熊本大学学位論文
間葉系幹細胞の心血管細胞保護 メカニズムに関する研究
2017 年 舘田 悠樹
A research on the mechanisms of cytoprotective effects of mesenchymal stem cells on cardiovascular cells
Yuki TACHIDA
A research on the mechanisms of cytoprotective effects of mesenchymal stem cells on cardiovascular cells
Yuki TACHIDA
Mesenchymal stem cells (MSCs) are tissue stem cells capable of differentiating into mesenchymal lineage cells such as osteoblast, chondrocyte and adipocyte, and are isolated from various tissues including bone marrow, adipose tissue and dental pulp. MSCs have been applied to regenerative medicine and cell therapy from a long time ago, since they have proliferative ability and multilineage potential. Actually, MSCs have been reported to show therapeutic effects on a wide range of diseases such as bone and cartilage damage, nerve injury and cardiovascular disease. In addition, MSCs have low immunogenicity and appear to evade allogeneic rejection, indicating that it is possible to clinically apply formulated allogeneic MSCs as well as autologous MSCs. Many clinical trials have been tried to determine whether MSCs have a therapeutic effect on diseases such as myocardial infarction, which are difficult to treat with drugs. In fact, administration of MSCs has been reported to exert a certain therapeutic effect. The therapeutic action of MSCs was initially supposed to be mediated by migration into the damaged tissue and the following differentiation into appropriate cells. Recently, it has been indicated that factors secreted from MSCs and cell-cell interaction factors on MSCs play an important role in cytoprotective effects of MSCs. However, the cytoprotective molecules from MSCs and detailed mechanisms of cytoprotection are still not clear. This is an obstacle to standardize and evaluate the quality of formulated MSCs.
Therefore, the aim of this study is to identify the secreted protein possessing therapeutic effects of MSCs and to clarify the molecular mechanisms of cytoprotection by MSCs. In this study, I identified the candidate proteins exerting cardioprotective effects by comprehensive analysis of proteins secreted from various MSCs. Moreover, I found the possibility that vasoprotective effects of MSCs are attributed to direct cell-cell interaction between vascular endothelial cells and MSCs.
I. Comprehensive analysis of secreted proteins derived from MSCs
It has been reported that secreted proteins derived from MSCs are important for the therapeutic effect on myocardial infarction. By mass spectrometry, I tried to comprehensively identify the proteins included in conditioned media (CM) of three MSCs, bone marrow derived-MSCs (BMSCs), adipose tissue derived-MSCs (ASCs) and dental pulp derived-MSCs (DPSCs), all of which had cardioprotective effects. Then, I totally identified 1,533 proteins secreted in either of MSCs, and 124 proteins of 1,533 proteins were common among three MSCs. These common proteins included already well-known proteins whose functions are linked to biological effects of MSCs and newly identified proteins whose roles are not well investigated. Pathway analysis of these common proteins showed correlation with the functions of MSCs. These data indicate the possibility that the secreted proteins possessing cardioprotective effects of MSCs are included in these 124 proteins.
II. Identification of the cardioprotective protein released from MSCs
Proprotein convertase subtilisin/kexin type 5a (PCSK5A) belongs to the proprotein convertase (furin/PCSK) family and is responsible for the conversion into active proteins by the proteolytic cleavage. I focused on PCSK5A among identified 124 common proteins, since many of cardioprotective proteins possess the furin/PCSK cleavage site. I found that the recombinant furin as well as ASC-CM decreased cell death of cardiomyocytes by oxygen-glucose deprivation (OGD), an ischemia model. An inhibitor of the furin/PCSK-like enzyme abolished the cardioprotective effects in both the recombinant furin and ASC-CM, indicating that furin/PCSK enzyme activity is important for suppression of cardiomyocyte cell death induced by ischemia. In addition, knockdown of PCSK5A in ASCs decreased both the furin/PCSK protease activity and cardioprotective activity in the CM. These findings suggest the importance of PCSK5A in the anti-ischemic activity of ASCs.
III. Evaluation of the protective effect of MSCs on vascular endothelial cells through direct cell-cell interaction
Therapeutic effects of MSCs on myocardial infarction were derived from not only protection of cardiomyocytes but also angiogenesis and vascular protection in infarct sites. Since MSCs is localized in perivascular area in vivo and have phenotype of mural cells, elucidation of the role of interaction of MSCs with vascular endothelial cells (ECs) may lead to understanding of the mechanism of angiogenesis and vascular protection. I focused on the response of ECs to bone morphogenetic protein 9 (BMP9) and investigated influence of the response on direct cell-cell interaction between ECs and MSCs, since BMP9 has trophic effect on ECs and is a causative gene of angiopathies such as Osler-Rendu-Weber disease and pulmonary hypertension.
As a result, the activation of Notch pathway in ECs and MSCs was induced in a coculture- and BMP9-dependent manner. In ECs, the vascular protective genes induced by BMP9 were significantly and synergistically induced in the presence of 3 types of mural cells (pericytes, fibroblasts and MSCs). These synergistic upregulations were greatly reduced in a non-contact condition. In MSCs, the expression of platelet-derived growth factor receptor type-beta (PDGFRB), which is considered to be important for the self-differentiation and interaction with ECs, was potently induced in the coculture with HUVECs, and BMP9 additively enhanced this upregulation. Taken together, these results suggest that MSCs potentiates the BMP9 signaling in ECs by direct cell-cell interacting with ECs and may promote the vascular protective effect.
In conclusion, I elucidated a part of the molecular mechanisms of cardioprotection mediated by secreted factors from MSCs and of vascular protection mediated by direct cell-cell interaction between ECs and MSCs.
These findings lead to clarify the mechanisms of MSCs therapy action, suggest the possibility of application to quality evaluation of MSCs and expansion of indication of formulated MSCs, and largely contribute safety improvement and promotion of MSCs therapy.
論文要旨
間葉系幹細胞の心血管細胞保護メカニズムに関する研究 舘田 悠樹
間葉系幹細胞(Mesenchymal stem cell: MSC)は、骨髄、脂肪、歯髄といった様々な組織に存在し、骨、
軟骨、脂肪等の間葉系細胞へ分化する組織幹細胞として広く知られている。MSC は、増殖能と分化能をもつ ことから再生医療・細胞治療に古くから応用され、骨および軟骨損傷、神経損傷、心血管疾患などの幅広い疾 患に治療効果を示すとされている。MSC は免疫原性が低く投与による拒絶反応を起こしにくいため、自己組 織から採取した細胞のみならず、製剤化された他家MSCを用いた臨床応用も可能である。とくに心筋梗塞な どの薬物治療が困難な疾患に対して、治療効果を示すかどうかの臨床試験が試みられ、実際、MSC 投与が 一定の治療効果を発揮している。MSCが治療効果を発揮する機序については、本細胞が広範な細胞への分 化能を保持することから、損傷組織への定着・分化による組織再生が本体と考えられてきた。近年、MSCに由 来する分泌因子や細胞間相互作用因子が、細胞保護作用に重要な役割を果たすことが示唆されてきたが、
その詳細なメカニズムは現在も不明である。また、MSC は採取組織や培養方法によって治療効果が異なると 指摘されているが、これは、治療効果の指標となる分子が不明瞭なことに依るところが大きく、MSC 細胞製剤 の規格化や品質評価の障害となっている。そこで、著者は、MSC 治療効果の指標となる分子を同定すること で、細胞保護作用の分子機構を明らかにすることを目指した。本研究では、MSC の分泌因子を網羅的に探 索し、心筋保護作用を発揮する候補分子を同定した。さらにMSCの血管保護作用が血管内皮との直接相互 作用に起因する可能性を見出した。
第1章:間葉系幹細胞に由来する分泌因子の網羅的探索
MSCが心筋梗塞に対して治療効果を発揮するためには、本細胞に由来する分泌因子が重要であると報告 されている。そこで著者は、骨髄、脂肪、歯髄の3種類の組織からMSCを採取し、培養上清中のタンパク質を 網羅的に探索し、3種の細胞に共通する心筋保護活性成分を同定した。
ラットの骨髄、脂肪、歯髄よりMSCを単離・培養し、その培養上清における心筋保護活性をin vitro虚血再 灌流障害モデルにより評価した。その結果、3種類のMSCの培養上清は、いずれも虚血再灌流により誘導さ れる心筋細胞死を抑制した。そこでLC-MS/MSを用いて3種類の培養上清中のタンパク質を網羅的に解析 したところ、1,533種のタンパク質を同定し、このうち共通して検出された分泌タンパク質は124種類であった。
これらの分泌因子の中には、MSC の分泌因子として既報の因子だけでなく、新規因子も多数含まれた。培養 上清中に含まれる分泌タンパク質を Western blotにより定量したところ、3 種類のMSC における存在量は
LC-MS/MSの定量結果とよく相関した。124種類の検出タンパク質をパスウェイ解析すると、MSC機能との相
関が認められた。従って、今回同定したタンパク質の中には MSC の細胞保護活性を発揮する分泌因子が含 まれる可能性があると考えられた。
第2章:間葉系幹細胞に由来する心筋保護分泌因子の同定
前章で同定した MSC 共通の 124 の分泌因子のうち、PCSK5A に注目した。PCSK5A は furin/PCSK (proprotein convertase subtilisin/kexin) familyに属するserine proteaseであり、さまざまな前駆タンパク 質を切断することで成熟体へと変換し、その活性発現に重要な役割を果たす。MSCが分泌する多くの心筋保 護性因子がPCSK切断配列をもつことから、培養上清中のfurin/PCSK酵素活性がMSCの心筋細胞死抑 制に重要であると考えた。まずin vitro虚血再灌流障害モデルにおいてrecombinant furinの影響を調べ た結果、recombinant furin は単独で心筋細胞死を抑制した。また、脂肪由来 MSC の培養上清は、
furin/PCSK 活性を有し、その心筋細胞死抑制作用は furin 阻害剤の添加で消失した。次にマイクロアレイ
およびWestern blot解析により脂肪由来MSCの培養上清中に含まれるPCSK family タンパク質7種類 の発現を調べたところ、PCSK5Aが最も多く発現していた。脂肪由来MSCのPCSK5A発現をノックダウンし たところ、培養上清中のfurin/PCSK活性と心筋細胞死抑制作用はともに低下した。これらの結果から、MSC の心筋保護作用には、本細胞に由来するfurin/PCSK活性が重要であると考えられた。
第3章:間葉系幹細胞との直接相互作用による血管内皮保護作用の評価
心筋梗塞における MSC の治療効果には、心筋細胞の保護作用以外に、梗塞部位における血管新生・保 護作用が重要である。MSC は生体内において血管内皮周囲に存在し、血管内皮周囲細胞としての形質を持 つことから、血管内皮細胞との相互作用の役割解明が、MSCの血管新生・保護作用メカニズムの理解への早 道である。Bone Morphogenetic Protein 9 (BMP9) は、Transforming growth factor-βファミリーに属す る増殖因子であり、血管内皮の栄養因子として作用すると考えられている。実際、BMP9 は、オスラー病や肺 高血圧症など血管異常症の原因遺伝子として知られる。そこで著者は、BMP9 による血管内皮応答に着目し、
MSCによる直接相互作用が本応答に及ぼす影響を調べた。
まず、ヒト臍帯静脈内皮細胞 (HUVEC) とヒト骨髄由来MSCとの共培養系を用いて、BMP9を本培養液 中に添加した。BMP9は両細胞を共培養した場合、いずれの細胞においてもNotch シグナル経路を相乗的 に活性化した。HUVEC単独培養でも、BMP9は幾つかの血管新生・保護因子を誘導したが、このBMP9に よる発現誘導はMSCと共培養することにより亢進した。MSCの代わりに血管周囲細胞である線維芽細胞、ペ リサイト、血管平滑筋細胞を共培養しても、内皮の BMP9 応答は増強された。血管周囲細胞による内皮 BMP9応答の増強作用は、非接触性の共培養では見られなかった。次に、MSCの分化や血管内皮との相互 作用に重要とされるPlatelet-derived growth factor receptor type-beta (PDGFRB) の発現を調べたとこ ろ、MSCにおけるPDGFRB 発現は HUVECとの共培養で誘導され、BMP9添加で増強された。従って、
MSCは、血管内皮と相互作用することで、内皮におけるBMP9シグナルを増強し、血管保護作用を促進する ものと考えられた。
以上、本研究で、著者は、MSC の分泌因子を介した心筋細胞保護ならびに内皮との直接相互作用による 血管保護の分子機序の一端を明らかにした。これらの知見は、MSC による細胞治療の作用機序解明の端緒 となるのみならず、MSC の品質評価への応用や他の疾患への適用拡大の可能性を示唆するものであり、
MSC治療の安全性向上や普及促進に大きく寄与するものである。
本稿は以下の論文に発表した内容を基礎とするものである
Proteomic comparison of the secreted factors of mesenchymal stem cells from bone marrow, adipose tissue and dental pulp
Journal of Proteomics & Bioinformatics, 8, 266-273 (2015)
Yuki Tachida, Hidetaka Sakurai, Junichi Okutsu, Koji Suda, Ryusuke Sugita, Yumiko Yaginuma, Yuji Ogura, Kohei Shimada, Fujio Isono, Kazuishi Kubota, Hideki Kobayashi
Secreted factors from adipose tissue-derived mesenchymal stem cells suppress oxygen/glucose deprivation-induced cardiomyocyte cell death via furin/PCSK-like enzyme activity
Biochemistry and Biophysics Reports, 7, 266-272 (2016)
Yuki Tachida, Koji Suda, Hiroyuki Nagase, Kohei Shimada, Fujio Isono, Hideki Kobayashi
Mutual interaction between endothelial cells and mural cells enhances BMP9 signaling in endothelial cells
Biology Open., 6(3), 370-380 (2017)
Yuki Tachida, Nanae Izumi, Toyo Sakurai, Hideki Kobayashi
目次
略語 緒論
第1章 間葉系幹細胞に由来する分泌因子の網羅的探索 1-1 序論
1-2 結果 1-3 考察
第2章 間葉系幹細胞に由来する心筋保護分泌因子の同定 2-1 序論
2-2 結果 2-3 考察
第3章 間葉系幹細胞との直接相互作用による血管内皮保護作用の評価 3-1 序論
3-2 結果 3-3 考察 総括
実験の部 参考文献 謝辞
1 略語
本文中、図表中で使用した略語は以下の通りである
ACVRL1 :activin A receptor like type 1
ADAMs :a disintegrin and metalloproteinases
AIMP1 :aminoacyl tRNA synthetase complex interacting multifunctional protein 1 ALK1 :activin A receptor like type 1
ASC :adipose tissue derived-MSC ASC-CM :ASC-conditioned medium BMP1 :bone morphogenetic protein 1 BMP9 :bone morphogenetic protein 9 BMP10 :bone morphogenetic protein 10 BMPER :BMP binding endothelial regulator BMPR2 :BMP type-II receptor
BMSC :bone marrow derived-MSC C1QTNF3 :C1q and TNF related 3
CADASIL :cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy CCL2 :C-C motif chemokine ligand 2
CDH5 :cadherin 5
CLEC11A :C-type lectin domain containing 11A
CM :conditioned medium
Ct :threshold cycle
CTGF :connective tissue growth factor
DKK3 :dickkopf WNT signaling pathway inhibitor 3 DLL1 :delta like canonical Notch ligand 1
DLL4 :delta like canonical Notch ligand 4 DMEM :Dulbecco's modified eagle medium DPBS :Dulbecco's phosphate-buffered saline DPSC :dental pulp derived-MSC
DTT :dithiothreitol
EC :endothelial cell
2 EDTA :ethylenediaminetetraacetic acid EGF :epidermal growth factor EGR1 :early growth response 1
ENG :endoglin
FBS :fetal bovine serum FGF-2 :fibroblast growth factor 2
FMOD :fibromodulin
FN1 :fibronectin 1
FSTL1 :follistatin like 1
GAPDH :glyceraldehyde-3-phosphate dehydrogenase GAS6 :growth arrest specific 6
GDF-15 :growth differentiation factor 15 HAEC :human aortic endothelial cell HDGF :heparin binding growth factor
HEPES :4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid HEY1 :hairy/enhancer-of-split related with YRPW motif protein 1 HHT :hereditary hemorrhagic telangiectasia
HIF1α :hypoxia inducible factor 1 alpha subunit HRP :horseradish peroxidase
HTRA1 :HtrA serine peptidase 1
HUVEC :human umbilical vein endothelial cell IGF-1 :insulin like growth factor 1
IL-6 :interleukin 6
INHBA :inhibin beta A subunit IPA :ingenuity pathway analysis
JAG1 :jagged 1
JAG2 :jagged 2
LC-MS/MS :liquid chromatography-tandem mass spectrometry Lefty :left-right determination factor
LTBP :latent transforming growth factor beta binding protein MACS :magnetic-activated cell sorting
MC :mural cell
3 MEMα :minimum essential medium alpha MGP :matrix gla protein
MMPs :matrix metalloproteinases MSC :mesenchymal stem cell MYDGF :myeloid derived growth factor NOV :nephroblastoma overexpressed NPC1 :Niemann-Pick disease type C1 protein NPC2 :Niemann-Pick disease type C2 protein OGD :oxygen glucose deprivation
PAH :pulmonary artery hypertension PCSK :proprotein convertase subtilisin/kexin PCSK5 :proprotein convertase subtilisin/kexin type 5 PDGF :platelet derived growth factor
PDGFRB :platelet derived growth factor receptor beta
qRT-PCR :quantitative reverse transcription polymerase chain reaction
RBP-Jk :recombination signal binding protein for immunoglobulin kappa J region SDS-PAGE :sodium dodecyl sulfate-poly acrylamide gel electrophoresis
SERPINE1 :serpin family E member 1 SERPINF1 :serpin family F member 1 SERPINH1 :serpin family H member 1 SIGLEC9 :sialic acid binding Ig like lectin 9 SMC :smooth muscle cells
SOD3 :superoxide dismutase 3
TBST :tris-buffered saline containing 0.1% Tween 20 TGF- :transforming growth factor beta
THBS1 :thrombospondin 1
TMEM100 :transmembrane protein 100 TSG-6 :TNF alpha induced protein 6
TWIST1 :twist family bHLH transcription factor 1 VEGF :vascular endothelial growth factor Wnt3a :Wnt family member 3A
4 緒論
幹細胞とは自己増殖と多分化能を有する細胞であり、胚性幹細胞(ES細胞)や人工多能性細胞(iPS 細胞)などがよく知られている。生体内にも恒常性維持のために自己修復能を有している組織幹細胞 と呼ばれる幹細胞が存在する。組織幹細胞の中で、近年広く研究が行われ、再生医療・細胞治療の細 胞ソースとして注目されている細胞に間葉系幹細胞 (Mesenchymal stem cell: MSC) がある。MSC はヒト骨髄にその存在が確認されてから、現在では様々な組織に存在することが明らかになってきた。
MSCは自己複製能、骨芽細胞、軟骨細胞、脂肪細胞への分化能、複数の表面抗原の発現により定義さ れており、骨髄、脂肪組織、歯髄、滑膜、胎盤といった組織から単離・培養され、広く研究に用いら れている (Figure 1)。
Figure 1. Origin and characteristics of MSC
MSCは低侵襲に採取可能な組織より単離培養が可能なこと、自己増殖能・分化能をもつことから再 生医療・細胞治療に古くから応用されてきた。MSCは免疫原性が低く投与による拒絶反応を起こしに くいため、自己組織から採取した自家MSCだけでなく、他家MSCを用いた臨床応用も可能であるこ ともその一因となっている。当初は MSC を分化させた細胞を移植することで損傷組織を修復するこ とが試みられたが、MSCの全身投与により様々な薬効を示すことが明らかとなってきた。液性因子に よる免疫調節作用の報告は多く、すでに製剤化された MSC の全身投与が移植片対宿主病の治療に用 いられ始めている。また骨軟骨損傷、神経損傷、心血管疾患など他の疾患にも幅広く薬効を示すとさ れ、多くの臨床試験が試みられ功を奏している。薬物治療が難しい心筋梗塞については特に MSC 治 療の応用が期待されている。近年心筋梗塞後の生存率は改善しつつあるが、心筋細胞の脱落の進行や
5
慢性心不全が重大な課題として残っている1)。MSCを用いた細胞治療や細胞移植は心筋細胞の補充や 心筋梗塞後の損傷心臓の心機能を改善する新しい治療の選択肢として有望視されている。非臨床試験 だけでなく、臨床試験においても MSC が心筋梗塞後の心機能を改善させ、損傷心臓の再生を促進す ると報告されている2), 3)。
このように様々な薬効を示すとされるMSCだが、その治療メカニズムについては不明な点が多い。
MSCは広範な細胞への分化能を保持することから、損傷組織への定着・分化による組織再生が薬効の 本体と考えられてきた。実際心筋梗塞において梗塞・障害部位へ MSC が遊走し、心筋細胞や血管内 皮細胞へ分化するとの報告もある。一方近年 MSC が心筋細胞、神経細胞、膵β細胞、血管内皮細胞 など様々な細胞に対して細胞保護効果を示すとされている。またMSC培養上清がMSC投与と同様の 効果を示すことから、これら細胞・組織保護効果において、MSCの分泌因子や細胞間相互作用因子が 重要な役割を果たすことが示唆されている (Figure 2)。心筋保護作用を示すような分泌因子(抗炎症 サイトカインや成長因子といった分泌因子)が MSC 薬効の本体であるとする報告も多数あるが、こ れら因子はMSC薬効を代替するまでには至っていない。MSCは採取組織や培養条件により薬効が異 なることが指摘されているが、心筋細胞保護効果は骨髄由来、脂肪組織由来、歯髄由来の MSC で共 通して認められている。そこで第1章では心筋保護作用を示すMSCの共通分泌因子に着目し、MSC の細胞保護作用の分子メカニズムを明らかにする目的で培養上清中のタンパク質の網羅的解析を行い、
MSC 共通分泌因子の同定を試みた。第 2 章では同定したタンパク質の中から PCSK5 に着目しその MSCの心筋保護作用への関与について検討した。
Figure 2. Multiple effects of MSC-secreted factors
6
MSCの組織・細胞保護効果において、心筋細胞以外にも血管への保護作用が注目されている。心筋 梗塞へのMSCの移植・投与により梗塞部位の血管数が増加することが報告されている2),4)。MSCは 血管新生因子であるVEGFの分泌や、内皮細胞との相互作用により血管を再生・維持する可能性が示 唆されている。また MSC は下肢虚血や脳虚血に対しても血流改善作用を示すことが報告されている
5),6)。MSCは生体内において血管周囲に存在することからも、VEGF等の液性因子の分泌だけでなく、
血管内皮細胞との細胞間相互作用により、血管形成、維持に重要な役割を果たしていると考えられて いる。しかしながら細胞投与によるMSCの血管内皮への作用については不明な点が多い。
近年血管内皮細胞 (Endothelial cell: EC)とECの周囲を覆う間葉系細胞である壁細胞 (Mural cell:
MC) との相互作用が血管のintegrityの維持に非常に重要であるとの報告が多くなされている7), 8), 9),
10) (Figure 3)。MCとはペリサイトや血管平滑筋細胞といった内皮細胞を裏打ちする細胞の総称であ
り、MSCもその局在、マーカー発現、形質の観点からMCとして定義されるため、ECとの相互作用 研究においてもよく用いられている11), 12)。EC、MC 2種類の細胞の相互作用の破綻は糖尿病性網膜症、
静脈奇形、遺伝性の脳卒中といった疾患への関与が示唆されている10), 13), 14)。EC-MCの相互作用は、
直接または分泌因子を介して、様々な細胞の機能や、遺伝子発現、細胞増殖・分化を制御しており、
特にBMP9シグナルがEC-MCの相互作用に重要な役割を果たしているとされている8), 14)。そこで第 3 章では、血管形成・保護における MSC のメカニズムを解明することを目的とし、血管内皮細胞と MSCを含むその周囲細胞の細胞間相互作用とBMP9経路のクロストークに注目した検討を行った。
Figure 3. Direct cell-to-cell interaction between EC and MC
7 第1章
間葉系幹細胞に由来する分泌因子の網羅的探索
1-1 序論
間葉系幹細胞(Mesenchymal stem cell : MSC)は骨髄、脂肪、歯髄といった様々な組織から単離 され、間葉系細胞への分化能、増殖能を持つ 15)。MSC は再生治療における細胞ソースとして有望と され、特に骨軟骨疾患、皮膚創傷、神経損傷、心血管疾患における再生治療への応用が期待されてい る16)。当初、移植MSCの治療効果はMSCの損傷組織への遊走と、適切な細胞への分化を介してい ると考えられていたが、近年、MSCは栄養因子の分泌によって治療効果を発揮するとの報告が多数な されている17)。これらの因子は血管新生、免疫応答、組織保護を制御しており、創傷治癒、炎症反応 の軽減、虚血組織障害の保護を促進する15), 17)。
MSC細胞治療は骨軟骨疾患、自己免疫疾患、心血管系疾患等の様々な疾患に適用されている18)。細 胞治療は有望な治療方法であるが、他家細胞の場合は感染や免疫適合性の問題が、自家細胞の場合は 必要細胞数の増殖拡大に時間がかかるといった問題がある。そのためMSC治療効果がMSC由来の分 泌因子によって代替されることには大きな利点がある。このような治療因子の候補として MSC の培 養上清からVEGF、IL-6、FGF-2、TSG-6といった分泌因子が同定されたが、どの因子についても単 独で完全にはMSCの効果を代替しなかった19), 20)。このことは未同定の重要な因子が存在する、また は複数因子がMSC治療の代替には必要であることを示唆している。それ故にMSC治療の代替のため にはMSCから産生される分泌因子のプロファイルを明らかにすることが重要である。
MSCの分泌因子を同定するために、マイクロアレイ解析等を用いた網羅的遺伝子発現解析が数多く 実施されている 21)。このような細胞の遺伝子発現解析は網羅的な解析が可能だが、miRNA による翻 訳制御、局在・分泌制御、翻訳後タンパク質の修飾/切断といった種々の翻訳後制御を受けるため、
mRNAの発現プロファイルは完全にはタンパク質の発現プロファイルを反映しない22)。 従って直接 タンパク質の解析を行うことが望ましい。実際 MSC 培養上清のプロテオーム解析は報告されている が23)、同定されたタンパク質数は限られている23)。さらに異なる組織から採取したMSCの分泌因子 を比較した報告はほとんどない。
本研究では骨髄由来 MSC (Bone marrow derived-MSC: BMSC)、脂肪組織由来 MSC (Adipose tissue derived-MSC: ASC)、歯髄組織由来MSC (Dental pulp derived-MSC: DPSC) の培養上清の質 量分析により、MSC 分泌タンパク質の網羅的解析を行った。近年 DPSC はその入手のしやすさ、準 備の簡便さ、治療効果の強さから幹細胞の細胞供給源として他のMSCと同様に注目されている24), 25)。 しかしながらDPSCの分泌因子についてはタンパク質レベルで十分な解析がされていない。そこで第
8
1章ではDPSCを含むMSCの分泌因子のプロファイルを明らかにすること、共通分泌因子の機能を 特徴付けること、MSCの定義のための新しい指標になるタンパク質群を同定することを目的として実 験を行った。
9 1-2 結果
1-2-1 BMSC、ASC、DPSCの性質検討
MSCは様々な種類の治療効果を持つ多くの因子を分泌する。異なる組織由来のMSC培養上清に含 まれる分泌因子を比較するために、BMSC、ASC、DPSCの3種類のMSCを調製した。これら3種 類の細胞は線維芽細胞様の紡錘状の形態を示した。フローサイトメトリー解析により MSC の表面マ ーカーの発現を調べたところ、CD29、CD44、CD90陽性、CD45、CD11b/c、CD31陰性を示し (Figure
4A)、報告されているMSC表面マーカーのパターンと一致した 26)。次にMSC培養上清における心筋
保護効果を確かめるために、H9c2細胞を用いたoxygen glucose deprivation (OGD) 誘導性の心筋細 胞 死 評 価 系 を 構 築 し た 。 生 細 胞 が 染 色 さ れ る calcein-AM と 死 細 胞 が 染 色 さ れ る ethidium homodimer-1をH9c2細胞に同時添加し、OGD誘導性の細胞死への影響を観察した。その結果、DPSC の培養上清は細胞生存率を回復させた (Figure 4B-D)。BMSC、ASC由来の培養上清についても同様 の結果が得られた (data not shown)。これらの結果を定量するためにCellTiter-Gloを用いて細胞数 をカウントしたところ、BMSC、ASC、DPSCの培養上清はH9c2細胞の生存率を66.1%からそれぞ れ81.5%、83.4%、89.3%まで回復させた (Figure 4E)。以上より、今回採取したBMSC, ASC, DPSC は以前より報告されていたMSCと同様の性質をもつことが示された。
10 Figure 4. Characterization of three types of MSCs.
(A) BMSCs, ASCs, and DPSCs were stained with antibodies against indicated antigens (red line) and isotype control (blue line), and analyzed by flow cytometry. (B-D) H9c2 were stained by calcein-AM and ethidium homodimer-1. In the no OGD condition, living H9c2 cells were positive for calcein-AM and negative for ethidium homodimer-1 (B). OGD/reperfusion induced cell death (C), and the conditioned medium from DPSCs increased living cardiomyocytes (D). (E) Cell viability was determined using the CellTiter-Glo luminescent cell viability assay. Data are shown as the means ± SD (N = 3, ∗𝑃 <0.05 versus control by Student’s t test). Bar = 50m.
11
1-2-2 マススペクトル測定によるBMSC、ASC、DPSC培養上清のプロテオミクス解析
次にBMSC、ASC、DPSCの培養上清のマススペクトル測定によるショットガンプロテオミクス解
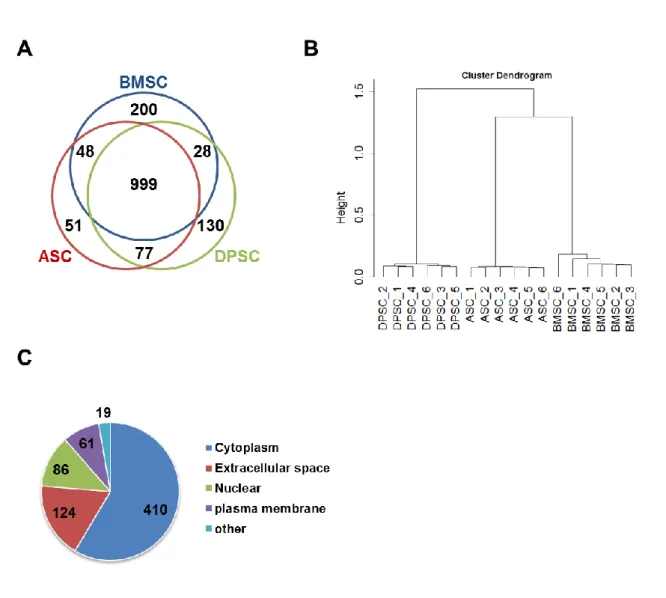
析を実施した。3種類のMSC培養上清中から全部で1,533のタンパク質が検出され、999のタンパク 質が3種類に共通して検出された (Figure 5A)。これらタンパク質群の階層クラスター解析を行った ところ、試料調製における再現性が示され、上清中のタンパク質発現パターンにおいてBMSCがDPSC よりもASCに近いことが示された (Figure 5B)。MSCに共通する分泌タンパク質に注目するために、
すべての試料(BMSC、ASC、DPSC、それぞれn = 6、計 18 サンプル)において発現が認められた、
検出信頼度の高い700のタンパク質についてIPA software によりannotation情報を付加した。その 内、発現部位が細胞外に分類されるタンパク質は124タンパク質であった (Figure 5C, Table 1)。培 養上清中の細胞外に分類されるタンパク質の割合は、概ね他のプロテオミクス解析の結果と一致した
27)。この分泌タンパク質群の中にはMSCの分泌因子としてよく知られるCTGF、SERPINE1、HTRA1、
THBS1、DKK3、MYDGF等の因子だけでなく23), 28)、AIMP1、CLEC11A、GAS6、HDGF、INHBA、
PCSK5といったMSCとの関連が報告されていないタンパク質も多く含まれていた。これらプロテオ
ミクス解析の結果を検証するために、Western blotにより8つの代表的な分泌タンパク質、CTGF、
SERPINE1、HTRA1、THBS1、FMOD、FN1、INHBA、PCSK5Aの発現を確認した。その結果、
8つすべてのタンパク質においてMSC培養上清中で発現が認められた (Figure 6)。以上の結果より、
今回同定された124のタンパク質は3種類のMSCに共通する分泌タンパク質として信頼性が高いこ とが示された。
12
Figure 5. Proteome analysis of the conditioned media from BMSCs, ASCs and DPSCs by mass spectrometry.
(A) Venn-diagram of the detected proteins in the identified datasets. (B) Unsupervised hierarchical clustering of 18 LC-MS/MS datasets. (C) The 700 proteins detected in all 18 datasets were classified by their cellular locations.
13
Table 1. Common secreted proteins identified by secretory proteome analysis of BMSCs, ASCs and DPSCs.
Major Protein ID Gene symbol Protein name
Signal intensity
BMSC ASC DPSC
XP_003749876.1 A2M alpha-2-macroglobulin 0.0279 0.0597 16.3715
XP_006248295.1 Abi3bp ABI gene family, member 3 (NESH) binding protein 0.2176 0.0028 0.0009
NP_077376.2 ADAMTS1
ADAM metallopeptidase with thrombospondin type 1 motif, 1
0.0574 0.0151 0.0166
NP_001099890.2 ADAMTS12
ADAM metallopeptidase with thrombospondin type 1 motif, 12
0.0486 0.0234 0.0064
NP_446209.1 AIMP1
aminoacyl tRNA synthetase complex-interacting multifunctional protein 1
0.0181 0.0256 0.0352
NP_112641.1 ATRN attractin 0.0371 0.0285 0.0339
NP_058783.1 BGN biglycan 8.0566 11.4552 2.7284
NP_112613.1 BMP1 bone morphogenetic protein 1 0.0089 0.0080 0.0212
XP_008769221.1 BTD biotinidase 0.0312 0.0378 0.0410
NP_001127908.1 C1QTNF3 C1q and tumor necrosis factor related protein 3 0.6521 0.0051 0.0713 NP_001128027.1 C1R complement component 1, r subcomponent 0.2321 0.5779 0.6503 NP_620255.1 C1S complement component 1, s subcomponent 0.0290 0.0571 0.0703
NP_757376.2 C2 complement component 2 0.4788 0.2556 0.7073
NP_001012477.1 CLEC11A C-type lectin domain family 11, member A 0.1712 0.0219 0.1242
NP_037249.1 COL11A1 collagen, type XI, alpha 1 4.2557 1.6553 2.4899
XP_001060689.2 COL12A1 collagen, type XII, alpha 1 3.0851 1.6315 24.2579 NP_001124020.1 COL14A1 collagen, type XIV, alpha 1 0.0185 0.0187 0.0213
XP_003749981.1
Col15a1/
LOC100909752
collagen, type XV, alpha 1 0.4020 2.3500 0.2256
XP_006239015.1 COL16A1 collagen, type XVI, alpha 1 0.0020 0.0042 0.0159
NP_445756.1 COL1A1 collagen, type I, alpha 1 47.7847 23.1591 74.0662
NP_114474.1 COL3A1 collagen, type III, alpha 1 5.7285 5.3784 7.0606
NP_001128481.1 COL4A1 collagen, type IV, alpha 1 0.2338 0.5403 0.1713
XP_008769639.1 COL4A2 collagen, type IV, alpha 2 0.0320 0.1698 0.0649
XP_006257375.1 Col4a5 collagen, type IV, alpha 5 0.0114 0.0210 0.0281
NP_604447.1 COL5A1 collagen, type V, alpha 1 1.5493 0.9353 1.5618
NP_445940.1 COL5A2 collagen, type V, alpha 2 3.5963 1.7975 4.3577
14
XP_008764201.1 COL5A3 collagen, type V, alpha 3 0.0113 0.0069 0.0122
XP_215375.5 COL6A1 collagen, type VI, alpha 1 1.3206 0.3482 0.7185
NP_001094211.1 COL6A2 collagen, type VI, alpha 2 0.6199 0.2134 0.4512
XP_006227002.1 COL6A3 collagen, type VI, alpha 3 0.7366 0.7000 0.4342
XP_233542.4 COL8A2 collagen, type VIII, alpha 2 0.0136 0.0075 0.0118
XP_008763639.1 CPQ carboxypeptidase Q 0.2642 0.4984 0.2369
NP_001099776.1 CPXM2 carboxypeptidase X (M14 family), member 2 0.0288 0.0091 0.0115 NP_076471.3 CSF1 colony stimulating factor 1 (macrophage) 0.0272 0.0632 0.0325
NP_036969.1 CST3 cystatin C 0.2887 0.4258 0.3429
NP_071602.1 CTGF connective tissue growth factor 0.1518 0.4688 0.0308
NP_112617.2 CYR61 cysteine-rich, angiogenic inducer, 61 0.0287 0.1708 0.0546
XP_006241347.1 DCN decorin 0.1304 0.0527 0.1963
XP_006230071.1 DKK3 dickkopf WNT signaling pathway inhibitor 3 1.1948 0.9247 1.4676
NP_446334.1 ECM1 extracellular matrix protein 1 0.1242 0.8841 0.1032
XP_006230806.1 EFEMP2
EGF containing fibulin-like extracellular matrix protein 2
0.0864 0.2105 0.2969
NP_001100180.1 EMILIN1 elastin microfibril interfacer 1 0.0506 0.0465 0.1624 XP_008758824.1 ERAP1 endoplasmic reticulum aminopeptidase 1 0.0315 0.0344 0.1193 XP_008768399.1 FAM114A1 family with sequence similarity 114, member A1 0.0136 0.0088 0.0211 XP_008763723.1 FAM49B family with sequence similarity 49, member B 0.0079 0.0099 0.0187
XP_008774018.1 FBLN2 fibulin 2 0.0929 0.8117 0.2353
NP_062026.2 FBLN5 fibulin 5 0.0051 0.0227 0.2153
XP_008760425.1 FBN1 fibrillin 1 0.1030 0.5587 1.0240
NP_114014.1 FBN2 fibrillin 2 0.0032 0.0013 0.0790
NP_542429.1 FMOD fibromodulin 0.0551 0.0215 0.0597
XP_006245214.1 FN1 fibronectin 1 18.0532 11.9231 7.4117
XP_006232016.1 FST follistatin 0.0103 0.0179 0.0271
NP_077345.1 FSTL1 follistatin-like 1 2.2374 2.6397 3.2233
NP_001004218.1 FUCA2 fucosidase, alpha-L- 2, plasma 0.0041 0.0093 0.0045
NP_476441.2 GAS6 growth arrest-specific 6 0.1123 0.0756 0.0333
NP_997475.1 GPI glucose-6-phosphate isomerase 0.5396 0.7572 0.8015
XP_008766203.1 GRN granulin 0.0106 0.0357 0.0131
XP_006234102.1 GSN gelsolin 0.0708 0.0695 0.1030
NP_446159.1 HDGF hepatoma-derived growth factor 0.0351 0.0294 0.0864
NP_001258221.1 HMCN1 hemicentin 1 0.0811 0.1225 0.1697
15
NP_113909.1 HTRA1 HtrA serine peptidase 1 0.1468 0.2708 0.3753
NP_036720.2 IGFBP3 insulin-like growth factor binding protein 3 0.0381 0.1236 0.0256 NP_001004274.1 IGFBP4 insulin-like growth factor binding protein 4 0.0194 0.0241 0.1587 NP_037236.1 IGFBP6 insulin-like growth factor binding protein 6 0.0187 0.0930 0.0672 NP_001013066.1 IGFBP7 insulin-like growth factor binding protein 7 1.1682 0.9196 0.7393
XP_008769938.1 INHBA inhibin, beta A 0.0649 0.0647 0.0385
XP_003753537.1 LAMA4 laminin, alpha 4 0.3678 0.5865 0.2223
XP_006240059.1 LAMB1 laminin, beta 1 0.3529 0.7615 0.3332
NP_446418.1 LAMC1 laminin, gamma 1 (formerly LAMB2) 0.4637 0.9408 0.3888 NP_063969.1 LGALS1 lectin, galactoside-binding, soluble, 1 1.0603 1.1178 1.8905 NP_114020.1 LGALS3 lectin, galactoside-binding, soluble, 3 0.0060 0.0300 0.0306 XP_008763191.1 LOC103692716 heat shock protein HSP 90-alpha 0.3629 0.6495 1.1855
NP_001099517.2 LOXL2 lysyl oxidase-like 2 0.1879 0.4050 0.2144
NP_067598.1 LTBP1
latent transforming growth factor beta binding protein 1
0.0776 0.1057 0.1217
XP_006240412.1 LTBP2
latent transforming growth factor beta binding protein 2
0.2818 1.3109 0.2345
NP_112312.1 LUM lumican 0.5346 0.0021 3.1152
XP_006243837.1 Manf
mesencephalic astrocyte-derived neurotrophic factor
0.0134 0.0108 0.0300
XP_006248588.1 Masp1 mannan-binding lectin serine peptidase 1 0.0611 0.2367 0.1086 NP_001008346.1 MESDC2 mesoderm development candidate 2 0.0054 0.0077 0.0215 NP_036943.1 MFGE8 milk fat globule-EGF factor 8 protein 0.1749 0.7081 0.3723
NP_001100629.1 MMP19 matrix metallopeptidase 19 0.0083 0.0194 0.0407
NP_112316.2 MMP2 matrix metallopeptidase 2 2.4294 1.4391 6.5126
XP_008756141.1 MYDGF myeloid-derived growth factor 0.0168 0.0172 0.0399
NP_001002851.1 NENF neudesin neurotrophic factor 0.0118 0.0129 0.0226
XP_213954.4 NID1 nidogen 1 1.5978 2.8422 1.1877
NP_001012005.2 NID2 nidogen 2 (osteonidogen) 0.0965 0.3051 0.0779
NP_110495.1 NOV nephroblastoma overexpressed 0.1144 1.0559 0.0114
NP_775141.2 NPC2 Niemann-Pick disease, type C2 0.3482 0.7171 0.5390
NP_001099573.1 OGN osteoglycin 0.3674 0.2553 0.2353
XP_006233139.1 OLFML3 olfactomedin-like 3 0.0212 0.0213 0.0368
XP_006238331.1 PAPPA
pregnancy-associated plasma protein A, pappalysin 1
0.0534 0.0054 0.0123
16
XP_006221574.1 PAPPA2 pappalysin 2 0.1094 0.1008 0.9496
NP_062110.1 PCOLCE procollagen C-endopeptidase enhancer 1.8591 2.8958 3.1640 NP_446275.1 PCSK5 proprotein convertase subtilisin/kexin type 5 0.0146 0.0373 0.0872 NP_640348.2 PLBD2 phospholipase B domain containing 2 0.0737 0.1281 0.0550 NP_001162015.1 PLTP phospholipid transfer protein 0.0296 0.0358 0.0322
NP_001101892.1 PLXDC2 plexin domain containing 2 0.0690 0.0444 0.0453
XP_006232398.1 POSTN periostin, osteoblast specific factor 3.2539 0.5683 0.4740
NP_112348.2 PROS1 protein S (alpha) 0.0740 0.0919 0.1198
NP_001177167.1 PSAP prosaposin 0.2440 0.5507 0.3535
NP_001258190.1 PXDN peroxidasin 0.3106 0.6939 0.2733
XP_006251969.1 RNASE4 ribonuclease, RNase A family, 4 0.2844 0.5855 0.3817
NP_001100048.1 SEMA3C
sema domain, immunoglobulin domain (Ig), short basic domain, secreted, (semaphorin) 3C
0.0163 0.0473 0.0433
NP_001098103.1 SEMA3D
sema domain, immunoglobulin domain (Ig), short basic domain, secreted, (semaphorin) 3D
0.1098 0.0325 0.3249
XP_008760920.1 SEMA3E
sema domain, immunoglobulin domain (Ig), short basic domain, secreted, (semaphorin) 3E
0.0419 0.0236 0.0416
NP_036752.1 SERPINE1
serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1), member 1
2.0028 7.4714 0.2493
NP_808788.1 SERPINF1
serpin peptidase inhibitor, clade F (alpha-2 antiplasmin, pigment epithelium derived factor), member 1
2.9882 3.9966 2.3547
NP_058869.1 SERPINH1
serpin peptidase inhibitor, clade H (heat shock protein 47), member 1, (collagen binding protein 1)
0.1892 0.2190 1.1739
NP_001005539.1 SMPDL3A sphingomyelin phosphodiesterase, acid-like 3A 0.0640 0.0199 0.0141 NP_037012.1 SOD3 superoxide dismutase 3, extracellular 0.1297 0.0874 0.0123 XP_008765885.1 SPARC secreted protein, acidic, cysteine-rich (osteonectin) 15.7040 19.1240 15.7550 NP_612542.1 SPON2 spondin 2, extracellular matrix protein 0.0143 0.0101 0.0422
XP_008768218.1 SPP1 secreted phosphoprotein 1 0.5236 0.0787 0.1310
XP_006251432.1 TCN2 transcobalamin II 0.3218 0.4342 0.6680
NP_067589.1 TGFB1 transforming growth factor, beta 1 0.0225 0.0140 0.0215 XP_006250510.1 TGFB2 transforming growth factor, beta 2 0.0796 0.0928 0.0260 XP_006240428.1 TGFB3 transforming growth factor, beta 3 0.0361 0.0779 0.0105
NP_001013080.1 THBS1 thrombospondin 1 2.6504 1.1193 0.3630
XP_008759460.1 THBS3 thrombospondin 3 0.0206 0.0104 0.0131
17
XP_008771256.1 TIMP1 TIMP metallopeptidase inhibitor 1 0.7176 1.3352 0.5195 NP_068824.1 TIMP2 TIMP metallopeptidase inhibitor 2 0.8361 0.7614 1.4598
XP_008761979.1 TNC tenascin C 0.2648 6.2157 0.4773
NP_001164030.1 VCAN versican 0.0188 0.0417 0.0439
NP_001014157.1 WDR1 WD repeat domain 1 0.3081 0.3749 0.4758
Signal intensity shows LFQ intensity of the protein normalized to that of apomyoglobin.
18
1-2-3 BMSC、ASC、DPSCの共通分泌因子における生物学的機能の解析
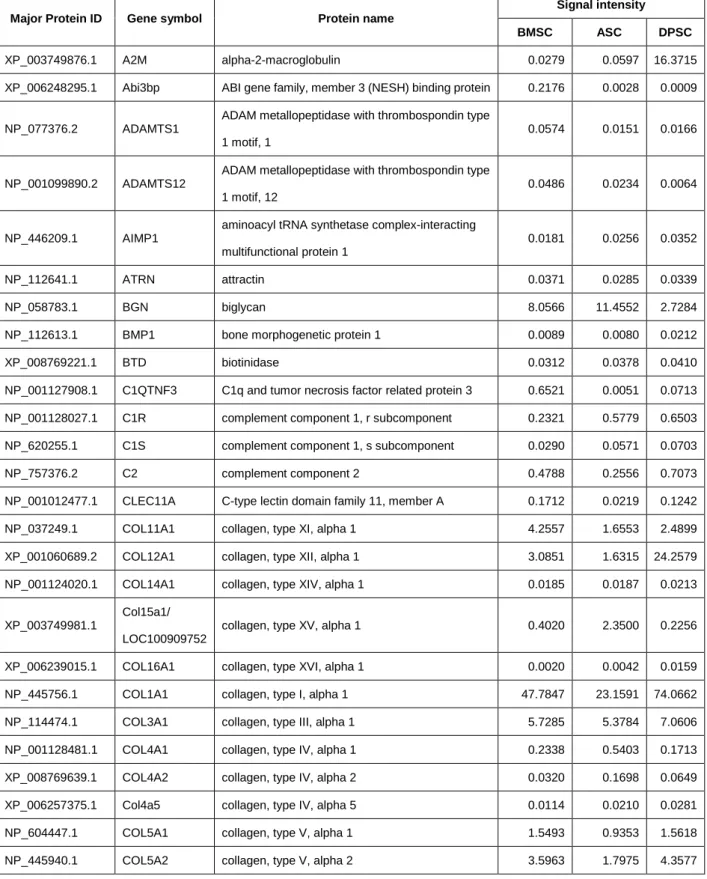
今回同定された124の分泌タンパク質の性質を調べるために、まずPanther classification system を用いてmolecular functionの分類を行った。Extracellular matrix proteinに分類されるタンパク質 が最も多く、他にもsignaling molecule、receptorやhydrolase、protease、oxidoreductaseといっ た酵素に分類されるタンパク質も多く認められた (Figure 7A)。次にこれら分泌タンパク質群と protein function との関連性について IPA software を用いて解析したところ、組織再生にかかわる angiogenesis、migration、inflammatory responseといった機能だけでなく、MSCの治療効果とし て知られるossification、sprouting、organ survival 29), 30) といった機能とも大きく関連していること がわかった (Figure 7B)。さらにIPA softwareによりupstream regulator 解析を行った結果、これ ら124の分泌タンパク質の発現制御因子の中に、MSCの形質維持に重要なTWIST1、HIF1、EGR1 が含まれることが示された (Figure 7C)。以上より、今回同定した124の分泌因子には組織再生効果 や組織保護効果などのMSCの治療効果を示す候補タンパク質が含まれる可能性が示唆された。
19
Figure 6. Validation of the mass spectrometry-based proteome result by Western blotting.
(A) The concentrated CM from BMSCs, ASCs and DPSCs were subjected to SDS-PAGE analysis, and CTGF, SERPINE1, FN1, HTRA1, FMOD, THBS1, INHBA, and PCSK5A were detected by Western blotting using each antibody. (B) The bar charts show the mean signal intensity of each protein in BMSCs, ASCs and DPSCs.
20
Figure 7. Characterization of 124 proteins identified as common secreted factors by secretory proteome analysis of BMSCs, ASCs and DPSCs.
(A) Protein classification of 124 secreted factors based on Panther. (B-C) The lists of protein functions (B) and upstream regulator (C) significantly altered were generated using the Ingenuity Pathway Analysis software.
21 1-3 考察
多くの実験や臨床試験において MSC の投与が様々な疾患に対して効果を示すことが報告されてい るが、その詳細なメカニズムについていまだに不明な点が多い。近年、MSCが損傷組織の保護や再生 を担う分泌因子を産生することで治療効果を発揮する可能性が示唆されている17)。実際今回の研究に おいても心筋梗塞のin vitroモデルであるOGD-assayにおいてMSC由来の培養上清が心筋保護作用 をもつことを示した (Figure 4E)。MSC由来の分泌因子は注目されてきているが、いくつかの問題が あるためMSCの機能を代替する因子の報告は多くない。
1つ目の問題点として、MSC由来の分泌因子の報告が限られていることが挙げられる。MSC分泌 因子は網羅的遺伝子解析、マス解析によるプロテオミクス解析、サイトカインアレイ等を用いて精力 的に解析されているが、治療効果をもつ正確な分泌因子のプロファイルについてはまだ明らかになっ ていない。著者は今回ショットガンプロテオミクスを用いて、BMSC、ASC、DPSCの培養上清中か
ら1,533タンパク質を同定した。同定数が1,000以下であった以前の報告23)と比較して、今回の実験
ではより多くのタンパク質を同定した。網羅的な分泌因子の解析がされていないDPSCも含めた3種 類のMSCの比較によりMSC共通の分泌因子を同定した。これらの分泌因子は過去の報告に治療効果 のある因子として同定されているものだけでなく、MSCの効果と直結しない新しい生理活性因子が含 まれていた。MSCの分泌因子を介した、根底にある治療メカニズムを理解する上で重要な手がかりと なると考えられる。
もう1つの生物学的に重要な因子を同定するための問題は、単一因子でなく多因子が MSC を完全 に代替するために必要である点である。例えばDPSCから産生されるCCL2とSIGLEC9の組み合わ せが脊髄損傷の治療因子として重要であることが報告されている 31)。今回MSCの共通の分泌因子と して、心筋保護作用を示すとされるCTGF、MYDGF、DKK3、FSTL1を同定した28), 32), 33), 34)。しか しながらCTGF、DKK3、FSTL1は単独ではin vitro OGD-assayにおいて細胞生存率を回復しなか った (data not shown)。今回同定したMSC共通の分泌因子群については、その組み合わせも考慮し て絞り込むことによりMSC治療のメカニズム解明に有用であると考えられる。
今回同定した因子のリストには MSC 機能との関係性が報告されていない AIMP1、C1QTNF3、
CLEC11A、INHBA、PCSK5といった因子も含まれており、MSCの治療メカニズムを明らかにする
新たな展望を与えると考えられる。例えばPCSK5はproprotein convertase subtilisin/kexin (PCSK) familyに属し、R/K Xn R/K配列をもつ様々な前駆タンパク質を切断し活性化するとされる35)。PCSK5 は分泌型のPCSK5Aと膜型のPCSK5Bの2種類のisoform を持つ 35)。MSCの培養上清においては PCSK5AがWestern blotより検出された (Figure 6)。今回MSC培養上清中に同定されたタンパク質 は PCSK 切断配列と推定される配列を持つものが多く含まれており (data not shown)、PCSK5 が
22
MSCからの分泌タンパク質の放出を制御している可能性が考えられる。スーパーオキシドラジカルか ら過酸化水素への反応を触媒するSOD3はヘパリン結合領域にPCSK認識配列 (RKKR↓R235) をも ち、切断型と非切断型として産生される36)。非切断型のSOD3はヘパリン結合領域を介して細胞外マ トリクスに結合しているため、PCSKによるSOD3の切断は細胞表面からの放出に重要である37)。さ らにTGF-、LTBP、fibrillinと複合体を構成し、TGF-を細胞外マトリクスにとどめるとされる38) fibronectinもまたPCSK認識配列 (RRAR↓R1911) を持つ。従ってSOD3 やTGF-複合体はPCSK5 によって MSC の細胞外マトリクスから培養上清中への放出が促進されるのかもしれない。SOD3 が もつ抗酸化機能やTGF-がもつ再生促進機能はMSCの主要機能であるため、PCSK5によるこれらタ ンパク質の切断が、培養上清中の心筋保護作用に重要な役割を果たす可能性がある。MSCから産生さ
れるPCSK5はMSCそのものだけでなく、周囲の細胞から産生される様々なタンパク質の局在や機能
を制御し、様々な生物学的作用を引き起こすのかもしれない。このような制御機構によって MSC が 広範な治療効果を示すと推測できるが、そのためには更なる研究が必要である。
今回同定した分泌因子群はMSC治療の新しい疾患への適応拡大の可能性を示唆している。BMP1、
SERPINF1、SERPINH1、NPC2を含む遺伝性疾患に関連するタンパク質がMSCの分泌因子として
同定された。BMP1、SERPINF1、SERPINH1の変異は骨の脆弱性、脊柱や長管骨の変形、発育不全 を特徴とする骨形成不全症を引き起こすことが知られている39)。また他家MSC を骨形成不全症患者 に投与することで骨芽細胞の分化を促進し、その症状が改善することが報告されている40), 41)。骨芽細 胞の供給源であることに加えて、BMP1、SERPINF1、SRPINH1 を分泌する MSC は、骨形成不全 症患者におけるこれらタンパク質の補充にも寄与しうるかもしれない。NPC1、NPC2 変異が原因の
Niemann-Pick type C病は細胞内にコレステロールが異常凝集する神経変性疾患である。MSC投与が
NPC1変異をもつNiemann-Pick type C病モデルマウスの神経機能を改善したとの報告がある42)。ま た recombinant NPC2がNPC2変異ヒト線維芽細胞においてコレステロールの凝集を回復させたと の報告もされている43)。それ故にNPC2を含むMSCの培養上清はNPC2変異のNiemann-Pick type C病の治療に役立つ可能性が考えられる。同様にMSCやMSC培養上清は欠乏タンパク質を補充する ことにより、他の遺伝性疾患にも同様の効果を発揮するかもしれない。このように MSC の分泌因子 の情報を利用して、MSCで治療可能な疾患を新たに明らかにすることは興味深い治療アプローチであ る。
著者は今回DPSCを含む3つのMSCの共通分泌因子を同定した。これらの因子にはすでにMSC の治療因子として報告のあるサイトカインや成長因子が含まれており、これらの全体解析は、MSCの 治療効果に関連する複数の機能と関連することを示した。このタンパク質リストは新しい MSC 治療 の代替因子の同定、MSC治療メカニズムの解明、MSC治療の新規適応疾患の予測に有用であると考
23
えられる。分泌因子のより詳細な解析が MSC 治療の更なる可能性を明らかにし、臨床応用の発展に つながることが期待される。