Pharma
VISION NEWS
No. 7
Introduction
長洲 毅志 1 (エーザイ株式会社)バイオイメージング:生命機能の統合的理解に向けて
清末 優子 2 (カン研究所)新規機能性蛍光プローブの論理的開発
浦野 泰照 9 (東京大学大学院薬学系研究科、JSTさきがけ)FRETプローブによるGタンパク質の活性化イメージング
中村岳史 ,青木一洋 18 (大阪大学・微生物病研究所・情報伝達分野)RNA蛍光イメージングによって拓かれる細胞核研究のフロンティア
谷 時雄 ,安東 知子 23 (熊本大学理学部理学科)オートファジー:細胞内の大規模分解系
水島 昇 30 (財団法人 東京都医学研究機構 東京都臨床医学総合研究所)薬学研究ビジョン部会からのお知らせ
35Index
( February 2006 )
Introduction
長洲 毅志 エーザイ株式会社
ゲノミクスに始まってトランスクリプトミ クスやプロテオミクス、メタボロミクスなど の名のもとに、網羅的な解析が行われるよう になってきております。ヒトをはじめとして 多くの生物の設計図は入手できましたし、精 度よく網羅的な発現解析が mRNA で出来る ようになりました。網羅的とまではいえませ んがタンパクレベルでも精度よく定量的な発 現解析が可能になりつつあります。かつて分 子生物学の実験を実際にやっていた頃を思い 出すと研究のやり方そのものが大きく変わっ てしまったと実感いたしますし、実際に皆様 も共感していただけるのではと思います。 では、この「オミクス」は創薬を変革した のでしょうか。残念ながらその答えは「否」 といわざるを得ません。もちろんまだ変えて いないという意味であって、方向が間違って いると言っているのではありません。 病気に関連する遺伝子が発見され、創薬の新 しいターゲット候補と書かれている記事は多 く見かけます。しかしながら、実際には遺伝 子発見から創薬への道のりは大変に長いもの なのです。 遺伝子発見までは簡単とは言わないまでも、 網羅的な手法が確立してきました。しかしな がら、その遺伝子が新しいものであればある ほどに、機能解析に非常に時間がかかります。 RNA 干渉という手段が開発され非常に有効 な技術となっています。しかしこれは遺伝子 の機能の発現を止める手段であってそのとき に生体や細胞に何が起こっているのかを突き 止めなければ機能解析にはなりません。 つまり、遺伝学的な言い方をすれば、遺伝 子に何らかの変化を与えたときに表現型がど うなるのかを調べることが必要なわけです。 個人的には、ゲノムから始まりプロテオーム やメタボロームと複雑さを増すオミクス分野 が広がっていますが、表現型の網羅的解析(フ ェノミクス)が確立されて始めて完結するの ではないかと思っております。 近年イメージング技術の発展は非常にすばら しいものがあり、遺伝子の機能解析に非常に 重要な技術として取り上げられるようになり ました。フェノミクスというような中身の不 確かな言葉が先行することは専門の先生方の お叱りを受けることになりますので、大上段 に振りかぶるつもりはありません。しかしな がらイメージング技術はしっかりと地に足の 着いた重要な技術であることは論を待たない と思います。そして将来的にはフェノミクス のような概念が認知されてくることを信じて おります。 そこで今回は、近年非常に重要な技術とな っているイメージングに焦点を絞って特集し、 5 名の先生方に寄稿していただきました。内 容は多岐にわたっており、私のような門外漢 が論評すべきではありませんが、総説から技 術開発や応用研究と、内容も濃くかつ幅広い 内容となりました。お忙しい中でご協力いた だきました先生方にはこの場を借りましてお 礼申し上げます。私は、創薬研究の中でもイ メージングが次の重要技術であることは確信 しておりまが、その気持ちを今回の特集で少 しでもご理解いただければと思っております。 ◆略 歴◆ 長洲 毅志 (Takeshi NAGASU):1979 年東大理・修士課程終了、エーザイ入社、1982 年ワシン トン大学留学、1999 年エーザイシーズ研所長、2003 年東大薬博士号取得、2005 年創薬研究本部副本部長バイオイメージング : 生命機能の統合的理解に向けて
清末 優子 株式会社カン研究所
はじめに 最近の動向として、生命を理解するために は生きたままの状態で解析することが求めら れるようになってきている。遺伝子やタンパ ク質の機能を調べるとき、これまでの生化学、 分子生物学の手法では、まず、試験管の中で の分子の特性を調べるが、生きた細胞や個体 の中で様々な因子と相互作用しあう環境でも 同様のことが起こっているのかどうか、それ がどのくらい生命機能に意味があるのかを in vivo で示さなければ本当の意味の生命現 象は理解できないという考え方が一般的にな ってきたからだ。 今や、生きたままの細胞あるいは個体の中 での分子の振る舞いの観察を可能にしたバイ オイメージングは、生命機能を理解する上で 必要不可欠な技術である。近年の急速なイメ ージング技術、特に蛍光タンパク質の発見・ 応用と検出機器の進歩は、生きた細胞・個体 での分子の生理機能を調べるための革新的な 発展をもたらした。本稿では、まず、in vivo における主要なイメージング法を紹介し、後 半では蛍光イメージングに焦点をあてて、蛍 光タンパク質を用いた分子イメージングにつ いて概説する。 1.in vivo イメージングGFP(green fluorescent protein)に代表 される、それ自身が蛍光を発することのでき るタンパク質の発見は、現在のバイオロジー における最も大きなブレイクスルーの一つで ある。遺伝子工学の手法を用いて人為的に蛍 光タンパク質を細胞や個体に発現させること で、生体の中で非侵襲的に分子を可視化する ことができる。しかし、多くの生き物は不透 明であり光を通さないので個体深部の蛍光シ グナルの検出は不可能であるため、蛍光タン パク質の用途は、培養細胞や個体・組織の表 面近傍、ゼブラフィッシュ等の透明な生物に 限られてしまう。個体のより深部のシグナル を検出するためには、化学発光を用いる方法 が普及しつつある。一方、元来は臨床検査用 の装置として開発された PET、SPECT や MRI がその応用範囲を広げられ、基礎科学的 研究や医薬品開発研究における重要なツール として脚光を浴びている。以下にそれぞれの イメージング方法の特徴を比較する。 ①蛍光イメージング 現在、蛍光イメージングに最も汎用されて いるのは蛍光タンパク質である。蛍光タンパ ク質をコードする遺伝子を操作することで、 興味ある遺伝子に融合して細胞に発現させた り個体の一部で発現させたりして自在に扱う ことができるため、遺伝子工学技術の浸透や 蛍光シグナル検出機器の進歩とあいまって、 応用範囲が急速に拡大している。詳細は後述 する。蛍光タンパク質以外では、細胞内オル ガネラや一部の組織を特異的に染色する試薬、 あるいはカルシウムなどに対する蛍光イオン 指示薬を用いることができる(表1)。 ◆略 歴◆ 清末 優子(Yuko Mimori-Kiyosue):カン研究所 主幹研究員 1992 年、御茶の水女子大学修士卒後、大阪大学基礎工学部。同時に松下電器産業・国際研究所に席をおき、極低 温電子顕微鏡による鞭毛の構造解析。1995 年、理学博士。 1997 年から、ERATO 月田・細胞軸プロジェクトにて、 イメージングを中心とした細胞内 の微小管の研究を始める。2000 年より現所属、カン研究所、主幹研究員。微 小管配向 の分子機構を探る研究を続けている。
核 DAPI, Hoechst
ミトコンドリア MitoTracker (Molecular Probes) リソソーム LysoTracker (Molecular Probes) ゴルジ体 BODIPY brefeldin A, BODIPY ceramide 細胞膜 DiI, DiO, DiA
エンドサイトーシス FM1-43 カルシウム指示薬 Fura2
血管 AngioSense680 (VisEn Medical) 腫瘍部位、炎症部位 ProSense(VisEn Medical) 骨代謝部位 OsteoSense(VisEn Medical) 表1 生体を標識する蛍光試薬の例 蛍光イメージングの大きなメリットは、目 的の分子を特異的に、非常に高い時間・空間 解像度で観察できることである。シグナル検 出技術の進歩により、現在、細胞や固体にお いてまで1分子レベルでのイメージングが可 能となっている。最大のデメリットは、不透 明な固体中では観察できないことであろう。 マウスのような透明性の低い検体の中で蛍光 を観察する場合、光が組織を通過するときの 散乱により励起・蛍光の光量減衰が生じるた め、深部のシグナルを検出することができな い。また、組織の表面近傍のシグナルを検出 しようとする際にも、可視域の短い波長で分 子を励起しようとすると、ヘモグロビンなど が光を吸収してしまい励起光が深部まで届き にくい上、ヘモグロビン自体をはじめとした 自家蛍光物質により背景光が高くコントラス トが低下する。このような問題を低減するた めに、生体内自家蛍光の少ない長波長の蛍光 (赤色~近赤外)を利用すれば、わずかであ れ光の浸透性が良くなることもあり、シグナ ルの改善が期待される。あるいは、光ファイ バーの先端に小さな対物レンズ取り付けて生 体に挿入し、直接に深部のシグナルを検出す るような顕微鏡が開発されつつある。 ②生物発光イメージング ルシフェリン-ルシフェラーゼ等の発光性 のプローブは、生化学では日常的に使用され ているが、蛍光よりもより生体深部に存在す るシグナルを検出することができるメリット が注目され、イメージングにおける有用性が 再認識されている。蛍の発光システムである ルシフェリンとルシフェラーゼは、ATP、マ グネシウムイオン、酸素の存在下で発光する。 ルシフェラーゼを人為的に細胞に発現させて ルシフェリンを投与すると、発光が生じ、そ こでルシフェラーゼが発現していることがわ かる。動物の体内からでの発光も、非常に感 度の高い検出器を用いることで体外から検出 することが可能である。例えば、ルシフェラ ーゼを発現させた癌細胞を動物に移植した後、 動物を生かしたまま経時的に癌細胞の増殖や 転移を体外のカメラで捉えることができる (http://www.xenogeny.com)。 デメリットは、感度と解像度が低いことで ある。発光した光は組織を通過する過程で著 しく散乱し、減衰する。従って、検出できる 像は非常にボケたものとなり、定量性も得に くい。そのような限界の中では、発光イメー ジングは、生体深部にある標的分子を特異的 に検出するためには有効な手段である。 ③PET(positron emission computed tomography:ポジトロン断層撮影)、SPECT
(single photon emission computed tomography:シングルフォトン断層撮影) PET、SPECT は、放射性同位体をプロー ブとして分子局在を測定する装置で、元来核 医学検査のために開発されたものである。短 半減期で生体に影響を与えず、かつ対外検出 可能な放射線を出す放射性同位元素で標識さ れた物質を生体に投与し、その生体内挙動を 検出することにより、生体内での生理学、生 化学、薬理学、分子生物学的現象を定量・画 像化する。 PET/SPECT の最大の特徴は感度が優れて いる点で、非常に少ない分子数で局在を見る ことができる。しかしデメリットは、分子プ ローブの設計が技術的に非常に高度であり、 いまのところ限られたプローブしか作製され ていないので、検出できる現象が限られてい ることである。薬剤の体内動態であれば、薬 剤を放射能標識することで直接測定すること ができるが、PET/SPECT で遺伝子発現をモ ニターするためには、レポーター遺伝子産物 に特異的親和性を有するPET/SPECT 用放射 性薬剤が必要である。これまでに報告された レポーター遺伝子と PET 薬剤の組み合わせ はまだ数種類しかない。今後、PET/SPECT 用のツールの開発が期待される。
④MRI(magnetic resonance imaging:核 磁気共鳴画像法) 1H-MRIは、水分子の水素原子の核磁化を捉 え画像化するもので、検体に何ら操作を加え ることなく、温度や流れ、水分含量や脂肪含 量といった物理化学的パラメータから画像コ ントラストを得る。空間分解能は数十平方ミ クロン程度であるが、測定対象を選ばず、全 くの非侵襲的に組織学的な状態を見ることが できる。13C、31P、23Naなどの多核種MRIを 用いれば、様々な生理学的パラメータを得る こともできる。 MRI では、検体のあるがままの状態での物 理量を測定しているため、分子イメージング には遠い。動物に何らかの操作を施した際の 影響を調べるためには、組織内の物質の物理 化学的パラメータを変化させる必要がある。 例えば生体内での水の透過性を調べる場合、 核磁化の緩和時間定数を変化させる緩和試薬 を投与し、緩和時間定数を強調する画像化を 行うことで、水透過性の高い部位を高信号領 域として検出する。MRI を使用するためには 生体への理解や高度なデータ解釈が必要であ るため、研究者が目的の測定対象に適した方 法を開発しようとすると、MRI への深い知識 を持ち、特定の生理機能検出を行うことので きる指示薬(緩和試薬)も作らなければなら ない。研究の幅広い用途に応用するためには 今後、多様な目的に応じたスタンダードな測 定法の確立が必要である。 2.蛍光タンパク質を用いたバイオイメージ ング GFP の登場は、間違いなく、近年の細胞生 物学に大きな転機をもたらした。それまで、 細胞抽出液の生化学的解析でしか調べること ができなかった細胞内の現象が、生きた細胞 の中で直接可視化することが可能になり、驚 くべき発見があいついだ。Tsien 博士(カリ フォルニア大)や宮脇博士(理化学研究所) のグループが中心となって GFP の改良や新 しい蛍光タンパク質の探索が続けられ、広い スペクトルにわたる多様な蛍光タンパク質が 開発されただけではなく、様々な細胞内シグ ナリングをモニターするためのプローブも作 製されてきた。本セクションでは、蛍光タン パク質の種類と用例を紹介した後、GFP 融合 タンパク質で微小管細胞骨格を可視化した実 例を紹介する。 ①蛍光タンパク質(fluorescent proteins, FPs)
GFP(green fluorescent protein)は 1962 年に下村博士によって、オワンクラゲの発光 器官から単離された。その 30 年後にcDNA が単離されて以後、改良が重ねられ、様々な モデル生物や細胞で用いることができる有用 なツールとなった(1,2)。現在までに、オワンク ラゲ以外の刺胞動物や節足動物からも蛍光タ ンパク質遺伝子が単離されて、青色から遠赤 色までのバリエーションがある(表2)。 らに、光刺激により活性化されて蛍光を 発 用途としては以下のような例がある。 1 2) P 3) P を 4) P 変異 5)分子の動態を見るマーカー:光刺激によ り蛍光を発するようになる GFP 変異体 表2 代表的な蛍光タンパク質(fluorescent proteins, FPs)
Color Protein (ref.) Source Excitation/Emission(nm)
Far-red mPlum (1) Dr. Tsien 590/649
Red mRFP1 (2) Dr. Tsien 584/607
mCherry(3) Dr. Tsien 587/610
Orange mKO MBL Intl. 548/559
Yellow-green Venus (4) Dr. Miyawaki 515/528
EYFP Clontech (TaKaRa) 514/527
Green EGFP Clontech (TaKaRa) 487/509
Cyan ECFP Clontech (TaKaRa) 433/475
Reference:(1) PNAS 2004; 101: 16745-16749, (2) PNAS 2002; 99: 7877-7882, (3)Nat. Biotechnol. 2004; 22: 1567-1572, (4) Nat. Biotechnol. 2002; 20: 87-90 さ す る よ う に な る タ ン パ ク 質 (photoactivated GFP, PA-GFP)や、色が変 わるもの(photoconvertible proteins)など も開発されて、一部のポピュレーションのみ をマークしてその動態を追跡することができ る。 主な )局在マーカー:既知あるいは未知の蛋白 に結合させて分子の局在を調べる。 細胞マーカー:特定の細胞にのみ GF を発現させその細胞を追跡する。 転写活性マーカー:寿命の短い GF 使って遺伝子発現を検出する。 pH レポーター:pH 感受性の GF 体を利用してpH の変化を調べる。
を使って分子動態(ターンオーバー)を 捉える。 分子間相互作用マーカー: FRET 現象 (後述)を 6) 利用して分子間の距離を調べ
②F fluorescence resonance energy transfer:蛍光共鳴エネルギー移動) もう る。 RET( FRET とは、2 つの蛍光分子が存在すると き、一方の蛍光分子の励起エネルギーが 一方の分子にトランスファーされる現象であ り、その結果エネルギーをトランスファーさ れた分子から蛍光が発せられる。蛍光を出す 方をドナー、受け取る方をアクセプターと呼 ぶ(図1)。 図1:FRET
CFP と YFP を用いた FRET プローブ。CFP がドナー、YFP がアクセプターで、CFP を励起(440 nm)したときに YFP するとエネルギーがYFP に移動し、YFP の蛍 光分子間の距離に依 存する。蛍光物質間の距離が 5nm のときに ka-u.ac.jp/biken/shuy ndex.htm ③微小管伸長ダイナミク 蛍光タンパク質の最もポピュラーな使用法 が近傍に位置 光(530 nm)が検出される。 FRET 効率は 2 つの蛍 FRET 効率がおよそ 50%で、それより距離が 離れるにつれで急激に減衰するため、ちょう ど、タンパク質分子がぴたりとくっつくよう な距離でしか起きない現象であることから、 タンパク質の相互作用や構造変化をモニタリ ングするために最適である。蛍光タンパク質 を用いる場合は例えば、GFP の遺伝子改変に よって作製されたシアン(青緑)色の CFP (Cyan Fluorescent Protein)と黄色の YFP (Yellow Fluorescent Protein)のペアを用い ると、FRET 現象を生じさせることができる。 FRETは、単にタンパク質間相互作用のモ ニタリングという目的だけではなく、シグナ ル伝達系の活性化を可視化する手段としての 有用性が脚光を浴びている。その先駆的なも のがFRETを利用したカルシウムセンサー “cameleon(カメレオン)”である(3)。Cameleon は、カルシウムの結合に依存して相互作用す るカルモジュリンとM13 ペプチドを連結し、 されにそれをGFPの青緑色変異体と黄色変 異体でサンドイッチした構造を持つタンパク 質である(図1)。その後、松田博士(大阪大 学微生物病研究所)のグループで、Rhoファ ミリーGタンパク質の活性化をモニターする プローブ、Raichu(Rasスーパーファミリー G蛋白の活性化モニター)やPicchu(チロシ ンリン酸化酵素の活性化モニター)などのシ リーズが次々と作製され、シグナル伝達が細 胞の状態に応じて様々な変化を見せることが 明らかにされている (松田研究室HP: http://www.biken.osa ouvirus/phogemon/i 及び中村先生の稿)。 スのイメージング は、タグとして興味あるタンパク質に融合し てその分子の局在を検出することである。筆 者らは、微小管細胞骨格の構成成分であるチ ュブリンや、微小管の先端に特異的に局在す る分子を GFP と融合したタンパク質を作製 し、微小管のダイナミクスを可視化してきた。 細胞運動や形態形成、分裂などの複雑な細 胞活動を担うため、細胞内に縦横に張り巡ら されているタンパク質繊維、細胞骨格が重要 である。細胞骨格のひとつ微小管は、チュブ リンが重合して形成される、プラス端とマイ ナス端を有する極性のある管状構造である。 細胞内では一般に、マイナス端が細胞の中心 付近にアンカーされ、プラス端が外側に向か って放射状に配置されている(図 2a)。微小 管の極性を利用した方向性のある物質輸送や オルガネラの正しい配置により細胞全体の極 性が形成されるため、微小管の配向は適切に 制御される必要がある。微小管はそれ自体が 主にそのプラス端においてダイナミックに伸 長と短縮を繰り返す性質を持ち、非対称な 様々な微小管ネットワークのパターンを形成 するためにはプラス端における微小管の安定 性が制御されている。
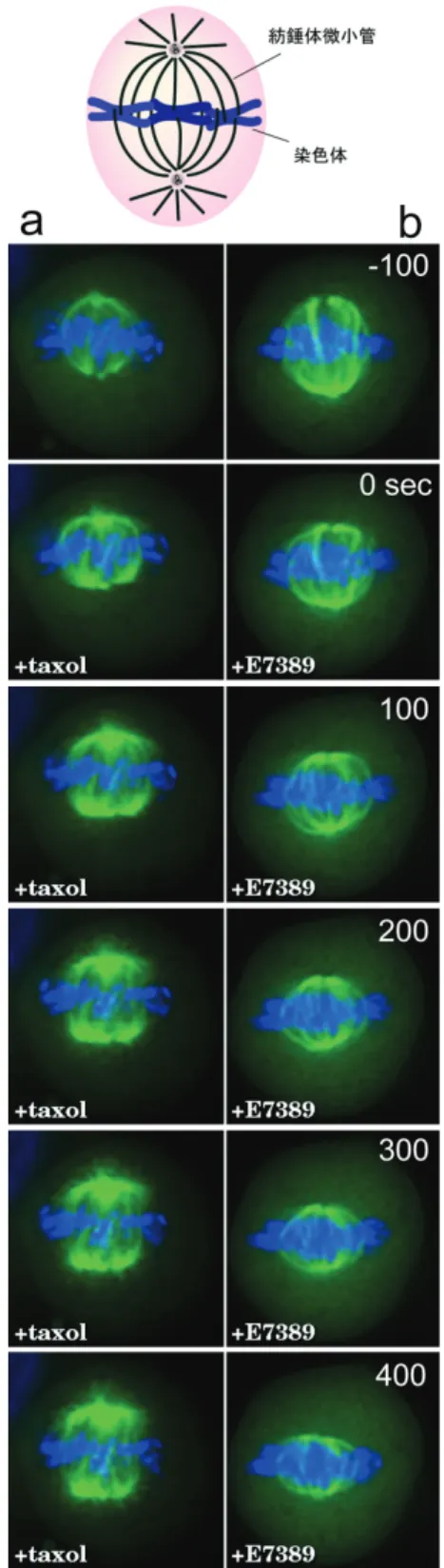
図 2:細胞内の微小管の配向と微小管の伸長端をマークす るEB1-GFP a)微小管は一般に、マイナス端が細胞の中心近傍に位置し、 プラス端を細胞周辺方向に向けて配置される。主にプラス 端において、伸長と短縮のダイナミクスを繰り返している。 b)GFP を融合した微小管結合タンパク質 EB1(EB1-GFP) の細胞内分布。EB1-GFP は、微小管全長にわたって弱く分 布するのと同時に、一部のプラス端にコメット状に集積し ている。c)EB1-GFP を発現する細胞をタイムラプス撮影 し、微小管先端を拡大して並べたもの。左上は時間(秒)。 筆者らはGFPを用いたイメージングによ り、微小管プラス端にチュブリンが重合して 新たに形成された先端に特異的に結合する分 子群が存在することを見出した。癌抑制因子 APC(adenomatous polyposis coli)遺伝子 産物や、その結合タンパク質EB1 である(4, 5)。 これらの分子以外にも微小管伸長端に結合す る分子が相次いで見つかり、現在、分子の動 態 の 特 徴 か ら“ 微 小 管 プ ラ ス 端 集 積 因 子 (+TIPs:プラスティップス)”と総称されて いる。現在までの研究で、+TIPsが微小管の 配向を制御するために重要な因子であること が明らかにされている(6)。 +TIPsの生理的役割はさておき、微小管の 伸長端をマークするという動的特性は、微小 管ダイナミクスをモニターするツールとして 優れている。EB1-GFPの動態は、筆者らのホ ー ム ペ ー ジ で 公 開 し て い る ( http://www.tsukita.jst.go.jp/kiyosue/eb1. html)。分子量が小さく扱いやすいこともあ り、現在、GFP融合EB1 (EB1-GFP)は、様々 な細胞、モデル生物において広く用いられて いる。EB1-GFPは微小管が伸長するフェーズ のみを検出するので、微小管の密度が高い部 位において微小管一本一本を分離して検出で きない場合にも、個々の先端を特定すること ができる。例えば、細い神経突起の中で微小 管がどちら向きに伸びていくのかを直接調べ る方法は無かったが、EB1 ホモログEB3 の GFP融合蛋白質を使って、神経細胞の樹状突 起や軸索における微小管伸長の方向をイメー ジングしたところ、樹状突起の細胞体側では およそ6:4の割合で細胞体方向に微小管が 伸長するのに対し、樹状突起の先端近傍と軸 索ではほとんど先端方向に伸長することが明 らかになった(7)。また、微小管制御因子を同 定する目的で、EB1-GFPを発現する線虫のト ランスジェニック株を樹立し、初期胚におい てEB1-GFPの動態を変化させる因子、即ち微 小管ダイナミクスを変化させる因子をRNAi 法によりスクリーニングすることで、微小管 の本数を制御する因子が同定されている(8)。 EB1-GFPは、微小管ダイナミクスを変化さ せる低分子量化合物の解析にも有用である。 抗癌剤タキソールは、微小管フィラメントを 安定化して伸長を促進し短縮を阻害すること で、結果的に微小管全体のターンオーバーを 抑制して伸長/短縮の両方のダイナミクスを 阻害する。EB1-GFPを発現する細胞にタキソ ールを添加すると、EB1-GFPコメットが数分 で消失するので、微小管の伸長が停止したこ とがわかる。現在臨床試験中の抗癌剤E7389 (微小管伸長阻害剤)は、重合していないチ ュブリンのプールを凝集させて重合できない 状態に不活性することで、微小管ダイナミク スを阻害すると考えられている(9, 10)。E7389 を細胞に添加してもEB1-GFPコメットは消 失し、微小管の伸長が停止したことが確認さ れた(図3)。
図3:E7389 による微小管伸長阻害 EB1-GFP を発現する細胞のタイムラプス撮影中に、抗癌剤 E7389 を添加した(3コマ目)。数分以内に EB1-GFP コメ ットが減少し、微小管の伸長が抑制されたことがわかる。 EB1-GFP のシグナルだけでは、微小管が 短縮していく過程は観察しにくいので、次に、 GFP を融合したチュブリンで微小管自体を 可視化した細胞で、細胞分裂中の紡錘体微小 管の長さの変化を追跡した(図4)。タキソー ルでは、チュブリンの重合が促進されて微小 管の本数、長さとも増加したが、E7389 では 微小管が次第に短縮して紡錘体全体が小さく なる様子が捉えられた。 おわりに 本稿では、実例として微小管ダイナミクス のイメージング・ツールを簡単に紹介したが、 色々な分子や細胞内構造を可視化することに より、多種多様な細胞現象を生きたままモニ ターすることができる。何を可視化しそこか らどのような情報を抽出するかはアイデア次 第であり、無尽蔵の可能性がある。イメージ ング技術は、これまでの生化学的・生物学的 解析とを並行して進め相補し合い、今後集積 していくであろう膨大なデータベース解析を 加えることにより、生命現象の統合的理解を 助け、基礎生命科学及び産業応用に貢献して いくことが期待される。 図4:E7389 により微小管が短縮する
GFP-α-tubulin を発現する細胞に taxol(a)あるいは E7389 (b)を添加した。青色が染色体、緑色が GFP-α-tubulin によって可視化された紡錘体微小管。Taxol を添加すると微 小管密度が増加するのに対し、E7389 を添加した細胞では 微小管が次第に短くなり、紡錘体が小さくなる。
◆参考文献◆
(1) Tsien RY.: Annu Rev Biochem. 67: 509-544, 1998 (2) Miyawaki A.: Neuron. 48: 189-199, 2005
(3) Miyawaki A, et al. : Proc Natl Acad Sci U S A. 1999 (4) Mimori-Kiyosue, Y. et al.: J Cell Biol., 148: 505-18, 2000 (5) Mimori-Kiyosue, Y. et al.: Curr Biol., 10 : 865-868, 2000
(6) 清末優子,月田承一郎 : わかる実験医学シリーズ『細胞骨格・運動がわかる』、三木裕明/編 : pp63-72, 羊土 社、2004
(7) Stepanova, T. et al. : J Neurosci. 23 : 2655-2664, 2003 (8) Srayko, M. et al. : Dev Cell 9 : 223-236, 2005. (9) Towle, MJ. et al. : Cancer Res. 61 : 1013-1021, 2001 (10) Jordan MA. et al. : Mol Cancer Ther. 4 : 1086-95, 2005
新規機能性蛍光プローブの論理的開発
浦野 泰照 東京大学大学院薬学系研究科、JST さきがけ
1.はじめに 生物は、各種刺激に対応して適切な応答を することで生命を維持している。ある外界か らの刺激に対して、どのような生理活性物質 が、どのタイミングで、どの場所で生成ある いは消去しているのかを明らかにすることは、 生命科学の本質に迫るものであり、現在も多 くの研究者が精力的に研究を行っている。こ の生物の本質に迫るためにはもちろん「武器」 が必要であり、HPLC、電気泳動、ELISA な ど、数多くの分析技術がこれまでに開発され、 生物研究領域に供給されてきた。特に近年は、 各種技術の進歩に伴って、細胞レベルでの詳 細な機能解析が盛んに行われている。しかし ながらこれらの分析技術は、細胞抽出液など を対象とするものであり、「生きている」状態 の生物試料を「生きたまま」観察するには不 向きであった。 このような中、「生きている」細胞を「生き たまま」観測する技術として、蛍光プローブ、 蛍光顕微鏡を用いた観察手法が開発された。 この手法は、蛍光顕微鏡下で生きている細胞 の挙動・応答を直接観察できるため、細胞内 外情報伝達系の解析など生物機能解明のため の最も強力な手法として、近年盛んに用いら れるようになってきている。さらにレーザー 共焦点蛍光顕微鏡をはじめとするハードウェ アの進化により、時空間分解能、感度などが ここ数年の間に激的に進歩したことも、本技 法の優位性をますます高めている。その原理 を図1左上に簡潔にまとめた。観測対象生理 活性分子(▽)の生成・消去を検知しようと した場合、ほとんどの生理活性物質は無色で あるため、光学顕微鏡でただ観察してもその 動きを知ることは出来ない。ここで▽と反 応・結合することで蛍光が現れる分子(蛍光 プローブ)を細胞内に存在させることで、▽ の動きを蛍光の変化として追うことが可能と なる。すなわち蛍光顕微観察において、ソフ トウェア的性質を持つ蛍光プローブは、ハー ドウェアである顕微鏡と同じ重みで重要な役 割を持っており、その種類の多さはそのまま 観測することが出来る分子の数となる。例え ばCa2+プローブFluo-3、Fura-2 無くしては、 Ca2+のセカンドメッセンジャーとしての役 割がここまで理解されることはなかったであ ろう。このような意味で有用な蛍光プローブ は生物現象の理解に決定的な役割を果たすが、 本当の意味で有用な「使えるプローブ」は実 は非常に少ない。これは、これまでの蛍光プ ローブはほぼすべて、Fluo-3 以来全く変化の ない単一の経験則に依存して開発されている ことに原因がある。つまり新たな蛍光プロー ブ の 開 発 と い う 観 点 か ら 見 る と 、Fluo-3, Fura-2 以来ブレークスルーと言える進歩は なく、普遍的な開発原理と呼べるものは存在 しない。よって自由自在なプローブ設計は不 可能であり、未だに生物領域の「かゆいとこ ろ」に手が届いていない。 ◆略 歴◆ 浦野 泰照(Yasuteru URANO、薬学博士):東京大学大学院薬学系研究科 助教授 経歴 1995 年東京大学大学院薬学系研究科博士課程修了、同年日本学術振興会特別研究員(PD)、97 年東京大学大 学院薬学系研究科助手、2004 年より科学技術振興機構さきがけ「構造機能と計測分析」領域研究員(併任)、2005 年より現職。平成16 年度日本薬学会奨励賞受賞 研究テーマ 生細胞の機能解析に有用な光機能性小分子の設計、開発とその応用筆者らはこのような現状を打破し、目的の 機能を有する蛍光プローブを論理的に精密に 設計することを目標とした研究を行ってきた。 そ の 結 果 、 光 誘 起 電 子 移 動(Photoinduced Electron Transfer; PeT)を設計原理とする蛍 光プローブの論理的なデザイン法の確立に成 功した。また実際このデザイン法に基づいて 種々のプローブの開発にも成功した。以下、 デザイン法の概要と開発に成功したプローブ を紹介する。 2.PeT による蛍光特性の制御 蛍光プローブとは、観測対象分子と「特異 的に」反応・結合することで、その蛍光特性 が大きく変化する機能性分子である。すなわ ち、「特異的な反応・結合」を「蛍光特性の変 化」に結びつけることができて初めて、論理 的な開発が可能となる。しかし残念ながら、 任意の化合物の蛍光特性を予想することは、 量子化学の進歩した現在でもほぼ不可能であ るため、これまでは主にtrial and error 方式 でプローブの開発は行われてきた。ではtrial and error を回避し、論理的に「観測対象分 子との反応、結合」を「蛍光特性の変化」に 結びつけるにはどうしたらよいのであろう か?その有効な方法として、蛍光を発する部 位とそれ以外の部位(標的分子と反応する部 位)とを分け、蛍光団の蛍光量子収率を蛍光 団以外の部分によりコントロールする方法が 挙げられる。具体的には、蛍光団の近傍に強 い電子供与体を配置すると、励起蛍光団から の蛍光発光よりも速くPeT が起こり、蛍光を 発しなくなる事象が物理化学領域ではよく知 られており、プローブ設計の基本原理として 有効であると考えられる。しかしこれまでの PeT 研究の対象のほとんどはアントラセンを はじめとする紫外域の蛍光団であり、有名な 蛍光の教科書には長波長励起蛍光団ではPeT による蛍光制御は不可能であるとの記述も存 在するほどであった。 3.フルオレセインを母核とする蛍光プロー ブの論理的なデザイン 紫外光領域で機能する蛍光プローブは、励 起光の照射による生細胞の障害が大きく、ま た自家蛍光も強く観測されてしまうため、生 物領域研究に用いることは難しく、実用性に 乏しいと言わざるを得ない。よって真に実用 ▼ ▽ ▽ ▽ ▽ ▽ ▽ ▼ ▼ ▼ ▼ ▼ ▼ ▽ ▽ ▽ ▽ ▽ ▽ 蛍光プ ロ ーブ ▼ ▽ ▽ ▽ ▽ ▽ ▽ ▼ ▼ ▼ ▼ ▼ ▼ ▽ ▽ ▽ ▽ ▽ ▽ 蛍光プ ロ ーブ 蛍光プ ロ ーブ と は、 ・ 観測対象分子と 「 特異的に」 反応・ 結合し ・ そ の前後で 蛍光特性が大き く 変化する 機能性分子で ある 。 ( 上図は細胞を 模式的に表し た も の。 ▽の分子のみを 特異的に検出する 。 ) 安息香酸部位 (電子供与部) キサンテン環部位 (蛍光団)
e
-無蛍 光 強 蛍光性 のPeT Donor電子密度と蛍光量子収率との関係 COOH -0.26460 COOH NH2 H2N -0.20498 COOH NH N N -0.26056 HOMO level (B3LYP, hartrees) 0.85 0.92 Φfl COOH NH2 -0.20422 0.015 COOH NHCOPh -0.23159 0.79 -0.19374 0.005 -0.22744 COOH 0.003 0.89 PeT Donor COOH O O HO R COOH O HO O COOH = R フルオレセインは分子内に蛍光団と電子供与部を併せ持つ分子であり、その蛍光特性は 安息香酸部位の電子密度に依存した光誘起電子移動(PeT)過程により制御可能である。 図1:蛍光プローブとはどのよう な機能性分子か?(左上) 代表的な長波長励起蛍光分子であるフルオレセインは2部位に分割して考えるこ とができ(左下)、その蛍光量子収率は光誘起電子移動により精密に予測するこ とが可能である(右)。的な蛍光プローブを開発するためには、500 nm 程度以上の長波長の可視光で機能するこ とが必須条件となる。筆者らは、このような 長波長励起蛍光団においても PeT による蛍 光のコントロールは可能と考え、フルオレセ インをターゲットとした物理化学的検討をま ず行った。フルオレセインは水系溶媒中で高 い蛍光量子収率を持ち、かつ生体に対する毒 性も少ないため、蛍光プローブの母核として 理想的な分子の1つである。実際、フルオレ セインを蛍光母核とするプローブは比較的多 く開発されてきたが、論理的にデザインされ た例はなく、そのほとんどはアミノフルオレ セインの蛍光量子収率が非常に低いことを利 用しているのが現状であった。 筆者らは種々のフルオレセイン誘導体を合 成しその蛍光特性を精査することで、一見リ ンカー構造の見あたらないフルオレセインで はあるが、実は分子をベンゼン環部位と蛍光 団であるキサンテン環部位の2部位に分けて 考えることが可能であり(図1左下)、分子内 PeTによりその蛍光特性を精密に制御可能で あることを見出した(1-3)。具体的には、ベンゼ ン環部位の電子密度がある値よりも大きいフ ルオレセイン誘導体はほぼ無蛍光であり、電 子密度がそれよりも低いとフルオレセインと 同等の強い蛍光を発することが明らかとなっ た(図1右)。本知見は、どのようなフルオレ セイン誘導体であっても、量子化学計算によ りベンゼン環部位の電子密度を求めることで、 その蛍光量子収率を正確に予測可能であるこ とを示す画期的なものである。さらに本知見 を最大限に活用することで、蛍光プローブの 論理的なデザインが初めて可能となった。す なわち、フルオレセインに標的分子と反応す る蛍光マスク部位を組み込み、この電子状態 が標的分子との特異的な反応・結合により変 化し、蛍光を発するようになるというプロー ブデザイン法を確立した(図2左上)。 言い換えるならば、ある検出対象分子に特異 Fluorophore
e
-Fluorophore Specific reaction ほぼ無蛍光 強い蛍光を発するPeT Donor No more
PeT Donor 蛍光プローブの論理的精密設計法 その1 Ex. max : Em. max : Φfl : 492 nm 515 nm 0.81 Ex. max : Em. max : Φfl : Ex. max : Em. max : Φfl : 492 nm 515 nm 0.81 492 nm 515 nm 0.81 Ex. max : Em. max : Φfl : 492 nm 517 nm 0.015 Ex. max : Em. max : Φfl : Ex. max : Em. max : Φfl : 492 nm 517 nm 0.015 492 nm 517 nm 0.015 1O 2 DMAX DMAX-EP O O HO COOH CH3 H3C O O HO COOH CH3 H3C O O 一重項酸素に特異的な化学反応 1O 2 O O R R R R 電子密度高 電子密度低 開発に成功した新規一重項酸素蛍光プローブ 図2:確立した蛍光プローブの論理的設計法その1(左上)。 開発に成功した一重項酸素蛍光プローブの概要(右)とその鍵反応(左下)。 的であり、かつ反応の前後で電子密度が変化 する化学反応が存在すれば、その反応部位自 身 は 全 く 蛍 光 を 持 た な く て も そ れ を 蛍 光 On/Offの変化に論理的に結びつけ、蛍光プロ ーブ化することが可能となった。本プローブ 設計法は、もちろんフルオレセイン以外の蛍 光団にも適用可能な汎用性の高い設計法であ る。実際ローダミン、BODIPYなどの蛍光特 性を精密に制御することに成功し、例えば BODIPY 骨 格 の NO 蛍 光 プ ロ ー ブ DAMBO-pH(4)などの開発にも成功している。 以下筆者らの設計法に基づく代表的な開発例 である、フルオレセイン骨格を持つ活性酸素 種検出蛍光プローブについていくつか紹介す る。
4.活性酸素種を種特異的に検出可能な蛍光 プローブの論理的開発 活性酸素種(ROS)は、炎症、ガンなど多 くの疾患に関わるとされ、また近年では細胞 内情報伝達物質としての役割も持つとの指摘 もあり、ますます注目を集めている。一口に ROSと言っても、スーパーオキシド、過酸化 水素、ハイドロキシルラジカル、一重項酸素 など多くの種が存在し、これらはそれぞれ特 徴的な化学反応性を持つことから、生体内に おいても異なる役割を持つ可能性も高い。 ROS検出用蛍光プローブは、筆者らの研究以 前にもいくつか開発され、中でもジクロロフ ルオレセインの2電子還元体であるDCFHが 広く用いられてきた。しかしながらDCFHに はROS間の特異性は全くなく、また励起光を 当てるだけでROSの有無にかかわらず大き く蛍光が増大してしまう欠点を持っており、 生物学的に意味あるデータを得ることは困難 であった。そこで筆者らは、上述の蛍光プロ ーブデザイン法を活用し、ある特定の活性酸 素種のみを検出可能な蛍光プローブの精密設 計を試みた。以下、一重項酸素(1O2)蛍光 プローブとOHラジカルなどの高い活性を持 つROSを特異的に検出可能な蛍光プローブ を紹介する。 1O2は、2重結合への特異的な付加反応 (ene反応)を引き起こすことがよく知られ ている特色あるROSの1つであり、その生体 内での発生、役割、影響に注目が集まってい る。このene反応の代表的な基質としてアン トラセンが挙げられるが、ene反応前後で大 きな電子密度の変化を伴うことが計算から予 測された(図2左下)。そこでこの化学反応を 上述のプローブデザイン法と組み合わせ、ジ フェニルアントラセンをベンゼン環部位とす る1O2蛍光プローブDPAX(5)、および図2右に 示したジメチルアントラセンをベンゼン環部 位とするDMAX(1)を設計、開発した。写真か らも明らかな通り、DMAXは1O2との反応前 はほぼ無蛍光であるが、1O2と反応すること で顕著な蛍光増大が観測され、デザイン通り 1O2蛍光プローブとして機能することが確か められた。ene反応は1O2特異的な反応であり、 またその反応性も高いため、DMAXを用いる ことで1O2を特異性高く、高感度に検出する ことが可能となった。もちろん、このような 長波長で機能する1O2蛍光プローブは今回 我々が開発したものが初めてである。 次に、OHラジカルなどの高い活性を持つ ROS(highly reactive oxygen species、以下 hROS)に対する蛍光プローブHPF, APFを紹 介する。前述した通り、ROSには非常に多く の種類があり、その反応性も大きく異なるは ずであるが、生物領域研究では現在でも「酸 化ストレス」として一括りに扱われることが ほとんどである。例えば、過酸化水素自身は ほとんど酸化力を持たないが、パーオキシナ イトライトは強い酸化力を持つ。よってもし Specific bond-scission Fluorophore
e
-ほぼ無蛍光 PeT Donor Fluorophore 強い蛍光を発する No more PeT Donor 蛍光プローブの論理的精密設計法 その2 高い活性を持つ酸化活性種(hROS)に特異的な化学反応 hROS ほぼ無蛍光 強い蛍光を発する COOH O O O HX COOH O HO O X=O; HPF X=NH; APF PeT Donor HPF APF DCFH •OH ONOO– –OCl NO O2– H2O2 各種ROSに対する反応性の比較 ROS 1O 2 6 8 6 5 9 2 <1 17 2 6 <1 <1 <1 ROO• Autoxidation 730 1200 7400 120 560 6600 3600 86 67 26 190 710 150 2000 強い活性を持つ 酸化活性種(hROS) APFを用いた、ブタ好中球の 次亜塩素酸産生イメ ージング <HPF、APFの特長> ・hROSとその他のROSとを 区別して検出可能 例:ONOO-とNOを区別可能 ・光自動酸化による蛍光上昇 がない→連続観測が可能 O O HO + HX bond-scission X p-位に水酸基、アミ ノ 基を有するジフェニルエーテル類は、OHラジカル、P450の酸化活性種 などの強い酸化活性を有するROSと選択的に反応し、エーテル結合の切断が起こる。 X=O, NH 図3:確立した蛍光プローブの論理的設計法その2(左上)。 観測対象である高い活性を持つ活性酸素種に特異的な化学反応(左下)と、 開発に成功した活性酸素種蛍光プローブの特長(右)。両者が生細胞内で等量産生したならば、細胞 に与える酸化ストレスは後者の方が圧倒的に 大きいはずである。そこで筆者らは、後者の ような強い酸化力を持つROS(hROS)のみ を検出可能な蛍光プローブの開発に着手した。 まずhROS特異的な化学反応として、図3左 下に示したジアリルエーテル類のipso置換反 応を採用した。この反応は、P450 の酸化活 性種やOHラジカルなどのhROSでは進行す るが、過酸化水素などその他のROSでは進行 しないことが、筆者らの研究(6)で明らかとな っており、プローブのhROS特異性を確保す るのに最適であると考えた。この化学反応は エーテル結合の開裂を伴うものであるため、 この特徴を生かすことが可能な第2のプロー ブ設計法をまず確立した(図3左上)。これは、 PeTによる蛍光消光に必要な電子密度の高い 部位をhROSとの反応により開裂する結合に より蛍光団に導入することで、hROSとの反 応前は非常に蛍光が弱く、かつhROS特異的 に電子密度の高い部位が蛍光団から切り離さ れることで蛍光が回復することを狙ったもの である。実際この設計法に則り、図3右に示 した2種の新規蛍光プローブHPF, APFの開 発に成功した(7)。HPF, APFにより検出可能な ROSを図3内の表にまとめたが、従来用いら れてきたDCFHがほぼ全てのROSと反応す る の に 対 し て 、 こ れ ら 2 種 の プ ロ ー ブ が hROSのみを特異的に検出可能である事がお 分かりいただけるかと思う。例えばHPFや APFを用いれば、NOとパーオキシナイトラ イトを区別して検出することが可能であり、 また両者の反応性が若干異なることを利用す れば次亜塩素酸の特異的検出も可能である。 さらにHPF, APFの最大の特長として、光依 存的な蛍光の増大が全く見られないことも挙 げられる。DCFHは可視光が当たるだけで顕 著な蛍光の増大が見られてしまうため、試料 調整には細心の注意が必要なばかりでなく、 同一視野の連続観測は不可能であるという大 きな欠点を有していた。今回開発に成功した HPF, APFはいずれも可視光照射では全く蛍 光が変化しないため、同一細胞内でのhROS 産生を系時的に、信頼性高く検出することが 可能となった。HPF, APFは国内では第一化 学薬品から、海外ではMolecular Probesなど から発売されており、既にこの特長を生かし た研究が論文として報告されるようになって きた。今後、HPF, APF類を用いることで、 酸化ストレス研究が大きく展開することが期 待される。 5.TokyoGreen 骨格の発見とこれに基づく セインの骨格構 新たな論理的設計法の確立 さらにごく最近、フルオレ 造を大胆に見直すことで、新たな蛍光プロー ブデザイン法に繋がる誘導体群の創製に成功 した。フルオレセインは 19 世紀から知られ る著名な蛍光性化合物にもかかわらず、積極 的な化学構造の変換が行われた例はほとんど 存在せず、例えばベンゼン環部位のカルボキ シル基はほぼ全てのフルオレセイン誘導体に おいて保存されていた。しかし上記のPeTの 考え方によれば、カルボキシル基は他の官能 基に変換することが可能なはずである。そこ でまず、カルボキシル基をメチル基、メトキ シ基へと変換することを試みた。その結果、 単にカルボキシル基を水素に置き換えると蛍 光は減弱する一方で、メチル基、メトキシ基 など他の官能基に置換した誘導体では、フル オレセインと同等の高い蛍光性を保つことが 明らかとなった(図4左)(8)。これはPeTの 観点からは納得のいく結果であり、またメチ ル基といった小さな置換基であってもベンゼ ン環部位と蛍光団の直交性を保つことが可能 であることが明らかとなった。驚いたことに これらの単純なフルオレセイン誘導体は新規 化合物であり、また以下に詳述するようにこ れらは極めて有用な蛍光プローブ母核となる ものであったため、我々はこれらの新規蛍光 骨格をTokyoGreen(以下TGと略す)と命名 した。ごく単純な2-Me TGですら新規化合物 であったという事実は、これまでいかにフル オレセイン分子に手が加えられてこなかった か端的に表すものである。TGsは設計自由度 の高い新規フルオレセイン分子群であると上 述したが、これは従来のフルオレセイン誘導 体は、レゾルシノールと無水フタル酸誘導体 の縮合により合成されていたため、ベンゼン 環部位の特定の位置に各種置換基を導入した 誘導体を収率良く合成することは困難であっ たのに対し、TGsはカルボキシル基を持たな いため、Grignard試薬等を用いた通常のC-C 結合生成反応による合成が可能であることに
由来する(図4右)。実際各種置換基をベンゼ ン環部位に導入した一連のTGsを高収率かつ 高選択的に合成することに成功した。次にこ れらTGsの蛍光特性を精査した結果、ベンゼ ン環電子密度の上昇により蛍光量子収率が減 少するというPeTの原理に一致した結果が得 られ、その蛍光On/Offの境界を初めて精確に 決定することに成功した(図5左下)。さらに そればかりでなく、蛍光On/Offの境界はキサ ンテン環部位の水酸基がアニオン型である場 合と、分子型である場合で大きく異なること も明らかとなった。図4左下の写真から明ら かなように、キサンテン環の水酸基がプロト ネートしている場合は、アニオン型である場 合に比べて蛍光On/Offの境界が電子密度の より低い側に移動する。この原因は、PeT過 O HO O COOH O HO O HO O O Me H Fluorescein, Φfl= 0.85 (既知化合物)
Hyd rox y p heny l fluorone, Φfl= 0.20
(既知化合物)
2-Me Tok yoGreen , Φfl= 0.84 (新規化合物) 図4:新たに開発した新規フルオレセイン系蛍光骨格TokyoGreen(左)。 TokyoGreen類は従来のフルオレセイン誘導体に比べ、1つのアイ ソマーを高収率で合成可能である(右)。 単一生成物、高収率 複数の生成物、低収率 <TokyoGreen類の合成> <通常のFluorescein誘導体の合成> O O TBDMSO OTBDMS Br R O HO O R tBuLi or Mg H+ + HO OH O R O O + Heat, H+ O HO O R COOH O HO O COOH R +
TG-βGal with GP293 / +lacZ Upper; TG-βGal GP293 / -lacZ Lower; FDG GP293 / +lacZ O O -O CH3 OCH3 O O CH3 OCH3 TG-βGal (almost non-fluorescent, Φfl = 0.002) 2-Me-4-OMe TG (strongly fluorescent, Φfl = 0.84) β-Galactosidase pH 7.4 O HO H H HO H H OH H O OH O O -O COO -O O βGal-O COO -O βGal-O O O O-βGal FDG
(non-fluorescent) (moderately fluorescent)FMG (strongly fluorescent)Fluorescein
cf. O -O O R Anion form O HO O R CH3 CH3 OCH3 CH3 OCH3 OCH3 OCH3 H3C H3CO H3C H3CO H3CO Electron density LOW HIGH COOH CH3 CH3 R' R' Neutral form 蛍光プローブの論理的精密設計法 その3 Fluorophore e -Fluorophore Specific reaction
PeT Donor PeT DonorNo more
-ほぼ無蛍光 強い蛍光を発する Fluorophore e -Fluorophore Specific reaction
PeT Donor PeT DonorNo more
-ほぼ無蛍光 強い蛍光を発する
図5:新規フルオレセイン誘導体TokyoGreensの構造と蛍光特性(左下)と、こ こ から確立した蛍光プローブの論理的設計法その3(左上)。 本設計法に基づき、高感度に生細胞内のβ-ガラクトシダーゼ活性を可視化可能な初めての蛍光プローブTG-βGalの開発に成功した(右)。
TG-βGal with GP293 / +lacZ Upper; TG-βGal GP293 / -lacZ Lower; FDG GP293 / +lacZ O O -O CH3 OCH3 O O CH3 OCH3 TG-βGal (almost non-fluorescent, Φfl = 0.002) 2-Me-4-OMe TG (strongly fluorescent, Φfl = 0.84) β-Galactosidase pH 7.4 O HO H H HO H H OH H O OH O O -O COO -O O βGal-O COO -O βGal-O O O O-βGal FDG
(non-fluorescent) (moderately fluorescent)FMG (strongly fluorescent)Fluorescein
cf. O -O O R Anion form O HO O R CH3 CH3 OCH3 CH3 OCH3 OCH3 OCH3 H3C H3CO H3C H3CO H3CO Electron density LOW HIGH COOH CH3 CH3 R' R' Neutral form 蛍光プローブの論理的精密設計法 その3 Fluorophore e -Fluorophore Specific reaction
PeT Donor PeT DonorNo more
-ほぼ無蛍光 強い蛍光を発する Fluorophore e -Fluorophore Specific reaction
PeT Donor PeT DonorNo more
-ほぼ無蛍光 強い蛍光を発する
図5:新規フルオレセイン誘導体TokyoGreensの構造と蛍光特性(左下)と、こ こ から確立した蛍光プローブの論理的設計法その3(左上)。 本設計法に基づき、高感度に生細胞内のβ-ガラクトシダーゼ活性を可視化可能な初めての蛍光プローブTG-βGalの開発に成功した(右)。
程の起こりやすさは電子ドナー側の電子密度 だけでなく、電子を受け取るアクセプター側 がどのくらい電子を受け取りやすいかにも大 きく依存することを考えれば、容易に理解で きる。すなわち例えばm-メトキシトルエンは、 プロトネート型のキサンテン環に対しては PeTドナーとして機能して蛍光消光を起こす 一方、より電子を受け取りにくいと考えられ るアニオン型のキサンテン環に対してはPeT による消光を起こすことが出来ないと言える (図5左下囲み内)。実際プロトネート型のキ サンテン部位は、アニオン型に比べて 0.4 V 以上還元されやすいことが酸化還元電位測定 により確かめられた。以上の知見、およびキ サンテン環のフェノール性水酸基のpKaは約 6.2 であり、中性水溶液中ではTGsはほぼア ニオン型として存在する事実を合わせること で、従来とは全く異なる発想に基づく新たな 蛍光プローブの論理的設計法が確立された (図5左上)。具体的には、2-Me-4-OMe TG のフェノール性水酸基が特定の保護基により 保護された化合物はPeTによりほぼ無蛍光で あるが、これが中性緩衝液中で各種加水分解 酵素によって脱保護反応が起こったならば、 強蛍光性であるアニオン型の 2-Me-4-OMe TGを生成するため、単段階加水分解反応によ り最大の蛍光増加を引き起こす蛍光プローブ として機能するというものである。 6.新規 β-ガラクトシダーゼ蛍光プローブ クトシ 7.新規アルカリフォスファターゼ蛍光プロ ガラクトシダーゼだけでな TG-βGal の開発と生細胞への応用 上記の設計法に基づき、まずβ-ガラ ダーゼ蛍光プローブTG-βGal(図5右上)を 開発した(8)。β-ガラクトシダーゼはレポータ ー酵素として最も汎用されているが、その検 出法はX-Gal染色などの吸光法を原理とする ものがほとんどであり、原理的に高感度であ るはずの蛍光法が用いられることは非常に少 なかった。これは、代表的な蛍光性プローブ であるFDG(図5右中段)が1分子内に2つ の糖部位を有し、強い蛍光を発するために2 段階の反応を必要とすること、また高い水溶 性のため細胞膜を透過せず、生細胞イメージ ングができないことが主な原因であった。 FDGを基質とした場合の2段階反応の初期 では、弱い蛍光性を示すFMGが生成し、さら に反応が進行して初めて強蛍光性のフルオレ セインを生成するため、β-ガラクトシダーゼ 量と蛍光強度上昇との間の線形性が乏しいば かりでなく、短い反応時間では低感度である という欠点を持つ。一方今回開発に成功した TG-βGalは分子内に糖部位を1つしか持た ず、これが加水分解される1段階の反応で、 FDGの2段階分に相当する最大の蛍光強度 変化を生じるように設計されているため、非 常に感度良くβ-ガラクトシダーゼ活性を検 出することが可能である。さらにTG-βGal は比較的脂溶性が高いため、細胞膜を容易に 透過するという最大の特長を有し、本プロー ブの開発により生細胞でのβ-ガラクトシダ ーゼ活性の可視化が初めて可能となった(図 5右下)。β-ガラクトシダーゼはクローン選 択マーカーとして汎用されており、これまで は細胞を固定してX-Gal染色を行う必要があ ったため、同じ 96 穴プレートを2枚用意す るなど繁雑な操作が必要であった。今回開発 に成功したTG-βGalを用いれば、β-ガラク トシダーゼ発現の有無を生きている状態の細 胞で検知可能であり、またX-Galに比べて高 感度検出も可能であることから、今後本プロ ーブを用いたクローニング操作が一般化する ものと予想される。(なおTG-βGalも第一化 学薬品から昨年末より市販されるようになっ た。) ーブ TG-Phos の開発とウェスタンブロッテ ィングへの適用 本設計法はβ-く、もちろん加水分解酵素全般に適用可能で ある。そこで次に、アルカリフォスファター ゼ(ALP)活性を高感度に検出可能な蛍光プロ ーブの開発を試みた。ALPは腫瘍マーカーと して用いられるなど臨床的に重要な酵素であ るが、基礎生物学研究においても、ウェスタ ンブロッティング(WB)やELISAの検出用酵 素として広く汎用される酵素である。ALP活 性 検 出 に は 、 古 く か ら 比 色 性 プ ロ ー ブ の BCIP-NBTが用いられてきたが、最近では化 学発光を原理とする検出試薬も汎用されるよ うになってきた。蛍光法を原理とするプロー ブとしてはFluorescein diphosphate (FDP) が良く知られているが、後述するように感度 面、ハンドリング面での問題から、ブロッテ
ィング膜上での検出にはほとんど用いられて いない。しかし蛍光法は原理的には十分高感 度であり、また同一WB膜上での多重染色に よる一斉分析も可能であることから、魅力あ る検出原理である。そこで筆者らは、既存の フルオレセインに比べてTGsの脂溶性が上昇 していることに着目し、WB膜上での高感度 ALP活性検出が可能なプローブとして、図6 上に示したTG-Phos(9)を設計、合成し、光学 特性及びプローブとしての特性を精査した。 その結果、開発したTG-Phos自身はほぼ無蛍 光であり、またpH 7.4 緩衝液中ALP非存在下 では化学的に安定であり、かつここにALPを 添加することで顕著な蛍光増大が観測された ことから、ALPプローブとして機能すること が示された。さらに従来最も高感度なALPプ ローブとされてきたFDPよりも、高感度かつ 高い線形性でALP活性を検出できることも 明らかとなり、in vitro検出系として極めて優 秀なプローブであることが明らかとなった。 次にWBへの適用を図るべく、ALPによる TG-Phos 由 来 の 蛍 光 性 生 成 物 で あ る 2-Me-4-OMe TGのWB膜への吸着性を検討 した。その結果、FDPからの生成物であるフ ルオレセインは水溶性が高く拡散してしまう ために検出が難しかったのに対し、TG誘導体 は適度な脂溶性を有するためにブロット膜に 吸着され、目的のバンドの検出が可能である ことが明らかとなった。そこで実際のWB操 作を行い、CYP3A2 をALP修飾2次抗体と TG-Phosを用いて検出したところ、数ngレベ ルの高感度検出が可能であることが示された。 TG-Phosの検出感度は、WB法での蛍光検出 ALP プ ローブ として 現在 汎用さ れて いる ELF97-phosphate、DDAO-phosphateよりも 高感度であることも明らかとなった。さらに TG-PhosとBODIPY TR-Xを用い、CYP3A2 と全タンパクの同時染色を試みたところ、各 バンドを鮮明かつ高感度に得ることに成功し、 蛍光法の利点である一斉分析が十分可能であ ることも示された(図6下)。 Protein of interest
Western blot membrane Alkaline Phosphatase Me OMe O O O P -OO O-Me OMe O O -O Protein of interest
Western blot membrane Alkaline Phosphatase Me OMe O O O P -OO O-Me OMe O O -O O Me O P O O OMe -O O- O Me -O O OMe ALP pH 7.4 TG-Phos (Φfl=0.011, almost non-fluorescent) 2Me-4-MeO TG (Φfl=0.84, strongly fluorescent) 図6:蛍光プローブの論理的設計法その3に基づいて開発された新規 アルカリフォスファタ ーゼ蛍光プローブ(上)と、こ のウエスタ ンブロット 法への適用例(下、全タンパクとの同時染色例)。 8.おわりに 以上述べてきたように筆者らは、PeTを基 本原理とした蛍光プローブの論理的なデザイ ン法を確立し、この原理に基づくことで全く 新たな機能を有する蛍光プローブの開発に成 功した。これらのデザイン法はフルオレセイ ン以外の蛍光団に対しても適用可能であり、 実際数多くの蛍光プローブの開発に既に成功 している。本稿で紹介したプローブの検出対 象のバラエティの豊かさ、およびその実用性 を見ていただければ、確立した設計法が極め て有用であることをお分かりいただけたかと 思う。(10-13)その他開発に成功したプローブ の例は、研究室HP: http://www.f.u-tokyo.ac.jp/~tlong を ご 参 照いただければ幸いである。 9.謝辞 本研究は、東京大学大学院薬学系研究科薬 品代謝化学教室にて行われたものである。長 野哲雄教授及び実験の苦楽を共にしてくれた 歴代の学生さん達に、この場を借りて深謝し たい。
◆参考文献◆
(1) Kumi Tanaka et al., J. Am. Chem. Soc., 123, 2530-2536 (2001). (2) Tetsuo Miura et al., J. Am. Chem. Soc., 125, 8666-8671 (2003). (3) Tasuku Ueno et al., J. Am. Chem. Soc., 126, 14079-14085 (2004). (4) Yu Gabe et al., J. Am. Chem. Soc., 126, 3357- 3367 (2004).
(5) Naoki Umezawa et al., Angew. Chem. Int. Ed., 38, 2899-2901 (1999). (6) Yasuteru Urano et al., J. Am. Chem. Soc., 119, 12008-12009 (1997) (7) Ken-ichi Setsukinai, et al., J. Biol. Chem., 278, 3170-3175 (2003). (8) Yasuteru Urano et al., J. Am. Chem. Soc., 127, 4888-4894 (2005). (9) Mako Kamiya et al., Angew. Chem. Int. Ed., 44, 5439-5441 (2005). (10) 浦野泰照 他, 現代化学, 381, 44-50 (2002).
(11) 浦野泰照 他, ぶんせき, 426-432 (2003).
(12) 浦野泰照 他, アレルギー・免疫, 12, 745-751 (2005).