博士学位論文
環境微生物を用いた微生物農薬に関する研究
近畿大学大学院
生物理工学研究科 生物工学専攻
大池 達矢
1
目次
第 1 章 緒論 ... 5 第2章 放線菌を用いた微生物農薬の研究 ... 10 2-1 序論 ... 10 2-2 材料および方法 ... 13 2-2-1 微生物 ... 13 2-2-2 放線菌を用いた植物病原菌に対する対峙培養試験 ... 13 2-2-3 放線菌の同定 ... 14 2-2-4 放線菌 KT 由来抗真菌活性物質の抽出および精製 ... 14 2-2-5 放線菌 AR10 由来抗真菌活性物質の抽出および精製 ... 15 2-2-6 KT 由来抗真菌活性物質の LC-MS 分析 ... 16 2-2-7 AR10 由来抗真菌活性物質の LC-MS 分析 ... 16 2-2-8 抗真菌活性物質の NMR 分析 ... 17 2-2-9 キュウリを用いた KT による植物病原菌防除試験 ... 17 2-2-9-1 R. solaniに対する感染防御試験 ... 17 2-2-9-2 B. oryzaeに対する感染防除試験 ... 18 2-2-10 キュウリを用いた AR10 による植物病原菌防除試験 ... 18 2-3 結果 ... 20 2-3-1 放線菌の単離および選抜 ... 202 2-3-2 16S rDNA 系統解析による放線菌の同定 ... 22 2-3-3 KT の生産する化合物の精製 ... 25 2-3-4 Streptomyces sp. KT の生産する化合物の LC-MS 分析 ... 27 2-3-5 AR10 の生産する化合物の精製 AR10 ... 31 2-3-6 AR10 の生産する化合物の LC-MS 分析 ... 34 2-3-7 精製した抗真菌活性物質のNMR分析 ... 36 2-3-8 KT を用いたR. solani の感染防除試験 ... 44 2-3-9 KT を用いたB. oryzae の感染防除試験 ... 50 2-4 考察 ... 52 2-5 結論 ... 56 第3章 未利用植物残渣を用いたBacillus subtilis RB14 の固体培養 ... 57 3-1 序論 ... 57 3-2 材料および方法 ... 60 3-2-1 微生物 ... 60 3-2-2 残渣の調整 ... 60 3-2-3 微生物の培養 ... 60 3-2-4 残渣懸濁液体培地を用いた RB14 の培養 ... 61 3-2-5 固体発酵培養における RB14 の培養 ... 61 3-2-6 HPLC を用いた iturin A 分析 ... 61 3-2-7 抗菌活性物質の熱耐性 ... 62
3 3-2-8 RB14 培養物を用いた植物病原菌抑制試験 ... 62 3-2-9 植物病原菌感染防除試験 ... 63 3-2-10 RB14 固体発酵培養物がキュウリの初期成育に与える影響 ... 63 3-3 結果および考察 ... 64 3-3-1 残渣液体培地を用いた RB14 の培養 ... 64 3-3-2 固体発酵培養における RB14 の培養 ... 66 3-3-3 固体発酵培養物添加培地による各種植物病原菌の生育抑制試験 ... 68 3-3-4 固体発酵培養物によるR. solaniの感染防除試験 ... 70 3-3-5 RB14 固体発酵培養物がキュウリの初期成育に与える影響 ... 72 3-4 結論 ... 74 第 4 章 低栄養細菌を用いた微生物農薬の研究 ... 75 4-1 序論 ... 75 4-2 材料および方法 ... 77 4-2-1 微生物 ... 77 4-2-2 微生物の培養および抗真菌活性試験 ... 77 4-2-3 抗真菌活性物質の抽出および精製 ... 78 4-2-4 精製した化合物の LC-MS 分析 ... 79 4-2-5 KS 由来抗細菌活性物の GC-MS 分析 ... 79 4-2-6 低栄養細菌の同定 ... 80 4-3 結果および考察 ... 81
4 4-3-1 低栄養細菌の選抜と抗真菌活性試験 ... 81 4-3-2 抗真菌活性物質の精製と分析 ... 84 4-3-3 抗菌活性物質の LC-MS 分析 ... 86 4-3-4 KS 由来抗細菌活性物質の GC-MS 分析 ... 88 4-3-5 KS および DE の同定 ... 91 4-4 結論 ... 95 第 5 章 総括 ... 96 参考文献 ... 99 原著論文 ... 112 謝辞 ... 113
5
第 1 章 緒論
国連によると、現在の世界人口は 75 億人を超え、2050 年には 97 億人にまで増加し、 2100 年には 111 億人に達すると推測されている(FAO, 2017)。特に、アフリカ地域の 26 か国の人口増加は著しく、その人口は 2050 年まで約 2 倍、2100 年には 3.5 倍の 44 億人に なると考えられている。また、この地域は貧困諸国が集中している地域であるため、人口増 加は様々な問題を悪化させる可能性がある。また、土壌の低肥沃度や乾燥などの環境に関す る問題なども抱えているため、作物を安定的に栽培する技術を必要とするが、そのような技 術はまだ確立されていないのが現状である。これらのことから、世界の人口増加を支える食 料生産は危急の課題となっている。棟居らによると、120 億人の世界人口レベルにおける農地必要量は 26 億 ha 以上であると推定されている(Munesue and Masui, 2009)。2000 年に
おける世界の耕作地の合計は 15 億 ha と試算されており、これに加えてさらに 11 億 ha 以 上の耕作地の確保が必要となる。また同時に、農作物生産に適した土地の面積は、世界全体 で必要量を十分に上回るだけ存在するが、森林保全や十分な灌漑の必要性などを考慮する と大きく制限を受けることが考えられ、限られた土地をどのように利用し、如何に作物栽培 における技術の改良を行うかが非常に重要な課題となる。また、FAO によると現状の食料 生産は、世界全体でみると約 25 億 t の穀物が年間に生産されており、ひとりあたりの食料 としては十分な量が生産されていると考えられているが、その一方で作物栽培における自 然災害や病害虫の影響、輸送における損失は非常に高いものとなっている。実際に、アメリ カにおいては、農薬などによる管理措置が実施されているにもかかわらず、毎年、農作物の 被害額は、病害によって 91 億ドル、昆虫によって 77 億ドル、雑草によって 62 億ドルが失
6 われていると推定されている(Agrios, 2005)。また、世界全体の作物生産量における生物的 被害による損失割合は、植物病原菌によるものが 14%、昆虫由来が 10%、雑草による損失 が 12%となっており、作物生産の 3 割程度が損失してしまうことが問題となっている (Agrios, 2005)。このような植物病害や害虫問題を解決する手段のひとつとして、一般的に は農薬が使用されている。 近年の農薬は、低使用量、人畜および作物に対する安全性の向上、環境負荷の低減が要求 されており(Kozo, 2013)、世界中で様々な対策が行われているが、農薬の使用には現状で
はまだまだ多くの問題を抱えている。OECD(Organisation for Economic Co-operation and
Development:経済協力開発機構)のデータをまとめた報告によると、問題点として、農薬 の継続的な使用は病害虫や雑草などの抵抗性の発現や農薬散布者や周辺地域住民の健康問 題、飲料水や地下水などへの農薬汚染が挙げられている(Kozo, 2013)。また同時に、農薬 に対するリスク評価を行うにあたり、対象とする生物が農薬に暴露する可能性のある濃度 を正確に知ることは難しいとも報告している。このような背景から、環境に対する負荷を低 減するため、従来の化学農薬のみに依存した病害防除から、耕種的、生物的、物理的および 化学的な防除方法を合理的に組合せた総合的病害虫および雑草管理( Integrated Pest Management: IPM)の確立が求められている。近年、これらの技術のひとつとして病害虫 に対して防除効果を有する微生物を利用した微生物農薬の研究が注目されている。
微生物農薬(biological control agent, biocontrol agent)に利用される微生物は、植物根圏
に定着し、植物の生育に対して様々な有益な効果をもたらす。抗菌活性物質や酵素の生産な
7
除の研究、植物生長促進効果をもつ微生物(plant growth promoting rhizobacteria: PGPR,
plant growth promoting fumgi: PGPF)を用いた作物栽培など、近年になってそれらの研究
が急速に進んできた。特に、微生物農薬の候補株としてよく用いられているのは、細菌では
Bacillus属、Pseudomonas属の2種、真菌ではTrichoderma属であり、これらの微生物の
研究は他の候補株と比較すると多くなっている(Hyakumachi et al., 2014)。Bacillus属細菌
は、環境中に普遍的に存在しており、生物農薬資材として数多く菌株が研究されてきた。特
に日本では、古くから伝統的な食文化として納豆を食することから、納豆菌(Bacillus
subtillis var. natto)の研究は多く、またその農業への利用は個人レベルでも行われてきた。
実際に、市販化された日本の微生物農薬登録数としては本属を用いたものが最も多くなっ
ている。本菌株の特徴は、芽胞を形成し、様々な環境条件下でも生存することが可能な点で
あり、製剤化の際、菌の処理が比較的容易であることが挙げられる。また、本菌株は抗真菌
活性物質である iturin, fengycin, plipastatin, surfactin(Huan et al., 2015)などの界面活性
作用をもつ環状リポペプチド化合物を生産し、宿主植物への病原真菌の感染防除や、病害抵
抗性誘導などの役割も担っていると報告されている(Ongena et al., 2007)。Pseudomonas属
細菌も同様に、幅広い環境中に普遍的に存在している。本属は、一部に植物病原性を有する
種も含むが、市販化された製剤数としてはBacillus属細菌に次ぐ。特に近年、Pseudomonas
fluorescensから派生し、新しく分類されたPseudomonas protegensは植物病原菌から植物
体を保護(protect)するとして命名された。本菌株も強い抗真菌活性物質の生産が多数報告
されており、感染防除・病害抵抗性誘導・植物成長促進などの研究がよく行われている
8
す る カ ビ と し て 広 く 研 究 が 行 わ れ て い る 。Trichoderma 属 は 、 木 材 腐 朽 菌 で あ る
Trichoderma reeseiにおけるセルラーゼの研究(Gusakov, 2011)やTrichoderma viride、
Trichoderma harzianum、Trichoderma atrovirideなどから生理活性物質として 100 種を超
える化合物の単離が報告されている(Mukherjee et al., 2012)。また、キノコ類に関しては
病原菌となり得るが、その幅広い物質生産能力により、植物病害防除の研究も行われている
(Freeman, et al., 2004, Howell, 2003)。日本では、Trichoderma asperellum(T. atroviride)
を用いた菌寄生型の微生物農薬として、製造・販売されている(Watanabe et al., 2007,
Yoshioka et al., 2012)。近年、微生物農薬の研究が増えつつある微生物として放線菌が挙げ
られる。放線菌は、グラム陽性の細菌で、菌糸や胞子を形成するという特徴を持っている。
また、放線菌は古くから抗菌活性物質の研究が進んでおり、1940 年代にはアクチノマイシ
ンやストレプトマイシンといった抗生物質が発見された(Waksman et al., 1958, Wainwright,
1991)。その後、数多くの生理活性物質が放線菌から単離され、医学分野の発展に貢献し、 抗生物質の実用化は 20 世紀における大きな発明のひとつとされている。農業においても、 放線菌は重要な役割を担っていると考えられており、環境中では腐葉土の分解において主 要な微生物のひとつとされ、堆肥作製時においても重要な菌であると考えられている。また、 放線菌の多くがキチナーゼを生産すると報告されており、そのため植物病原菌や昆虫など の細胞壁を分解することで、植物病原菌やセンチュウのなどといった病害の抑制に繋がる
と考えられる(Singh and Gaur, 2016)。このような利点があるにも関わらず、放線菌を用い
た微生物農薬に関する研究は、現在のところ他の微生物に比べるとまだ少ないのが現状で
9 これらの微生物の他にも多くの微生物が病害虫防除に有効であるとされ、研究がおこな われており、微生物農薬の研究は今後さらに拡大し、普及していくと考えられる。微生物農 薬の開発は、「食の安全」「食の安心」が重要な課題である中、消費者のみならず、農薬汚染 や病害虫の抵抗性発現など、環境への負荷軽減に繋がると考えられる。このような背景から、 本研究では、微生物農薬の開発を目指し、土壌などの自然環境中から単離したいくつかの微 生物について微生物農薬の候補株として植物病原菌に対する防除効果を試験した。
10
第2章 放線菌を用いた微生物農薬の研究
2-1 序論 第1章でも述べた通り、植物病原菌は作物へ感染し病害を引き起こす微生物のことを示し、 農作物の収穫に多大な被害を与えている。特に、土壌伝染性糸状菌である Rhizoctonia solani は土壌伝染性の植物病原菌として報告されており、立枯病や紋枯病など世界中において 様々な作物へ被害を与える植物病原菌として知られている(Summerell et al, 2011)。R.solaniは、イネ (Jayaprakashvel et al., 2010)、エンドウ (AI-Askar and Rashad, 2010)、ダイ
ズ(Oliveira et al., 2013)、ジャガイモ(Jayaprakashvel et al., 2010)、トマト(Bartz et al., 2013)、
キュウリ(Huang et al., 2011)、ピーナッツ(Summerell et al., 2011)など様々な作物に対し
て感染し、病害を引き起こす。実際に、イネの場合、この病原菌による紋枯病による収量の
損失は、アジアでは 5-10%と推定され、さらに、R. solaniの生育に適した栽培条件下で感
受性の品種が栽培された場合は、収量に対する損失が 50%まで増加したと報告されている
(Lee and Rush, 1983, Marchetti and Bollich, 1991, Savary et al., 2000)。ジャガイモにおい
ては、枝枯病によって 30%の収量損失が報告されている(Banville, 1989)。キュウリの場 合、米国の年間平均損失は約 7-9%であり、これは約 4-5 百万ドルの損失に相当する(Lewis and Papavizas, 1980)。これらのことから本病原菌を防除することは農業生産性を向上させ るための重要な課題となっている。 近年の農業は、植物病原菌から植物を保護するための方法として、化学農薬が広く使用さ れている(Deepa et al., 2011)。しかし、農薬の無差別な使用は目標病害以外にも影響を与え、
11 おける化学農薬および化学肥料の過剰な使用は、昆虫や植物病原菌における薬剤耐性や飲 料水や地下水などへの農薬汚染などへと繋がったと報告されている (Mizumoto et al., 2006)。さらに、農薬が検出された割合や基準値を超えた割合のいずれも極めて低いという 結果にも関わらず、「食」の安全や「食」の安心などといった考え方に対する間違ったリス クコミュニケーションにより、化学農薬に対する信頼性は低下している(梅津, 2011)。こ の問題に対処するために最近では生物農薬と有機質肥料が、化学農薬と化学肥料の代替も
しくは併用の選択肢としてそれぞれ注目されている(Segarra et al., 2008, Mizumoto et al.,
2006)。特に微生物を使用した植物病原菌の生物的防除は、化学農薬に換わる実現可能な選 択肢として考えられており、その毒性についても動物や人間に対して比較的低いとされて いる(Zucchi et al., 2014)。生物農薬は防除対象の生物以外には影響が少なく選択性が高いと されている。また、自然界に存在する生物を病害虫防除に利用したものであるため残留毒性 の懸念がなく、環境に対する負荷を軽減しながら防除効率を維持して農業生産に貢献する ことが可能となる。しかし、現在までの研究では生物農薬を使って全ての病害虫の防除を行 うことは困難である。そのため化学農薬と微生物農薬の併用が必要と考えられている
(Chandler et al., 2011, Vemmer and Patel, 2013)。
放線菌は培地上のコロニーが放射状に広がることから古くは放射状菌ともよばれ、土壌
に多く存在する高 GC 含量(70%以上)グラム陽性桿菌である。また原核生物でありながら
糸状菌のように菌糸を出し複雑な構造をしており、菌糸の先端に胞子を形成する。菌糸は寒
天培地中に伸長する基底菌糸と寒天表面上から大気中に伸長する気菌糸の二種類に分けら
12 物質を産出する菌が多く存在し、これまで発見された抗生物質のうち約 3 分の 2 は放線菌 由来の生産物とされており、その抗生物質から殺虫剤や動物薬として実用化された物質も 少なくない(Zhu et al., 2014)。また、放線菌は抗菌活性物質だけでなく、リン酸の可溶化や シデロフォア、IAA などの植物ホルモンの生産による植物生長促進作用を持っている (Jog et al., 2012)。さらに、多くの放線菌は耐久性胞子の形成能を備えおり、中には袋状の胞子 嚢を形成する種類や運動性胞子を形成する種類も存在するため、高温や乾燥などに強いと されている(野々村, 1989)。これらの研究から、近年は植物病原菌に対する農薬としての利 用も進められている。 本研究では放線菌を用いた微生物農薬の開発を目的とし、土壌由来放線菌 KT および AR10 の植物病原菌に対する抑制能の評価を行った。また両菌株において抗真菌活性物質を 同定し、さらに植物を用いた植物病原菌の防除試験を行った。
13 2-2 材料および方法
2-2-1 微生物
放線菌 KT は、和歌山県高野山の樹の根圏から土壌を採取し、単離を行った。その後
potato dextrose broth agar (PDA, Difco; agar 1.5%)培地にて培養し 24 ℃で生育を行った。
菌株の保存はグリセリンを用いた凍結保存にて行った。AR およびその他の放線菌 52 株は、
アリの巣周辺土壌および他の環境土壌中から単離し、PDA 培地で培養および継代を行った。
対 峙 試 験 に 使 用 し た 植 物 病 原 菌 は 、 以 下 の 8 種 (Bipolaris oryzae MAFF 305382,
Colletotrichum echinochloae MAFF 305460, Colletotrichum orbiculare MAFF 306685,
Fusarium oxysporum MAFF 103038, Fusarium solani MAFF 235170, Monilinia fructigena
MAFF 305640, Phytophthora infestans MAFF 236324, Rhizoctonia solani MAFF 235846)
を使用し、培養および継代は PDA 培地で行った。これらの植物病原菌は、研究室で保存し ていた農業生物資源ジーンバンクから得たものを使用した。 2-2-2 放線菌を用いた植物病原菌に対する対峙培養試験 放線菌の植物病原菌に対する抑制活性は、寒天培地による対峙培養試験によって評価し た。PDA 培地で培養した植物病原菌をφ7 mm のステンレスカップでくり抜き、新しい PDA 培地の端から 15 mm の位置に植菌した。放線菌は、植物病原菌を植菌した反対側に植菌し、 7-14 日間、24℃で培養を行った。植物病原菌の抑制面積は、植物病原菌がプレート全面に 広がった際に、対峙培養したプレートにおける植物病原菌の増殖阻止面積を測定した。抑制 評価は、抑制面積が 10 cm2未満(-)、10 cm2以上 20 cm2未満(+)、20 cm2以上 30 cm2未
14
満(++)、30 cm2以上(+++)の 4 段階で評価した。
2-2-3 放線菌の同定
放線菌の同定は、16S rDNA 系統解析および形態学的観察から行った。株式会社テクノス
ルガ・ラボに委託し、得られた塩基配列は DB-BA11.0(Techno Suruga Laboratory)および
国際塩基配列データベース(DDBJ/ENA(EMBL)/GenBank)に対する BLAST 相同性検索を 参照して同定し、分子系統樹の推定は近隣結合法にて行った。形態観察は、寒天培地上にお けるコロニーの色調、気菌糸および気中菌糸の形成、胞子形成を観察した。 2-2-4 放線菌 KT 由来抗真菌活性物質の抽出および精製 1 L フラスコ(IWAKI)を使用して、放線菌を PDB 培地 800 mL で 15 日間、30 mm スタ ーラー(アズワン株式会社)を用いて攪拌培養(800 rpm)を行った。その後、吸引濾過にて 菌液を除去し、菌体を回収した。菌体を 50 mL 遠心チューブに移し、等量のメタノールを 加えて抽出を行った。その後、遠心分離にて菌体を沈殿させ、有機層を回収し、減圧乾固し た。抽出は計 3 回行った。その後、抽出で得られた KT 由来抗真菌活性物質を蒸留水に懸濁 し、遠心分離後、上清を回収した。得た上清は、逆相カートリッジカラム(Bond Elut 1000
mg )に供し、MeOH : H2O = 0 : 100、MeOH : H2O = 20 : 80、MeOH : H2O = 40 : 60、
MeOH : H2O = 60 : 40、MeOH : H2O = 80 : 20、MeOH : H2O = 100 : 0、 各 6 mL で溶出
した。溶出により得た画分を減圧乾固し、抗真菌活性が認められたメタノール 80 %および
100 % 画分をメタノールにて溶解し HPLC に供した。HPLC のシステムは LC2000(日本
15 社)を用いた。pump A には蒸留水、pump B には CH3CN(ナカライテスク株式会社)を使用 し、 A / B = 70 / 30 – 30 min – 60 / 40 (v/v) の条件で溶出し、流速は 1 mL / min で検出 波長 UV 320 nm で分析および分取を行った。 2-2-5 放線菌 AR10 由来抗真菌活性物質の抽出および精製 AR10 を 1 L フラスコに入った PDB 培地 800 mL で 1000 rpm、24 ℃、10 日前後、30 mm のスターラーを用いて攪拌培養した。分液漏斗に菌液を入れ、酢酸エチルを等量加え 分配抽出を行い、有機層を回収した。この作業を 3 回繰り返した。有機層は減圧乾固し、粗 抽出物を得た。AR10 の培養液から酢酸エチルで抽出した粗抽出物を調整し、500 µg を TLC
(Silica gel 60 F254)に供した。トルエン:酢酸エチル:酢酸=16:4:1 の展開溶媒で展開し、UV
254 nm で観察した。バイオオートグラフィー試験では、TLC を 5 つに切り分け、PDA 培 地の端から 15 mm の位置に TLC を置き、反対側にR. solani を植菌した。24 ℃で数日間 培養後、観察を行った。また。分取 TLC にて抗真菌活性が観察された画分を回収し、粗精 製物を得た。TLC で粗精製した抗真菌活性物質を HPLC に供し、化合物の精製を行った。 HPLC のシステムは LC2000(日本分光)を使用し、カラムは Inertsil ODS-3(φ 4.6×150 mm、ジーエルサイエンス株式会社) を使用して、流速 1.0 mL/min、検出波長 254 nm で 分析を行った。溶媒は ポンプ A に 蒸留水、ポンプ B に CH3CN を使用し、A/B=50/50 で 分析を行った。検出強度が最も高かったピークを分取し、対峙培養にて抗真菌活性物質の確 認を行った。
16 2-2-6 KT 由来抗真菌活性物質の LC-MS 分析
精製した KT 由来抗真菌活性物質をメタノールにて溶解し、LC-MS 分析に供した。LC
のシステムは、LC-20A (SHIMADZU)を使用し、 カラムは Inertsil ODS-3 (φ2.1×150
mm , ジーエルサイエンス株式会社)を使用した。分析は、流速 0.2 mL /min、 カラムオー
ブン 40℃、検出波長 320 nm で行った。溶出溶媒は、pump A に 0.1 %ギ酸添加超純水、
pump B に CH3CN を使用し、 A / B = 70 / 30 – 30 min – 60 / 40 (v/v) の条件で溶出し
た。MS は Triple TOF 5600 + (AB SCIEX) を使用し、ESI 法の positive mode にてイオン
化させ、Q1 scan ( m/z 100 - 2000 )の範囲で IDA(Information Dependent Acquisition )に
て測定を行った。
2-2-7 AR10 由来抗真菌活性物質の LC-MS 分析
精製した抗真菌活性物質をメタノールに溶解し、LC-MS 分析に供した。LC のシステム
は、LC-20A システム(SHIMADZU)を使用し、カラムは Inertsil ODS-3 (φ2.1×150 mm ,
ジーエルサイエンス株式会社)を使用した。分析は、流速 0.2 mL/min、カラムオーブン 40 ℃、
検出波長 254 nm で行った。溶出溶媒は、ポンプ A に 0.1%ギ酸水溶液、ポンプ B に CH3CN
を使用し、A/B=70/30 (v/v) -50 min- 20/80(v/v)の条件で溶出した。MS のシステムは
Qtrap ESI-MS を使用し、positive mode の Q1 scan (m/z : 100-800)の範囲にて検出を行っ
17 2-2-8 抗真菌活性物質の NMR 分析
精製した抗真菌活性物質は、DMSO-d6 (KT)もしくは CDCl3(AR10) で溶解した。KT の
抗真菌活性物質は、NMR Anova-500 (BRUKER Bio Spin) にて1H-NMR の分析を行った。
また、Peak 1 のサンプルには重水( D2O )を添加した。AR10 の抗真菌活性物質は、NMR
Anova-600 (BRUKER, Bio Spin) にて1H-NMR および13C-NMR の分析を行った。また、
2 次元 NMR については、H-H COSY、 HMBC における1H-1H および1H-13C カップリン グを検出した。どちらの分析においてもケミカルシフトの構成は使用した溶媒の残留プロ トンシグナルにて設定した。 2-2-9 キュウリを用いた KT による植物病原菌防除試験 2-2-9-1 R. solaniに対する感染防御試験 植物病原菌による作物への感染防除を試験するため、キュウリおよびチンゲンサイ用い て実験を行った。植物種子は、70%エタノールおよび 1%次亜塩素酸水溶液で殺菌処理を行 い、滅菌水にて 3 回洗浄した。殺菌した種子は、素寒天培地上で 3 日間栽培し、発芽させ た。発芽後、種子を改変ハイポネックス寒天培地(微粉 Hyponex ; 0.0375%, sucrose; 0.35%, agar; 0.8%, pH6.0)に移植した。種子はキュウリの場合は 3 個体、チンゲンサイの場合は 5 個体移植した。放線菌 KT は、PDB 培地で攪拌培養を 3 日間行い、培養液 200 µL を移植日 と同日に植菌した。移植から 5 日後にステンレスカップでくり抜いたφ0.7 cm のR. solani の寒天断片をアグリポットに中心に植菌し、感染防除試験を行った。試験は小型人工気象器 内で行い、15-20 日間、24℃で 12 時間の明暗周期で栽培した。また、同時に未滅菌の培養
18
土を用いて同様の試験を行った。アグリポットに添加した未滅菌土壌に植物を移植し、KT
の菌糸体を 2.0g添加した。病害重症度は 0-5 の 6 段階で評価し(0: no disease, 1: browning
of root, 2: browning of stem, 3: wilting of plants, 4: lodging of plants, 5: plant death)、式1で
算出した。
2-2-9-2 B. oryzaeに対する感染防除試験
B. oryzaeに対する感染防除試験は Detached Leaf Assay によって行った。植物はエンバ
クを用い、本葉の 2-3 葉期を使用した。回収した葉を洗浄し、15 mm の長さに切断した。
切断面をパラフィンで塞ぎ、1%次亜塩素酸水溶液で殺菌した。殺菌後、滅菌水で洗浄を 3
回行い、KT 培養液に 10 分間浸漬した。浸漬後、葉を湿らせたガーゼを敷いた 12 well プレ
ートに移した。B. oryzaeを培養したプレートから 4 mm2の寒天断片をくり抜き、葉切片に
植菌した。栽培は 25℃、12 時間の明暗周期で 12 日間栽培した。病害重症度は 0-4 の 5 段
階で評価し (0: no disease, 1: hyphal formation around the agar pieces, 2: hyphal coverage of
less than 50%, 3: hyphal coverage of 50-80%, 4: completely covered with hyphe)、式1で算
出した。
Disease index =Σ(score of disease ∗ number of plants)
total number of plants ・・・(1)
2-2-10 キュウリを用いた AR10 による植物病原菌防除試験
試験管に PDB 培地を 2 mL 分注し、そこに AR10 を植菌し、3 日間培養した。菌液を 1.5
19
たアグリポットに素寒天培地を 80 mL 作製した。新鮮重を 0.01 g に揃えた菌体を滅菌水
100 µL で懸濁し、素寒天培地に全面塗布した。1 つのアグリポットあたりに殺菌したキュ
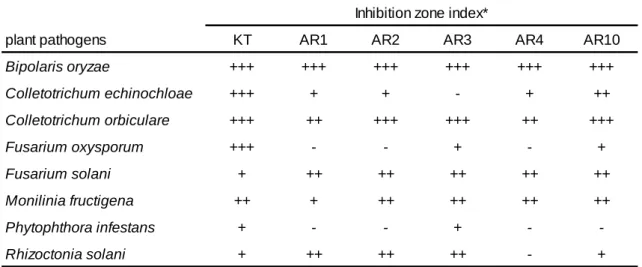
20 2-3 結果 2-3-1 放線菌の単離および選抜 18 か所の環境土壌サンプルから単離し、形態観察により合計 53 株の放線菌を単離した。 各放線菌について、R. solani に対する対峙培養試験を行い強い抗菌活性を示す菌株を選抜 した。その結果、6 株を選抜し、それぞれ KT、AR1、AR2、AR3、AR4 および AR10 とし た。これらの放線菌に対して、8 種の植物病原菌を用いた対峙培養試験を行った(Table 1)。 その結果、最も様々な植物病原菌に抑制を示したのは KT であり、特に B. oryzae、C.
echinochloae、C. orbiculare、F. oxysporumの 4 株に対して強い抑制活性が認められた。ま
た、次に AR10 がB. oryzaeおよびC.orbiculareに対して強い抑制活性を示し、他の AR 株
よりもわずかながら抑制活性は強かった。KT は比較的どの植物病原菌に対しても抑制活性
が認められたが、F. solani に対しては他の放線菌の方が強い抑制を示した。逆に、F.
oxysporumに対しては、KT 以外の放線菌はあまり強い抑制活性を示さなかった。これらの
21
Table 1 選抜した放線菌の植物病原菌に対する抗真菌活性試験
plant pathogens KT AR1 AR2 AR3 AR4 AR10
Bipolaris oryzae +++ +++ +++ +++ +++ +++ Colletotrichum echinochloae +++ + + - + ++ Colletotrichum orbiculare +++ ++ +++ +++ ++ +++ Fusarium oxysporum +++ - - + - + Fusarium solani + ++ ++ ++ ++ ++ Monilinia fructigena ++ + ++ ++ ++ ++ Phytophthora infestans + - - + - -Rhizoctonia solani + ++ ++ ++ - +
Inhibition zone index*
22 2-3-2 16S rDNA 系統解析による放線菌の同定
KT および AR10 の微生物種の同定を行った。KT は、寒天培地上で表面は白色、裏面は
褐色のコロニーを形成し、気菌糸を形成したが胞子の形成は確認されなかった。テクノスル
ガ・ラボ微生物同定システムを用いた DB-BA11.0 および国際塩基配列データベースに対す
る BLAST 相同性検索の結果、KT の 16S rDNA 塩基配列はStreptomyces morookaensis の
基準株 NBRC 13416T (アクセッション番号 AB184878 および NR_112529 )に対し相同率 99.8 % の相同性を示した。また、S. lavenduligriseus NRRL B-3173T (NR_043824 )に対し 相同率 99.6 % の相同性を示した。DB-BA11.0 および国際塩基配列データベースに対する 相同性検索で得られた塩基配列を基に解析した分子系統樹において、KT は Streptomyces 属で構成されるクラスター内に含まれ、Streptomyces lavenduligriseus NRRL B-3173T (NR_043824)とクラスターを形成したが、両者の間にはやや距離が認められ互いに異なる
分子系統学的位置を示した(Fig. 1)。以上のことから KT は Streptomyces lavenduligriseus
に近縁なStreptomyces sp.と同定した。一方、AR10 は培地上で綿状のコロニーを形成し、
グラム染色の結果、放線菌特有の菌糸や連鎖胞子の形成が確認された。DB-BA11.0 および
国際塩基配列データベースに対する BLAST 相同性検索の結果、AR10 の 16S rDNA 塩基配
列はStreptomyces lanatus の基準株 NBRC 12787T(AB 184845)に対し相同率 98.7%の最も
高い相同性を示した。また、AR10 の塩基配列と相同性検索で得られた塩基配列を基に解析
し、作成した分子系統樹において、Streptomyces 属で構成されるクラスター内に含まれた
ため、Streptomyces sp.であると推定したが、作製した分子系統樹内のいずれの既知種とも
23
Fig. 1 KT の 16S rDNA 塩基配列に基づく分子系統樹
24
Fig. 2 AR10 の 16S rDNA 塩基配列に基づく分子系統樹
25 2-3-3KT の生産する化合物の精製 放線菌 KT からの抗真菌活性物質の抽出は菌糸体から行った。PDB 液体培地で攪拌培養 を行った KT の培養液を濾過し、菌糸体を回収した。回収した菌糸体にメタノールを添加 し、抗真菌活性物質を抽出した。その結果、粗抽出物 500 µg で植物病原菌R. solani に対 して抑制を示した。その粗抽出物を用いて逆相カートリッジカラムに供し、抗真菌化合物の 分離を行ったところメタノール 80%および 100% の画分にて抗真菌活性物質が認められた。 これらの画分を合一し、HPLC に供した。保持時間 18.0 分、21.1 分、 23.6 分、 25.6 分、
27.7 分、28.9 分に peak が検出された(Fig. 3)。これら 6 つの peak を分取したところ、peak
1、peak 3 および peak 4 にて抗真菌活性が認められた(Fig. 3)。それぞれの peak は 100
26
Fig. 3 KT 粗抽出物の HPLC 分析
菌体から酢酸エチルで抽出した粗抽出物を HPLC に供した。番号は Peak 番号を示し、分取した各々の Peak
について抗真菌活性試を行った。分取した化合物は 100 µg/disk に調整した。写真下の番号は各 Peak 番号 を示す。
27 2-3-4 Streptomyces sp. KT の生産する化合物の LC-MS 分析 HPLC により精製した KT 由来抗真菌活性物質をメタノールにて溶解し、LC-MS に供し た。保持時間 18.0 分の Peak 1 の MS スペクトルを解析したところ、抗真菌活性物質の母骨 格と思われるシグナルm/z 766.4 ([M+H]+ ) が得られた。また、MSMS から水分子および 糖の脱離が見られるフラグメントイオンm/z 748.4 ([M+H-H2O]+)、m/z 603.3
([M+H-R-H2O]+) が検出された(Figs. 4A,B)。 保持時間 23.6 分の Peak 3 の MS スペクトルを解析し
たところ、m/z 780.4 ([M+H]+ )、m/z 762.4 ([M+H-H
2O]+)、m/z 617.3 ([M+H-R-H2O]+) が検出された(Figs. 5A,B)。保持時間 25.6 分の Peak 4 の MS スペクトルを解析したところ、
m/z 780.4 ([M+H]+ )、m/z 762.4 ([M+H-H
2O]+)、m/z 617.3 ([M+H-R-H2O]+) が検出さ れた(Figs. 6A,B)。
28 Fig. 4 KT 由来抗真菌活性物質 Peak 1 の LC-MS 分析 分取した Peak 1 の LC-MS 分析(A)および LC-MS/MS 分析(B)を示し、矢印は化合物の母骨格および フラグメンテーションのシグナルを示す。 A B [M+H]+ (m/z 766.4) (m/z 748.4) [M+H-H2O] + (m/z 603.3) [M+H-R-H2O] +
29 Fig. 5 KT 由来抗真菌活性物質 Peak 3 の LC-MS 分析 分取した Peak 3 の LC-MS 分析(A)および LC-MS/MS 分析(B)を示し、矢印は化合物の母骨格および フラグメンテーションのシグナルを示す。 A B [M+H]+ (m/z 780.4) [M+H-R-H2O] + (m/z 617.3) (m/z 762.4) [M+H-H2O] +
30 Fig. 6 KT 由来抗真菌活性物質 Peak 4 の LC-MS 分析 分取した Peak 4 の LC-MS 分析(A)および LC-MS/MS 分析(B)を示し、矢印は化合物の母骨格および フラグメンテーションのシグナルを示す。 A B [M+H]+ (m/z 780.4) [M+H-R-H2O] + (m/z 617.3) (m/z 762.4) [M+H-H2O] +
31 2-3-5AR10 の生産する化合物の精製 AR10 AR10 の生産する抗真菌活性物質を抽出および検出するため、TLC 分析を行った。TLC 分析の結果、複数のスポットが検出された(Fig. 7)。これらの化合物の中に抗真菌物質が含 まれているかを確認するため、バイオオートグラフィー試験を行った。その結果、Rf=0.44 の化合物で抗真菌活性が認められた(Fig. 7)。抗菌活性が認められたこの化合物を TLC で 分取し、HPLC で分析を行った。回収した抗真菌化合物の HPLC 分析結果を Fig. 8 に示す。 保持時間 11 分にピークが検出され、このピークを再度抗真菌活性試験に供した結果、抗菌 活性が認められた。
32 Fig. 7 TLC 分析による抗菌活性物質の検出 酢酸エチル抽出物を TLC 分析に供した。TLC プレート(Silica gel 60 F254)は UV 254 nm で観察した。展 開溶媒は、トルエン:酢酸エチル:酢酸=16:4:1 の溶媒を使用した。TLC は 5 画分に切断し、バイオオ ートグラフィーに供した。番号は切断した画分の番号を示し、バイオートグラフィーの写真と一致する。 植物病原菌にはR. solaniを使用した。 2 1 3 4 5 2 1 4 5 3
33
Fig. 8 AR10 粗精製物の HPLC 分析および抗真菌活性試験
TLC 分析で検出した抗真菌活性物質を回収し、HPLC 分析に供した。分析は 50%CH3CN のアイソクラテ
ィック溶出で行った。3 画分と洗浄部分を回収し、抗真菌活性試験を行った。番号は回収した画分を示す 1: 保持時間 0-10 分、 2: 10.5-11.5 分、3: 12-20 分。
34 2-3-6 AR10 の生産する化合物の LC-MS 分析
HPLC で精製し、抗真菌活性が認められた化合物の LC/MS 分析を行った。LC/MS 分析
の結果、保持時間 24 分にピークが検出され、この化合物の MS スペクトルを取得したとこ
35
Fig. 9 AR10 由来抗真菌活性物質 LC-MS 分析
分取した抗真菌活性物質の UV クロマトグラム(A)、トータルイオンクロマトグラム(B)および MS ス
ペクトル(C)を示す。MS スペクトルの矢印は化合物の母骨格のシグナルを示す。分析は 30%-40%CH3CN
36 2-3-7 精製した抗真菌活性物質のNMR分析
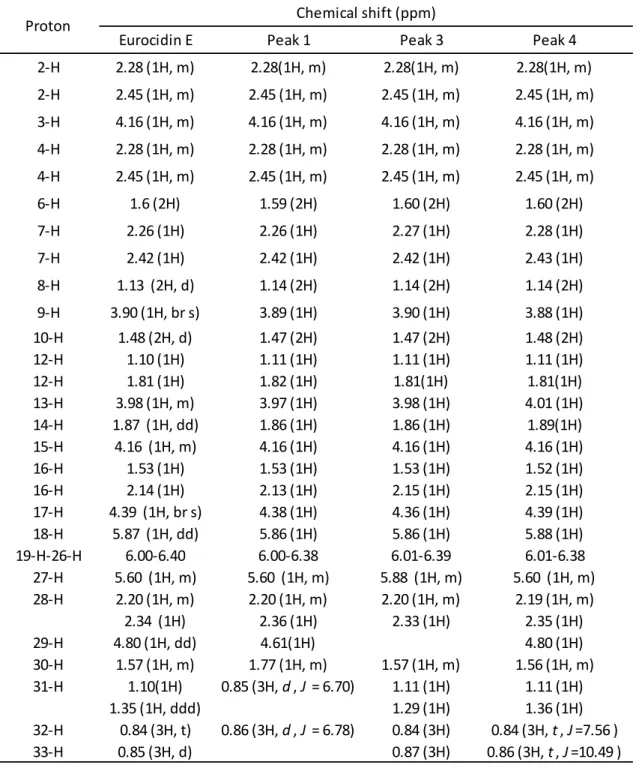
化合物の構造決定を行うため、LC-MS 分析と同時に NMR 分析を行った。KT 由来抗真
菌活性物質を DMSO-d6 にて溶解し NMR に供した。Peak 1 の1H-NMR 解析を行ったと
ころ、(Nakagomi et al., 1990) らが発見した eurocidin E と類似するシグナルが検出された
( Figs. 10A,B)。eurocidin E と Peak1 の化合物の化学構造とケミカルシフトを比較したとこ
ろ、eurocidin E の 30 位のケミカルシフトが 1.57 ppm のマルチプレット、31 位が 1.10
ppm のマルチプレット、32 位が 0.84 ppm のトリプレット、33 位が 0.85 ppm のダブレッ
トであったのに対し、今回得た Peak1 の化合物の 30 位のケミカルシフトが 1.77 ppm のマ
ルチプレット、31 位が 0.85 ppm のダブレット、32 位が 0.86 ppm のダブレットであったこ
とから、Peak 1 は Eurocidin E の 29 位アルキル側鎖が sec - ブチル基から、iso -プロピ
ル基に置換された化合物であると同定した ( Figs. 10A,B,E,D および Table 2)。 また、Peak
3、Peak 4 の1H-NMR 解析結果から、Nakagomi らが報告した eurocidin E と酷似するシグ
37
A eurocidin E eurocidin E (Nakagome, 1990)
B peak 1
38
Fig. 10 KT 由来抗真菌活性物質の1H-NMR 解析
Nakagome らによって報告された eurocidin E(A)および抗真菌活性が認められた peak 1(B), peak 3(C), peak 4(D)の NMR 解析の結果を示す。NMR 解析によって構造決定された eurocidin E(E)および今回単 離した新規 eurocidin(D)を示す。
D peak 4
E
39
Table 2 KT 由来抗真菌活性物質の1H-NMR(500 Mhz, DMSO-d6)
Eurocidin E Peak 1 Peak 3 Peak 4
2-H 2.28 (1H, m) 2.28(1H, m) 2.28(1H, m) 2.28(1H, m) 2-H 2.45 (1H, m) 2.45 (1H, m) 2.45 (1H, m) 2.45 (1H, m) 3-H 4.16 (1H, m) 4.16 (1H, m) 4.16 (1H, m) 4.16 (1H, m) 4-H 2.28 (1H, m) 2.28 (1H, m) 2.28 (1H, m) 2.28 (1H, m) 4-H 2.45 (1H, m) 2.45 (1H, m) 2.45 (1H, m) 2.45 (1H, m) 6-H 1.6 (2H) 1.59 (2H) 1.60 (2H) 1.60 (2H) 7-H 2.26 (1H) 2.26 (1H) 2.27 (1H) 2.28 (1H) 7-H 2.42 (1H) 2.42 (1H) 2.42 (1H) 2.43 (1H) 8-H 1.13 (2H, d) 1.14 (2H) 1.14 (2H) 1.14 (2H) 9-H 3.90 (1H, br s) 3.89 (1H) 3.90 (1H) 3.88 (1H) 10-H 1.48 (2H, d) 1.47 (2H) 1.47 (2H) 1.48 (2H) 12-H 1.10 (1H) 1.11 (1H) 1.11 (1H) 1.11 (1H) 12-H 1.81 (1H) 1.82 (1H) 1.81(1H) 1.81(1H) 13-H 3.98 (1H, m) 3.97 (1H) 3.98 (1H) 4.01 (1H) 14-H 1.87 (1H, dd) 1.86 (1H) 1.86 (1H) 1.89(1H) 15-H 4.16 (1H, m) 4.16 (1H) 4.16 (1H) 4.16 (1H) 16-H 1.53 (1H) 1.53 (1H) 1.53 (1H) 1.52 (1H) 16-H 2.14 (1H) 2.13 (1H) 2.15 (1H) 2.15 (1H) 17-H 4.39 (1H, br s) 4.38 (1H) 4.36 (1H) 4.39 (1H) 18-H 5.87 (1H, dd) 5.86 (1H) 5.86 (1H) 5.88 (1H) 19-H-26-H 6.00-6.40 6.00-6.38 6.01-6.39 6.01-6.38 27-H 5.60 (1H, m) 5.60 (1H, m) 5.88 (1H, m) 5.60 (1H, m) 28-H 2.20 (1H, m) 2.20 (1H, m) 2.20 (1H, m) 2.19 (1H, m) 2.34 (1H) 2.36 (1H) 2.33 (1H) 2.35 (1H) 29-H 4.80 (1H, dd) 4.61(1H) 4.80 (1H) 30-H 1.57 (1H, m) 1.77 (1H, m) 1.57 (1H, m) 1.56 (1H, m) 31-H 1.10(1H) 0.85 (3H, d , J = 6.70) 1.11 (1H) 1.11 (1H) 1.35 (1H, ddd) 1.29 (1H) 1.36 (1H) 32-H 0.84 (3H, t) 0.86 (3H, d , J = 6.78) 0.84 (3H) 0.84 (3H, t , J =7.56 ) 33-H 0.85 (3H, d) 0.87 (3H) 0.86 (3H, t , J =10.49 )
40
AR10 の抗真菌活性物質も同様に NMR 解析に供した。化合物は重水素化クロロホルム
(CDCl3)に溶解し、NMR に供した。1H-NMR の結果、27 個のシグナルが確認され、13
C-NMR の結果、18 個のシグナルが確認検出された(Fig. 11 および Table 3)。H-H COSY と
HMBC による解析を行ったところ、H-H COSY では、[5.83 ppm - 6.84 ppm]、[5.73 ppm - 5.61 ppm]、[5.61 ppm - 4.03 ppm]、[1.60 ppm - 5.24 ppm]、[5.24 ppm - 2.10 ppm]、 [5.24 ppm - 1.80 ppm]、[2.10 ppm - 1.19 ppm]、[2.10 ppm -1.13 ppm]、[1.80 ppm - 1.19 ppm]、[1.80 ppm - 1.13 ppm]、[1.19 ppm – 1.40 ppm]、[1.13 ppm – 1.40 ppm]、[1.40 ppm - 0.84 pm]、[1.40 ppm - 1.17 ppm]の間で相関が認められた(Fig. 12)。 HMBC において [4.51 ppm - 166.3 ppm]、[6.84 ppm - 166.3 ppm]、[5.83 ppm - 73.0 ppm]、[5.73 ppm - 73.0 ppm]、[5.61 ppm - 1.49 ppm]、[4.03 ppm - 56.9 ppm]、[4.03 ppm - 13.9 ppm]への相関が 認められた。以上の結果から、AR10 の生産する抗真菌活性物質は albocycline であると同 定した。
41
Fig. 11 AR10 由来抗真菌活性物質の NMR 解析
分取した抗真菌活性物質の1H-NMR(A)および13C-NMR(B)。
A
42 Table 3 KT 由来抗真菌活性物質の1H-NMR および13C-NMR(600 MHz, CDCl-d) d 1H d 13C 1 166.3 2 5.83(1H,
d
,J
=15.4) 115.3 3 6.84(1H,d
,J
=15.5) 154.8 4 73.0 5 5.73(1H,dd
,J
=0.5,16.2) 135.9 6 5.61(1H,dd
,J
=6.2,16.2) 130.6 7 4.03(1H,d
,J
=6.2) 84.8 8 136.5 9 5.24(1H,t
,J
=6.3) 129.1 10 1.80(1H,m
) 24.6 10' 2.10(1H,m
) 11 1.13(1H,m
) 34.2 11' 1.19(1H,m
) 12 1.40(1H,m
) 39.0 13 4.51(1H,dq
,J
=6.4,8.5) 75.5 4-Me 1.49(3H,s
) 26.9 7-OMe 3.26(3H,s
) 56.9 8-Me 1.60(3H,s
) 13.9 12-Me 0.84(3H,d
,J
=6.9) 15.6 13-Me 1.17(3H,d
,J
=6.4) 17.843
Fig. 12 AR10 由来抗真菌活性物質の H-H COSY および HMBC
分取した抗真菌活性物質の1H-NMR および13C-NMR(A)、H-HCOSY(C)、HMBC(D)。解析した H-H
COSY および HMBC より認められた各シグナルの相関を albocycline に照らし合わせたものを示す(B)。
A B
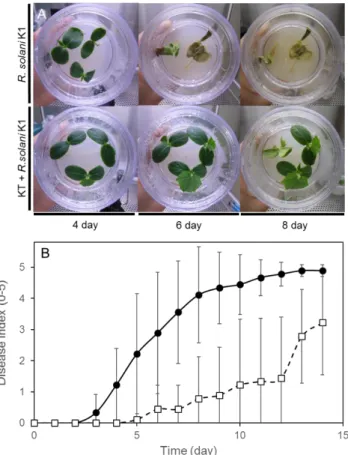
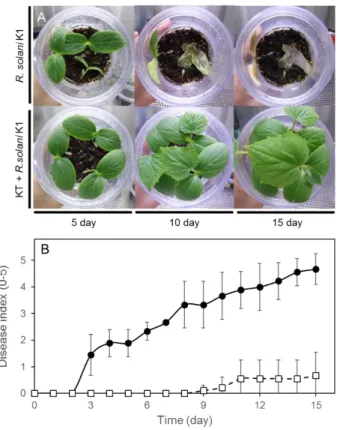
44 2-3-8KT を用いたR. solani の感染防除試験 本試験で最も抑制活性が高かった放線菌 KT を用いてキュウリ個体におけるRhizoctonia による立枯病の感染防除試験を行った。接種後 3 日目に対照区ではR. solaniの初期感染が 認められ、6-8 日目には枯死などの重篤な状態を示す個体が観察された(Fig. 13A)。しかし、 KT を処理した区では、12 日において病害重症度 1 程度であり、感染個体は認められたも のの、対象区に比べ病害重症度低く、重篤な症状は観察されなかった(Fig. 13B)。キュウリ を用いた試験では、R. solaniの感染を明らかに抑制していたため、次にアブラナ科の植物種 であるチンゲンサイを用いて同様の試験を行った。
45
Fig. 13 KT によるキュウリ立枯病(R. solani )の感染防除試験
写真は、KT によるR. solaniの感染防除を示し、上段がR. solaniのみ、下段に KT とR. solaniを植菌した
ものを示す(A)。グラフは、時間経過における感染防除を病害重症度で示した(B)。R. solani のみ(●)
R. solaniおよび KT(□)を接種したものを示す。 病害重症度は、試験中の各期間に基づき、算出した。
46
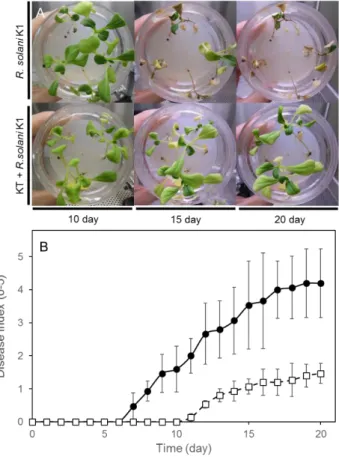
チンゲンサイを用いた試験において最初の感染は 7
日目に対象区で観察され、その後、15-20 日目にはキュウリと同様に重度の病害が観察された。KT を処理した区では、10 日目ま
では感染が認められず、 R. solaniによる感染を防除していた。11 日目に始めてR. solaniの
感染が観察され、その後、徐々に感染が認められたが、最終的に病害重症度は 1 程度と対象
区に比べ低い値であった(Fig. 14A,B)。これらの結果から、KT による R. solaniの著しい
感染防除効果が認められため、実際の圃場と同様の未殺菌土壌を用いて同様の実験を行っ
た。Fig. 15, 16 に示すように、 未滅菌土壌においても KT はR. solaniの感染に対する顕著
47
Fig. 14 KT によるチンゲンサイ立枯病(R. solani )の感染防除試験
写真は、KT によるR. solaniの感染防除を示し、上段がR. solaniのみ、下段に KT とR. solaniを植菌した
ものを示す(A)。グラフは、時間経過における感染防除を病害重症度で示した(B)。R. solani のみ(●)
R. solaniおよび KT(□)を接種したものを示す。 病害重症度は、試験中の各期間に基づき、算出した。
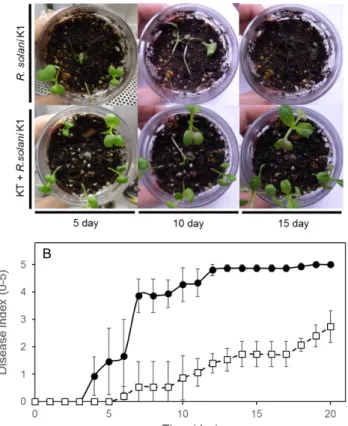
48
Fig. 15 KT によるキュウリ立枯病(R. solani )の感染防除試験(未滅菌土壌)
写真は、KT によるR. solaniの感染防除を示し、上段がR. solaniのみ、下段に KT とR. solaniを植菌した
ものを示す(A)。グラフは、時間経過における感染防除を病害重症度で示した(B)。R. solani のみ(●)
R. solaniおよび KT(□)を接種したものを示す。 病害重症度は、試験中の各期間に基づき、算出した。
49
Fig. 16 KT によるチンゲンサイ立枯病(R. solani )の感染防除試験(未滅菌土壌)
写真は、KT によるR. solaniの感染防除を示し、上段がR. solaniのみ、下段に KT とR. solaniを植菌した
ものを示す(A)。グラフは、時間経過における感染防除を病害重症度で示した(B)。R. solani のみ(●)
R. solaniおよび KT(□)を接種したものを示す。 病害重症度は、試験中の各期間に基づき、算出した。
50 2-3-9 KT を用いたB. oryzae の感染防除試験
R. solaniに対する明確な抑制効果が観察されたことから、次にイネ、トウモロコシおよびコ
ムギなどを含むイネ科植物における重要な植物病原菌としてB. oryzaeを試験に使用し、試
験を行った。KT がB. oryzaeを防除できるかどうかを評価するために、エンバク葉を使用
し、detached leaf aasay を行った。その結果、KT 培養液を浸漬処理した葉は、植物病原菌
の増殖を抑制し、葉に病害はほとんど認められなかったが、無処理である対象区の葉は病原
体によって覆われていた(Fig. 17A)。 KT を処理した葉は、試験の最終日にもB. oryzaeの
51
Fig. 17 KT による葉枯病(B. oryzae)の感染防除試験
写真は、KT によるB. oryzaeの感染防除を示し、上段がB. oryzaeのみ、下段に KT とB. oryzaeを植菌し
たものを示す(A)。グラフは、時間経過における感染防除を病害重症度で示した(B)。B. oryzaeのみ(●)
B. oryzaeおよび KT(□)を接種したものを示す。 病害重症度は、試験中の各期間に基づき、算出した。
52 2-4 考察 植物の根圏には様々な微生物が存在する。植物と微生物との間の相互作用は、植物の生育 に良い影響だけでなく悪い影響を及ぼす可能性がある。微生物農薬は、植物の生長を促進し、 植物病原菌を抑制することができると考えられている。放線菌は、植物病害の潜在的な生物 学的防除剤の 1 つであり、有用な抗菌活性物質を産生することがすでに知られている。 本研究では、環境中から 53 株の放線菌を単離した。KT は樹の根圏から単離したが、AR1、 AR2、AR3、AR4、AR10 はアリの巣周辺から単離した。これは Haeder らの研究において、 ハキリアリと放線菌が共生し(Haeder et al, 2009)、ハキリアリの巣の環境を整えると報告 されていたことからアリの巣周辺土壌には植物病害防除に有用な放線菌が存在しているの ではないかとの考えで行った。ハキリアリは自らの餌(キノコ)を栽培するという特徴を持 つが、餌となるキノコの病原菌を抑制するため、放線菌を利用している。このハキリアリの 生活形態は、微生物農薬の概念と一致しているのではないかと考え、AR 株の単離を行った。 その後、選抜した KT と AR10 について微生物同定を行った。 テクノスルガ・ラボ微生物同定システムを用いた DB-BA11.0 および国際塩基配列データ
ベースに対する BLAST 相同性検索によると、KT と Streptomyces lavenduligriseus NRRL
B-3173T (NR_043824)の 16S rDNA 塩基配列間には 6 塩基の相違点が認められ、その内の
2 塩基は挿入/欠損によるものであり両者の明確な差として捉えることは困難であったが、
残りの 4 塩基は明らかに異なっていた。このことから KT は Streptomyces lavenduligriseus
に近縁なStreptomyces sp.と同定された(Fig. 1)。Streptomyces lavenduligriseusを用いた微
53
された。同様に AR10 の同定の結果、AR10 も同様にStreptomyces 属で構成されるクラス
ター内に含まれたため(Fig. 2)、Streptomyces sp.であると推定したが、作成した分子系統 樹 内 のい ず れの 既知 種と も 異な る 分子 系統 学的 位 置を 示 した ため 、 AR10 は新 種 の Streptomycesではないかと考えられた。 今回、単離および選抜した放線菌 KT および AR10 は対峙培養試験において多くの植物 病原菌を抑制し、幅広い抗真菌スペクトルを示した(Table 1)。これらの結果は、広範囲の 植物病原体を制御するために複数の抗真菌代謝産物の存在を示唆している。これらの抗真 菌活性物質を単離し、構造決定を試みた結果、KT からは eurocidin 類が同定され、AR10 か
らは albocycline が同定された。KT から精製した抗真菌活性物質である Peak 3 と Peak 4 の
MS スペクトルを解析したところ、抗真菌活性物質の母骨格と思われるシグナルm/z 780.4 ([M+H]+ ) が得られた(Figs. 5,6)。これらの化合物を MS スペクトルの同位体比から組成を 割り出して化合物検索を行った所 C40H61NO14 を示す化合物の可能性が高いと考えられた。 同様に Peak 1 の MS スペクトルを解析したところ、m/z 766.4 ([M+H]+ ) のシグナルが得 られ、MS スペクトルの同位体比より C39H59NO14 を示す化合物の可能性が高いと考えられ た(Fig. 4)。LC-MS 分析の結果による組成からは物質の同定まで至らなかったため NMR 分 析にて抗真菌活性物質の同定を試みた。NMR 分析の結果、今回単離した化合物は 1990 年
に Nakagomi が Streptoverticillium eurocidicum から単離した eurocidin E の異性体 (Fig.
10)ではないかと推測された。既知である化合物 eurocidin E の 29 位アルキル側鎖が sec -
ブチル基(-CH(CH3)CH2CH3)から、iso -プロピル基(-CH(CH3)2)に置換された新規化
54
なカップリングを示すことが出来ないため、今後サンプル量を増やし詳細な構造を見てい
く必要がある。eurocidin 類は放線菌由来の物質として 1955 年に、Nakazawa が報告してお
り(Nakazawa, 1955)、その後、eurocidin A、B、C、D、E というポリケチド化合物が Nakagome
らによって発見された(Nakagomi et al., 1990) 。これらの物質はマウスの肥満細胞脱顆粒抑
制、抗炎症作用などの研究が行われている。抗真菌・抗原虫活性としてはCandida albicans、
Rhizoctonia solani、 Trichomonas vaginalisなどに対する活性が既に知られている(Pandey
and Rinehart, 1976)。しかしながら、eurocidin 類は 1954 年から 1990 年代までは研究が行
われていたが、それ以降の研究報告はほとんど行われていない。eurocidin E の物性として
は 淡 黄 色 の 粉 で メ タ ノ ー ル に 溶 け る が 、 水 や ヘ キ サ ン に は 難 溶 で あ る 。 ま た 、S.
lavenduligriseus からの eurocidin 生産に関する論文はこれまでに報告されていない。
一方、AR10 の生産する albocycline は、すでに多くの報告がなされており、Streptomyces
maizeus UC-5130 や Streptomyces venezuelae、Streptomyces bruneogriseus など様々な
Streptomyces属やStreptomyces属以外の放線菌であるPropionicimonas sp. が生産してい
ることが報告されているが(Slechta et al., 1978, Zucchi et al., 2014, Harada et al., 1984)、
今回作成した系統樹内の AR10 周辺の Streptomyces 属については albocycline の生産が報
告されていない。
微生物同定の結果と物質生産から AR10 はこれらの菌に近縁な新種の菌である可能性が
ある。微生物農薬の候補として両菌株の同定および化合物の単離および構造決定を行い、次
に微生物農薬としてより応用的な試験を行うために、KT を用いて植物病原菌の防除試験を
55 キュウリまたはチンゲンサイ感染防除試験によって、KT の病害抑制能を試験した。 KT は R. solani による立枯病からキュウリまたはチンゲンサイを保護することができた(Fig. 13、14)。感染防除試験は、Hyponex 培地および未殺菌土壌の両方で行った。両方の試験で 植物病原体に対する KT の抑制効果が認められた(Fig. 15、16)。これまでの研究では、培 地や滅菌土を使用することが多いが、滅菌されていない土壌を使用する報告は少ない。未滅 菌土壌を用いた実験では、未滅菌土壌中に存在する微生物の影響を避けることができず、目 的微生物の増殖が困難である。しかし、KT は未滅菌土壌条件下で植物病原菌の増殖を抑制 することができた。この結果は、KT 培養による前処理が、一般的な圃場においても植物病 原菌の感染を減少させることができる可能性を示唆するものである。 本研究では、R. solaniによる立枯病だけでなく、B. oryzae による葉枯病についても感染 防除試験を試みた。 B. oryzaeはイネ科植物の葉に褐色の斑点を引き起こすことが知られて おり、葉枯病は 1-10%の間の重大な損失を引き起こす(Sarvary et al., 2000, 2006)。葉枯病 は、いもち病や紋枯病(Matsumoto et al., 2016)に続いて、日本ではイネの三大病害として
知られており、これらの病害防除が重要な課題である。KT は、detached leaf assay におい
てB. oryzaeの感染を大幅に遅らせることができた。したがって、KT は、今回使用した 8 種の植物病原菌全てに対して抑制効果が認められたことも考慮すると、多くの植物病原菌 を防除する微生物農薬として期待でされる。また、AR10 は抗菌スペクトルの広さや抗菌活 性の強さでは KT に劣るが、今後同様の植物試験を行い、より圃場に近い条件下での試験や KT との組合せによる病害防除効果の向上が認められるかなど、また違った方法を検討する ことによって新規の微生物農薬の開発に貢献できるのではないかと考えられる。
56 2-5 結論
本章では放線菌であるStreptomyces sp. KT およびStreptomyces sp. AR10 を単離および
同定した。両菌株は、単離した 53 株の放線菌の中でも植物病原菌に対して強い抑制能を有
していた。各菌株の生産する抗真菌活性物質を抽出および精製し、構造決定を行った結果、
KT からは、eurocidin E(Peak 3 および 4,同位体)および新規だと示唆される eurocidin が
得られた。eurocidin の研究は 1980 年代からほぼ行われておらず、今回の報告は、この化合 物の微生物農薬として新たな研究を指し示すことに繋がるかもしれない。一方、AR10 が生 産する抗真菌活性物質は既知物質である albocycline であった。しかしながら、両菌株の近 縁 種 に お い て 各 化 合 物 の 生 産 は 報 告 さ れ て お ら ず 、 同 時 に 各 化 合 物 を 生 産 す る Streptomyces 属を用いた微生物農薬の報告もまだなされていないため、これらの菌を用い た農薬の研究は新規微生物農薬の開発に貢献すると考えられる。実際に、応用的な研究とし て、KT を用いた植物病原菌の防除試験では、R. solaniおよびB. oryzaeに対して高い防除 効果を発揮した。どちらの植物病原菌も世界各国で重篤な病害を引き起こし、作物生産に多 大なる影響を及ぼす病原菌であることから、これらを防除することができる KT を使用し た微生物農薬の開発は、今後の作物生産に貢献するのではないかと考えられる。AR10 を始 めとする AR 株は、今後同様の植物病感染防除試験を行い、微生物農薬候補株として評価す る必要があると考えている。
57
第3章 未利用植物残渣を用いた
Bacillus subtilis RB14 の固体培養
3-1 序論 緒論でも述べた通り、作物生産にとって農薬は必要不可欠である。近年では、化学農薬の 使用を減らし、微生物農薬などの生物学的防除剤との併用が支持され始めている。微生物農 薬としてよく利用されている菌に Bacillus 属細菌が挙げられ、日本でも同菌を用いた製剤 が市販されている。Bacillus subtilisは、数種類の作用によって微生物農薬としての効果を発 揮している。その中でも特に強い抗真菌活性を有する化合物に非リボソームペプチド合成酵素(Non-ribosomal peptide synthetase)によって生合成される環状リポペプチドがある。こ
の環状リポペプチドは、一定数のアミノ酸と脂肪酸残基が結合した両親媒性の化合物であ
り、植物病原菌の生体膜に作用し、細胞流出による細胞死を起こす。(Zeriouh et al., 2014)。
Bacillus属の生産する環状リポペプチドは iturin、fengycin、surfactin の 3 クラスに分類さ
れ、中でも、iturin と fengycine は強い抗真菌活性を示す。また、Bacillus属の特徴としてバ
イオフィルム形成能が挙げられ、Bacillus属はこの能力によって植物体で強靭なコロニーを 保持するのに役立てている。特に植物個体の表面部分は温度や湿度の急激な変化、また栄養 の枯渇などのため微生物の生育に適さないが、幅広い環境で生存可能な Bacillus 属細菌を 微生物農薬として用いることで植物個体の表面部分から侵入するような病原菌の防除が可 能となると報告されている(Zeriouh et al., 2014)。しかしながら、微生物農薬として利用す るためには、一定の菌数が必要となり、そのために比較的安価な培養基質が必要となる。本 研究では、培養基質としてJatropha curcasの種子残渣に着目した。
58 る植物である。BDF はカーボンニュートラルな資源からのエネルギー生産であるため、温 室効果ガスの削減に貢献できると期待されている。BDF は主に菜種、コーン、パーム油、 大豆、サトウキビといった従来食料として利用しているものが多く、燃料として利用した際 に食料との競合が懸念される。そこで、非食料用油からの BDF 製造が求められており、J. curcasのような非食用で油脂生産能の高い植物が注目されている。 J. curcas はトウダイグサ科の落葉低木である。生育速度が速く、干ばつや貧栄養土壌に も強いといった特徴を有しており、50 年に渡り種子生産が可能である(Sotolongo et al.,2007)。J. curcasの種子は 41%の外殻と 59%の仁で構成されており、仁の 40-50%は油
脂で構成されている (Saetae and Suntornsuk, 2011)。J. curcas 種子由来の油は優れた BDF
の候補であるが、多量の種子採油残渣廃棄物が生じる問題がある(Liang et al., 2010)。仮 に、J. curcas 種子からバイオディーゼル燃料を生産しようとすると、種子 1 kg あたり 500 g の種子残渣が生じる(Sotolongo et al.,2007)。 種子残渣物の 30%程度はタンパク質であ り(Makkar et al., 1997)、採油処理後の残渣物は、高い栄養と豊富なタンパク質が含有して いるが、種子には抗栄養因子であるサポニン、フィチン酸、トリプシン阻害物、グルコシノ レート、アミラーゼ阻害物などや(Rakshit et al. 2008)、毒性物質であるホルボールエステ ル類なども含まれているため、家畜飼料としては適さず、付加価値の低い植物用肥料または 廃棄物として処理されている。またJ. curcas がもつホルボールエステル類はテルペン類化 合物であり、ホルボールエステルが細胞膜を透過し、細胞内にあるジアセチルグリセロール を介した細胞増殖を促すシグナル伝達系に作用することで、細胞内の本シグナル系にある プロテインキナーゼ C を活性化し、発がんのプロモーターとなることが知られている
59 (Blumberg, 1988)。 本研究では、微生物農薬候補株であるB. subtilis RB14 の培養基質として油脂抽出後のJ. curcas種子残渣を利用し、微生物農薬として農業分野への応用を目的とした。また、培養方 法としては一般的な液体振盪培養を行った後、iturin の高生産が見込める固体培養で行い、 B. subtilis RB14 の培養物を利用した植物病原菌からの感染防除試験などを行った。
60 3-2 材料および方法
3-2-1 微生物
Iturin A を生産し、植物病原菌に抑制を示す微生物としてB. subtilis RB14 を、植物病原
菌 と し て Bipolaris oryzae、Fusarium oxysporum、Monilinia fructigena、Phytophthora
infestans、Colletotrichum echinochloae、Fusarium solani、Penicillium digitatum、Rhizoctonia
solaniを使用した。B. subtilis RB14 は、Trypto-Soya Agar (TSA)培地 (2.0% polypeptone、
0.25% glucose、0.25% K2HPO4、0.5% NaCl、1.5 % agar) にて保存した。植物病原菌の保
存は Potato Dextrose Agar(PDA)培地 (2.4% potato dextrose broth、1.5% agar) で行い 25℃
で培養した。
3-2-2 残渣の調整
J. curcas種子の外殻を除去し HIGH-FLEX HOMOGENIZER(SMT)を用いて約 30 秒間
18,000 rpm にて破砕した。破砕した種子重量の 5 倍量のn-hexane を加え懸濁し、一晩以上 静置した。上清をろ紙 (No.5A 110 nm, ADVANTEC)を用いて吸引濾過し、濾液を減圧留 去した。n-hexane 抽出後の残渣に acetone を用いて同様の操作を行い、中性脂質を除去し た。溶媒抽出後の残渣を乾燥させ、ふるいにて均等な大きさにしたものを種子採油残渣とし て各実験に使用した。また、残渣は使用前に乾燥機(55℃)で乾燥処理を行った。 3-2-3 微生物の培養
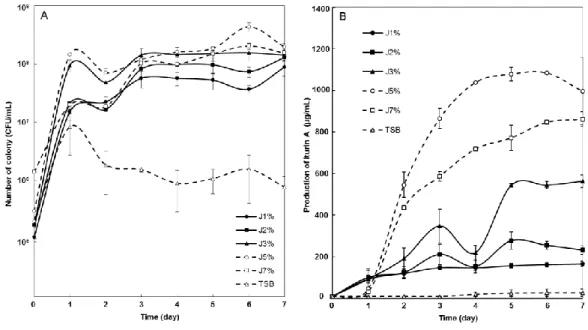
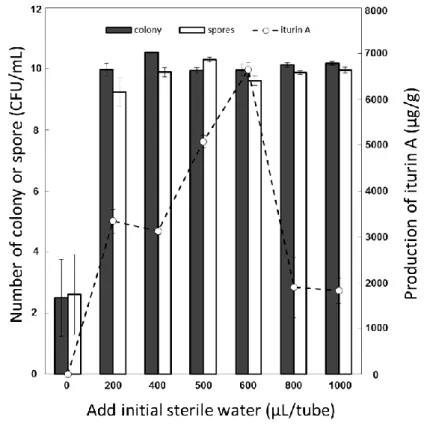
61 30℃、200 rpm で 16 時間振盪培養を行った。培養後 8,000×gで 15 分間遠心分離し、上清 を捨て等量の滅菌水を加え撹拌したものを各実験に使用した。 3-2-4 残渣懸濁液体培地を用いた RB14 の培養 100 mL フラスコに 20 mL の蒸留水とJ. curcas残渣 1 %, 2 %, 3 %, 5 %, 7 %(w/v)をそれ ぞれ混合し、121℃、15 分間オートクレーブ処理を行った。作製したジャトロファ液体培地 に培養液を 1 %植菌し、30℃、200 rpm にて振盪培養を行い、生菌数および iturin A 生産の 経時変化を測定した。 3-2-5 固体発酵培養における RB14 の培養 試験管に採油残渣を 0.2 g 入れ 121 ℃、15 分間のオートクレーブ処理を行った。水を除 去するため乾燥機(55℃)にて一晩乾燥させた。残渣培地に滅菌水をそれぞれ 0, 200, 500, 400, 600, 800, 1000 µL 加え、固体発酵培地を作製した。培養液を 20 µL 加えて、室温(24℃)、 静置にて 5 日間培養し、培養後に生菌数算定を行った。培養後の残渣培地に、培養前に加え た滅菌水との合計が 2 mL になるように滅菌水をそれぞれ加えよく攪拌した。得られた菌懸 濁液の生菌数算定を行った。 3-2-6 HPLC を用いた iturin A 分析 培養後の培養液および固体発酵残渣を用いて iturin A の抽出および HPLC 分析を行った。 培養液と等量の 35 %アセトニトリル水溶液を混合し、10 分間撹拌した。10,000×gにて 10
62
分間遠心を行い、得た上清をフィルター濾過(13JP, ADVANTEC)した。濾過後、上清を
HPLC に供し、iturin A 生産量を測定した。固体発酵残渣の抽出は、生菌数算定に使用した
培養液を用いて培養液と同様の方法で行った。HPLC は日本分光の LC-2000 システムを使
用した。カラムは逆相系 Chromolith Performance RP-18e (φ4.6 mm×100 mm)を使用し、
溶出溶媒は、pump A:10 mM CH3COONH4 、pump B:CH3CN を使用した。溶出は、流速
2 mL/min、カラムオーブン 40 ℃、検出波長 205 nm、分析時間は 10 分間 pump B 35%で 測定した。 3-2-7 抗菌活性物質の熱耐性 固体発酵残渣を用いて残渣に含まれる抗菌活性物質の熱耐性試験を行った。固体発酵残 渣を 121℃、2 分間のオートクレーブ処理を行った。その後、0.1 g を 1/10 PDA 培地と 1/10 TSA 培地の端から 1.5 cm に置いた。オートクレーブ未処理の固体発酵残渣と、未植菌の残 渣も同様に試験した。残渣を置いた後、培地中央にR. solani寒天断片を植菌した。24℃、 静置にて 5 日間培養し、植物病原菌に対する抑制能を測定した。 3-2-8 RB14 培養物を用いた植物病原菌抑制試験 RB14 固体発酵培養物が 20%(w/v)となるように PDA 培地を作製し、RB14 培養物含有 培地とした。培地の中心に前述の植物病原菌を植菌し、25℃で培養し、菌糸の成長を観察し た。