博士論文
CD63 発現の増加は BRAF 変異メラノーマ細胞の
BRAF 阻害剤耐性を抑制する
第 2 章 略語
AKT : AKT serine/threonine kinase BSA : Bovine Serum Albumin CD : Cluster of differentiation CDK4 : Cyclin-dependent kinase-4 cDNA : complementary DNA
CTLA-4 : Cytotoxic T-Lymphocyte Antigen-4 CXCR4 : C-X-C chemokine receptor type-4 EGFR : Epidermal growth factor receptor ERK : Extracellular signal-related kinase esiRNA : endoribonuclease-prepared siRNA FACS : Fluorescence activated cell sorting FAK : Focal adhesion kinase
FBS : Fetal Bovine Serum GTP : Guanosine triphosphate HA : Hemagglutinin antigen HGF : Hepatocyte growth factor HIV : Human Immunodeficiency Virus
IGF-1R : Insulin like growth-factor 1 receptor MAPK : Mitogen-activated Protein Kinase MDR-1 : Multidrug Resistance Protein-1 MEK : MAPK/ERK kinase
mRNA : messenger RNA
LAMP-1 : Lysosomal associated membrane protein-1 LAMP-2 : Lysosomal associated membrane protein-2 PBS : Phosphate Buffered Saline
PCR : Polymerase chain reaction PD-1 : Programmed cell death 1
PDGFR : Platelet derived growth factor receptor PMEL : Premelanosome protein
PI3K : Phosphatidylinositol 3-kinase PI4K : Phosphatidylinositol 4-kinase
PTEN : Phosphatase and Tensin Homolog Deleted from Chromosome 10 qRT-PCR : quantitative Reverse Transcription-Polymerase Chain Reaction SDS-PAGE : SDS-Polyacrylamide Gel Electrophoresis
SDC4 : Syndecan-4
siRNA : small interfering RNA RPN-2 : Ribopholin-2
TBS : Tris-Buffered Saline
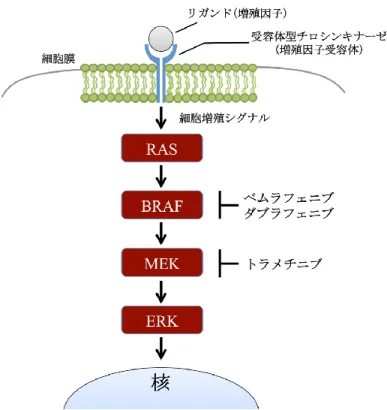
Figure 1. BRAF 変異および NRAS 変異によるメラノーマの悪性化促進機構 正常な細胞では増殖因子などの刺激があった場合のみ、RAS-RAF 経路を介して細胞内にシグナルが伝 達する。しかし、メラノーマ細胞では NRAS や BRAF に遺伝子変異が認められ、これらの変異により MEK-ERK 経路など下流シグナルが恒常的に活性化することでメラノーマ細胞の悪性化が促進されて いる。 3-2 実臨床におけるメラノーマ治療法 メラノーマの実臨床における治療では、根治切除可能な腫瘍は外科的手術によって 切除される。一方で、根治切除不能な腫瘍に対しては、これまで細胞障害性の薬剤を 用いた薬剤療法または放射線療法が一般的な治療法であった。しかし、メラノーマは 非常に悪性度の高いがん種であり、それらの治療法に高い抵抗性を示す。そのため、 ほとんど治療効果を望むことはできず、長年、より治療効果の高い薬剤の開発が望ま れていた。近年、様々な機構を標的とした新規薬剤が開発されたことにより、これま で外科的手術による根治が望めなかった後期ステージの進行性メラノーマに対しても、 治療の選択肢が持てるようになった。根治切除不能な腫瘍に対しては遺伝子変異の検 査が行われ、この際に BRAF に遺伝子変異が認められれば、BRAF 阻害剤を用いた薬 剤療法を行う。また、メラノーマの腫瘍内は免疫抑制性の微小環境であることから (Lizee et al., 2006; Khalili et al., 2012)、疲弊した免疫系を活性化することが有効な治療 法だと考えられている。そのため、BRAF 変異の有無に限らず、Programmed cell death-1 (PD-1) を 標 的 と す る ニ ボ ル マ ブ や ペ ム ブ ロ リ ズ マ ブ 、 Cytotoxic T-Lymphocyte Antigen-4 (CTLA-4) を標的とするイピリムマブなどの免疫チェックポイント阻害剤と 呼ばれる抗体薬を用いる場合もある (Fig. 2)。しかしながら、メラノーマにおける免 疫チェックポイント阻害剤の単剤奏効率は約 20~30%であり、効果を示さない患者も 少なくない (Postow et al., 2015; Yamazaki et al; 2017)。
された HGF は、その受容体である c-MET を介して腫瘍細胞に働き、Phosphoinositide 3-kinase (PI3K) - AKT serine/threonine kinase (AKT) 経路を活性化することでメラノー マ細胞に薬剤耐性を付与する。そのため、AKT 阻害剤または MET 阻害剤を BRAF 阻 害剤と併用することで、薬剤耐性を減弱させることが可能である (Wilson et al., 2012)。 また、がん微小環境に存在するがん随伴性線維芽細胞は、BRAF 阻害剤により活性化 し、細胞外マトリックス分子フィブロネクチンに富んだ微小環境を再構築することが 示された (Hirata et al., 2015)。この特徴的な微小環境はメラノーマ細胞におけるインテ
グリン1-Focal adhesion kinase (FAK) シグナル伝達を増強することで ERK の再活性化 および BRAF 阻害剤に対する耐性を引き起こす。そのため、BRAF 阻害剤と FAK 阻害 剤の併用は有望な治療候補になると考えられている。
一方で、メラノーマ細胞自体の自律的な薬剤耐性獲得機構は、薬剤療法として BRAF 阻害剤が投与される以前から先天的にメラノーマ細胞に備わっていた自然耐性と治療 過程でメラノーマ細胞自体が後天的に獲得した獲得耐性の 2 つに分類される。その中 で も 自 然 耐 性 に お け る 代 表 的 な も の が 、 が ん 抑 制 遺 伝 子 Phosphatase and Tensin Homolog Deleted from Chromosome 10 (PTEN) の 欠 失 お よ び 細 胞 周 期 制 御 分 子 Cyclin-dependent kinase-4 (CDK4) の活性化変異とその補助因子サイクリン D1 の増幅 である (Smalley et al., 2008; Nathanson et al., 2013) (Fig. 4)。PTEN の欠失は 10%以上の メラノーマで認められ (Paraiso et al., 2011)、PI3K-AKT 経路の活性化を増強すること で BRAF 阻害剤に対する耐性を引き起す (Trunzer et al., 2013)。CDK4 の活性化変異と サイクリン D1 の遺伝子増幅は BRAF 阻害剤が引き起こす細胞周期の停止を阻害する ことで、耐性を引き起こすと考えられている (Smalley et al., 2008)。国内初の CDK4 に
対する阻害剤が進行乳がん適応として数ヶ月前に承認されたばかりであり、BRAF 阻 害剤との併用で良好な効果を示すことが報告されていることから (Flaherty et al., 2012; Yadav et al., 2014; Yoshida et al., 2016)、今後メラノーマに対する適応拡大が期待 される。
細胞に備わっていた自然耐性と、治療過程でメラノーマ細胞自体が後天的に獲得した獲得耐性の 2 つ に分類される。
3-4-1 細胞膜 4 回貫通型タンパク質—テトラスパニン
テトラスパニンは膜貫通ドメインを 4 つ持ち、細胞外領域に大小 1 つずつのループ 構造を構築しているタンパク質の総称である (Fig. 5)。ヒトでは 33 種類、ショウジョ ウバエでは 37 種類同定されており (Charrin et al., 2014; Termini et al., 2017)、アメーバ や真菌などの単細胞生物においても存在していることが報告されているため (Albers et al., 2016)、広域な生物種で保存されている。生体内での分布においても非常に多様 であり、テトラスパニンファミリーメンバーCD9 や CD63 などは、ほとんどの組織や 細胞で一様に発現しているが、それ以外のメンバーの多くは特異的な組織や細胞に分 布を示す (Maeker et al., 1997)。この生体内の分布の多様性はテトラスパニンが様々な 状況において細胞におけるシグナル伝達を制御することを可能にしている。テトラス パニンは細胞接着やシグナル伝達に関与するタンパク質などとマイクロドメインを形 成し、それらのタンパク質のシグナル伝達の足場として機能することで細胞の増殖 (Franco et al., 2010; Sadej et al., 2010)、移動 (Funakoshi et al., 2003; Hong., et al., 2012; Seubert et al., 2015)、形態変化 (Kazarov et al., 2002; Gustafson-Wagner et al., 2013) など を制御している。
であるが (Atkinson et al., 1984; Hotta et al., 1988)、細胞においては主に後期エンドソー ムおよびリソソームなどの細胞内オルガネラに局在する。この細胞内オルガネラへの ソーティングには CD63 の C 末端領域に存在する lysosomal sorting motif-GYEVM が重 要であることが報告されている (Rous et al., 2002)。
CD63 は相互作用分子と直接的または間接的に複合体を形成することで、それらの 分子の局在や活性を変化させ、細胞機能を制御する。複合体を形成する相互作用分子 は多岐にわたっており、代表的なものとしては、細胞接着分子インテグリンやケモカ イン受容体を代表とした細胞膜受容体 (Berditchevski et al., 1996; Skubitz et al., 1996; Yoshida et al., 2008)、他のテトラスパニン分子 (Radford et al., 1996; Hammond et al., 1998; Israels et al., 2001)、Phosphatidylinositol 4-kinase (PI4K) やマトリックスメタロプ ロテアーゼなどの酵素が挙げられる (Berditchevski et al., 1997; Takino et al., 2003; Jung et al., 2006)。
CD63 の生理的機能として、メラノサイトタンパク質 premelanosome protein (PMEL) と複合体を形成し、PMEL のメラノソームへのソーティングを制御することで、メラ ノソームの成熟に関与している (van Niel et al., 2011)。一方で、CD63 はマトリックス
メ タ ロ プ ロ テ ア ー ゼ の 内 因 性 阻 害 因 子 Tissue inhibitors of metalloproteinases-1 (TIMP-1) の受容体でもあり (Jung et al., 2006)、細胞内にシグナルを伝達することで、 顆粒球産生などの生理機能にも寄与することが報告された (Kobuch et al., 2015)。しか し 、 CD63 遺 伝 子 欠 損 マ ウ ス の 示 す 表 現 型 は 軽 度 の 腎 障 害 の み で あ る こ と か ら (Schroder et al., 2009)、健常な生体内では CD63 が欠損した状態においても、その機能 を補う代替的な分子が存在することが考えられる。
とが報告されている (Yoshida et al., 2008)。そのため、CD63 の導入が HIV-1 感染に対
する遺伝子治療法として有望かもしれないと考えられている。さらに、がんとの関与 に対しても盛んに研究が行われており、メラノーマ、甲状腺がん、乳がん、急性骨髄 性 白 血 病 な ど の が ん 種 で 悪 性 形 質 の 亢 進 と CD63 の 関 与 が 報 告 さ れ て い る (Bommarito et al., 2011; Toricelli et al., 2013; Tominaga et al., 2014; Forte et al., 2017) 。そ の中でも最も研究が進んでいるがん種がメラノーマである。初期ステージのメラノー マ細胞では、メラノサイトと比較して CD63 の発現が増加し、インテグリン1 および TIMP-1 と複合体を形成するようになる。この複合体形成は下流の PI3K 経路を活性化 し、足場非依存的な細胞死に対する耐性をメラノーマ細胞に付与している (Toricelli et al., 2013)。一方で、CD63 はメラノーマの細胞移動、上皮間葉転換、腫瘍形性能を負に
CD63 は細胞外領域ループ構造に 3 つの N-型糖鎖付加部位を有する糖タンパク質である。小胞体で合 成され、糖鎖付加を受け、ゴルジ体に輸送後に高度に糖鎖がプロセシングされる。Melanoma-associated antigen (ME491) という別名でも知られる。CD63 は初期ステージのメラノーマ細胞では発現が増加す る一方、メラノーマの進行に伴ってその発現が減少することから、メラノーマのステージ特異的なが ん抗原だと考えられている。 3-5 本研究の背景と目的 私は予備的な実験から以下の 2 つの結果を得ていた。 ① Vemurafenib 処理によりメラノーマ細胞の CD63 平均分子量が増加する
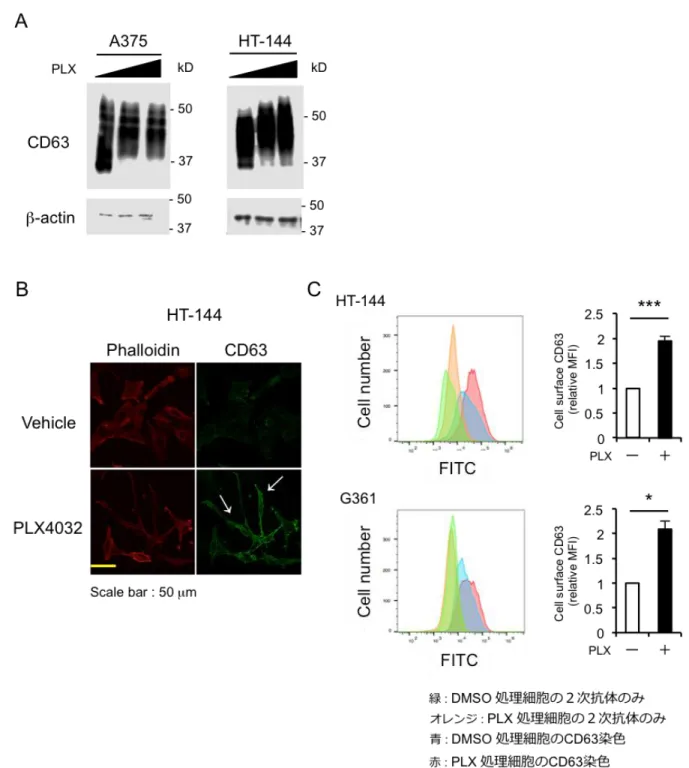
ヒト BRAF 変異メラノーマ細胞株 A375 細胞を Vemurafenib で処理したところ、 CD63 のタンパク質量に変化は無かったが、平均分子量の大幅な増加が認められた。 他のヒト BRAF 変異メラノーマ細胞株においても同様の現象が観察された。
② Vemurafenib 処理によりメラノーマ細胞の細胞膜上の CD63 量が増加する
ヒト BRAF 変異メラノーマ細胞株 A375 細胞を Vemurafenib で処理したところ、細 胞染色により細胞膜上の CD63 量の顕著な増加を見出した。他のヒト BRAF 変異メ ラノーマ細胞株においても同様の現象が観察された。 乳がん細胞において、小胞体で N-型糖鎖付加をする酵素複合体構成因子の 1 つであ る Ribopholin-2 (RPN-2) が微小管阻害薬であるドセタキセルへの薬剤耐性を増強して おり (Honma et al., 2008)、RPN-2 によって糖鎖付加が制御され、薬剤耐性に関与する 因子が CD63 だと報告された (Tominaga et al., 2014)。CD63 は糖鎖が付加されると細胞
膜に局在するようになり、その際に薬剤耐性関連分子 Multidrug Resistance Protein-1 (MDR-1) の細胞膜への移行を制御することで、ドセタキセルへの薬剤耐性を増強する (Tominaga et al., 2014)。また、CD63 は付加される糖鎖の種類により不均一な分子量を
第 4 章 実験方法
1) 抗体Mouse anti-HA antibodies, Mouse anti--actin antibodies, Mouse anti--tubulin antibodies は Sigma Aldrich (USA) から購入した。 Mouse anti-CD63 mAb antibodies は BioLegend (USA) から購入した。Rabbit anti-Flag antibodies, Rabbit anti-ERK1/2 antibodies, Rabbit anti-phospho-ERK1/2 (Thr202/Tyr204) antibodies, Rabbit anti-Akt antibodies, Rabbit anti-phospho-Akt (Ser473) antibodies, Rabbit anti-GAPDH antibodies は Cell Signaling Technology (USA)から購入した。Mouse anti-Syndecan-4 antibodies 5G9 は Santa Cruz Biotechnology (USA) から購入した。Alexa Fluor labeled secondary antibodies は Dako (USA) から購入した。EZ Blot anti-rabbit IgG HRP は Gene Tex (USA) から購入した。
2) 細胞培養
ヒト BRAF 変異メラノーマ様細胞株 A375、HT-144、G361、SK-MEL28 (American Type Culture Collection (ATCC)), ヒト膵がん細胞株 PANC-1 (ATCC), ヒト乳がん細胞株 MDA-MB231 (ATCC), レトロウイルスパッケージング細胞株 PlatA は 10% 非働化ウシ 胎児血清 (FBS,Biosera, France)、1.5 mM L-glutamine、100 U/ml penicillin、
streptomycin を含む RPMI 1640 培地 (Thermo Fisher Scientific, USA) 中、37℃、5% CO2
下で培養された。ヒト角化細胞株 HaCaT (ATCC) は上記と同じ条件下で DMEM (高グ ルコース) (和光純薬工業, Japan) 中で培養された。A375, HT-166, SK-MEL-28 細胞由来 Vemurafenib 耐性株は、以前に当研究室で作製された (Satow et al., 2017)。実験に応じ て、各種細胞は、示された濃度の PLX4032 (Vemurafenib) (AdooQ BioScience, USA), UO126 (Cell Signaling Technology), Tunicamycin (Sigma Aldrich) が添加された増殖培地 中で、示された時間培養された。
3) Small interfering RNA (siRNA) を用いた遺伝子発現抑制
メラノーマ細胞を12 well dish (Thermo Fisher Scientific) に8×104 cells/well となるよう
播種した。24 時間後、ネガティブコントロール siRNA (All stars; Qiagen, USA) ある い は 3GNT-2 に 対 す る 2 種 の siRNA (si3GNT-2#1:Hs_3GNT2_2; si3GNT-2#2:Hs_3GNT2_3; Qiagen) を Lipofectamine RNAiMAX 試 薬 (Invitrogen, USA) を用いて導入した。導入方法は製品プロトコールに順じた。細胞は、2日間培養 後に解析した。
4) プラスミド作製
トロールとしてベクターのみを Lipofectamine 2000 (Invitrogen) を用いてメラノーマ 細胞に導入した。導入方法は製品プロトコールに順じた。A375およびHT-144 は 2 mg/ml, G361 は1 mg/mlのG418 (Life Techonologies, USA) でそれぞれのプラスミド発 現細胞を選択し、A375細胞はその後、限界希釈法によりクローン化を行った。 A375細胞へのSDC4 ET-CD63遺伝子 (キメラタンパク質遺伝子) 導入についても hCD63-Flag遺伝子導入と同様の条件で行った。キメラタンパク質遺伝子がサブクロー ニングされたpMXs-IP SDC4ET-hCD63 (WT), pMX-IPs SDC4ET-hCD63 (AAA) あるい はコントロールとしてpMXs-IP SDC4-ET, ベクターのみを導入した細胞を3 g/mlの Puromycin (Sigma Aldrich) で処理することで、それぞれのプラスミド発現細胞を選択 した。
6) ウェスタンブロッティング
目的の処理を施した細胞を冷PBSで洗浄後、可溶化バッファーと超音波処理により 可溶化し、22,140 x g、20 分間、4℃ の遠心分離により得られた上清を細胞可溶化液 とした。これにLaemmli sample buffer を加え、100℃で 5 分間加熱し、ウェスタンサ ンプルとした。SDS-PAGE で分離したタンパク質を PVDF 膜 (BioRad, USA) に転写 し、膜を10% スキムミルク (Yukijirushi, Japan) /TTBS もしくは 5% BSA (和光純薬工 業) /TTBS でブロッキングした。一次抗体を 5% スキムミルク/TTBS もしくは 5% BSA/TTBS もしくは Can Get Signal Solution 1 (TOYOBO Life Science) で適当な濃度 に希釈し、室温で 1 時間もしくは 4℃ で一晩反応させた後、メンブレンを TTBS で 洗 浄し た 。 その 後 HRP-標識 二 次 抗体 を 5% ス キ ム ミル ク /TTBS も し くは 5% BSA/TTBS もしくは Can Get Signal Solution 2 (TOYOBO Life Science) で希釈し、室温 で 1 時間反応させた。続いて TTBS で洗浄し、ECL Western blotting Detection Reagent (GE Healthcare Life Sciences, UK) もしくは ImmunoStar LD (和光純薬工業) を用いて 検出した。検出は、C-DiGit Blot Scanner (LI-COR, USA) を用いて行った。解析は、Image Studio (LI-COR) を用いて行った。
RNeasy Tissue Kit (Promega, USA) を用いて、キットプロトコルに従いtotal RNAを調 製した。 のRNAからReverTra Ace qPCR RT Master Mix(TOYOBO Life Science)を 用いてcDNAを合成した。このcDNAをテンプレートとしてTHUNDERBIRDTM SYBR
qPCR Mix (TOYOBO Life Science) と 以 下 に 示 す プ ラ イ マ ー を 用 い て 、 C1000TM
Thermal Cycler (Bio-Rad CFX 96 TM Real-time System, USA) によりPCR反応を行った。 データは全て内部標準として用いたGapdhにより補正した。実験は3回繰り返した。
ヒト CD63: Forward 5’- ATGCAGGCAGATTTTAAGTGCT -3’ Reverse 5’- GTTCTTCGACATGGAAGGGATTT -3’ ヒト3GNT-2 Forward 5’- ATACTGGAACCGAGAGCAAG -3’ Reverse 5’- TCAGGTTCGCAGTAGTTCAG -3’ ヒト GAPDH Forward 5’- GTCGCTACAGTGACCTGCTG -3’ Reverse 5’- GTCTTTGAGCGTCTGGTTGA -3’
8) フローサイトメトリー法 (Fluorescence activated cell sorting : FACS) による細 胞表面抗原の解析
9) Clonogenic assay 6 ウエルプレート1ウエルに 400 個の細胞を播種し、一晩培養した後、PLX4032 (5 M) またはコントロールとして DMSO を添加した培地で培養した。培地交換は 2 日 または 3 日毎に行い、10 日後、細胞を PBS で洗浄し、2% Glutaraldehyde (和光純薬工 業) /PBS で 10 分間固定した。PBS で洗浄後、0.1% Crystal Violet 溶液 (和光純薬工業) で細胞を 20 分間染色し、1% SDS 溶液で溶出、プレートリーダー (Corona Electric, Japan) で 590 nm の吸光度を計測することで、生存細胞数を定量化した。 10) 免疫染色法 5×104 個のメラノーマ細胞を 6 well plate 中の 12 mm ガラスカバースリップ (Matsunami, Japan) 上に播種した。目的の処理を行った後、48 時間培養し、細胞を 4% PFA/PBS で固定、PBS で 2 回洗浄し、ブロッキング溶液 (5% BSA/PBS) を加えて 30 分間ブロッキングを行った。一次抗体あるいはビオチン化 LEA レクチン (Vector Laboratories) を 5% BSA/PBS で適当な濃度に希釈し、4℃ で一晩反応させた後、細胞 を PBS で洗浄した。その後 0.1% Alexa Fluor 標識二次抗体 (Thermo Fisher Scientific) あるいは 0.1% Alexa Flouor 標識 streptavidin (BioLegend) と 0.5% Alexa Fluor 標識 Phalloidin (Invitrogen) (F-actin 細胞骨格染色) および Hoechst 33342 (Dojindo) (核染色) を 5% BSA/PBS で希釈し、室温で 1 時間反応させた。PBS で 3 回洗浄した 後 に 、 ProLong Diamond Antifade Moutant (Invitrogen) に て 封 入 し 、 蛍 光 顕 微 鏡 (BZ-9000/Keyence, Japan、Plan Apo 20×/0.75, Nikon, Japan、Plan Apo 60×/1.40 Oil, Nikon) あるいは共焦点顕微鏡 (FV1000/Olympus, Japan、UPlanSApo 60×/1.35 Oil, Olympus、 UPlanSApo 100×/1.40 Oil, Olympus) を用いて観察した。
11) 細胞膜タンパク質のビオチン化法
LS-biotin (Pierce, USA) / PBSを添加して、4 ℃で 30 分間反応させ、細胞表面をビオチ ン標識した。続いて、細胞を 100 mM グリシン/ PBS で4℃、10 分間処理することで クエンチングを行い、冷 TBS で洗浄した。その後、可溶化バッファー (1% Triton-X100、 0.25% SDS、1 mM EDTA、1x protease inhibitor cocktail / TBS) で可溶化し、22,140 x g、 5 分間、4℃ で遠心分離に供した。上清を回収後、この溶液 16 ul 分に sample buffer 4 ul を添加し、これを input 画分とし、残った溶液のうち 200 l に 100 l のストレ プトアビジンビーズ (GE Healthcare) を添加し、4℃で 1 時間反応させた。用いたス トレプトアビジンビーズの量は、ビオチン化されたすべてのたんぱく質をプルダウン するのに十分であることは確認した。ビーズを可溶化バッファーで3回洗浄後、ビー ズに20 lのsample bufferを加え、100℃で 5 分間加熱処理を行い、ビオチン標識タン パ ク 質 画 分 と し た 。 Flag 抗 体 を 用 い た ウ エ ス タ ン ブ ロ ッ ト に よ り 、 input 中 の CD63-Flag に対するビオチン化された CD63 量を定量化した。
12) Lycopersicon Esculentum (Tomato) レクチンビーズを用いたポリグルコサミン 陽性タンパク質のプルダウン法
G361細胞またはHT-144細胞を5 MのPLX4032あるいはDMSOを添加した培地で48 時間処理し、PBSで2回洗浄後、Cell Dissociation Bufferで細胞を浮遊させた。無血清培 地に懸濁した細胞を 37℃で30分間静置したのち、5x105 個の細胞を 500 l の 1%
Triton X-100 含有バッファーA (20 mM Hepes pH7.5、150 mM NaCl、0.1 mM CaCl2、1x
sample buffer に溶解し、糖溶出画分とした。CD63 抗体を用いたウエスタンブロット により、処理による LEA 結合画分中の CD63 量の変化を解析した。
13) 免疫沈降およびN-型糖鎖遊離酵素消化
CD63 (WT)-FlagあるいはCD63 (AAA) -Flagを発現したA375 細胞を5 MのPLX4032 あるいは DMSO を添加した培地で72時間処理した。細胞を冷 TBS で洗浄後、可溶 化バッファー (50 mM Tris-HCl pH7.4、150 mM NaCl、1% Triton X-100、0.25% SDS、 1mM EDTA、protease inhibitor cocktail) で可溶化し、その溶液を22,140 x g、5 分間、4℃ で遠心分離し、得られた上清を細胞可溶化液とした。一部は input 画分として調製し、 残りの溶液には抗 Flag 抗体 (Cell Signaling) を結合させた protein A-sepharose (GE Healthcare) ビーズを添加し、4℃で 1 時間反応させた。ビーズを可溶化バッファーで 3 回洗浄後、変性バッファー (0.5% SDS、1% 2-mercaptoethanol/20 mM Tris buffer PH 8.8) を加え、100℃で 3分間加熱処理を行うことで、結合タンパク質をビーズから溶 出した。溶出したタンパク質を酵素消化バッファー (3.5% NP-40、protease inhibitor cocktail /20 mM Phosphate buffer) 中でN-glycosidase (PNG) F (Takara Bio, Japan) と 37 度で一晩反応させ、sample buffer 添加後 100℃で 5分間加熱処理することで反応を停 止させた。Flag 抗体と非変性 rabbit IgG を認識する EZ Blot 抗 rabbit IgG HRP (Gene Tex, USA) を用いたウエスタンブロットにより分子量変化を解析した。
14) 統計解析
全ての数値的 な実験 データに対し て、有 意差検定ソフ トウェ ア SigmaPlot○R 12.5
第 5 章 実験結果
5-1 Vemurafenib 処理により CD63 の平均分子量および細胞膜局在が増加する
CD63 は、細胞膜上で様々なパートナー分子と相互作用することで細胞機能を制御 している (Berditchevski et al., 1995; Toricelli et al., 2013; Park et al., 2015)。さらに近年、
がん細胞の細胞膜上での CD63 の増加が抗がん剤への耐性に寄与していることが報告 されている (Tominaga et al., 2014)。そのため、Vemurafenib 処理がメラノーマ細胞にお
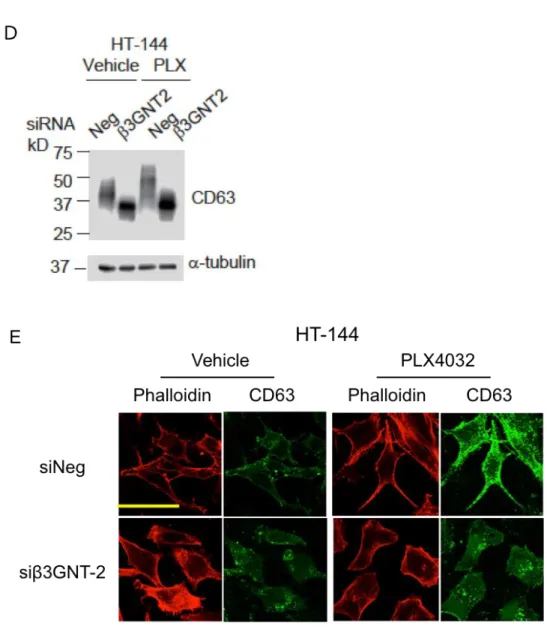
Figure 7. Vemurafenib 処理により CD63 の平均分子量および細胞膜局在が増加する
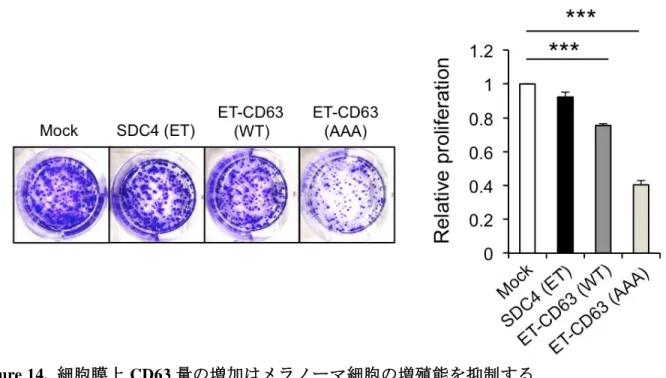
Figure 9. CD63 過剰発現により Vemurafenib 存在下でのメラノーマ細胞の増殖が抑制される
照細胞およびhCD63–Flag過剰発現細胞の各クローン (400個) を播種し、PLX4032 存在下あるいは非存 在下で10日間培養し、生存した細胞数を比較した。グラフはMock #1の値を1として相対的に示した。 PLX4032 存在下で、hCD63-Flag 発現細胞の生存数は対照に比べ著しく低いことが認められた。(平均 値±SEM) *:P < 0.05 (D) 対照細胞およびhCD63–Flag発現A375細胞の各クローンを PLX4032 存在下あ るいは非存在下で 48 時間培養し、ERKとAKTの活性をWBで解析した。PLX4032非存在化において、 Mock細胞とhCD63–Flag発現細胞の間に、リン酸化ERKおよびリン酸化AKT量の変化は認められなかっ た。一方、薬剤処理によりリン酸化ERK量の減少が観察されたが、Mock細胞とhCD63–Flag発現細胞の 間に差は認められなかった。 5-4 Vemurafenib 処理による CD63 平均分子量の増加は糖鎖構造の変化による BRAF 変異メラノーマ細胞への Vemurafenib 処理はメラノーマ細胞の CD63 の平均分 子量および細胞膜局在を増加させることを見出した (Fig. 7)。CD63 は糖鎖が付加され ると細胞膜での局在が増加し、細胞膜の CD63 量の増加が乳がんにおいては抗がん剤 への薬剤耐性を増強することが報告されている (Tominaga et al., 2014)。また、一般に 糖タンパク質に付加される糖鎖は不均一であるため、糖タンパク質は不均一な分子量 を示し(Liang et al., 2014; Boman et al., 2016)、CD63 も例外ではない。これらのこと
Figure 10. Vemurafenib 処理による CD63 平均分子量の増加は糖鎖構造の変化による
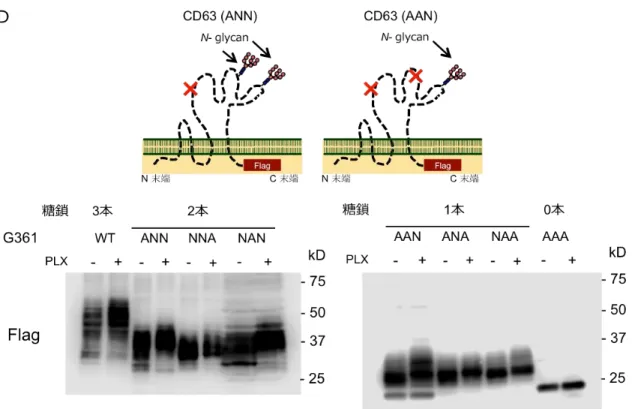
(A) HT-144 細胞を PLX4032 (5 M) 単独あるいは Tunicamycin (0.1 g/ml) 共存下で 48 時間培養し、 CD63 の分子量を WB により解析した。PLX4032 処理で増加した分子量は、低濃度の Tunicamycin 共 存下では観察されず、CD63 のコアペプチドに相当する 25kDa 分子種が検出された (白線は、ブロット が同じ膜の異なる部分から得られていることを示す)。(B) CD63 の N-型糖鎖付加部位変異型 hCD63 (AAA) -Flag および CD63 (WT) -Flag をそれぞれ発現する A375 細胞を作製した。hCD63 (WT) -Flag で は、PLX4032 処理により内在性 CD63 と類似した平均分子量の増加が観察された一方、hCD63 (AAA) -Flag では hCD63 のコアペプチドと一致する 25kDa タンパク質が検出され、薬剤処理による分子量変 化が認められなかった。(C) hCD63 (WT) -Flag および hCD63 (AAA) -Flag 発現細胞を PLX4032 存在下で 72 時間培養後、hCD63-Flag タンパク質を抗 Flag 抗体により免疫沈降した。これを溶出し、N-型糖鎖遊 離酵素ペプチド N-グリコシダーゼ F (PNG) 消化を供し、WB により解析した。薬剤の存在に関わらず、 WT の分子量が PNG 消化により AAA と同じ 25kDa まで減少した。 (D) WT, AAA に加え、CD63 の N-型糖鎖付加部位変異型 hCD63-Flag (ANN, NNA, NAN, AAN, ANA, NAA) -Flag をそれぞれ発現する G361 細胞を樹立した。PLX4032 (5 μM) 存在下あるいは非存在下で 48 時間培養し、CD63 の分子量を WB により解析した。薬剤処理により AAA では分子量の変化が認められなかった一方、少なくとも 1 ヶ所 の糖鎖修飾部位を有する変異型 hCD63 (ANN, NNA, NAN, AAN, ANA, NAA) では薬剤処理による平均 分子量の変化が認められた。
加が糖鎖修飾系の変化によるのかを Tunicamycin を用いて検討した。Vemurafenib 処理 への感受性が高い HT-144 細胞および G361 細胞をそれぞれ Vemurafenib 単独あるいは Tunicamycine 共存下で培養し、免疫染色およびフローサイトメトリーにより細胞表面 上の CD63 量を比較した。免疫染色の結果より、Vemurafenib 処理で増加する細胞膜上 の CD63 量が、Tunicamycin 共存下により減少し、対照細胞と比べても増加が認められ なかった (Fig. 11A)。さらに、フローサイトメーターを用いた HT-144 細胞の解析で も、Vemurafenib 処理細胞での増加は、低濃度の Tunicamycin 共存下で検出されなかっ た (Fig. 11B)。これらの結果から、Vemurafenib 処理による細胞膜上 CD63 の量の増加 には、糖鎖の付加が必要であることが示唆された。 次に、薬剤存在下での細胞膜上における野生型 hCD63 (WT) -Flag の増加に糖鎖付加 が必要か検討するため、変異型 hCD63 (AAA) -Flag と野生型 hCD63 (WT) -Flag をそれ ぞれ発現する HT-144 細胞を作製した。これらの細胞の細胞表面上のタンパク質をビ オチン標識し、ストレプトアビジンビーズで沈降することで、細胞膜上の hCD63-Flag 画分を得た。WB により、総 flag タンパク質あたりの細胞膜上での hCD63-Flag 存在比 を解析することで、薬剤処理による細胞膜上の CD63 量の増加に糖鎖の付加が重要か 検討した。薬剤存在下において、細胞膜上での hCD63 (WT) -Flag の存在量は対照に比 べ平均で 2.5 倍増加したが、hCD63 (AAA) -Flag ではむしろ約半分に減少した (Fig. 11C)。このことから、薬剤処理による細胞膜上 CD63 量の増加には CD63 への糖鎖の 付加が必要で、Fig. 10 の結果とあわせ、その構造の変化が関与することが示唆された。
Figure 12. Vemurafenib 処理により CD63 に付加するポリラクトサミン量が増加する
やかな変化であった。これらのことから、Vemurafenib 処理による CD63 の平均分子量 および細胞膜局在の増加には、部分的に MEK-ERK 経路の阻害が寄与していることが 示唆された。
Figure 13. Vemurafenib 処理が引き起こす CD63 の平均分子量および細胞膜上の量の増加は部
分的に MEK-ERK 経路の阻害を介する
が予想された。
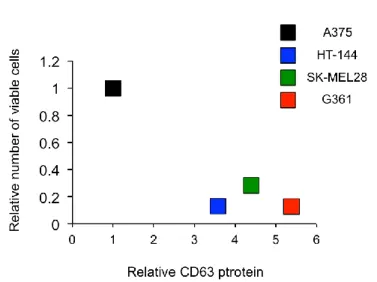
Figure 14. 細胞膜上 CD63 量の増加はメラノーマ細胞の増殖能を抑制する
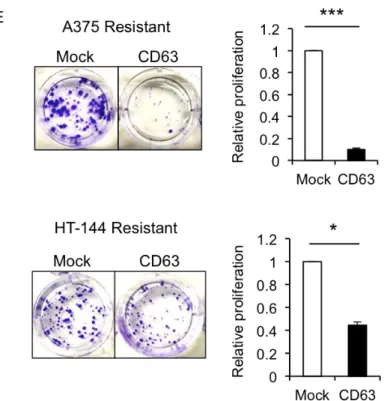
討した。まず、A375 細胞、HT-144 細胞および SK-MEL28 細胞の Parental 細胞と Resistant 細胞における CD63 タンパク質量をWBにより解析したところ、それぞれの細胞由来 の Resistant 細胞クローン 2 つでは、Parental 細胞と比較して、CD63 量の顕著な減少が 認められた (Fig. 15A)。37~50kDa のスメアバンドとして検出される Parental 細胞での CD63 と異なり、Resistant 細胞の発現する CD63 は分子量 50kDa 付近を示す、高分子 CD63 が主なものであった。さらに、Resistant 細胞での CD63 タンパク質の減少が mRNA レベルで起きているのか検討するため、Parental 細胞および Resistant 細胞での CD63 mRNA 量の差を定量的 RT-PCR を用いて測定した。その結果、A375 細胞由来の Resistant 細胞では Parental 細胞と比較して、40%程度の CD63 mRNA 量の減少が観察され、 HT-144 細胞由来および G361 細胞由来の Resistant 細胞では 40~60%の減少が認められ た。(Fig. 15B)。次に、フローサイトメトリーにより、Resistant 細胞における細胞表面 上の CD63 存在量の変化を解析した。フローサイトメーターを用いた解析から、HT-144 細胞由来および SK-MEL28 由来の Resistant 細胞では、Parental 細胞と比較して、細胞 表面上の CD63 存在量が 40%~60%減少していることが観察された (Fig. 15C)。またこ の際、元来 CD63 の発現量の低い A375 細胞由来の Resistant 細胞では有意な細胞表面 上の CD63 存在量の減少は認められなかった。A375 Parental 細胞は元々、他の細胞に 比べ CD63 発現量が顕著に低いことから、Parental 細胞と Resistant 細胞間での減少す る総 CD63 量が少量であり、細胞膜上 CD63 量の減少に反映していないことが考えら れる。これらの結果から、Resistant 細胞では Parental 細胞に比べ、mRNA レベルで CD63 の発現が減少しており、いくつかの細胞では細胞膜上の CD63 量も相関して減少して いることが示唆された。
Figure 15. CD63 過剰発現は Vemurafenib 耐性メラノーマ細胞の増殖能を抑制する
第 6 章 考察
するのかもしれない。また、Galectin の N-型糖鎖に対する親和性は、コア構造からの 分 岐 の 数 と N- ア セ チ ル ラ ク ト サ ミ ン の 数 に 比 例 す る こ と が 報 告 さ れ て い る (Hirabayashi et al., 2002; Lau et al., 2007)。このことから、本研究で示唆された長鎖の
Resistant 細胞での低い CD63 量 Parental 細胞に比較して Resistant 細胞で認められる CD63 の顕著に低い発現は薬剤 存在下における耐性細胞の増殖促進機構の 1 つであろうと考える (Fig. 15)。この CD63 発現量の変化は mRNA レベルで認められたことから、CD63 の上流に存在する転写因 子の関与が示唆された。メラノーマの薬剤耐性に関与する因子であり、CD63 の上流 に存在する転写因子として、小眼球症関連転写因子 MITF (Microphthalmia-associated transcription factor) がある (Ploper et al., 2014; Muller et al., 2014; Ji et al., 2016)。 薬剤 耐性を獲得したメラノーマ細胞では MITF の発現減少が高頻度で検出されており (Ji et al., 2016)、薬剤耐性患者由来のメラノーマ組織でも、その発現減少が認められてい
る (Muller et al., 2014)。そのため、薬剤耐性細胞における MITF の発現減少が下流の
CD63 mRNA 量の減少を引き起こしているのかもしれない。
耐性を予想するマーカーについての報告はいくつかなされているが (Baudy et al., 2012; Muller et al., 2014; Richard et al., 2016)、エクソソームについての知見はない。そ
第 7 章 参考文献
1. Albers T, Maniak M, Beitz E, et al. The C Isoform of Dictyostelium tetraspanins localizes to the contractile vacuole and contributes to resistance against osmotic stress. PLos ONE. 2016; 11:e0162065
2. Atkinson B, Ernst CS, Ghrist BF, et al. Immunoassay for melanoma-associated proteoglycan in the sera of patients using monoclonal and polyclonal antibodies. Cancer Res. 1984; 44:2577-2581
3. Baudy AR, Dogan T, Flores-Mercado JE, et al. FDG-PET is a good biomarker of both early response and acquired resistance in BRAFV600 mutant melanomas treated with vemurafenib and the MEK inhibitor GDC-0973. EJMMI Res. 2012; 2:22
4. Berditchevski F, Bazzoni G, Hemler ME. Specific association of CD63 with the VLA-3 and VLA-6 integrins. J. Biol. Chem. 1995; 270:17784-17790
5. Berditchevski F, Zutter MM, Hemler ME. Characterization of novel complexes on the cell surface between integrins and proteins with 4 transmembrane domains (TM4 proteins). Mol. Biol Cell. 1996; 7:193-207
7. Berditchevski F, Odintsova E, Sawada S, et al. Expression of the palmitoylation-deficient CD151 weakens the association of alpha 3 beta 1 integrin with the tetraspanin-enriched microdomains and affects integrin-dependent signaling. J. Biol. Chem. 2002; 277:36991-37000
8. Berditchevski F, Odintsova E. Tetraspanins as regulators of protein trafficking. Traffic. 2006; 8:89-96
9. Boman A, Svensson S, Boxer A, et al. Distinct lysosomal network protein profiles in parkinsonian syndrome cerebrospinal fluid. J. Parkinsons. Dis. 2016; 6:307-315
10. Bommarito A, Richiusa P, Carissimi E, et al. BRAFV600E mutation, TIMP-1 upregulation, and NF-κB activation: closing the loop on the papillary thyroid cancer trilogy. Endocr. Relat. Cancer. 2011; 18:669-685
11. Braeuer RR, Shoshan E, Kamiya T et al. The sweet and bitter sides of galectins in melanoma progression. Pigment Cell Melanoma Res. 2012; 25:592-601
12. Chapman PB, Hauschild A, Robert C, et al. Improved survival with vemurafenib in melanoma with BRAF V600E mutation. N. Engl. J. Med. 2011; 364:2507-2516
13. Chapman PB, Robert C, Larkin J, et al. Vemurafenib in patients with BRAFV600 mutation-positive metastatic melanoma: final overall survival results of the randomized BRIM-3 study. Ann. Oncol. 2017; 28:2581-2587
15. Charrin S, Jouannet S, Boucheix C, et al. Tetraspanins at a glance. J. Cell Sci. 2014; 127:3641-3648
16. Claas C, Stipp CS, Hemler ME. Evaluation of prototype transmembrane 4 superfamily protein complexes and their relation to lipid rafts. J. Biol. Chem. 2001; 276:7974-7984
17. Codina J, Li J, Dubose TD Jr. CD63 interacts with the carboxy terminus of the colonic H+-K+-ATPase to decrease [corrected] plasma membrane localization and 86Rb+ uptake. Am. J. Physiol. Cell Physiol. 2005; 288:1279-1286
18. Davies H, Bignell GR, Cox C, et al. Mutations of the BRAF gene in human cancer. Nature. 2002; 417:949-954
19. Duffield A, Kamsteeg EJ, Brown AN, et al. The tetraspanin CD63 enhances the internalization of the H,K-ATPase beta-subunit. Proc. Natl. Acad. Sci. USA. 2003; 100:15560-15565
20. Fedorenko IV, Abel EV, Koomen JM, et al. Fibronectin induction abrogates the BRAF inhibitor response of BRAF V600E/PTEN-null melanoma cells. Oncogene. 2016; 35:1225-1235
21. Flaherty KT, Lorusso PM, Demichele A, et al. Phase I, dose-escalation trial of the oral cyclin-dependent kinase 4/6 inhibitor PD 0332991, administered using a 21 -day schedule in patients with advanced cancer. Clin. Cancer Res. 2012; 18:568-576
23. Forte D, Salvestrini V, Corradi G, et al. The tissue inhibitor of metalloproteinases-1 (TIMP-1) promotes survival and migration of acute myeloid leukemia cells through CD63/PI3K/Akt/p21 signaling. Oncotarget. 2017; 8:2261-2274
24. Franco M, Muratori C, Corso S, et al. The tetraspanin CD151 is required for Met-dependent signaling and tumor cell growth. J. Biol. Chem. 2010; 285:38756-38764
25. Friedman AA, Amzallag A, Pruteanu-Malinici I, et al. Landscape of targeted anti-cancer drug synergies in melanoma identifies a novel BRAF-VEGFR/PDGFR combination treatment. PLoS ONE. 2015; 10: e0140310
26. Funakoshi T, Tachibana I, Hoshida Y, et al. Expression of tetraspanins in human lung cancer cells: frequent downregulation of CD9 and its contribution to cell motility in small cell lung cancer. Oncogene. 2003; 22:674-687
27. Girotti MR, Pedersen M, Sanchez-Laorden B, et al. Inhibiting EGF receptor or SRC family kinase signaling overcomes BRAF inhibitor resistance in melanoma. Cancer Discov. 2013; 3:158-167
28. Gustafson-Wagner E, Stipp CS. The CD9/CD81 tetraspanin complex and tetraspanin CD151 regulate α3β1 integrin-dependent tumor cell behaviors by overlapping but distinct mechanisms. PLos ONE. 2013; 8: e61834
30. Hammond C, Denzin LK, Pan M, et al. The tetraspan protein CD82 is a resident of MHC class II compartments where it associates with HLA-DR, -DM, and -DO molecules. J. Immunol. 1998; 161:3282-3291
31. Hauschild A, Grob JJ, Demidov LV, et al. Dabrafenib in BRAF-mutated metastatic melanoma: a multicentre, open-label, phase 3 randomised controlled trial. Lancet. 2012; 380:358-365
32. Hemler ME. Targeting of tetraspanin proteins--potential benefits and strategies. Nat. Rev. Drug Discov. 2008; 7:747-758
33. Hirabayashi J, Hashidate T, Arata Y, et al. Oligosaccharide specificity of galectins: a search by frontal affinity chromatography. Biochim.Biophys.Acta. 2002; 1572:232-254
34. Hirata E, Girotti MR, Viros A, et al. Intravital imaging reveals how BRAF inhibition generates drug-tolerant microenvironments with high integrin β1/FAK signaling. Cancer Cell. 2015; 27:574-588
35. Hong IK, Jeoung DI, Ha KS, et al. Tetraspanin CD151 stimulates adhesion-dependent activation of Ras, Rac, and Cdc42 by facilitating molecular association between β1 integrins and small GTPases. J. Biol. Chem. 2012; 287:32027-32039
36. Honma K, Iwao-Koizumi K, Takeshita F, et al. RPN2 gene confers docetaxel resistance in breast cancer. Nat. Med. 2008; 14:939-948
antigen associated with early stages of melanoma tumor progression. Cancer Res. 1988; 48:2955-2962
38. Israels SJ, McMillan-Ward EM, Easton J, et al. CD63 associates with the alphaIIb beta3 integrin-CD9 complex on the surface of activated platelets. Thromb. Haemost. 2001; 85:134-141
39. Ji Z, Erin Chen Y, Kumar R, et al. MITF modulates therapeutic resistance through EGFR signaling. J. Invest. Dermatol. 2016; 135:1863-1872
40. Jung KK, Liu XW, Chirco R, et al. Identification of CD63 as a tissue inhibitor of metalloproteinase-1 interacting cell surface protein. EMBO J. 2006; 25:3934-3942
41. Kazarov AR, Yang X, Stipp CS, et al. An extracellular site on tetraspanin CD151 determines alpha 3 and alpha 6 integrin-dependent cellular morphology. J. Cell Biol. 2002; 158:1299-1309
42. Khalili JS, Liu S, Rodríguez-Cruz TG, et al. Oncogenic BRAF(V600E) promotes stromal cell-mediated immunosuppression via induction of interleukin -1 in melanoma. Clin. Cancer Res. 2012; 18:5329-5340
43. Kimura A, Terao M, Kato A, et al. Upregulation of N-acetylglucosaminyltransferase-V by heparin-binding EGF-like growth factor induces keratinocyte proliferation and epidermal hyperplasia. Exp. Dermatol. 2012; 21:515-519
cells show different glycan profiles. J. Proteome Res. 2014; 13:1021-1033
45. Kobuch J, Cui H, Grünwald B, et al. TIMP-1 signaling via CD63 triggers granulopoiesis and neutrophilia in mice. Haematologica. 2015; 100:1005-1013
46. Lajoie P, Partridge EA, Guay G, et al. Plasma membrane domain organization regulates EGFR signaling in tumor cells. J. Cell. Biol. 2007; 179:341-356
47. Lau KS, Partridge EA, Grigorian A, et al. Complex N-glycan number and degree of branching cooperate to regulate cell proliferation and differentiation. Cell. 2007; 129:123-134
48. Liang Y, Eng WS, Colquhoun DR et al. Complex N-linked glycans serve as a determinant for exosome/microvesicle cargo recruitment. J. Biol. Chem. 2014; 289:32526-32537
49. Liu L, He B, Liu WM, et al. Tetraspanin CD151 promotes cell migration by regulating integrin trafficking. J. Biol. Chem. 2007; 282:31631-31642
50. Lizée G, Radvanyi LG, Overwijk WW, et al. Immunosuppression in melanoma immunotherapy: potential opportunities for intervention. Clin. Cancer Res. 2006; 12:2359-2365
51. Lupia A, Peppicelli S, Witort E, et al. CD63 tetraspanin is a negative driver of epithelial-to-mesenchymal transition in human melanoma cells. J. Invest. Dermatol. 2014; 134:2947-2956
53. Maecker HT, Todd SC, Levy S. The tetraspanin superfamily: molecular facilitators. FASEB J. 1997; 11:428-442
54. Manzano JL, Layos L, Bugés C, et al. Resistant mechanisms to BRAF inhibitors in melanoma. Ann.Transl.Med. 2016; 4:237
55. Marjon KD, Termini CM, Karlen KL, et al. Tetraspanin CD82 regulates bone marrow homing of acute myeloid leukemia by modulating the molecular organization of N-cadherin. Oncogene. 2016; 35:4132-4140
56. Montagut C, Sharma SV, Shioda T, Montagut et al. Elevated CRAF as a potential mechanism of acquired resistance to BRAF inhibition in melanoma. Cancer Res. 2008; 68:4853-4861
57. Mourad-Zeidan AA, Melnikova VO, Wang H, et al. Expression profiling of Galectin-3-depleted melanoma cells reveals its major role in melanoma cell plasticity and vasculogenic mimicry. Am. J. Pathol. 2008; 173:1839-1852
58. Müller J, Krijgsman O, Tsoi J, et al. Low MITF/AXL ratio predicts early resistance to multiple targeted drugs in melanoma. Nat. Commun. 2014; 5:5712
59. Nakagawa S, Arai Y, Mori H, et al. Small interfering RNA targeting CD81 ameliorated arthritis in rats. Biochem. Biophys. Res. Commun. 2009; 388:467-472
Clin. Cancer Res. 2013; 19:4868-4878
61. Nazarian R, Shi H, Wang Q, et al. Melanomas acquire resistance to B-RAF(V600E) inhibition by RTK or N-RAS upregulation. Nature. 2011; 468:973-977
62. Ohannesian DW, Lotan D, Thomas P, et al. Carcinoembryonic antigen and other glycoconjugates act as ligands for galectin-3 in human colon carcinoma cells. Cancer Res. 1995; 55:2191-2199
63. Paraiso KH, Xiang Y, Rebecca VW, et al. PTEN loss confers BRAF inhibitor resistance to melanoma cells through the suppression of BIM expression. Cancer Res. 2011; 71:2750-2760
64. Park SA, Kim MJ, Park SY, et al. TIMP-1 mediates TGF-β-dependent crosstalk between hepatic stellate and cancer cells via FAK signaling. Sci. Rep. 2015; 5:16492
65. Partridge EA, Le Roy C, Di Guglielmo GM, et al. Regulation of cytokine receptors by Golgi N-glycan processing and endocytosis. Science. 2004; 306:120-124
66. Ploper D, Taelman VF, Robert L, et al. MITF drives endolysosomal biogenesis and potentiates Wnt signaling in melanoma cells. Proc. Natl. Acad. Sci. USA. 2014; 112:420-429
67. Postow MA, Chesney J, Pavlick AC, et al. Nivolumab and ipilimumab versus ipilimumab in untreated melanoma. N. Engl. J. Med. 2015; 372:2006-2017
69. Prieto VG, Mourad-Zeidan AA, Melnikova V, et al. Galectin-3 expression is associated with tumor progression and pattern of sun exposure in melanoma. Clin. Cancer Res. 2006; 12:6709-6715
70. Qiu H, Duan WM, Shu J, et al. B3GNT2, a polylactosamine synthase, regulates glycosylation of EGFR in H7721 human hepatocellular carcinom a cells. Asian Pac. J. Cancer Prev. 2014; 15:10875-10878
71. Radford KJ, Thorne RF, Hersey P. CD63 associates with transmembrane 4 superfamily members, CD9 and CD81, and with beta 1 integrins in human melanoma. Biochem. Biophys. Res. Commun. 1996; 222:13-18
72. Radford KJ, Thome RF, Hersey F, et al. Regulation of tumor cell motility and migration by CD63 in a human melanoma cell line. J. Immunol. 1997; 158: 3353-3358.
73. Richard G, Dalle S, Monet MA, et al. ZEB1-mediated melanoma cell plasticity enhances resistance to MAPK inhibitors. EMBO Mol. Med. 2016; 8:1143-1161.
74. Rous BA, Reaves BJ, Ihrke G, et al. Role of adaptor complex AP-3 in targeting wild-type and mutated CD63 to lysosomes. Mol. Biol. Cell. 2002; 13:1071-1082
75. Sadej R, Romanska H, Kavanagh D, et al. Tetraspanin CD151 regulates transforming growth factor beta signaling: implication in tumor metastasis. Cancer Res. 2010; 70:6059-6070
77. Satow R, Nakamura T, Kato C; et al. ZIC5 drives melanoma aggressiveness by PDGFD-mediated activation of FAK and STAT3. Cancer Res. 2017; 77:366-377
78. Schröder J, Lüllmann-Rauch R, Himmerkus N, et al. Deficiency of the tetraspanin CD63 associated with kidney pathology but normal lysosomal function. Mol. Cell. Biol. 2009; 29:1083-1094
79. Sekulic A, Haluska P Jr, Miller AJ, et al. Malignant melanoma in the 21st century: the emerging molecular landscape. Mayo Clin. Proc. 2008; 83:825-846
80. Seubert B, Cui H, Simonavicius N, et al. Tetraspanin CD63 acts as a pro-metastatic factor via β-catenin stabilization. Int. J. Cancer. 2015; 136:2304-2315
81. Shen L, Yu M, Xu X, et al. Knockdown of β3GnT8 reverses 5-fluorouracil resistance in human colorectal cancer cells via inhibition the biosynthesis of polylactosamine -type N-glycans. Int. J. Oncol. 2014; 45:2560-2568
82. Silvie O, Charrin S, Billard M, et al. Cholesterol contributes to the organization of tetraspanin-enriched microdomains and to CD81-dependent infection by malaria sporozoites. J. Cell Sci. 2006; 119:1992-2002
83. Skubitz KM, Campbell KD, Iida J, et al. CD63 associates with tyrosine kinase activity and CD11/CD18, and transmits an activation signal in neutrophils. J. Immunol. 1996; 157:3617-3626
2008; 7:2876-2883
85. Straussman R, Morikawa T, Shee K, et al. Tumour micro-environment elicits innate resistance to RAF inhibitors through HGF secretion. Nature. 2012; 487:500-504
86. Suzuki M, Tachibana I, Takeda Y, et al. Tetraspanin CD9 negatively regulates lipopolysaccharide-induced macrophage activation and lung inflammation. J. Immunol. 2009; 182:6485-6493
87. Takawale A, Zhang P, Patel VB, et al. Tissue inhibitor of matrix metalloproteinase-1 promotes myocardial fibrosis by mediating CD63-integrin β1 interaction. Hypertension. 2017; 69:1092-1103
88. Takeda Y, He P, Tachibana I, et al. Double deficiency of tetraspanins CD9 and CD81 alters cell motility and protease production of macrophages and causes chronic obstructive pulmonary disease-like phenotype in mice. J. Biol. Chem. 2008; 283:26089-26097
89. Takino T, Miyamori H, Kawaguchi N, et al. Tetraspanin CD63 promotes targeting and lysosomal proteolysis of membrane-type 1 matrix metalloproteinase. Biochem. Biophys. Res. Commun. 2003; 304:160-166
90. Termini CM, Gillette JM. Tetraspanins function as regulators of cellular signaling. Front. Cell Dev. Biol. 2017; 5:34
166:7309-7318
92. Togayachi A, Kozono Y, Ishida H, et al. Polylactosamine on glycoproteins influences basal levels of lymphocyte and macrophage activation. Proc. Natl. Acad. Sci. USA. 2007; 104:15829-15834
93. Togayachi A, Kozono Y, Kuno A, et al. Beta3GnT2 (B3GNT2), a major polylactosamine synthase: analysis of B3GNT2-deficient mice. Methods Enzymol. 2010; 479:185-204
94. Tominaga N, Hagiwara K, Kosaka N, et al. RPN2-mediated glycosylation of tetraspanin CD63 regulates breast cancer cell malignancy. Mol. Cancer. 2014; 13:134
95. Toricelli M, Melo FH, Peres GB, et al. Timp1 interacts with beta-1 integrin and CD63 along melanoma genesis and confers anoikis resistance by activating PI3 -K signaling pathway independently of Akt phosphorylation. Mol. Cancer. 2013; 12:22
96. Trunzer K, Pavlick AC, Schuchter L, et al. Pharmacodynamic effects and mechanisms of resistance to vemurafenib in patients with metastatic melanoma. J. Clin. Oncol. 2013; 31:1767-1774
97. van Niel G, Charrin S, Simoes S, et al. The tetraspanin CD63 regulates ESCRT-independent and -dependent endosomal sorting during melanogenesis. Dev. Cell. 2011; 21:708-721
IGF-1R/PI3K. Cancer Cell. 2010; 18:683-695
99. Wang WC, Lee N, Aoki D, et al. The poly-N-acetyllactosamines attached to lysosomal membrane glycoproteins are increased by the prolonged association with the Golgi complex. J. Biol. Chem. 1991; 266:23185-23190
100. Wang J, Huang SK, Marzese DM, et al. Epigenetic changes of EGFR have an important role in BRAF inhibitor-resistant cutaneous melanomas. J. Invest. Darmatol. 2015; 135:532-541
101. Weilner S, Keider V, Winter M, et al. Vesicular Galectin-3 levels decrease with donor age and contribute to the reduced osteo-inductive potential of human plasma derived extracellular vesicles. Aging. 2016; 8:1156-1157
102. Wilson TR, Fridlyand J, Yan Y, et al. Widespread potential for growth-factor-driven resistance to anticancer kinase inhibitors. Nature. 2012; 487:505-509
103. Yadav V, Burke TF, Huber L, et al. The CDK4/6 inhibitor LY2835219 overcomes vemurafenib resistance resulting from MAPK reactivation and cyclin D1 upregulation. Mol. Cancer Ther. 2014; 13:2253-2263
104. Yang X, Kovalenko OV, Tang W, et al. Palmitoylation supports assembly and function of integrin-tetraspanin complexes. J. Cell Biol. 2004; 167:1231-1240
2017; 108:1223-1230
106. Yoshida T, Kawano Y, Sato K, et al. A CD63 mutant inhibits T-cell tropic human immunodeficiency virus type 1 entry by disrupting CXCR4 trafficking to the plasma membrane. Traffic. 2008; 9:540-558
107. Yoshida T, Ebina H, Koyanagi Y. N-linked glycan-dependent interaction of CD63 with CXCR4 at the Golgi apparatus induces downregulation of CXCR4. Microbiol. Immunol. 2009; 53:629-635