熊本大学学位論文
膜−水相界面におけるα
1-酸性糖タンパク質の 構造及び機能特性の評価
2004 西 弘 二
Evaluation of structural and functional characteristics of α
1-acid glycoprotein on membrane–water phase
Koji Nishi
Evaluation of structural and functional characteristics of
1-acid glycoprotein on membrane–water phase
Koji Nishi
Human α1-Acid glycoprotein (AGP), a member of the lipocalin family, is a polypeptide with 2 disulfide bonds and 5 carbohydrate chains, which account for about 45% of its total mass of 40 kD. It is a major binding protein for neutral and basic ligands. Although its three-dimensional structure and biological functions are still unknown, circular dichroism (CD) measurements and molecular modeling lead to the conclusion that this protein has a largely β-sheet structure in aqueous solution.
The hypothesis that membrane transport of a drug depends on the concentration of
the non-bound drug is widely accepted. However, because this hypothesis does not fully explain the uptake mechanism of some AGP-binding drugs, a protein-mediated uptake system has been proposed. In such a system, structural changes in the protein due to the interactions with the membrane surface decrease its drug-binding capacity.
Here, we report structural and functional properties of AGP in the interaction with the membrane and various factors involved in it.
1) In reverse micelles used as the membrane model, new peaks for AGP at 209nm and 222nm appeared in the CD spectrum. This result suggests that, as the result of the interaction with the membrane, the structural conformation of AGP changes to an
α- helix structure from the original
β-sheet structure in native state. From experiments of its drug-binding properties, the binding capacity of AGP to three representative drugs in reverse micelles becomes significantly decreased. These results support the validity of the AGP mediated ligand-uptake model.
2) Based on experiments using liposomes under mildly acidic conditions, it appears that
AGP is converted into an
α-helix structure rich form, following its binding to themembrane by the electrostatic interaction, and the unique structural conformation of
AGP under mildly acidic conditions becomes a trigger for its interaction. The binding
capacity of AGP to progesterone was significantly decreased as the result of the
interaction with the membrane. These results also strongly support the view that some
AGP-binding drugs are taken up into the tissue in the AGP mediated ligand-uptake model. In addition, an expression system of recombinant AGP (rAGP), which had not been previously established, was constructed using yeast (Pichia pastoris), and the findings indicate that rAGP had similar structural and functional properties to AGP. On the examination using tryptophan (Trp, W) mutants (W25A, W122A and W160A) produced by this expression system, Trp25 and Trp160 appear to be present in the hydrophobic region in the interior of the membrane and Trp122 was on the outside.
3) In mildly acidic condition (pH4.0), methanol and other alcohols significantly induced an α-helix structure formation in AGP. It was also observed that this
α-helix structurewas formed under strong acidic (pH2.0) and highly ionic (1.5M NaCl) conditions.
These results indicate that hydrophobic interaction involve in the formation of an
α- helix structure for AGP, and an
α-helix structure of AGP observed in its interaction with the membrane is formed inside the membrane.
4) Since the structural properties of reduced asialo-AGP (RA-AGP) are similar to molten-globule structure observed under strongly acidic (pH2.0) and highly ionic (1.5M NaCl) conditions and in the presence of alcohol, RA-AGP might serve as a useful model of an
α-helix structure form, which was observed in its interaction with themembrane. The results obtained herein using RA-AGP indicate that, in the formation of a molten-globule structure of AGP, two steps are involved, that is, the first is a fast step (seconds to minutes) and the second a slow one (〜hrs). Moreover, using histidine mutants and chemically modified AGP, the findings indicate indicated that His97 and His100 are important residues in the formation of an α-helix structure of AGP.
These results provide useful basic information for the appropriate use of an AGP
binding-drug, in which the protein-binding rate is its index. Moreover, it would also be
interesting to evaluate the relationship between the biological function of the molecule
and structural characteristics.
本論文で使用した略語一覧
AGP
α1-酸性糖タンパク質
α1-Acid glycoprotein
A-AGP
アシアロAGP Asialo-AGP
Ala; A
アラニンAlanine
AOX1
アルコールオキシダーゼ1 Alcohol oxidase 1 Asn
アスパラギンAsparagine
1,4-Bu(OH)
21,4-ブタンジオール 1,4-Butandiol
BSA
ウシ血清アルブミンBovine serum albumin
CD
円二色性Circular dichroism
CYP
チトクロームP450 Cytochrome P450 DMI
デシプラミンDesipramine Es
エストラジオールEstradiol
ESR
電子スピン共鳴Electron spin resonance
EtOH
エタノールEthanol
Et(OH)
2 エタンジオールEthandiol
GPF
グリコペプチダーゼF Glycopeptidase F
hAGP
ヒトAGP Human AGP
His; H
ヒスチジンHistidine
HSA
ヒト血清アルブミンHuman serum albumin IMP
イミプラミンImipramine
MeOH
メタノールMethanol
NMR
核磁気共鳴Nuclear magnetic resonance
PC
フォスファチジルコリンPhosphatidyl-choline PE
フォスファチジルエタノールアミンPhosphatidyl-ethanol-
amine
PG
フォスファチジルグリセロールPhosphatidyl-glycerol PL
プロプラノロールPropranolol
PrOH
プロパノールPropanol
1,2-Pr(OH)
21,2-プロパンジオール 1,2-Propanediol
PS
フォスファチジルセリンPhosphatidyl-serine
RA-AGP
還元型アシアロAGP Reduced asialo-AGP
rAGP
組換え型AGP Recombinant AGP
R-AGP
還元型AGP Reduced-AGP
TFE 2,2,2-トリフルオロエタノール Trifluoroethanol
Trp; W
トリプトファンTryptophan
WT
野生型Wild type
本論文は学術雑誌に掲載された次の論文を基礎とするものである。
(1)
Structural and Drug-binding Properties of
α1-Acid Glycoprotein in Reverse Micelles
Biochim. Biophys. Acta,1601, 185-191 (2002).
Nishi K, Sakai N, Komine Y, Maruyama T, Halsall H.B, Otagiri M.
(2)
Binding of α
1-Acid Glycoprotein to Membrane Results in Unique Structural Change and Ligand Release
Biochemistry, submitted for publication.
Nishi K, Maruyama T, Handa T, Halsall H.B, Otagiri M.
(3) Role of Histidine Residues on the Formation of α-Helix Structure of α1
-Acid Glycoprotein
In preparation.
Nishi K, Fukunaga N, Komine Y, Maruyama T, Halsall H.B, Otagiri M.
目次
第
1
章 緒論---1第
2
章 α1-酸性糖タンパク質(AGP)と逆相ミセルの相互作用に
関する検討---5第
1
節 序---5
第
2
節AGP
と逆相ミセルの相互作用---5
2-1
逆相ミセル中のAGP
の立体構造の評価---52-2
逆相ミセル中のAGP
の薬物結合性の評価---6
第
3
節 小括---8
第
3
章 AGPとリポソームの相互作用に関する検討---10
第
1
節 序---10
第
2
節AGP
の結合性評価---10
2-1
電荷及びpH
の影響---10
2-2
シアル酸の影響---14
第
3
節AGP
の構造特性評価---15
3-1
電荷の影響---15
3-2 pH
の影響---17
3-3
シアル酸の影響---18
第
4
節AGP
の機能特性評価---19
第
5
節AGP-リポソーム複合体の構造特性評価 ---20
5-1
トリプトファン(Trp; W)残基の蛍光スペクトル---20
5-2
組換え型AGP
の発現系の構築---21
5-3
組換え型AGP
の構造・機能特性の評価---25
5-4 Trp
変異体を用いた複合体の構造予測---27
第
6
節 小括---28
第
4
章 AGPのα-ヘリックス構造形成機構に関する検討 ---31
第
1
節 序---31
第
2
節 アルコールによるAGP
のα-ヘリックス構造の誘導---31
2-1
メタノールによるα-ヘリックス誘導効果---312-2 AGP
の α-ヘリックス構造形成におけるアルキル鎖の影響---32
第
3
節 酸性・高イオン強度下におけるAGP
のα-ヘリックス構造 の形成---34
3-1 AGP
の酸性モルテン・グロビュール状態---343-2 Stern-Volmer
プロット解析によるトTrp
残基の環境評価---35
第
4
節 小括---36
第
5
章 AGPのα-ヘリックス構造形成における各種因子の検討 ---38
第
1
節 序---38
第
2
節 ジスルフィド結合及びシアル酸の影響---38
2-1
円二色性(CD)スペクトルによる各種修飾AGP
の構造 特性評価---38
2-2
キナルジンレッドの結合性評価---39
2-3 CD
スペクトルの時間依存的変化---40
2-4
α-ヘリックス構造形成に及ぼすヒスチジン(His; H)残基 の影響---41
第
3
節 小括---43
第
6
章 総括---45実験の部
---48
謝辞
---54
参考文献
---55
第
1
章 緒論α1
-酸性糖タンパク質(AGP)は、183
個のアミノ酸残基と五本の糖鎖からなる分子量約
40kD
の血清糖タンパク質である 1)。このタンパク質の糖鎖含量は約
45%と他の糖タンパク質のなかでも極めて高いため、 X
線結晶構造解析やNMR
解析等による立体構造の解明はなされていない。しかしながら、円二色 性(CD)スペクトルや分子モデルの設計によるデータから、水溶液中では β−シート構造に富んでいることが報告されている2,3)。
N
C
164
183
54 38 15
5
85 75
72 147
Figure 1. Structural model of AGP
:Glycan、■:-S-S- bond
このタンパク質の生物学的意義は現在までのところ不明であるが、疎水性化 合物の輸送担体として機能しているタンパク質、リポカリンファミリーの一員 として分類されている 4,5)。事実、血中では多くの塩基性薬物やステロイドホル モン類の主要結合担体として働き、結合薬物の体内挙動に影響を及ぼしている
1,6)。また、AGP は生体反応の急性期(ストレス、外傷、炎症、腫瘍等)に血中 及び組織中で著しく増加する急性相反応物質の一つでもあり 7-9)、Cytochrome
P450(CYP)をはじめとする一連の薬物代謝酵素を誘導するフェノバルビター
ル、リファンピシンなどの薬物の反復投与時にも上昇することが知られている10,11)。このような
AGP
の濃度変動は、AGPと結合性の高い薬物のタンパク結合に変化をもたらし、その体内動態、ひいては薬理効果に影響を与える可能性が 十分に考えられる。事実、従来の
in vivo
及びin vitro
の検討に加え、AGPトランスジェニックマウスを用いた検討で、血中の
AGP
濃度の上昇が、イミプラ ミンの非結合型濃度を減少させるため、その消失や分布、さらには薬理効果に まで影響を及ぼすことが確認されている12)。また新規抗癌剤UCN-01
((3R*, 8S*,9R*, 10R*, 12R*)-2, 3, 9, 10, 11, 12-Hexahydro-3-hydroxy-9-methoxy-8-methyl-10- (methylamino)-8, 12-epoxy-1H, 8H-2, 7b, 12a-triazadibenzo[a, g]–cyclonona[cde]
trinden-1-one; 7-Hydroxy-staurosporine)のヒトにおける体内動態(総クリアラン
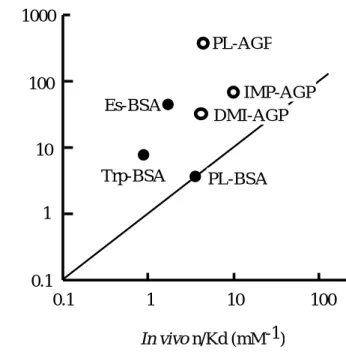
ス、血中消失半減期等)は、AGP との強固な特異的結合によって都合良く説明 されている13)。In vitro n/Kd (mM-1 )
0.1
1 10 100
1 10 100 1000
0.1
PL-AGP
PL-BSA Es-BSA
Trp-BSA
In vivo n/Kd (mM-1)
Figure 2. Relationship between in vivo and in vitro ligand binding (n/Kd) to plasma protein
14)The solid line represents a 1:1 relationship. Abbreviations are as follows: BSA, bovine serum albumin; PL, propranolol; IMP, imipramine; DMI, desipramine;
Es, estradiol; Trp, tryptophan.
IMP-AGP DMI-AGP
その一方で、これら
AGP
結合性薬物のうち、プロプラノロール、イミプラ ミン、デシプラミン等においては、in vivo でのAGP
への結合性が、in vitroで の結合性よりも小さくなることが知られており(Figure 2)14)、類似した減少が、同じ輸送タンパク質であるヒト血清アルブミン(
HSA)においても観察されて
いる 15-17)。このような背景には、タンパク質介在性のリガンドの組織移行機構
の存在の可能性が考えられる。これは、細胞表面に結合した一部の血中タンパ
ク質が生体膜との相互作用により構造変化を惹起し、次いで、結合型リガンド を解離・放出するというものである(
Figure 3)。実際、ヒト白血球(リンパ球、
顆粒球、単球)及び赤血球表面上での AGP の存在が蛍光電子顕微鏡により確 認されている 18,19)。最近、AGP は血管内皮細胞の表面に結合し、エンドサイト ーシス→エキソサイトーシスという過程(トランスサイトーシス)により血中 から消失するという報告もあり、この過程における
AGP
結合型リガンドの解 離の可能性も十分に考えられる 20)。また電子スピン共鳴(ESR)スペクトルに より、HSA も肝細胞表面で構造変化を起こし、それに伴い薬物結合能が低下す る可能性があることが報告されている21)。Membrane Blood Tissue
L
Protein L
L Ligand
Protein
L L
Figure 3. Proposed model for protein-mediated ligand uptake into tissue
最近、Bychkova らは、生体内でタンパク質が生体膜と相互作用する際、タン パク質の構造に一種の変性状態である 緩み が起こることから、この状態が 天然状態と変性状態の中間にあるのではないかという仮説を提唱している22,23)。 実際に、生理的条件下ではタンパク質の変性構造として、ランダムコイルのよ うな高度な変性状態は考えにくく、むしろ中間状態として様々な機能を果たし ていると考えるほうが、極めて合理的でありかつ魅力的である。このことは、
AGP
が細胞表面において膜と相互作用する際、中間状態を形成することにより、結合型リガンドを組織へ放出するのみでなく、生理活性を誘起させる可能性を も期待させる。
これまで、生体膜を構成している脂質二重膜や糖脂質等は細胞内小器官を内 封するためだけに存在する器官として、その機能は極めて消極的なものと考え られてきた。しかし最近の報告から、生体膜は非常に多彩な機能を持つことが 明らかにされつつある。そのなかの一つに生体内に存在するタンパク質との相 互作用がある。この場合、その因子として生体膜自身が持つ様々な物理化学的 性質が挙げられる。一般に生体膜表面は負の電荷を帯びることから、その静電 的ポテンシャルにより周囲のバルク相からプロトンが引き寄せられ、局所的に
pH
が低下していると考えられている 24)。このような環境は、周囲に存在する タンパク質の構造や機能に影響を及ぼすと考えられる。事実、そのような因子 が原因となり、生体膜との相互作用が観察されているタンパク質が多数報告されている 25-30)。そのため、AGP と生体膜の間にもこのような因子が関与し、相
互作用を促す可能性が十分に考えられる。
このような背景のもと、AGP と生体膜の相互作用様式とその際の
AGP
の構 造変化のメカニズムの解明を目的として本研究を行った。まず、第2
及び3
章 において生体膜のモデルである逆相ミセル及びリポソームをそれぞれ用いて、AGP
と生体膜の相互作用に伴う構造特性及び薬物結合能の変化について検討し た。さらに第4
章では、未だ確立されていないAGP
の発現系の構築を行うこ とにより、変異型AGP
を作製して、生体膜とAGP
の相互作用様式の詳細につ いて検討した。第5
章では、アルコール存在下や強酸、高イオン強度下での構 造特性の観点から、第6
章ではAGP
分子内に存在する種々の構造形成因子(S-S
結合、シアル酸、アミノ酸)の観点から、生体膜との相互作用によるAGP
の 構造転移のメカニズムについて検討した。以下に得られた知見を詳述する。第
2
章 1-酸性糖タンパク質( AGP)と逆相ミセルの相互作用に関する
検討第
1
節 序疎水性領域と親水性領域を兼ね備えた界面活性剤のミセルは、生体膜の最も 単純なモデルとして親水効果の研究過程にしばしば用いられる31,32)。なかでも、
逆相ミセル内部は、膜-水相界面のミミックとして数多く用いられてきた。この 逆相ミセルは、界面活性剤、有機溶媒、水の三種混合系であり、このうち
AGP
は膜の内部の水相部分に取り込まれている。生体膜表面には、負の電荷の存在 が確認されていることから、本研究では生体膜に近いミミックとしてアニオン 性のスルホコハク酸ジ-2-エチルヘキシルナトリウムを使用した。第
2
節 AGPと逆相ミセルの相互作用2-1 逆相ミセル中の AGP
の立体構造の評価これまで、CD スペクトルや分子モデルの設計から、
AGP
はβ-シートに富ん だ構造をとっていることが知られている 2,3)。そこで、水溶液中及び逆相ミセル 中におけるAGP
のCD
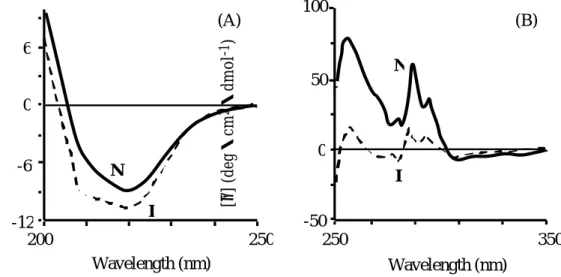
スペクトルを測定した(Figure 4)。水溶液中のAGP
は、遠紫外領域においては、219nm付近のピークと
209nm
付近にショルダーを持つ コットン効果を、近紫外領域においては秩序だった三次構造に起因する芳香族 アミノ酸残基由来のCD
バンドを示した。一方、生体膜モデルの逆相ミセル中 においては、近紫外領域で観察されたコットン効果は著しく減少し、三次構造 の崩壊、特に芳香族アミノ酸残基の固定の程度が著しく低下している可能性が 示唆された。通常、未変性タンパク質は、タンパク質コアでの側鎖のタイトな パッキング、遠距離疎水結合や塩架橋及びそれらのネットワーク等の分子内相 互作用により保持されているため、タンパク質独自の堅固な三次構造を持つ。したがって、逆相ミセル中で観察された三次構造の欠如は、これら分子内相互 作用が崩壊し側鎖の運動の自由度が高くなったことを示している。また興味深 いことに、遠紫外領域では、209nmと
222nm
付近にピークを有するα-ヘリックス構造に特有なスペクトルが増大した。これらの結果から、
AGP
は膜-水相界 面においてα-ヘリックス構造が新たに形成されるものの、三次構造は消失して いるという特殊な状態にあることが明らかとなった。-50
250 350
(B)
Wavelength (nm) -6
0 6
200 250
-12
Wavelength (nm)
0 50
[θ] (deg・cm2・dmol-1)
[θ]×10-3 (deg・cm2・dmol-1) 100
(A)
N I
N
I
Figure 4. Far (A) and near (B) UV CD spectra of AGP in native state (N; solid line) and membrane interaction mode (I; dashed line)
2-2 逆相ミセル中の AGP
の薬物結合性の評価これまでに、
AGP
分子上の薬物結合サイトに関する研究から、AGP
分子上 には酸性、塩基性薬物及びステロイドホルモンの少なくとも三つの結合サブサ イトが存在し、それらが互いに重なり合って、一つの幅広い結合領域を形成し ていると考えられている 33,34)。そこで、これらのサブサイトのマーカーとして 既にAGP
と強く結合することが知られている薬物、すなわち酸性薬物として ジクマロール、塩基性薬物としてクロルプロマジン、ステロイドホルモンとし てプロゲステロンを使用し、逆相ミセル中におけるこれらリガンドとAGP
と の結合性を調べた。なお、Figure 5にこれら薬物の構造式を示す。AGP は、その分子内に三つのトリプトファン(
Trp)残基を有しており、こ
のTrp
残基由来の蛍光スペクトルは、ジクマロールやクロルプロマジン及びプ ロゲステロンの濃度増大とともに減少した(Figure 6)。ジクマロール及びクロ ルプロマジンにおいてはほぼ70%、プロゲステロンでは 20%で消光に飽和が観
察された。したがって、AGP 分子内に存在する三つのTrp
残基のうち、クロルプロマジンやジクマロールにおいては二つの
Trp
残基が、プロゲステロンの場 合は一つのTrp
残基だけが結合部位近傍に位置し、このような蛍光消光に関与 している可能性が考えられる。このAGP
の蛍光消光を薬物結合に基づくもの と仮定し、各薬物の結合パラメータを算出した(Table 1)。CH3
CH3 C CH3
O
H H
H O
O
OH
O
OH OO
N S
Cl (CH2)3N(CH3)2
Chlorpromazine Dicumarol Progesterone
Figure 5. Chemical structures of chlorpromazine, dicumarol and progesterone
0 1
0 1 2 3 4 5 6
Relative Trp fluorescence intensity
Figure 6. Fluorescence quenching titration curves of various drug-binding to AGP
Each symbol represents chlorpromazine (■), dicumarol (▲) and progesterone (●) respectively.
[Drug] / [AGP]
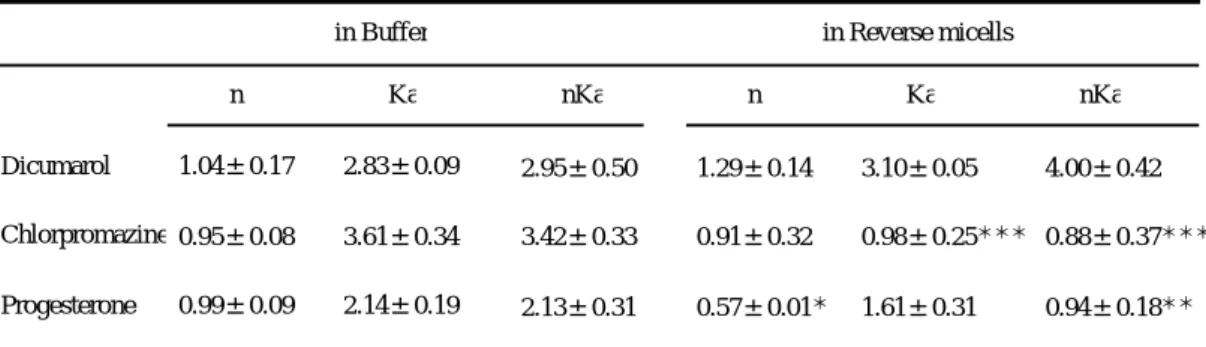
逆相ミセル中での結合性を水溶液中のそれと比較すると、塩基性薬物のクロ ルプロマジンは、結合サイト数の変化なしに結合定数の著しい低下が認められ た。対照的に、酸性薬物のジクマロールの結合性はほとんど変化しなかった。
また、プロゲステロンにおいては、わずかな結合定数の低下に加え、結合サイ ト数の有意な減少が認められた。事実、結合能の指標である
nKa
値はクロルプ ロマジン及びプロゲステロンにおいて逆相ミセル中で有意に低下していた。こ れらの結果は、AGP のα-ヘリックス構造への転移過程において、それぞれの結
合サブサイトが異なった挙動をとることを示唆しているものと思われた。in Buffer in Reverse micells
n Ka nKa
Dicumarol Chlorpromazine Progesterone
1.04±0.17 0.95±0.08 0.99±0.09
2.83±0.09 3.61±0.34 2.14±0.19
2.95±0.50 3.42±0.33 2.13±0.31
1.29±0.14 0.91±0.32 0.57±0.01*
3.10±0.05 0.98±0.25***
1.61±0.31
n Ka nKa
4.00±0.42 0.88±0.37***
0.94±0.18**
n; the number of binding sites, Ka; the association constant (x106M-1).
All values are the means ± S.D. (3-5 experiments)
*Significantly different from value in buffer, p<0.05
**Significantly different from value in buffer, p<0.01
***Significantly different from value in buffer, p<0.001
Table 1. Apparent binding parameters of drugs to AGP by fluorescence quenching method
第
3
節 小括本章では、生体膜のモデルとしてアニオン性の逆相ミセルを作製し、AGP と 逆相ミセルの相互作用について検討した。逆相ミセル中の
AGP
の構造特性をCD
スペクトルを用いて評価した。次に、逆相ミセル中のAGP
の機能特性につい て、酸性薬物、塩基性薬物及びステロイドホルモンを用いてAGP
の薬物結合 能を評価した。以下に、本章で得られた知見を要約する。1)逆相ミセル存在下、AGP
のCD
スペクトルにおいて、209nmと222nm
に新たなピークが観察された。これは
AGP
が生体膜と相互作用する際、本来のβ-シ ート構造からα-ヘリックス構造へ転移することを示唆しているものと考えられ た。2)逆相ミセルと相互作用した際、三種の薬物のうち塩基性薬物のクロルプロ
マジン及びステロイドホルモンのプロゲステロンに対してのみ、AGP の有意な結合能の減少が観察された。そのため、生体膜との相互作用による
AGP
の薬 物結合部位のサブサイトはそれぞれ異なった挙動を示すことが示唆された。これらの知見は、これまでに報告された
AGP
に対するin vivo
及びin vitro
で の薬物結合性の違いが生じる原因の一つとして、提唱されているAGP
介在性 のリガンド組織取り込み機構が存在する可能性を強く示唆するものであった。また、生体膜との相互作用により観察されたα-ヘリックス構造体には、これま で解明されていない
AGP
の生理機能発現との関わりが期待される。第
3
章 AGPとリポソームの相互作用に関する検討第
1
節 序人工脂質二重膜、リポソームは生体膜の最も単純なモデルとして、膜の物性 のみならず、タンパク質、ペプチド及び低分子化合物等との相互作用に関する 研究に汎用されている 35,36)。同じ生体膜モデルである逆相ミセルは、界面活性 剤や有機溶媒などタンパク質の性質に影響を及ぼしやすい物質から形成される が、一方でリポソームは実際に脂質二重膜を形成しているリン脂質から調製さ れるため、逆相ミセルよりもより生体膜に近いモデルであると言える。そこで、
中性リン脂質であるホスファチジルコリン(PC)、酸性リン脂質であるホスフ ァチジルグリセロール(PG)、ホスファチジルセリン(PS)及びホスファチジ ルエタノールアミン(PE)を用いて中性及びアニオン性のリポソームを調製し、
AGP
と生体膜の相互作用について各種検討を行った。さらに、Pichia 酵母を用 いてAGP
の各種変異体を作製し、AGP と生体膜の複合体構造の詳細について 検討を加えた。第
2
節 AGPの結合性評価2-1 電荷及び pH
の影響生体膜表面は、そのポテンシャルにより負の電荷を帯びており、さらにこれ がバルク相のプロトンを引き寄せることにより、生体膜近傍の環境は緩和な酸 性条件下にあることが報告されている 24)。このような条件が、AGPと生体膜の 相互作用に関与している可能性は高い。そこで、酸性領域の各
pH
において、AGP
とPG
あるいはPC
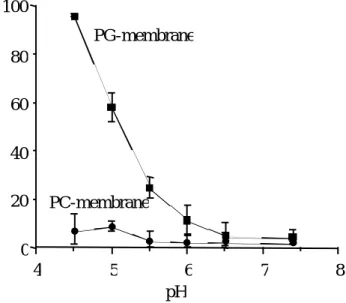
から調製したリポソーム(PG-、PC-membrane)との結 合実験を行い、結合率及び見かけの結合定数を算出した(Figure 7、Table 2)。その結果、pHの低下に伴う
PG-membrane
に対するAGP
の結合率の上昇が観察 された。一方、PC-membrane に関してはこのようなAGP
との有意な結合は観 察されなかった。これより、AGP と生体膜の結合には静電的相互作用が関与し ていることが示唆された。また、PG-membrane との結合がpH6.0
からpH4.5
の間で急激に上昇した理由として、このような
pH
領域でのAGP
分子のプロトン 化による正電荷への荷電が考えられる。0 100
4 5 6 7 8
PG-membrane
PC-membrane
% Bound AGP
pH
Figure 7. Effect of pH on AGP-membrane (PG (■)- and PC (●)-membrane) interaction 80
60 40 20
T a b l e 2 . A p p a r e n t a s s o c i a t i o n c o n s t a n t ( K a ) o f A G P w i t h P G - a n d P C - m e m b r a n e a t v a r i o u s p H s
Lipid
PG
PC
pH Ka (M
-1)
7.4 6.5 6.0 5.5 5.0 4.5 7.4 6.5 6.0 5.5 5.0 4.5
100 ± 40 140 ± 60 330 ± 30 830 ± 60 3,500 ± 280 63,000 ± 9500
60 ± 20
90 ± 10
80 ± 10
90 ± 10
240 ± 10
200 ± 230
一般にタンパク質と生体膜の結合は、タンパク質の正の電荷と生体膜の負の 電荷との静電的相互作用による場合が多い。事実、これまでに、生体膜との静 電的相互作用により相互作用を引き起こすタンパク質の存在が数多く報告され
ている 25-30)。このような静電的相互作用の関与を確認する方法としては、イオ
ン強度を変化させ、その影響を観察するのが最も一般的である。すなわち、
NaCl
の濃度を上げていき、イオン強度を上げることによって、膜とAGP
の結合が 抑制された場合は、両者の間に静電的相互作用が関与している可能性が高くな る。 そこで、pH7.4とpH4.5
においてそれぞれNaCl
を添加して、膜とAGP
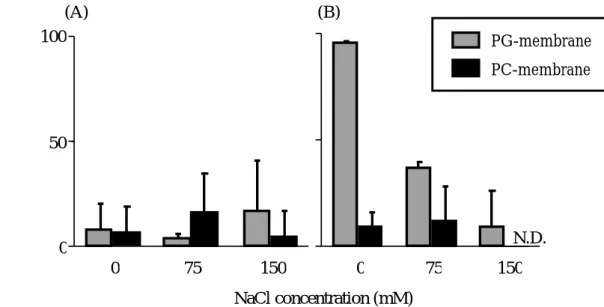
の 結合性の変化について検討した(Figure 8)。その結果、Figure 8(A)に示すよう に、pH7.4 ではPG-及び PC-membrane
ともに塩濃度に依存せず、有意な結合は 観察されなかった。一方、Figure 8(B)とTable 3
に示すように、pH4.5ではPG-
membrane
においてのみ塩濃度依存的に結合性の低下が観察された。Figure 7で得られた結果と合わせて考えると、AGP は膜近傍の環境と類似した緩和な酸性 条件下で膜との結合性を示し、そこには静電的相互作用が関与していることが 強く示唆された。
0 50 100
0 75 150 0 75 150
NaCl concentration (mM)
% Bound AGP
N.D.
(A) (B)
PG-membrane PC-membrane
Figure 8. Effect of ionic strength on the binding of AGP to membrane at pH7.4 (A) and pH4.5 (B)
N.D. : not determined
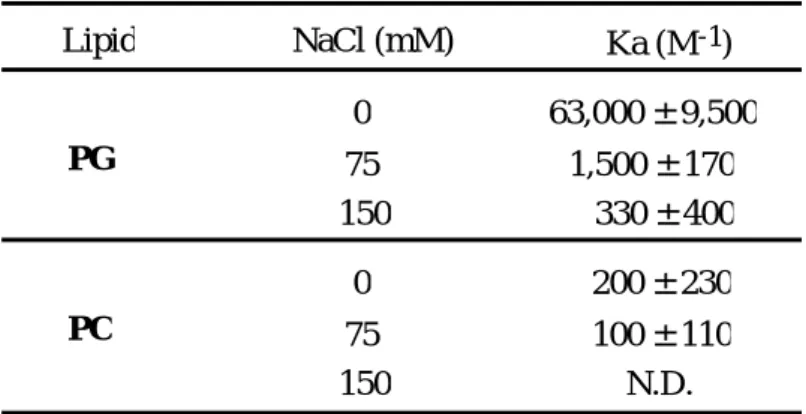
Table 3. Effect of ionic strength on the apparent association constant (Ka) of AGP to membrane at pH4.5
Lipid PG
PC
NaCl (mM) Ka (M
-1) 0
75
150 330 ± 400
1,500 ± 170 63,000 ± 9,500
0 75 150
200 ± 230 100 ± 110
N.D.
N.D. : not determined
さらに、PGと同じ酸性リン脂質である
PS、PE
を用いて調製したリポソーム とAGP
の結合実験を行った(Figure 9)0 100
PG PS PE PC
% Bound AGP
80 60 40 20
Figure 9. Binding of AGP to PG-, PS-, PE-, and PC-membrane at pH4.5
PS は
PG
と同等のアニオン性を有しており、PE もこれらに比べると極めて 低いが、わずかにアニオン性を有している脂質である。Figure 9
から明らかな ように、PS においてはPG
に匹敵する程のAGP
結合性を示した.また、PEに おいても弱い結合性が見られた.これらリポソームによるAGP
の結合性の違 いが、リポソームのアニオン性の程度に対応していることからも、AGP と生体 膜の結合に静電的相互作用が関与していることが強く示唆された。2-2 シアル酸の影響
AGP はその分子内の五つのアスパラギン(Asn)残基に枝分かれ構造を有す る糖鎖が結合しており,血清タンパク質のなかでも最も糖化されているタンパ ク質の一つである。いずれの糖鎖末端にも、負に帯電したシアル酸が数多く存 在しているため、最も小さな
pI
値を持つ血清タンパク質でもある 1)。一般にタ ンパク質の糖鎖は、それ自身の水溶性を高めるのみならず、タンパク質-タンパ ク質間相互作用に影響を及ぼしたり、多様な生理システムにおける分子認識や タンパク質のコンフォメーションの安定性に寄与していることが明らかにされている 37-40)。AGP の場合も、生理機能発現や自身の安定性の維持に糖鎖が関与
していると考えられているため、これらシアル酸が AGP と生体膜の相互作用 にどのように関与しているのかを明らかにすることは、非常に興味深いことと 思われる。そこで、AGP の糖鎖先端に存在するシアル酸を酵素的にすべて除去 したアシアロ AGP を調製し、PG-membrane との結合性について検討した(Figure 10)。
Figure 10. Effect of sialic acid on the interaction between AGP and PG-membrane at pH4.5
*Significantly different from value of native AGP, p<0.05
0 100
Native Asialo
% Bound AGP
80 * 60 40 20
その結果、AGP と比較してアシアロ AGP と膜との結合性は有意に低下した。
これは、AGP の糖鎖、特にシアル酸が生体膜との結合を促進する作用を有する 可能性を示唆する。本来、シアル酸は負に帯電しているため、同じく負に帯電 している生体膜とは互いに反発することが予想される。しかし得られた結果は
予想に反し、シアル酸の存在が AGP と生体膜との相互作用に正に作用すると いうものであった。その原因としては、pH4.5 といった緩和な酸性条件下にお ける AGP 分子のプロトン化による正電荷の増加や、それに伴う構造変化を、
シアル酸が促進したためと考えられた。
第 3 節 AGP の構造特性評価
3-1 電荷の影響
これまでの結果から、AGP は生体膜と静電的相互作用により結合している可 能性が示唆された。その場合、AGP がどのような構造状態をとって生体膜と結 合しているのかを明らかにすることは重要である。そこで、pH4.5において
PG-
及び
PC-membrane
の濃度を変化させた場合のAGP
の構造特性の変化を、CDスペクトル法を用いて、α-ヘリックス含量の指標となる
222nm
における平均モル 楕円率([θ])を測定することにより検討した(Figure 11)。Figure 11. Effects of PG- and PC-membrane on the conformational structur e of AGP at pH4.5
(A) Far UV CD spectra of AGP in the absence (solid line) or presence (dashed line) of PG- membrane at pH4.5 and (B) -[θ] at 222nm of AGP interacting with PG (■) - a n d P C (●) -
membrane.
- [θ ] x 10-3
at 222nm (deg·cm
2·dmol-1)
4 14
0 60
PG-membrane
PC-membrane
[Phospholipid] / [AGP]
12 10 8 6
20 40
-20 -10 0 10
200 250
Wavelength (nm)
[θ ] x 10-3
(deg·cm
2·dmol-1)
(A) (B)
pH4.5 において、PG-membrane では、その濃度に依存してα
-ヘリックス含量
の増加が観察され、その構造変化はβ-シート構造からα-ヘリックス構造への二 状態転移で近似できることが示唆された。一方PC-membrane
においては、この ような有意なα-ヘリックス含量の変化は観察されなかった。またデータには示
していないが、pH7.4においてはPG-及び PC-membrane
存在下、AGPの構造に 有意な変化は観察されなかった。これらの結果から、AGP と生体膜の相互作用 には、膜近傍の緩和な酸性環境と静電的相互作用が協同的に関与している可能 性が示唆された。ここで観察された静電的相互作用の関与を確認するために、
AGP
とPG-
membrane
との相互作用に対するNaCl
の影響について検討した(Figure 12)。その結果、
NaCl
の濃度依存的にα-ヘリックス含量の減少が観察されたことより、
静電的相互作用の関与が示唆された。また、NaClの濃度が
150mM
のときAGP
の遠紫外領域のCD
スペクトルはPG
非存在下と極めて類似しており、両者の 相互作用はほとんど抑えられていることが示唆された。6 14
0 50 100 150
NaCl concentration (mM)
Figure 12. Effect of ionic strength on the conformational structure of AGP
interacting with PG-membrane at pH4.5(A) Far UV CD spectra of AGP interacting with PG-membrane in the absence (solid line) or presence ( d a s h e d l i n e ) o f N a C l ( 1 5 0 m M ) a n d ( B ) - [θ] a t 2 2 2 n m o f A G P i n t e r a c t i n g w i t h P G - m e m b r a n e i n various concentrations of NaCl.
10 12
8
- [θ ] x 10-3
at 222nm (deg·cm
2·dmol-1)
-20 -10 0 10
200 250
Wavelength (nm)
[θ ] x 10-3
(deg·cm
2·dmol-1)
(A) (B)
3-2 pH
の影響生体膜と
AGP
の相互作用には静電的相互作用が大きく関与している一方で、このような相互作用が緩和な
pH
の低下に伴い惹起されることから、pH 低下が 生体膜との相互作用の引き金になり得る可能性が考えられる。そこで、酸性領 域の各pH
におけるAGP
と生体膜の相互作用について、CD スペクトル法を用 いて検討を行った(Figure 13)。その結果、PG-membrane存在下、pHの低下に伴ってα-ヘリックス含量の増加が観察された。一方、データには示していない
が、PG 非存在下では、pH の変化に関わらずα
-ヘリックス含量に有意な変化は
見られなかった。これらの結果から、AGP はpH
の低下に伴って、生体膜と相 互作用を受けやすくなることが示唆された。また、PC-membrane
を用いた同様 の検討では有意なα-ヘリックス含量の変化は観察されなかった。4 14
4.5 6.5
pH
PG-membrane
PC-membrane
Figure 13. Effect of pH on the interactio n between AGP and PG (●) - and PC (■) - membrane
12 10 8 6
5.0 5.5 6.0
- [θ ] x 10-3
at 222nm (deg·cm
2·dmol-1)
膜タンパク質の膜貫通ドメインの多くが、膜内部(疎水性領域)で疎水相互 作用によりα-ヘリックス構造を形成していることから、AGP と生体膜の相互作 用は、膜への結合からβ→α構造転移という過程を経ており、この構造転移の駆 動力として疎水相互作用が働いていると考えられる。また、この構造転移が
NaCl
の添加により抑制されたことからも、そのような過程が存在する可能性が強く示唆された。さらに、AGP と生体膜の相互作用において、両者の結合過程が律 速であり、そこに緩やかな
pH
の低下や静電的相互作用が関与しているものと 考えられる。また、PG-membrane との相互作用によるα
-ヘリックス含量の増加は、このよ
うな緩やかな酸性条件下での構造特性に起因している可能性が考えられる。そ こで、pH4.5 におけるAGP
の構造特性について、CD スペクトル法を用いて検 討を行った(Figure 14)。その結果、AGPはpH4.5
において三次構造は変化し ているものの(Figure 14(B))、二次構造は維持されている(Figure 14(A))と
いうユニークな構造状態をとっていることが示唆された。Figure 13において、AGP
とPC-membrane
との相互作用は観察されなかったことから、このような特殊な構造変化による
AGP
分子表面の正電荷の増加が相互作用の引き金にな っている可能性が示唆された。-10 0 10
200 250
-100 0 100
250 350
Wavelength (nm) Wavelength (nm)
[θ] (deg・cm2・dmol-1) [θ]×10-3 (deg・cm2・dmol-1)
Figure 14. The far-(A) and near-(B) UV CD spectra of AGP at pH7.4 (solid line) and 4.5 (dashed line)
(A) (B)
3-3 シアル酸の影響
リポソームと
AGP
の結合性に関する検討から、AGP の糖鎖先端に存在する シアル酸は、生体膜との結合を促進する因子であることが示唆された。そこで、このようなシアル酸の作用が、生体膜との相互作用における構造特性の変化に どのような影響を及ぼすかについて、PG-membrane との相互作用によるβ→α構
造転移を
CD
スペクトルにより検討した(Figure 15)。その結果、アシアロ
AGP
のα-ヘリックス構造への転移の程度はAGP
と比較 して有意に抑制された。この原因として、次の三つが考えられた。1)アシア
ロAGP
の膜への結合性の有意な低下。2)アシアロAGP
自身の構造特性。3)アシアロ
AGP
の等電点の上昇。これまでの報告によると、アシアロAGP
はシ アル酸が存在しないため分子間反発が減少し凝集しやすくなる一方で、AGP 分 子中に存在する五本の糖鎖間の反発(分子内反発)がなくなりペプチド部分が よりコンパクトになり、安定性が上昇することが知られている 1)。また、シア ル酸が存在しないことでAGP
の等電点が上昇するため、pH4.5 におけるAGP
とアシアロAGP
の構造特性に伴う電荷の状態の違いが関与している可能性も 考えられた1)。4 14
0 60
[PG] / [AGP]
12 10 8 6
20 40
- [θ ] x 10-3
at 222nm (deg·cm
2·dmol-1)
AGP
asialo AGP
Figure 15. Effect of sialic acid on the conformational structure of native (■) and asialo (●) AGP interacting with PG-membrane at pH4.5
第
4
節 AGPの機能特性評価生体膜との相互作用による
AGP
の構造特性の変化が、薬物結合性に及ぼす 影響を知ることは、AGP 介在性のリガンドの組織取り込み機構の存在の可能性 を確認することになる。そこで、pH7.4〜4.5、PG-membrane
存在下、中性リガ ンドであるプロゲステロンのAGP
に対する結合率を求め、各pH
でのα-ヘリックス含量に対してプロットした(
Figure 16)。その結果、生体膜との相互作用
により形成されるα-ヘリックス構造の含量とプロゲステロンの結合率に有意な 相関が観察されたことから、プロゲステロンの結合部位はAGP
のα-ヘリック ス構造形成部位に存在する可能性が示唆された(Figure 16)。また結果には示
していないが、PG-membrane非存在下ではpH7.4〜4.5
の間で有意な結合率の変 化は観察されなかった。Figure 16. Correlation between
binding of progesterone and
α-helixcontents of AGP interacting with PG-membrane at various pHs
60 70 80 90 100
6 7 8 9 10 11 12 13
-[θ] x 10-3 at 222nm (deg·cm
2·dmol
-1)
Progesterone bound to AGP (%)
第
5
節 AGP-リポソーム複合体の構造特性評価5-1 トリプトファン(Trp)残基の蛍光スペクトル
これまでの結果から、AGP は生体膜と結合しα-ヘリックス構造を形成するこ とが示唆されたが、果たして
AGP
のこのようなα-ヘリックス構造は膜の表面 もしくは内部のどちらで形成されているのだろうか。膜タンパク質の一般的な 構造的特徴から、α-ヘリックス構造は膜内部で形成されていると考えるのが妥当と思われる。そこで、PG-membrane 存在または非存在下、蛍光スペクトル法 を用いて
AGP
分子内に存在するTrp
残基の蛍光極大波長を測定することによ り、膜と相互作用した際のTrp
残基の環境評価を行った(Figure 17)。その結果、PG-membrane存在下、AGP分子の
Trp
残基の極大蛍光強度におい て有意な低波長シフト(blue shift)が観察された。このシフトは、NaClの添加 により抑制されたことから、AGP と膜の相互作用により生じたものであること が示唆された。したがって、膜と相互作用した際、Trp
残基は膜内部の疎水性 領域に存在することが推察された。0.8 1 1.2
335 340 345
PG+NaCl none PG
Fluorescence intensity (arbitrary unit)
Wavelength (nm)
Figure 17. Effect of PG-membrane on Trp fluorescence intensity of AGP
Each symbol represents fluorescence intensity and λmax of AGP at pH4.5 (▲), pH4.5 + PG (●) and pH4.5 + PG + NaCl (150mM) (■) respectively.
5-2 組換え型 AGP
の発現系の構築Trp
残基の蛍光スペクトルの結果から、AGP 分子内に存在する三つのTrp
残 基(Trp25、Trp122、 Trp160)のいずれかが膜内部に存在することが示唆された。
どの
Trp
残基がそのような環境に移行するかを明らかにすることは、AGPの膜 への結合状態での構造特性を明らかにするとともに、α-ヘリックス構造形成部位の解明にもつながるものと思われる。その場合、一般的には発現系を用いて
Trp
変異体を作製して検討を行うのが常法であるが、現在までに組換え型AGP
が作製された例は報告されていない。そのため、これまでは、それぞれのアミ ノ酸残基に比較的特異的な修飾剤を用いて非可逆的修飾を行う、いわゆる化学 修飾法が汎用されてきた。しかし、この場合修飾剤の特異性や修飾アミノ酸残 基の同定の問題等により、特定のアミノ酸残基の役割を詳細に検討することは 困難であると考えられてきた。pPIC9
8.0 kb
5'AOX1 3'AOX1 (TT)
HIS4
3'AOX1 ColE1
Ampicillin
-factor s.s
Bgl II
Bgl II
AGP cDNA
Xho I EcoR I
Linearization wtih Bgl II
Pichia genome (HIS
+) Pichia genome (his4)
Figure 18. Expression construct for the recombinant AGP (rAGP)
in Pichia pastoris (GS115)
そこで、異種タンパク質発現宿主として、多くの組換え型タンパク質発現系 に用いられている、メタノール資化酵母である
Pichia pastoris(GS115
株)を 用いて、組換え型AGP(rAGP)の発現系を構築した
41-45)。Figure 18 に今回構 築した発現ベクター(pPIC9)を示す。酵母と大腸菌のシャトルベクターpPIC9
上のアルコールオキシダーゼ1(AOX1)プロモーター及びシグナル配列(
α-factor)の下流に AGP cDNA
を接続し、産生されたrAGP
が酵母のタンパク質分泌機構により、培地中に効率よく分泌されるように構築した。この発現ベク ターを制限酵素サイト
Bgl II
で直線化後、エレクトロポレーション法を用い、メタノール資化能を有する
Pichia pastoris
の染色体DNA
上にsingle crossover
で 挿入し、酵母の形質転換を行った。形質転換体の大量培養は、まず
1L
のBMGY
培地(1% yeast extract, 2%
polypepton, 100mM potassium phosphate (pH6.0), 1.34% yeast nitrogen base with ammonium sulfate without amino acids, 4 x 10
-5% biotin, 1% glycerol)(growth phase)
で培養を行い、その後、発現誘導炭素源であるメタノールを含む
1L
のBMMY
培地(1% yeast extract, 2% polypepton, 100mM potassium phosphate (pH6.0), 1.34%yeast nitrogen base with ammonium sulfate without amino acids, 4 x 10
-5% biotin, 1%
methanol)(induction phase)へと培地を交換し、96
時間培養を行った。また、rAGP
の発現誘導を維持するため、12時間毎に最終濃度が1%となるようにメタ
ノールを添加した。組換え体の精製は、遠心分離(6,000 x g, 10min, 4°C)により菌を分離後、培
地上清を
65%硫酸アンモニウム分画(pH4.0)で濃縮・粗精製を行い、その後、
陰イオン交換、ゲルろ過クロマトグラフィーに付し、透析、凍結乾燥を経て、
最終的に
rAGP
を得た。Figure 19 (A) には、得られた
rAGP
の SDS-PAGEの結果を示した。ヒトAGP
(hAGP)と比較して、rAGP では分子量の低下が観察された。
hAGP
分子上には
N-結合型糖鎖が五本存在しており、そのどれも複合型糖鎖と呼ばれるもので
あるが、酵母の場合すべて高マンノース型糖鎖と呼ばれるマンノースに富んだ 糖鎖が付加される。一般に、高マンノース型糖鎖の一本当たりの分子量は複合 型糖鎖のそれより小さい。そのため、このような組換え型での分子量の低下は、
酵母で付加された糖鎖含量の低下によると考えられた。また、
rAGP
では複数 のバンドの出現が観察されたが、Western blotting の結果より、これらはすべてAGP
由来のペプチドを有することが示唆された(Figure 19 (B))。(A) (B)
20.1 30 45 66 97
1 2 1 2
KDa
Figure 19. SDS-PAGE (A) and Western blotting (B) of hAGP (lane 1) and rAGP (lane 2)
それでは、なぜ分子量の異なる複数のバンドが出現したのであろうか。原因 として、次のことが考えられる。
1)発現-分泌過程において何らかの原因でペ
プチド鎖長の異なる複数のAGP
が出現した。2)細胞内での糖鎖付加過程にお いて、それぞれ糖鎖含量の異なるAGP
が分泌された。1)については、酵母細胞 内の代表的なタンパク質分解酵素であるカルボキシル・ペプチダーゼによるC
末端の切断が考えられた。しかし、データには示していないが、GS115 株のカ ルボキシル・ペプチダーゼを欠損させた株であるSMD1168
株を用いた場合やC
末端にヒスチジン・タグを付けてrAGP
を精製した場合も複数のrAGP
のバ ンドが観察されたことから、1)の可能性は低いと思われた。そこで2)につい
て検討するため、糖鎖切断酵素であるglycopeptidase F
(GPF)でrAGP
の糖鎖 を処理し、SDS-PAGEを行った(Figure 20)。GPF で糖鎖をすべて除去すると、hAGP と
rAGP
の分子量はほぼ同一となっ た。これは、rAGP のペプチド部分はhAGP
と同等のものであり、複数のバン ドはそれぞれ糖鎖含量の異なるAGP
が発現したために出現したと考えられた。また、データには示していないが、糖鎖結合アミノ酸である
Asn
残基を欠損さ せた変異体を用いた検討から、糖鎖含量の違いはそれぞれ付加された糖鎖の本 数が異なっていることに起因することが示唆された。事実、顆粒球マクロファージコロニー刺激因子を
Pichia
酵母で発現させた場合、糖鎖本数の異なる複数 のバンドが確認されている46)。KDa
20.1 30 45 66 97
1 2
Figure 20. SDS-PAGE of hAGP (lane 1) and rAGP (lane 2) treated with glycopeptidase F
5-3 組換え型 AGP
の構造・機能特性の評価hAGP と異なる
rAGP
の糖鎖分子の構造や含量が、AGP 自身の構造や機能特 性に影響を及ぼす可能性は十分に考えられる。そこで、構造特性においてはCD
スペクトル及びTrp
残基の蛍光スペクトルを(Figure 21)、機能特性において は代表的なAGP
結合性薬物であるクロルプロマジン、ワーファリン及びプロ ゲステロンを用いて薬物結合実験を行った(Figure 22)。Figure 21(A)の遠紫外領域の CD
スペクトルの結果から、hAGP とrAGP
の間に有意な差は観察されなかった。これは、