分泌型抗原である p24 (GRA1) と膜結合型抗原であ る p30 (SAG1) が共に存在し、p30 に比べ p24 は活性化の程度は低いが、両抗原 ともに TLA 感作マウス脾臓内 T 細胞を活性化することから、p24 と p30 は TLA の Th1 細胞型免疫応答の誘導能に関係しているものと推測された。 Th1 細胞型免疫応答の誘導は、様々な免疫細胞における抗原提示に伴う接着分 子などの発現やサイトカインの産生などが指標となることが多くの研究で報告 されている (3, 9)。特に T 細胞からの IFN-γ や抗原提示細胞から産生されるイン ターロイキン 12 (Interleukin-12; IL-12) は、マウスにおいては Tp 感染の際、生体 内防御システムに重要な働きを持つと考えられている (3, 13-15, 17)。また、抗 原に対して血清中に産生される IgG サブクラスも Th 細胞型免疫応答誘導の指標 として知られており、マウスでは Th1 細胞型免疫応答では IgG2a が有意に産生 され、Th2 細胞型免疫応答では IgG1 が有意に産生されることが知られている (5, 45)。I章では p24 および p30 が TLA の Th1 型免疫応答誘導能に関係するもの と推察されたが、双方の抗原が Th1 型免疫応答誘導能を有するか否かは不明で
ある。そこで本章では rp24 と rp30 感作マウスにおいて、感作抗原刺激による 脾臓内 T 細胞と付着性細胞の IFN-γ や IL-12 mRNA の発現状況ならびに血清中 の感作抗原反応性 IgG サブセットを比較検討し、TLA の Th1 型免疫応答の誘導 を主に担っている抗原が rp24 と rp30 のいずれかを評価した。
材料と方法 1. マウス BALB/c 系の 6‐8 週齢雌マウス(日本クレア、東京)を実験に供した。 2. p24 および p30 組み換えタンパク質の作成 Ⅰ章で作製した rp24 ならびに rp30 を実験に供した。 3. 抗原感作マウスの作製 マウス 3 頭の腹腔内に組み換えタンパク質 rp24 あるいは rp30 を 50 μg (100 μg/ml 水溶液) をそれぞれ投与し、投与 2 週間後に同量の抗原をさらに投与して 2 週間を経過したものを rp24 あるいは rp30 感作マウスとして実験に供した。 4. 脾臓内 T 細胞ならびに付着性細胞の抗原刺激 非感作あるいは抗原感作マウス 3 頭から脾臓内 T 細胞ならびに付着性細胞 を調整して実験に供した。準備した脾臓内 T 細胞及び付着性細胞は 10% FBS 加 RPMI で各々2×106 個/ml、1×105 個/ml に調整し、24 穴プレートの各 well に 0.5 ml ずつ分注した。対照群ならびに各抗原添加群を設け、それぞれ対照群 では培養液のみ、抗原添加群では抗原含有培養液を 0.1 μg/ml および 1.0 μg/ml
となるように添加し、37℃、5% CO2で 5 日間培養を行った。培養後の細胞に ついて、感作抗原特異的増殖反応の活性を I 章と同様の方法で、またサイトカ イン mRNA の発現量を以下に示す方法で検討した。
5. サイトカイン mRNA 発現の検討
前述の項目 4 で培養した細胞のプレートを遠心し上清を除去後、Trizol 試薬 (Invitrogen, USA) 1 ml を添加して、I 章と同様の方法に準じて total RNA を回収 した。回収した total RNA のうち 40 μg を使用し、一本鎖 cDNA を合成した。逆 転写酵素 (Superscript II, Invitrogen) に付属の 5×RT 溶液 (4 μl) 、0.1 M DTT 溶 液 (2 μl) 、10 mM dNTPs (1 μl) 、Oligo dT25 primer (0.5 μl) を添加し、65℃で 2 分間加熱した。加熱後、氷冷して逆転写酵素及び RNase inhibior を各 1 μl ずつ 添加し、全量を 20 μl となるように DEPC 処理水にて調整した反応液を 42℃で 60 分間加熱して、逆転写反応を行った。TE buffer (pH 8.0) で 5 倍希釈した合成
cDNA 溶液 1 μl に、20 mM dNTPs 溶液 (2 μl) 、Taq DNA polymerase (0.1 μl;
Promega, USA) 、15 mM MgCl2液 (1.6 μl) 、10×PCR buffer (2 μl)、表 1 に示した IFN-γ (63)、 IL-12 (p40) (17) およびマウス β-actin の forward ならびに reverse
primer 液 (50 μM) 各 0.1 μl を添加し、全量 20 μl となるように調整した。PCR 反 応は、各 3 回行った。反応条件は IL-12 については、94 ℃で 3 分間を 1 cycle、
94 ℃で 20 秒間、55 ℃で 20 秒間ならびに 72 ℃で 45 秒間を 30 cycle、IFN-γ お よびβ-actin については 94 ℃ 30 秒間、 55 ℃ 30 秒間、72 ℃ 60 秒間 を 5 cycles 及び 94 ℃ 30 秒間、50 ℃ 30 秒間、72 ℃ 60 秒間を 29 cycles と設定し た。なお、本研究では IL-12 mRNA の発現を、恒常発現している p35 ではなく、 抗原刺激により発現誘導される p40 について検討した (17, 40)。PCR 反応後、得 られた産物をエチジウムブロマイド含有の 1.5%アガロースゲルで 100 V で 60 分間電気泳動した。 泳動後、ゲルにおける産物のバンドをトランスイルミネー ター上で撮影し、Scion Image (Scion Corporation, USA) を用いて、β-actin 増幅産 物に対する mRNA の発現量比を求めた。
6. rp24 および rp30 感作マウス血清の作製
3.と同様の方法で抗原感作マウス作製後、回収した血清を実験に供した。
7. 血清中における rp24 あるいは rp30 反応性 IgG サブクラスの測定
抗原感作マウス血清中の IgG サブクラスは、Bourguin ら (3)、Seng ら (55) の 報告を元に、以下に示すように間接 ELISA 法により測定した。
マイクロタイタープレート (Maxisorp, Nunc, Denmark) の各 well に、各抗原溶 液を 100 μl (2.0 μg/ml) 分注して室温で 2 時間固相化した。固相化後各 well を
0.02% Tween 添加 PBS (以下 T-PBS) により 3 回洗浄し、5%スキムミルク添加 PBS を各 well に 100 μl 分注して室温で 2 時間ブロッキング処理を行った。rp24 ある いは rp30 感作マウスの血清を PBS で 1/400 から 1/3,200 まで希釈し、それぞれ rp24 あるいは rp30 を固相化した well に 100 μl ずつ分注して 37℃で1時間静置 した。静置後、各 well を T-PBS で 3 回洗浄し、0.5%スキムミルク添加 T-PBS で 1,000 倍に希釈した HRP 標識抗マウス IgG1 (Zymed, USA) および IgG2a (コスモ バイオ、東京) 抗体溶液を各 well に 100 μl 加え 37℃で 1 時間静置した。静置後 各 well を T-PBS で 3 回洗浄し、基質テトラメチルベンジジン (3, 3’, 5, 5’-tetramethyl benzidine stabilized substrate; 以下 TMB、Dako、東京) 溶液を各 well に 100 μl ずつ分注して室温で 10 分間静置して発色を行った。発色後、1 N 硫酸 を 50 μl ずつ分注して反応を停止し、450 nm で各 well の吸光度 (OD 値) をプレ ートリーダー (A450i、東ソー、東京) にて測定した。各感作血清の測定は二重 測定で行い、各個体の OD 値の平均と標準偏差を算出し、比較した。
結果
1. rp24 および rp30 感作マウス脾臓内 T 細胞と付着性細胞の感作抗原刺激によ る IFN-γ および IL-12 mRNA の発現
rp24 および rp30 の脾臓内T細胞と付着性細胞に対する IFN-γ および IL-12 mRNA 発現誘導能を検討する上で、まず rp24 と rp30 感作マウスの脾臓内 T 細 胞が付着性細胞と共に培養した場合に、それぞれの感作抗原刺激に対して有意 な特異的増殖反応を示すことを確認した。0.1 および 1.0 μg/ml の rp24 を添加し た群では、増殖活性がそれぞれ 59,200.0 ± 3,897.0 RLU、69,587.0± 17,038.0 RLU と、無添加培養群の 7,805.0 ± 852.0 RLU に比較して有意に高値を示し、 その活性は抗原濃度に依存していた (図 5A ; p < 0.01)。また、0.1 μg/ml および 1.0 μg/ml の rp30 を添加した群においても、増殖活性がそれぞれ 34,060.3 ± 5,351.0 RLU、124,820.7 ± 291,29l.0 RLU と、無添加培養群の 3,098.7 ± 702.0 RLU に比較して有意な高値を示し、rp24 と同様に抗原濃度に高く依存していた (図 5B; p < 0.01)。以上のように、rp24 と rp30 感作マウス脾臓内 T 細胞は付着 性細胞と共に培養することにより、それぞれの感作抗原刺激に対して増殖反応 を示した。また、その増殖活性は濃度依存的であり、1.0 μg/ml の抗原添加によ り高い増殖活性が得られた。次に、各々の感作抗原を 1.0 μg/ml 添加培養した脾 臓内 T 細胞と付着性細胞について、サイトカイン mRNA 発現の検討を行った。
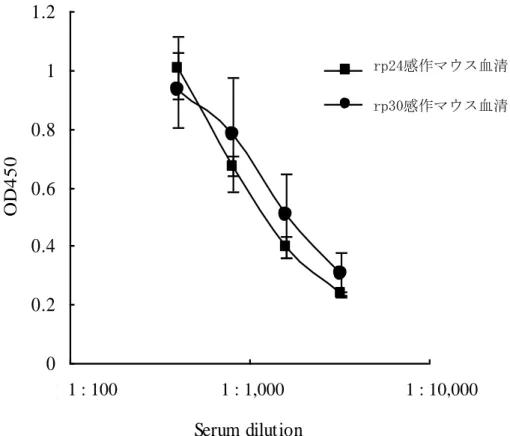
β-actin mRNA に対する IFN-γ mRNA の発現量比を算出したところ、非感作マウ スでは IFN-γ mRNA の発現が確認されなかったのに対して、rp24 感作マウスで は 0.15、rp30 感作マウスでは発現量比が 0.27 であった。また、β-actin mRNA に 対する IL-12 mRNA の発現量比は、非感作マウスでは 0 であったのに対して、rp24 感作マウスでは 0.33、rp30 感作マウスでは 1.0 であった。非感作マウス、rp24 および rp30 感作マウスの脾臓内 T 細胞と付着性細胞について同様の検討を 3 回 行ったが、いずれの場合も同様の結果であった。以上の結果から、rp24 および rp30 感作マウスの脾臓内 T 細胞と付着性細胞には、それぞれの感作抗原の刺激 により IFN-γ mRNA と IL-12 mRNA が誘導された。また、いずれのサイトカイ ン mRNA 発現量比も rp24 感作マウスより rp30 感作マウスの方が高値を示した (図 6)。 2. rp24 および rp30 感作マウス血清の感作抗原に対する総 IgG、IgG1 および IgG2a 抗体価の比較 rp24 および rp30 抗原感作マウス血清中の感作抗原に反応する抗体価を総 IgG にて比較した。 rp24 および rp30 感作マウス血清を 1/400、1/800、1/1,600 およ び 1/3,200 希釈した場合の総 IgG 抗体価を OD 値で比較検討した。図 7 に示すよ うに、総 IgG の OD 値は 1/400 希釈では rp24 感作, 1.007 ± 0.125; rp30 感作, 0.932
± 0.108、1/800 希釈では rp24 感作, 0.671 ± 0.195; rp30 感作, 0.779± 0.033、 1/1,600 希釈では rp24 感作, 0.395 ± 0.142; rp30 感作, 0.504 ± 0.037、1/3,200 希釈では rp24 感作, 0.24 ± 0.074; rp30 感作, 0.302 ± 0.006 であった。いずれの 希釈においても rp24 と rp30 感作マウス血清間の OD 値に有意な差異は認められ なかった。 次に rp24 ならびに rp30 感作マウス血清中の感作抗原に反応する IgG1 の抗体 価についても同様に比較検討した。 IgG1 の OD 値は、1/400 希釈では rp24 感作, 1.250 ± 0.090; rp30 感作, 0.820 ± 0.150、1/800 希釈では rp24 感作, 0.810 ± 0.080; rp30 感作, 0.501 ± 0.120、1/1,600 希釈では rp24 感作, 0.550 ± 0.100; rp30 感作, 0.150 ± 0.070 であった。1/400、1/800 および 1/1,600 の希釈においても rp24 感作マウス血清の方が rp30 感作マウス血清に比較して有意に高値を示していた (p < 0.05、図 8-A)。 一方、rp24 ならびに rp30 感作マウス血清中の感作抗原に反 応する IgG2a の抗体価についても同様に比較検討した。全ての希釈において、 rp30 感作マウス血清の方が rp24 感作マウス血清に比較して高値を示しており、 1/400 と 1/800 希釈では有意であった (1/400 希釈では rp24 感作, 0.219 ± 0.066; rp30 感作, 0.383 ± 0.009、1/800 希釈では rp24, 0.124± 0.037; rp30, 0.206 ± 0.036; p < 0.05、 図 8-B)。
考察 I 章では TLA 感作マウス脾臓内 T 細胞が rp24 および rp30 に対して有意な増殖 反応を呈していたが、その反応は rp24 に比較して rp30 の方が高かった。その理 由として、TLA 中における p24 と p30 の増殖活性誘導の相違が考えられた。そ こで本章では、rp24 および rp30 感作マウスを同一濃度条件で作製し、双方の感 作マウスの脾臓内 T 細胞と付着性細胞をさらに同一濃度の感作抗原で刺激して、 抗原特異的増殖反応を検討した。その結果、rp24 および rp30 感作マウス脾臓内 T 細胞と付着性細胞は、感作抗原に対して双方とも有意な特異的増殖反応を同 様に示した。このことからI章における TLA 感作マウス脾臓内 T 細胞の rp24 および rp30 に対する増殖活性の相違には、TLA 中に存在する他の増殖活性成分 との相互作用により各抗原の増殖活性誘導が影響を受けている可能性が推察さ れた。 感作抗原に対して同様な特異的増殖反応を呈した rp24 および rp30 感作マウス 脾臓内 T 細胞と付着性細胞について、Th1 細胞型免疫応答の誘導の有無を明ら かにするため、指標となる IFN-γ と IL-12 mRNA の発現状況を検討した。rp24 および rp30 感作マウスの脾臓内 T 細胞と付着性細胞は共に IFN-γ と IL-12 mRNA の発現が認められ、p24 および p30 が共に免疫細胞に IFN-γ の産生を誘導するこ と、また Th1 細胞型免疫応答の誘導能を持っていることが推察された。しかし
ながら、rp24 および rp30 共に同一濃度による感作刺激において、rp24 感作マウ スに比較して rp30 感作マウス脾臓内 T 細胞と付着性細胞は、IFN-γ と IL-12 mRNA の発現比が高かった。このことは、rp24 感作マウスに比較して、rp30 の 方が Th1 細胞型免疫応答誘導能が高いことを推測させる。 サイトカイン mRNA の発現状況による rp24 と rp30 の感作マウスの Th1 細胞 型免疫応答誘導能の判定に加え、各感作マウスの血清における感作抗原に反応 する総 IgG、IgG1 ならびに IgG2a の抗体価を比較検討した。血清中の感作抗原 反応性総 IgG の抗体価については、双方の感作マウス間に有意な差は認められ なかった。また IgG1 および IgG2a については rp24 および rp30 感作マウス血清 共に産生されていたが、IgG1 では rp24 感作マウスに比較して rp30 感作マウス の方が有意に低値を示し、IgG2a では rp24 感作マウスに比較して rp30 感作マウ スの方が有意に高値を示した。以上の結果から、いずれの抗原とも抗原特異的 IgG1 および IgG2a を産生するが、rp24 は IgG1 抗体産生を有意に誘導し、rp30 は IgG2a 抗体産生を有意に誘導するものと推察された。
以上の様に、rp24 と rp30 感作マウスの脾臓内 T 細胞と付着性細胞には感作抗 原刺激により IFN-γ と IL-12 mRNA の発現は誘導されるものの、その発現量は rp24 に比較して rp30 感作マウスの細胞の方が高かったこと、さらに血清中の感 作抗原反応性 IgG2a 抗体価が rp24 に比較して rp30 感作マウスの方が有意に高
かったことから、p30 は p24 に比較して Th1 細胞型免疫応答誘導能が高いもの と推察された。また、I章と本章の結果から TLA の Th1 細胞型免疫応答誘導能 を示す主な成分として p30 が関連していると推測された。 本章では、TLA の Tp 特異的 Th1 細胞型免疫応答の誘導には p30 が密接に関 係していることが判明した。しかしながら、p30 の Tp 特異的 Th1 細胞型免疫応 答誘導に関連する領域については明らかではない。そこで次章では p30 配列の 合成ペプチドを作製し、Th1 細胞型免疫応答誘導に関連する抗原領域を検討す る。
小活
TLA 中の存在が明らかとなった p24 と p30 の Th1 細胞型免疫応答誘導を比較 評価するため、rp24 と rp30 感作マウスの脾臓内 T 細胞と付着性細胞の各感作抗 原刺激後における IFN-γ および IL-12 mRNA 発現量と、両感作マウス血清中の感 作抗原反応性 IgG1 と IgG2a の抗体価を比較検討した。双方の感作マウスの脾臓 内 T 細胞と付着性細胞は、感作抗原刺激に対して IFN-γ と IL-12 mRNA を共に 発現したが、発現量は rp24 に比較して rp30 感作マウスの方が高値であった。ま た、血清中の感作抗原反応性 IgG1 と IgG2a の抗体価は、IgG1 では rp24 に比較 して rp30 感作マウスの方が低値を示し、IgG2a では rp24 に比較して rp30 感作マ ウスの方が高値を示した。以上の結果から、TLA の Th1 細胞型免疫応答誘導能 は、p24 に比較して主に p30 が担っているものと推察された。
Ⅲ章
Th1 細胞型免疫応答を誘導する p30 (SAG1) ペプチド領域と構造的特徴
序論
前章では、p24 (GRA1) と p30 (SAG1) の感作マウス脾臓内 T 細胞と付着性細 胞に対する IFN-γ ならびに IL-12 mRNA 発現誘導能および感作抗原特異的 IgG2a 抗体誘導能を検討したところ、TLA の Th1 細胞型免疫応答誘導能は主に p30 が 担っているものと推察された。
抗原タンパク質による Th1 細胞型免疫応答誘導には、Th1 細胞型免疫応答誘 導能を持つ抗原のペプチドが、抗原提示細胞の主要組織適合遺伝子複合体 (Major Histocompatibility Complex, MHC)と結合し、ヘルパーT 細胞へ提示され ることが重要と考えられている (21, 24, 39)。したがって、Th1 細胞型免疫応答 誘導能を持つ p30 のペプチド領域とその構造が明らかとなれば、今後 Tp 感染予 防のためのワクチン開発などに有用な情報をもたらすものと考えられる。しか しながら、p30 の Tp 特異的 Th1 細胞型免疫応答を誘導するペプチド領域を検討 した報告は、現在のところ認められないのが現状である。 そこで本章では、p30 のシグナル配列を除くアミノ酸全領域に対し、1 ペプチ ドあたり 20 アミノ酸残基数程度になるように 20 本の合成ペプチドを作成し、
rp30 感作マウス血清中 IgG2a が高い反応性を示す合成ペプチドと rp30 感作マウ ス脾臓内 T 細胞に高い IFN-γ 産生を誘導する合成ペプチドを検討し、Tp 特異的 Th1 細胞型免疫応答誘導能に密接に関係する領域を推測した。また、その推測 された領域について Chou-Faman 法 (7) によるパラメーターから 2 次構造モデ ルを予測し、その領域の抗原性と Th1 細胞型免疫応答誘導能との関係を考察し た。
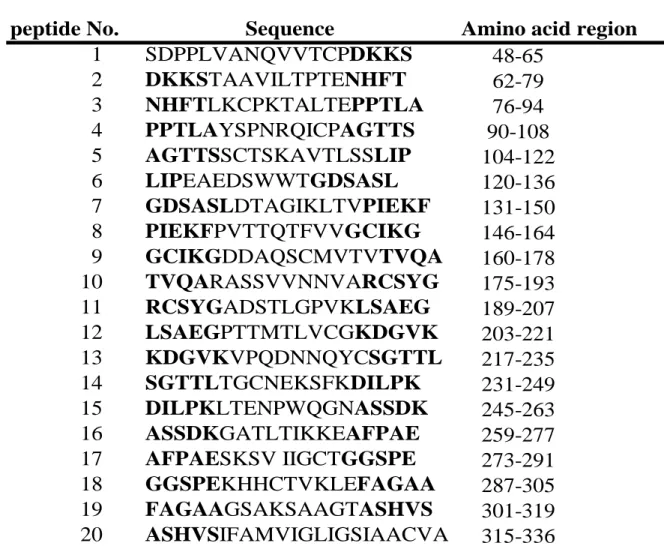
材料及び方法 1. マウス BALB/c 系の 6‐8 週齢雌マウス (日本クレア、東京) を実験に供した。 2. p30 組み換えタンパク質の作製 Ⅰ章で調製した p30 の組み換えタンパク質 (rp30) を実験に供した。 3. rp30 抗原感作マウスおよび rp30 抗原感作マウス血清の作製 Ⅱ章と同様にマウスの腹腔内に rp30 を 50 μg (100 μg/ml 水溶液) を投与し、投 与 2 週間後に同量の抗原をさらに投与して 2 週間を経過したものを rp30 抗原感 作マウスとして実験に供した。またこの抗原感作マウスから全採血した血清を rp30 抗原感作マウス血清として用いた。 4. ペプチド合成 本 実 験 に 用 い た p30 合 成 ペ プ チ ド は 、 p30 タ ン パ ク 質 (Accession No. CAA32244) の全長配列から、N 末端のシグナル配列 (アミノ酸 No. 1-47) を除 外した配列を対象として合成した。親水-疎水性アミノ酸の割合、合成効率、等 電点などを考慮し、前後配列の N 末端側および C 末端側の 3 から 6 アミノ酸ず
つオーバーラップさせ、17 から 22 アミノ酸残基数となる様に解析ソフト Genetyx(Genetyx、東京)を用いてデザインした。設計した合成ペプチドのアミ ノ酸配列は表 2 に示した。ペプチド合成は、Fmoc 法による固相合成で行った(シ グマアルドリッチジャパン、石狩)。 5. p30 合成ペプチドに対する rp30 抗原感作マウス血清中 IgG1 および IgG2a の 反応性の検討 マイクロタイタープレート (MaxisorpTM
, NalgeNunc, USA) の各 well に、0.05
M 炭酸ナトリウム緩衝液 (pH 8.5) にて 2 μM になるように溶解させた各合成ペ プチド溶液を 100 μl ずつ分注して室温で 2 時間固相化した。固相化後のブロッ キング操作はⅡ章と同様の方法で行った。rp30 抗原感作マウス血清を T-PBS に て 1,000 倍希釈し各ペプチド固相化 well に反応させた (37℃、1 時間)。洗浄の 後、西洋ワサビペルオキシダーゼ標識抗マウス IgG1 ならびに IgG2a を T-PBS に て 3,000 倍希釈し、各 well に 100 μl ずつ分注して反応させた (37℃、1 時間)。 反応後、洗浄し、発色基質 (TMB) を各 well に 100 μl ずつ添加し、室温で 10 分 放置後、1N H2SO4 にて発色を停止させた。発色は、450 nm の波長で測定した。 詳細はⅡ章の間接 ELISA 法と同様の方法で実施した。
6. 間接競合 ELISA 法による rp30 感作マウス血清の合成ペプチドに対する親和 反応の確認 rp30 感作マウス血清の p30 合成ペプチドに対する親和反応を確認するために、 間接競合 ELISA 法により検討を行った。親和反応の対象となる p30 合成ペプチ ドは、上述した項目 5 に準じてペプチド固相化プレートを作成した。Ⅱ章と同 様にブロッキング操作を行い、競合反応用抗原となるペプチドを 2,000、500、 125、31.25、7.8、1.95、0 nM になるように PBS で希釈したものを各 well に 50 μl ずつ添加した。更に、この各 well に rp30 感作マウス血清を T-PBS にて 1,000 倍 に希釈したものを 50 μl ずつ加えた。1 時間,37℃で反応後,T-PBS にて 3 回洗 浄を行い,西洋ワサビペルオキシダーゼ標識抗マウス IgG2a を 3,000 倍希釈して 各 well に 100 μl ずつ加えた。1 時間、37 ℃で反応後、同様に T-PBS で洗浄を行 った後、発色基質である TMB を各 well に 100 μl ずつ添加し、室温で 10 分放置 後、1 N H2SO4 にて発色を停止させた。発色は、450 nm の波長で測定した。抗 原を添加しない well の OD 値 (=Bound 0 nM) ; B0) に対して各濃度の抗原添加を 行った well (B) の OD 値を B/B0 として各々計算し、4 係数ロジスティックによ る計算式から標準曲線を求め、抗原未添加に対する各抗原濃度添加の競合反応 比率を求めた。
7. p30 ペプチド刺激脾臓内 T 細胞の IFN-γ 産生能の検討 Ⅰ章と同様の方法で未感作あるいは抗原感作マウスから脾臓内 T 細胞ならび に付着性細胞を調製して実験に供した。準備した脾臓内 T 細胞及び付着性細胞 は、10% FBS 加 RPMI で各々2×106 個/ml、1×105 個/ml に調整し、96 穴マイク ロプレートの各 well に 50 μl ずつ分注した。対照群ならびに各ペプチド添加群を 設け、それぞれ対照群では rp30 を終濃度 1.0 μg/ml、ペプチド添加群では RPMI に溶解した合成ペプチドを終濃度 5 μM となるように添加し、 37℃、5% CO2 で 3 日間培養を行った。培養後の上清を IFN-γ の測定に供した。測定は測定キ ット (Biosource, USA) 付属の取り扱い説明書に従い実施した。まず、回収した 培養上清を、抗マウス IFN-γ 抗体を塗布されたマイクロプレートに 50 μl 添加し、 室温で 1 時間培養した。培養後、プレートを T-PBS で 3 回洗浄し、ビオチン化 抗サイトカイン溶液を 50 μl 添加して更に室温で1時間培養した。 再度洗浄し、 西洋ワサビペルオキシダーゼ標識ストレプトアビジン溶液を 50 μl ずつ各 well に添加した。室温 1 時間培養後、T-PBS で洗浄 (3 回) し、発色基質である TMB を 50 μl ずつ添加、室温 10 分間反応後 1 N H2SO4を各 well に 50 μl 添加して反応 を停止させた。マイクロプレートリーダーで 450 nm の波長を測定した。測定は 2 重測定で実施し、各データは平均と標準偏差を算出した。
8. 二次構造予測
p30 アミノ酸配列の二次構造予測ならびに親水-疎水性領域の検索には、解析 ソフト Genetyx (Genetyx、東京)、並びに Uniprot (http://www.uniprot.org/ uniprot/) を用いた。また膜貫通領域の解析には SOSUI (http://sosui.proteome. bio.tuat.ac.jp) を用いた。
結果
1. p30 合成ペプチドに対する rp30 感作マウス血清中 IgG2a および IgG1 の反応 p30 由来合成ペプチド領域に反応する rp30 感作マウス血清中における IgG2a および IgG1 を比較した。血清中 IgG2a は、peptide 6 (アミノ酸 No. 120-136) と peptide 15 (アミノ酸 No. 245-263) に顕著に反応しており、OD 値は双方ともそれ 以外のペプチドの値に比較して有意に高値を示していた (p < 0.01、図 9-A) 。 一方、血清中 IgG1 は peptide 15 のみに顕著に反応し、OD 値はその他のペプチ ドに対する値に比較して有意に高値を示していた(p < 0.01、図 9-B)。
2. rp30 感作マウス血清中 IgG2a の p30 由来合成ペプチドに対する親和性
1.の結果において rp30 感作マウス血清中 IgG2a が顕著に反応を示した peptide
6 ならびに 15 に対する rp30 感作マウス血清中 IgG2a の親和性を競合反応により 検討した。50 %競合阻害を示す peptide 抗原の濃度 (50% inhibitory concentration; 以下 IC50) は、peptide 6 では約 100 nM、peptide 15 では約 500 nM であり、peptide 6 の方が peptide 15 に比較して阻害濃度が低く (図 10)、さらに競合阻害反応が 生じシグモイド形状を示していることから、生体は rp30 の peptide 6 の領域を抗 原決定基として高く認識し、rp30 感作により生体に産生された IgG2a の親和力 も peptide 6 の方が peptide 15 と比較して高いものと推察された(47,48)。
3. p30 由来合成ペプチド刺激に対する rp30 感作マウス脾臓 T 細胞の IFN-γ 産生 rp30 と各 p30 由来合成ペプチドの刺激における rp30 感作マウス脾臓 T 細胞の 培養上清中の IFN-γ 量を比較した。培養上清中の IFN-γ 量は、ほとんどのペプチ ド刺激では rp30 刺激と同等あるいはそれ以下であったが、peptide 3、4、5 なら びに 6 による刺激では、rp30 による刺激に比較して有意に高値を示し (peptide 3, 109.9± 17.2 pg/ml; peptide 4, 125.8 ± 16.3 pg/ml; peptide 5, 97.0 ± 5.1 pg/ml; peptide 6, 135.0 ± 29.0 pg/ml; rp30, 56.46 ± 24.75 pg/ml; p < 0.05) 、その中で も peptide 6 による刺激が最も高い値を示した (図 11) 。 4. 予測された活性ペプチド領域の二次構造 結果 2 および 3 で示された様に、peptide 6 は rp30 感作マウス血清中の IgG2a の反応性が高く、rp30 感作マウス脾臓内 T 細胞に対しても IFN-γ を最も強く誘 導した。この要因として peptide 6 を含む近傍の領域の立体構造が抗原性のある 領域として認識されやすい特徴があると推論した。そこで、p30 配列の二次構造 を予測するために、二次構造解析ソフトである GENETYX および Uniprot により 解析を行った。peptide 6 を含む p30 の N 末端側の領域は、β ターンが連続し電 荷を帯びるアミノ酸が多く存在していた。GENETYX による解析では、β ターン が最も予測される領域は peptide 6 を含むアミノ酸 No. 124-140 付近であり、一
方 Uniprot による解析では、アミノ酸 No. 136-139 の部位がターン構造予測部位 と算出された。更に peptide 6 の前後 3-6 アミノ酸のオーバーラップ領域を除外 した配列 (E A E D S W W T) は、親水性アミノ酸 (E, D) と疎水性アミノ酸 (A, W, T) の両方が存在していた。peptide 6 ならびに一部配列がオーバーラップして いる peptide 7 の配列内にはプロリン (P) 残基が存在していることからも、近傍 の配列は細胞表面に存在し、抗原性がある領域として認識され易いターン構造 を取る可能性が推測された。なお、p30 配列の C 末端には GPI アンカー結合部 位 (アミノ酸 No. 306-310) が存在するが、この GPI 結合予測部位以降のアミノ 酸 No. 314 から末端の 336 までの配列は、疎水性ペプチドが集中しており、SOSUI による解析では膜貫通領域として予測された。
考察 p30 は、アミノ酸配列の C 末端に GPI 骨格のエタノールアミンリン酸がアミ ド結合し、さらにその GPI のリン脂質部分が Tp の細胞表面に結合しているとい う SAG ファミリーに共通した構造をもつタンパク質である (41, 46)。p30 は Tp のタキゾイト、ブラディゾイトの表面を覆い、強毒株である RH 株、ならびに 弱毒株である P (LK) 株、Beverley 株などにおいても存在し (46)、宿主細胞への 侵入、寄生虫の宿主内における生存、免疫反応の調整 などに関わっている (66)。 Ⅱ章において、p30 は分泌型の p24 と比較して Th1 細胞型免疫応答誘導能が高 いことが推測された。そこで本章では、シグナルペプチド領域を除外した p30 全領域について、p30 の Th1 細胞型免疫応答の誘導に関与している抗原領域と、 その立体構造的特徴を検討した。 まず、Th1 細胞型免疫応答の誘導に関与している抗原領域を推測するため、 p30 由来合成ペプチドを用いて rp30 感作マウス血清中 IgG2a が反応する領域を 検討した。その結果、血清中 IgG2a は peptide 6 (アミノ酸 No. 120-136) と peptide 15 (アミノ酸 No. 245-263) 、特に peptide 6 に高い反応性を示したのに対し、血 清中 IgG1 は peptide 6 には反応せず peptide 15 のみに高い反応性を示した。また、 peptide 6 ならびに 15 に対する rp30 感作マウス血清中 IgG2a の親和性を競合反 応により検討した結果、50 %競合阻害 (IC50) を示すペプチド抗原の濃度は
peptide 6 の方が peptide 15 に比較して阻害濃度が低かった。Ngo ら (47)、 O’Sullivan らは (48)、競合阻害 ELISA においてペプチド抗原が抗体に対して親 和力を持ち、かつ抗原決定基として作用している場合の特徴として、1) 反応溶 液内において平衡状態にある抗原-抗体結合が解離、2) 添加した遊離の抗原濃 度に比例した競合阻害が生じ、グラフ形状もシグモイド曲線を呈すると報告し ている。rp30 感作血清においては、peptide 6 の方が peptide 15 と比較して、低 濃度でも競合阻害反応によるシグモイド形状のグラフを示したことから、生体 は rp30 の peptide 6 の領域を抗原決定基として高く認識し、rp30 感作により生 体に産生された IgG2a の親和性も peptide 6 の方が peptide 15 と比較して高いも のと推察された。これらのことから、rp30 感作マウス血清中 IgG2a は peptide 6 を特異的に認識するものと推測された。本論文では結果を掲載していないが、 peptide 6 には、強毒株である致死性の RH 株感染血清中の IgG2a も高い反応性 を示し (33)、弱毒株である Beverley 株感染血清中 IgG2a も peptide 1 から 6 ま での親水性に富んだ領域に高い反応性を示すことが判明している。 前章におい て、rp30 感作マウス脾臓内 T 細胞は、rp30 の刺激に対して IFN-γ mRNA を強く 発現したことから、次に rp30 と p30 由来合成ペプチド刺激に対する rp30 感作 マウス脾臓内 T 細胞の IFN-γ 産生誘導能を検討した。その結果、rp30 より有意 に IFN-γ 産生能を誘導するペプチド領域は、peptide 3 から 6 (アミノ酸 No.
76-136) であり、特に peptide 6 で顕著であった。rp30 感作マウス血清中 IgG2a において peptide 6 と共に反応が確認された peptide15 は、peptide 6 と比較しても IFN-γ 産生誘導能を示さなかった。peptide 15 は Th 細胞を提示し抗体産生を補 助する活性は示すものの、サイトカイン誘導に至るまで Th 細胞の分化には関 与していないと推測される。以上の様に、rp30 感作マウス血清中 IgG2a が peptide 6 に親和性が高く特異的に結合し、peptide 3 から 6 が rp30 感作マウス脾臓内 T 細胞に高い IFN-γ 産生を誘導することから、Tp 特異的 Th1 型細胞免疫応答誘導 には p30 の peptide 3 から 6 を含む N 末端側の前半領域が密接に関係しているこ とが推察された。 petide 1 から 6 (No. 48 から 136) までの範囲の N 末端領域は、β ターンが連続 するため親水性領域に富み、電荷を帯びるアミノ酸も多数存在し、抗原性とし て高く認識される可能性が推測された。この検索結果は、p30 には β 構造を取り やすい配列が多く存在し、p30 が β タンパク質に分類されている報告とも一致す る (16)。また、GENETYX による解析では、T 細胞エピトープとして予測され る領域が、peptide1 から 7 までの範囲に集中していた。T 細胞が抗原認識を高め る二次構造上の特徴としては、その配列内に親水性-疎水性、あるいは α へリッ クス-β シート構造あるいはターン構造を複合して持つと報告されている (18, 46)。peptide 6 配列のうち、前後 3-6 アミノ酸のオーバーラップ領域を除外した
配列 (E A E D S W W T) はこの特徴を有しており、電荷を帯びるアミノ酸が存 在し (E, D) 、更に親水性アミノ酸 (E, D) と疎水性アミノ酸 (W, T) で占められ ていた。また peptide 6 ならびに一部配列がオーバーラップしている peptide 7 の 配列内にはプロリン (P) 残基が存在しており、GENETYX による二次構造解析 では、細胞表面に存在しやすく抗原性を持ちやすいターン構造を取る可能性が 推測された。更に Uniprot による解析においても、p30 配列内でターン構造が最 も高頻度で予測される部位として、peptide 6 配列近傍のアミノ酸 No. 136-139 を示した。また peptide 6 配列前後にはリジン (K) 残基が存在するが、C 末端に リジン残基をもつ領域は、抗原がプロセッシングの過程で酵素消化を受けた後 でもαへリックスの構造が安定するという特徴があるため、ペプチドワクチン としての機能を有する可能性が高いとされる (57)。以上より、p30 の peptide 6 を含む N 末端側の領域は、二次構造においてもターン構造が配列近傍に存在し、 また溶解性が高いアミノ酸を多数持つなど、抗原性領域として認識される要素 を多く含み、Th 細胞型免疫応答の誘導に重要な配列である可能性が示された。
小活 p30 の Th1 細胞型免疫応答誘導に関連する抗原領域とその構造を明らかにす るため、シグナルペプチド領域を除外した p30 全領域について、1 つのペプチド あたり 20 アミノ酸残基数程度でなおかつ前後配列を数残基ずつ重複するように デザインした 20 本の合成ペプチドを作成し、rp30 感作マウス血清中 IgG2a の反 応性と rp30 感作マウス脾臓内 T 細胞に対する IFN-γ 産生誘導能を検討した。rp30 感作血清中 IgG2a が最も高く反応したペプチドは peptide 6 であった。また、 peptide 3 から 6 のペプチドで rp30 感作マウス脾臓内 T 細胞を刺激した場合にお いては、rp30 で刺激した場合と比較して有意に IFN-γ の産生を誘導した。さら に、p30 の N 末端領域の peptide1 から 6 までの範囲は、β ターン構造や T 細胞エ ピトープ予測領域が集中しており、特に peptide 6 の配列近傍は T 細胞が抗原認 識を高める二次構造上の特徴を有していることが判明した。以上の結果から、 p30 の peptide 6 を中心とした N 末端側の配列が p30 の Th1 細胞型免疫応答の誘 導に密接に関わる領域であることが推測された。