パッションフルーツ(Passiflora edulis Sims)の 花器及び果実の発育に関する研究
石 畑 清 武*
(1992年9月20日 受理)Studies on the Development of Flower Buds and Fruits in Purple Passion Fruit, Passiflora edulis Sims
Kiyotake ISHIHATA 目 次 第1章 緒 論 、 第2章 花芽分化及び発育 第1節 緒 百 第2節 新梢の生長と花芽の分化・発育 第3節 摘 要 第3章 花の形態と結果 第1節 緒 言 第2節 花型について 第3節 花型と花粉の発芽 第4節 花型と結果 第5節 摘 要 第4章 受粉及び受精 第1節 緒 百 第2節 人工培地上における花粉の発芽 第3節 柱頭上における花粉の発芽,花 粉管の伸長及び受精 第4節 摘 要
第1章 緒 論
第5章 人工受粉と果実の品質 第1節 緒 百 第2節 人工受粉と結果率の向上 第3節 人工受粉 第4節 摘 要 第6章 果実の発育 第1節 緒 百 第2節 果実の発育 第3節 仮種皮の発育と果実の品質 第4節 果実の形質と人工受粉による果 実の品質 第5節 摘 要 第7章 総 括 謝 辞 引用文献 Summary トケイソウ属(Passiflora)の多くは南米の熱帯,亜熱帯性気候の地域に分布しており,約600 種を越す14,26-28,46-51,55・73,128-130,144・145,160-162,164)といわれているが,これらのうち,キイロトケイソウ *指宿植物試験場(P.edulisSimsf.flavicarpaDeg.)とパッションフルーツ(P.edulis Sims)の2種類が世界で もっとも広い地域で栽培されている。キイロトケイソウは熱帯低地及び亜熱帯低緯度地方の低地で 栽培されている18,30,39,97,98,112,126,143,170)のに対して,パッションフルーツは熱帯低地では生育しにく く6,19,41,75,98,163)熱帯では標高500-3,000mの高地,亜熱帯,及び温帯の温暖な地域で栽培されて いる8,13,19,30,41,54,75,93,96,97,113,115・119,120,133,142,149,178)。我が国では,九州南部,高知県の海岸部及び東 京都八丈島においてパッションフルーツの露地栽培が可能である76,79,180)。 パッションフルーツの別名ムラサキクダモノトケイソウの名称は,成熟果の果皮が紫色であるこ とに由来するものである。我が国へ導入されたのは明治中期で33,86,142)第二次世界大戦終了頃ま では試験研究機関において標本植物として栽培されている程度であった。経済的な栽培は1949年 頃に鹿児島県で開始されたが,当初は青果利用を主目的としていたために栽培面積は僅かであっ た65)。ところが,果汁が種々の糖類16)ビタミン類131,132)ァミラーゼ20,132)香気成分76,92,109-111 のほかかノウムやカロチン94)数種のプロテアーゼを含有し43)トコフェロール生理活性が高い20) こと,さらに,種子には高濃度のリノール酸及び脂質95)大豆油様の油脂132)果皮にはペクチ ン131,134)等を含有することが明らかにされ,その果実の加工が注目され始め, 1964年には鹿児島県 指宿市にジュース加工工場が設立された。以来,ジュース加工を目的とした栽培が行われるように なり,次第に栽培面積が増加し, 1966年には主要生産地である指宿地方では年間の生産高が160 t に達した。 しかしながら,パッションフルーツの栽培にはブドウやキウイなどと同じく棚仕立てに多額の資 材費を要するため,主産地指宿地方においては温泉熱を利用して野菜や観葉植物などの施設栽培へ 作付け転換が行われ始め,パッションフルーツの栽培面積ならびに生産量は次第に減少していった。 その結果,ジュース加工業者は国内で十分な原料を確保することができなくなり,台湾やスリラン カ,ハワイ等で加工されたパッションフルーツの果汁を輸入し,再加工して販売するようになっ た1)。ところが,近年,食生活に対する要求が多様化し,風味や形状が特異な熱帯果実に対する晴 好が高まるにつれ,風味や品質が外国産より遥かに優れた国内産のパッションフルーツの果汁が再 び注目を浴びるようになった。 現在,九州南部,南西諸島及び沖縄県では青果及びジュース用としてパッションフルーツ,青果 用としてパッションフルーツとキイロトケイソウとの交配系(紅皮)の生産が増加しつつある。パッ ションフルーツは栽植後1年目から果実の収穫が可能なこともあって,奄美大島ではフライト農業 の基幹作目として育てようと;鹿児島県と地元が一体となって栽培に取り組んでいる。なお,キイ ロトケイソウはパッションフルーツに比べて耐寒性が弱いため栽培は極めて困難なようである。 このように,我が国の西南暖地においてパッションフルーツの栽培は盛んになりつつあるが,開 花期に降雨が多いことによる受粉障害や適当な花粉媒介昆虫がいないため,年によっては結果数が かなり少ないことがあり,作柄は安定していない。 本研究は,パッションフルーツの安定的な生産技術の確立に資する目的で, 1960年から行って きているものである。 「開花一受粉一結果」について生態,形態及び組織学的な面からその大要を 明らかにしたと思われるので,ここに成果をとりまとめて報告することにした。 本論文は7章よりなっている。第2章では花芽分化及び分化後の花芽発育を的確に把握するため に,年間の新梢の生育と花芽の発育との相互関係を明らかにした。第3章では開花期を通して開花 数の推移と花型の変異を気象との関連で経時的に調べ,次いで,第4章では受粉及び受精の実態を 調査し,開花時に花型の異なる3種類の花の柱頭上における花粉の発芽,及び花柱内への花粉管伸
長について明らかにし,第5章では人工受粉によって結果率及び果実品質が著しく向上することを 示した。第6章では受精後の果実の発育,とくに可食部である仮種皮の発達過程を経時的に調査す るとともに,品質についても言及した。なお,第7章ではこれらの結果を総括した。 第2章 花芽分化及び発育 第1節 緒 看 トケイソウ属の中で最も広く食用に利用されている種はパッションフルーツである。我が国では, 南九州の降霜の少ない地域から沖縄にかけて,主に果皮が紫色の系統が露地栽培されており,本研 究の実験・調査を行った鹿児島県指宿市では開花期は4月下旬から6月上匂(前期,夏実となる) と9月下旬から10月中旬(後期,冬実となる)の年2回である60,70)。オーストラリアのNewSouth Wales州, Western Australia州及びQueensland州南部,南アフリカのTransvaal及びNatal地 方では,夏実が主であるが冬実の収穫も出来ると報告されている8,106,107,120・176-178)。しかし,指宿 市では前期の初期に開花する花は自家稔性花の割合が高く60・65)品質の良い果実が得られるが,そ の後に分化する花菅は気温が高いために未発達のまま落下する。一方,冬実は完熟する前に低温に あうために収穫が不可能となる60,70)。 したがって,花芽の分化・発育についての解剖形態学的な知見は,結果調節の技術開発にとって 極めて重要であると考えられるが, Raiu138)のP.calcarataを材料とした配偶子形成と種子の発育 に関する研究以外に,トケイソウ属植物の花芽分化・発育については報告が見あたらない。 本章では,南九州の夏季高温時においてはパッションフルーツの正常な開花が行われない原因を 明らかにする目的で,とくに気温との関連で新梢の生長に伴う花芽の形成・発育について形態学的 及び解剖学的調査を行った。 第2節 新梢の生長と花芽の分化・発育 材料と方法 材料には鹿児島大学農学部附属農場指宿植物試験場の圃場に栽植された実生由来の3年生株を供 試し,調査は1977, 1978の両年に亘って行った。栽植密度は畦幅2m,株間1.5mとし, 1株当り 主枝4本,高さ2mの垣根仕立てで,側枝は無勢走とした。圃場の土性は壌土で,晴耕栽培であっ た。施肥は10∂当りN:P205:K20-16:16:16 の複合肥料(コスモホスカ,宇部化成株式会 社) 100kgと菜種油粕 の半量ずつを3月と6月の2回に分施した。その他の管理は当地方にお ける慣行法によった。 1)新梢の生長と花芽の着生 調査(1978年)には,標準的な生育をしている5株を選び,各棟から伸長させた4本の主枝新 梢のうちの1本について,休眠覚醒後の3月下旬から経時的に新梢の生長,花芽の発育及び落書の 調査を行った。なお,花芽についての調査は,新梢先端部の刺葉菜臓に発達する花芽が拡大鏡で確 認できる大きさ(花芽長:約0.05耽7n)に達するまでは,実体顕微鏡を用いて行った。
2)花芽の分化・発育 観察には, 1977年12月30日から1978年3月27日までは1977年に生長した新梢の基部第6節 から上の葉臓に着生した長さl-2mの側芽(側杖)を, 1978年4月1日から5月21日までは休眠 覚醒後の3月下旬より生長した新梢(側杖)の基部から第5-6節位に着生した花芽(新梢の第1 -4節位にはほとんどみられない)を用いた。花芽は 10個について剥離法により光学顕微鏡 下で調査した。 3)花芽の発育経過と到花日数 1983年4月19日に, 2 日後に開花すると思われる花芽が着生している新梢20枝を採取し, 各葉肢に着生する花芽の形状を調査した。その後,基部から第5-6節までの花芽はFAA フォ ルマリン:酢酸:アルコール:水-5:5:45:45%の混合液)で,第6-7節から茎頂までの花 芽はリン酸緩衝液(pH:7.2)に溶かした4%グルタルアルデヒドで固定し,それぞれ光学顕微鏡 及び走査型電子顕微鏡(SEM)による観察材料とした。なお, FAAで固定した材料は常法によ り厚さ12-14vmのパラフィン連続切片を作成し,デラフイールドのへマトキシリンで染色して永 久プレパラートを作成し鏡検した。一方,グルタルアルデヒドで固定した材料は,エタノールシリー ズで脱水した後,酢酸イソアミルによる臨界点乾燥(HITACHI, HCP-1)を行った。それを金属 片にド-タイトで接着し,イオンスバッター(日本電子製JHC-1,100)による金蒸着を行い,走 査型電子顕微鏡(日本電子製JSM-15型)を用い加圧電圧15AVで観察した。 なお,本実験の観察に供したと同じ伸長程度の新梢を選び,各葉肢に着生している花芽が開花に 到るまでの所要日数を調査した。 結 果 1)新梢の生長及び花芽の着生 新梢の伸長状況と葉臓に着生する花芽の発育及び実験期間中における日最低気温及び日最高気温 の変化を第1・表及び第1図に示した。 新梢の伸長は日平均気温(最低気温と最高気温の平均値)が15℃前後に達した4月上旬から始 まり,気温の上昇につれて旺盛となったが,日平均気温が25℃以上になった6月上旬以降は非常 に緩慢となった。さらに,日平均気温が25℃以下に低下した8月下旬より再び伸長したが, 11月 以降は伸長が緩慢となり,日平均気温が13℃以下に低下した12月中旬には伸長は完全に停止した。 花芽は各葉臓に1個着生しており(Plate I-1, 2, 3),新梢の基部から第5-7節の菓肢に着生 した花芽は5月下旬から6月上旬にかけて順次発育し開花した(第1図)。第8-9節から上位節 の葉臓に着生した花芽は,包葉長が0.5-9mro 花弁形成期∼子房形成期)に発育した後すべて落 下した。新梢の生長が非常に緩慢となった7-8月においても,乗数の増加が認められた枝梢の葉 肢には花芽が分化・着生したが,包葉長が1mにも達しないうちにすべて落下した。 12月上旬にな ると気温の低下とともに地温が低下し(第1表),新梢のみならず葉芽及び花芽の生長は完全に停 止し(第1図, 12月中旬以降は降霜のために花芽は凍害を受けて枯死した。 2)花芽の分化・発育 上述したように,露地栽培されている実生由来の3年生株について新梢の生長及び花芽の着生・ 開花について調査した結果では,花芽が正常に発育し開花・結果するのは,指宿地方では新梢の基 部から数えて第5-7節に着生した花芽に限られることが確認された。そこで,基部から第5-6
第1表 実験期間中の実験地の気象
Table 1. Climatic conditions of the experimental field Air temperature
Max. Mean Mm.
Soil temperature
Year Month Decade
In depth 5cm 1 0cm 暮し S 0 r F 1977 November Early Middle Late December Early Middle Late 1978 January Early Middle Late February Early Middle Late March e s cd ℃ ℃ 25.7 20.8 21.4 17.8 17.2 13.1 3 3 4 8 8 5 1 1 1 4 4 1 3 3 4 = ‖リ = 1 6 6 4 1 6 日 日 印 : 4 3 3 6 5 8 1 1 1 2 1 3 ● ● 2 4 1 1 1 1 4 3 9 ● ● 9 9 8 0 1 9 ● 9 8 9 9 5 2 ● 1 0 3 1 1 1 ℃ O > C ^ c T > O O r -i i f i N ^ O C 」 > O Q 。 0 0 0 . 0 L O ^ O O C D O > t > -L O L O ^ ^ ^ O O N I T ) 0 0 日 リ 日 印 ℃ ℃ 19.6 19.6 17.5 17.6 13.3 13.3 4 3 0 1 3 1 1 1 1 3 6 7 ● 0 9 8 1 1 8 7 9 8 8 2 2 9 ● 一 1 0 1 日 印 ‖ リ 日 日 4 3 0 1 3 1 1 1 1 1 8 7 0 9 8 1 0 8 6 ● 9 8 8 1 2 7 ● ● ● 1 0 1 日 H H コ 臼 y t i O O H ^ H O N C O C O C O C 」 > L O C 」 > O J -I < N l d 0 2 ( j ) a a n ^ B a a d E 8 1 -a y a a q E n u a p o v ( 1 1 1 0 ) m S u d ¥ j o o u c 0 0 O 5 日H
Mar. Apr. May. Jun. Jul. Aug. Sept. Oct. ぐ Month
第1図 パッションフルーツの新梢の伸長,花芽の発育及びそれらの消長並びに実験地の 気温1978.
Fig. 1. Shoot elongation, flower buds development and the abscission with days elapsed after bud differentiation in purple passion fruit, and air
tempera-●
ture of experimental held during March to December, 1978.
●
節に着生する花芽の発育について, 1978年3月2日から同年5月21日まで5日置きに調査を行っ た(第2表)。 新梢の生長は3月中旬より始まったが, 3月17日に採取した材料で包葉初生突起の形成が認めら れた(Plate 1-4, 5)。この時期が花芽分化期と思われる。 3月22日の材料では包葉(Plate 1-6), 3月27日の材料では花芽に等片の形成が認められたが Plate 1-7),この材料(基部より第5-6節)より上位節の菓臓に着生する材料では包葉突起が認められた。 4月1日の花芽では等片はか なり発育しており,専片の内側基部に花弁の形成が認められた(Plate 1-8)。 4月6日には等片 の内側に荊初生突起が(Plate 1-9), 4月11日には約の内側に雌蕊初生突起が認められた(Plate
n-i 。 4月16日の花芽は節及び雌蕊の形成期(Plate n-2), 4月21日の花芽は花柱形成期(Plate n-3)で,花弁基部の内側に副花冠初生突起が認められた(Platen-4)。 4月26日の花芽では子 房は形成期にあり,肱珠初生突起(PlateE-5)及び薪には4室の蔚胞(Plate E-6)が,専片の 内側には蜜腺(PlateH-7),花粉母細胞(Platen-8,9)の形成が認められた。 5月1日の花芽は 肱珠形成期で(Plate m-1, 2),薪胞内には械畿組織から分離した花粉四分子が観察された Plate Ⅲ-3,4 。 5月6日には肱珠は倒生状態に発育し(倒生肱珠),内種皮,珠心,肱嚢,合点及び珠 孔形成が認められた(Platem-5)。また,紡胞内には発育した小胞子(花粉粒)が認められた (Platem-6, 7)。 5月11冒には肱珠は発育肥大し,荊胞の相接する基部の内皮と械畿組織は崩壊 第2表 指宿地方におけるパッションフルーツの花芽の分化及び発育栓過(新梢の基部から第 5-6節位,各日5個の花芽を調査)
Table 2. Differentiation and development of flower buds at 5th or 6th node from the base of the new shoots in purple passion fruit. Five flower buds were observ-ed at each date
Month March April May
Date 7 17 22 27 1 6 ll 16 21 26 1 6 11 16 21 Undifferentiation stage Bracterial leaf formation stage Sepal differentiation stage Sepal formation stage Petal formation stage Stamen formation stage Pistil formation stage Corona formation stage Ovary formation stage Ovule formation stage Flowering 5 5 5 1 4 1 4 1 4 1 2 1 3 1 2 2 3 1 4 4 1 5 5 5 5
し,完成間近の花粉が認められた(Platem-8)。 5月16日には等片長は包葉長の約1.5倍に,花 柱長は子房縦径とほぼ同じ長さで完成期の約70%の大きさに,肱珠は縦径0.3-0.4i雄に,子房は 5.0-5.5ototに発育した。また,半荊(荊室)が形成され,副花冠の長さはほぼ13-14mに発育し, 基部側には紫斑点が観察された(PlateHI-9)。 5月20日の花芽では包葉,専片,花弁,花柱,花 糸ともに発育を終了しており,肱珠は縦径0.55-0.60**,子房は縦径5.5-6.0耽7nに発達していた (PlateIV-1,2, 3)。夕刻には専片頂部が少し開き始め,翌日(5月21日)には完全に開花した。 以上のように,花芽原基では最も外側に位置する包葉の初生突起が隆起し,この形成が進行する 過程でその内側に専片初生突起が形成され,尊片の発育過程でその内側基部に花弁初生突起が形成 された。さらに,花弁の発育の過程で内側基部に雄蕊初生突起が,雄蕊の発育過程でその内側基部 に雌蕊突起が形成されて発育した。つまり,各器官は外側から求心的に連続して形成されたが,副 花冠初生突起のみは雄蕊初生突起や雌蕊初生突起より遅れて花弁基部に形成された。一方,花芽に 隣接して形成された側枝の原基は次の開花期(9月下旬∼10月)または翌春まで生長しなかった。 3)花芽の発育経過と到花日数 第2図には1月より7月の調査期間中の日最高気温と日最低気温の変化を示した。このような温 度条件下における花芽の発育程度(縦径及び横径)と到花所要日数の間には,第3図及び第4図に みられるような負の相関関係がある。到花所要日数(Y)は花芽の縦径(XL)及び横径(Xw と の間には,それぞれ Y -1.73XL2-14.38XL+32.43及び Y -7.56Xw -32.37Xw+36.09 の式で推定が可能である。 新梢の生長が始まって間もない3月中旬,開花約60日前に,茎頂(PlateIV-4 から基部側へ5 -6節下位の葉臓に,花芽原基または包菓形成初期の花芽が認められた(PlateIV-5, 6)。この花 芽は開花約5週間前には縦径約0.2mmに発育し,草片及び花弁が発達した内側には薪が発育し,円 Jan. Feb. Mar. Apr. May. Jun. Jul. Month 第2図 実験地の1月から7月の気温1983.
5 0 ll SuTJ8MOTJ O一sAvn 0.4 0.8 1.2 1.6 2.0 2.4 2.8 3.2 3.6 4.0 4.4 Length of flower buds {CM)
第3図 パッションフルーツの花芽縦径と到花日数(1983.
Fig. 3. Relationship between the length of the flower buds and the
● ●
number of days to flowering in purple passion fruit, 1983.
筒状の柱頭が観察された(PlateIV-7)。開花約4週間前になると,柱頭は発育して内側に浅い溝 が形成されており PlateIV-8),荊胞内には械壌組織が認められ,花粉母細胞は移動期で(Plate IV-9, PlateV-1),肱珠原基が認められた PlateV-2)。開花約3週間前の花芽では縦径1.2-1.4cm,横径0.5-0.7cmに生長し,珠柄,内珠皮,外珠皮及び肱嚢母細胞 PlateV-3)が,薪胞 内には同時膜形成の花粉四分子(PlateV-4)などが認められた。開花約2週間前には花芽は縦径 1.3-1.7cm,横径0.8-1.lcmに生長し,荊胞内には織機組織の崩壊及び花粉粒の肥大が認められた (PlateV-5)。開花約1週間前の花芽は縦径2.0-3.0cm,横径1.1-1.5cmに生長し,肱珠は形成終 期であった(PlateV-6)。開花2-3日前になると花芽は縦径3.1-4.0cm,横径1.5-2.Ocwiにな り,旺珠,花糸(PlateV-7,8),節(PlateV-9 ,柱頭(PlateVI-1),花粉(PlateVI-2)な どは形成終期であった。開花前日には花芽は縦径4.5-5.0cm,横径2.3-2.5cmになり,翌日開花し た(PlateVI-3)。 考 察 本実験を行った指宿市では,パッションフルーツは3月中旬頃から新梢の伸長生長を始めるが,
S u T J 9 M O T J O ; S X B Q 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 Width of flower buds (cm)
第4図 パッションフルーツの花芽横径と到花日数(1983).
Fig. 4. Relationship between the width of the flower buds and the number of days to flowering in purple passion fruit, 1983.
● 当地の気象条件下では12月上旬の低温に遭遇するまで無限生長を行い,花芽は健全な充実した新 梢の基部から第5-6節以上の各葉臓に1個ずつ分化した。開花は4月下旬頃から始まったが, 7 -8月以降は高温のため花菅はすべて開花前に落下した。年によっては, 9-10月になり気温が 低下すると開花する花がみられるが,開花し,結果しても,充分に成熟した果実を収穫することは 不可能であった。 パッションフルーツの収穫を春期と秋期の2回行っているオーストラリアのQueensland州南 部177,178)においては,開花期は8-9月と3-4月の2シーズンで,平均気温はそれぞれ16.8-21.9℃と21.2-23.6℃である165)。指宿市における開花期は4-6月と9-10月の2シーズンで平 均気温は16.3-23.3℃と20.4-25.6℃であり64)y指宿市の気温はQueensland州南部の気温にほぼ 類似している。なお,奄美大島における開花期の3月下旬∼5月上旬は平均気温が17.2-22.3℃で, 指宿市の4-6月の気温にほぼ類似している。 Simon149'によれば,地温: 25℃,昼一夜の気温: 30-20℃-25℃では, 25-15℃-20℃におけ るより着花数が著しく少ないとされており,宇都宮169)は花芽分化には昼一夜の温度が33.8±1.1-24.5±0.5℃より23.5±0.8 -16.1±0.2℃が適し,高温下では着花数が減少すると報告している。 従って,本試験を行った指宿市において9-10月期に開花数が少なかったのは,花芽の発達期の
気温がやや高かったためであり,開花・結果しても収穫に到らなかったのは,結果後の気温の低 下65)によるものであろう。なお,著者ら61,68,69が実生苗の栄養生長に及ぼす地温の影響をガラス 温室内で調査した結果によれば,生長は地温20℃の場合が最も旺盛であった。 南九州に普及している主要な果樹の花芽分化から開花までの所要日数は以下のとおりである。ま ず落葉果樹のブドウは5月中・下旬117)から6月下旬32,34)ヵキは7月下旬に花芽を分化するが, 花器の形成は翌春に新梢が生長を開始するまでは行われず,新梢の生長過程で花芽は急速に発育す る44,103。結局,花芽が分化してから開花まで9カ月以上を要するとされている。ナシでは6月下 旬∼7月上旬に花芽を分化するが11,12,81)花器は翌春までの間に発達して完成する。クリは8月上 旬に花芽を分化し,花器は翌年の5月上旬までに完成し99-101)モモは7月下旬∼8月上旬に花芽 分化するが31,32)花器は秋に完成し翌春に開花するので,ナシ,クリ及びモモでは花芽の分化から 開花まで8カ月以上を要することになる。常緑果樹では,ウンシュウミカンは8月頃に茎頂の変化 が観測できるといわれているが56,105)実質的には1月下旬に花芽分化を開始して約100日後の5 月上∼中旬に開花し71,104,123)ビワは7月に花芽を分化し90-100日後に開花する122)。また,蔓性 のキウイは3月中旬に花芽形成が始まり,約2カ月後の5月中∼下旬に花器を完成して開花すると 報告されている173)。なお,ナシ11,12)では花芽形成には新梢の生長の停止が必要とされているが, 柑橘類ではシキキツにみられるように年数回着花する系統が知られており25,139)種や品種によっ て花芽形成に必ずしも生長の停止が必要ではない141)。 本実験の結果によれば,パッションフルーツは新梢の生長につれて3月下旬に茎頂近くの菓臓に 花芽を分化するが,花器の完全な発育は温度条件によって影響されることが認められた。 温度条件が好適であれば,花芽分化後花の各器官は求心的に連続して発達し,約60日後に開花 するので,花芽原基の形成から花器の完成・開花までの所要期間は,南九州地方で普通に栽培され ている木本性の果樹より著しく短く,キウイとほぼ同程度であるとみなされる。なお,パッション フルーツの花芽の分化・発育過程は,雌蕊初生突起分化後に花弁の基部に副冠の初生突起を分化す るという点を除けば,キュウリにおける花芽の分化・発育36)と極めて類似しているようである。 第3節 摘 要 パッションフルーツの南九州における花芽分化及び花芽の発育,開花に至る過程を明らかにする 目的で形態学的及び解剖学的・な調査観察を行った。 1.花芽は新梢の伸長に伴ない,茎頂近くの葉臓に順次分化する。花芽の器官形成は求心的に行わ れ,発育過程はキュウリの場合と非常に類似していた。 2.花芽形成は生長が停止する冬季を除き常時行われるが,高温長日の夏季には枝梢の生長,花芽 の発育ともに抑制され,花芽は縦径1.0耽7n位の太さに発育すると落下した。花芽の分化及び発育 の適温は13-25℃のようである。 3. 月に開花する花芽は3月下旬以降に分化し,約60日で花器を完成したものである。発 育中の花芽の縦径(XL)及び横径(X,と開花所要日数(Y)との間には,それぞれ負の相関が 認められ Y-1.73XL -14.38XL+32.43, Y-7.56Xw -32.37Xw+36.09の式が得られ た。 4.開花25日位前は花粉母細胞と肱珠形成始期, 14日位前は花粉,肱嚢及び珠心形成期, 7日位 前は肱珠及び花粉形成終期で,開花前日に花器は完成した。
Plate I
l 新梢の頂点近くの花芽. ×1.3.
Flower buds near the apex of shoot. Leaves are removed. ×1.3.
2 頂点近くの花芽(Plateト1の拡大). ×3.5.
Magnified version of plateト1 showing the flower bud near apex. ×3.5. 3 花芽と側枝. ×3.5.
Flower bud and lateral shoot. ×3.5.
4 生長点(矢印,縦断図). ×70.
Longitudinal section of growing point (arrow). ×70.
5 3月22日,包葉の初生突起形成(花芽分化,縦断図). ×44.
Flower bud differentiation, Longitudinal section of bract primordia formed on March 17. ×44.
6 3月22日,包菓形成期(縦断図). ×58.
Longitudinal section of bract formation stage on March 22. ×58.
7 3月27日,尊初生突起の形成(縦断図). ×58.
Longitudinal section of calyx primordia on March 27. ×58.
8 4月1日,花弁の初生突起形成(縦断図). ×25.
Longitudinal section of petal primordia on April 1. ×25.
9 4月6日,薪の初生突起(縦断図). ×18.
Plate n
1 4月11日,雌蕊初生突起の形成(縦断図). ×16.
Longitudinal section of pistil primordia on April ll. ×16.
2 4月16日,雌蕊及び荊形成期(縦断図). ×13.
Longitudinal section of pistil and anther formation stage on April 16. ×13.
3 4月21日,花柱伸長期(縦断図). ×10.
Longitudinal section of style elongation stage on April 21. ×10.
4 4月21日,子房形成及び副花冠初生突起形成(縦断図). ×5.
Longitudinal section of ovary formation and corona primordia on April 21. ×5.
5 4月26日,肱珠初生突起形成(縦断図). ×31.
Longitudinal section of ovule primordia on April 26. ×31.
6 4月26日,花粉嚢形成(横断図). ×5.
Cross section of pollen sac formation on April 26. ×5.
7 4月26日,蜜線形成(縦断図). ×13.
Longitudinal section of nectary formation on April 26. ×13.
8 花粉嚢内の花粉母細胞(PlateH-6の拡大). ×48.
Magnified version of plateII-6, showing pollen mother cells in pollen sac in
cross section. ×48.
9 4月26日,花粉母細胞(横断図). ×160.
Plate IE
1 5月1日,肱珠形成期(横断図). ×13.
Cross section of ovule formation stage on May 1. ×13.
2 肱珠形成期(Plate ffl-1の拡大). ×66.
Magnified showing of ovules (plate DI-1). ×66.
3 5月1日,械壌組織と花粉四分子(横断図). ×18.
Cross section of tapetum and pollen tetrad on May 1. ×18.
4 花粉四分子(PlateEl-3の拡大). ×146.
Magnified pollen tetrad (plate m-3). ×146.
5 5月6日,倒生肱珠(縦断図). ×110.
Longitudinal section of anatropous ovule on May 6. ×no.
6 5月6日,節,花粉嚢及び花粉(横断図). ×13.
Cross section of anther, pollen sac and pollen grains developed on May 6. ×13.
7 花粉(Platem-6の拡大). ×115.
Magnified pollen in plate IH-G. ×115.
8 5月11日,花粉嚢内の崩れた械壌組織と成熟間近の花粉(横断図). ×22.
Disrupted tapetum and almost matured pollen grains were found in pollen sac on May ll (cross section). ×22.
9 5月16日,花芽(縦断図). ×0.8.
Plate IV
1 5月20日,開花前日の花芽(縦断図). ×0.7.
Longitudinal section of a flower bud on May 20, one day before flowering. ×0.7.
2 5月20日,開花前日の肱珠. ×13.
0vules at one day before flowering. ×13.
3 開花前日の肱珠(PlateIV-2の拡大). ×35.
An ovule magnified from plate IV-2. × 35.
4 新梢頂部(走査型電子顕微鏡写真). ×66.
Shoot apex (SEM). ×66.
5 新梢頂部近くの花芽(走査型電子顕微鏡写真). ×9.
Flower bud formation near shoot apex (SEM). ×9.
6 新梢頂部近くの花芽(走査型電子顕微鏡写真). ×31.
Flower bud near shoot apex (SEM). ×31.
7 開花約5週間前の花芽(走査型電子顕微鏡写真). ×87.
Flower bud about 5 weeks before flowering (SEM). ×87.
8 開花24-26日前の花芽の柱頭,子房及び荊(走査型電子顕微鏡写真). ×35.
Flower bud 24 to 26 days before flowering, showing stigma, ovary and anther (SEM). ×35.
9 開花24-26日前デイアキネス期の花粉母細胞(横断図). ×50.
Cross section of pollen mother cell at diakinensis stage 24 to 26 days before flower-ing. ×50.
Plate V l 花粉母細胞(PlateIV-9の拡大). ×100.
Pollen mother cells magnified from plate IV-9・ ×100.
2 開花24-26日前の肱珠初生突起(縦断図). ×87.
Longitudinal section of ovule primordia 24 to 26 days before flowering. ×87・
3 開花約3週間前,肱嚢母細胞形成期(縦断図). ×115.
Longitudinal section of embryo sac mother cell formation stage about 3 weeks before flowering. ×115.
4 開花約3週間前,花粉嚢内の花粉四分子(横断図). ×142.
Cross section of pollen tetrads in pollen sac about 3 weeks before flowering. ×142.
5 開花約2週間前,花粉嚢内の花粉粒(横断図). ×115.
Cross section of pollen grains in pollen sac 2 weeks before flowering. ×115.
6 開花約1週間前,発育終期に近い肱珠(縦断図). ×44.
0vule about 1 week before flowering, showing longitudinal section of ovule near final formation stage. ×44.
7 開花数日前,花糸の上部(走査型電子顕微鏡写真). ×7.
Upper portion of filament a few days before flowering (SEM). ×7・
8 花糸と荊接合部(Plate V-7の拡大,走査型電子顕微鏡写真). ×13.
Filament connected with anther magnified from plate V-7. ×13・
9 開花数日前の荊(走査型電子顕微鏡写真). ×4.
Plate VI
l 開花数日前の柱頭(走査型電子顕微鏡写真). ×5.
Capitate stigma a few days before flowering (SEM). ×5.
2 開花数日前の花粉粒(走査型電子顕微鏡写真). ×93.
Pollen grains a few days before flowering (SEM). ×93.
3 開花(当日). ×0.3.
Flowering, showing 5 calyxes, corona, 3 stigmas and one ovary. ×0.3.
略語の説明(Plate I-VI)
an:前, b:包葉, C:尊, co:副花冠, emc:肱嚢母細胞, es:旺嚢, f:珠柄, fb:花芽, fi:花糸, g:子房柄> 9P:生長点, ii:内珠皮, Is:側枝, m:珠 孔, ∩:珠心, nee:蜜腺, nuc:核, o:子房, oi:外珠皮, ovu:肱珠, p: 雌蕊, pe:花弁, pm:花粉母細胞, po:花粉, ps:花粉嚢, pte:花粉四分子, S:柱頭, ta:械犠組織
Abbreviation:
an:anther, b:bract, c:calyx, co:corona, emcembryo sac mother cell, es:embryo sac, f:funiculus, fb:flower bud, fkfilament, g:gynophore, gp:growing point, ii:inner integument, ls:lateral shoot, m:micropyle, n:nucellus, nee:nectary, nuc:nucleus, o:ovary, oi:outer integument, ovu:ovule, p:pistil, pe:petal, pm:pollen mother cell, po:pollen, ps: pollen sac, pte:pollen tetrad, s:stigma, ta:tapetum.
第3章 花の形態と結果 第1節 緒 看 パッションフルーツは自家稔性植物とされているが3,76,112,114,145)放任状態では結果率が非常に 低い58,176・180)。このため栽培にあたっては結果率を高めるために人工受粉が行われているが181)結 果率は天候の影響を受けやすく,晴天時は90.6%,雨天時は6.7%という例が報告されている60)。 また,開花期の後半(5月中旬∼6月上旬)は前半(4月下旬∼5月上旬)に比べ結果率が低下し, その原因は,開花時の花の内的あるいは気象的要因によるものと推察される。 パッションフルーツの花柱は3本に分岐し,開花時には外側に曲り,ほぼ水平に展開した花糸の 先端で下向きになった薪と薪との間に位置している。ところが,花柱の形や花柱と薪の位置関係な どが異なる花が知られており,これら異なる形態の花の形成は気温や日長などの外的要因と関係が あると推察されている175。 本章では,形態(花型)が異なる花の形成とそれらの花における花粉発芽及び結果との関係を開 花期の気温との関連で究明した。
第2節 花型について 材料と方法 材料には指宿植物試験場内の垣根仕立て栽培の実生3 - 5年生株を供用した。 まず, 1978年5月12日と17日の両日,午前5時から午後9時までの間に開花した花について, 花器の展開状況を中心に経時的な観察を行った。次いで, 1979年4月22日から5月31日にかけ て,午後2時から3時に開花している花の花型を調べた。そして,各花型から30花ずつを採取し, 花器の長さと幅,子房の長径及び短径,柱頭の長径及び短径などを調査した。さらに, 1982年5 月1日から6月4日にかけて,開花前日の午後から翌日までの気温と開所時刻を調査した。材料は 1日20花宛供した。なお,長さ,幅,長径,及び横径の測定にはノギス(1/lOOraJI走可能)を用 いた。 結 果 膏は日の出とほぼ同時刻に等片と花弁が開き始め,まもなく開張し開花した。その後,花糸は花 床に水平に開き,薪は下向きになり開所した(第5図)。一方,花柱は大部分の花では開花後外側 へ水平に開くとともに薪の方-琴曲し,やがて柱頭が薪に接近または接触したが,一部の花では花 柱は直立していた。このように,柱頭が薪に接する接触型すなわち正常花(normal style type)
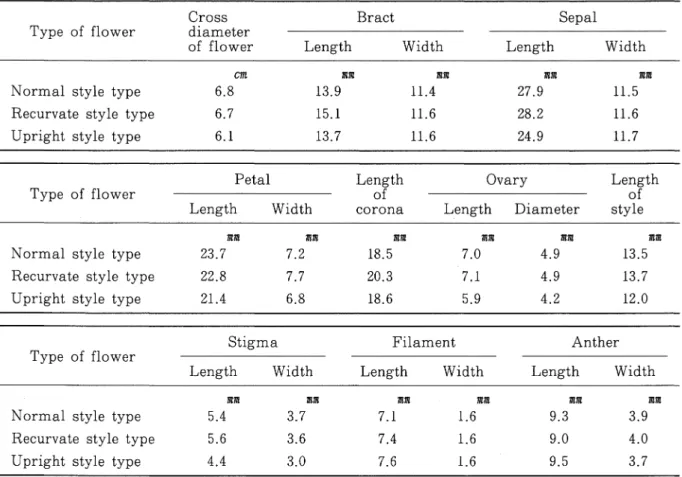
のほかに,柱頭が薪の位置まで達せず薪の上方に位置する接近型(recurvate style type),花糸 は前二者と同様に展開するが花柱が琴曲しない直立型(upright style type)の3型が見出された (第6図)。ところが,いずれの型の花においても等片,花弁,副花冠(corona)6,13,39,121)花糸及び
第5図 パッションフルーツの花の形態. ×1.0.
Fig. 5. Floral organs of purple passion fruit. ×1.0.
Left: face view, right: longitudinal section, a: anther, c: calyx, co: corona, f: filaments, g: gynophore, n: nectary, o: ovary, p: petal, s: stigma.
第6図 パッションフルーツの花型. ×0.7.
Fig. 6. Three flower types in purple passion fruit. ×0.7. Left: normal style type, center: recurvate style type, right: upright style type.
花柱は夕刻には立ち上り,夜半にかけて開花し,翌日再び開花することはなかった。なお,直立型 においても花糸と薪は接触型及び接近型のそれらと同様に正常に展開して裂開したが,花柱は開花 時から開花時まで直立したままであった(第6図)。 第3表は開花日における各花型の出現状況を経時的に調べた結果である。直立型は日中の気温 が24-25℃であった1978年5月12日, 21.5-21.?℃の5月17日の両日とも午前7時に認められ, 出現率は3.4%と7.1%で大差はなく,しかも終日(7:00-21:00)一定であった。一方,接触型と 接近型の出現率は経時的に変動したが,両者の合計数は1日中一定であった。すなわち,両日とも 接近型は直立型と同じく午前7時に出現したが,接触型は9時に初めて出現した。その後は,接触 型の数だけ接近型が減少し,気温が高かった5月12日には午後1時に,気温がやや低かった5月 17日には午後3時に接触型の出現率は最高に達し,その後は減少に転じ,両日とも午後9時には 接触型は認められなくなった。このように,接触型は接近型と可逆的に変化する花型で,接近型に は潜在的に接触型が含まれていることが明らかになった。 次に,毎日定時(午後2時から3時までの間)に調査した日別総開花数と花型の出現率の推移を 第7図に,また,期間中の気温を第8図に示した。調査期間中に開花した総花数は6,053花で,そ れらは接触型が76%,接近型が17%,直立型が7%であった。 4月下旬の開花期初期から5月中 旬の開花期中期にかけては接触型の出現率が高く, 5月下旬の開花期後期には接近型及び直立型の 出現率の増加がみられた。 花の形態的調査結果では,第4表のように接触型と接近型とは形態的差異は認めにくいが,直 立型は接触型及び接近型に比べ花の直径,琴片及び花弁の長さが小さく,特に雌蕊の花柱長,柱頭 の長径及び短径とも著しく小さかった。子房の長径では接触型花の7.0mm,接近型花7.1mに比較し て直立型は5.9耽7nとやや小さいようである。 3花型間の雄蕊には形態的差異は認められなかった。
第3表 パッションフルーツの開花日における各花型花出現の経時的推移
Table 3. Changes of the flower types with the time elapsed after flowering in purple passion fruit
May 12, 1978
Number of flowers observed
Time Air
temperature Weather Normal Recurvate Upright
style style style type type type
ooooooooooo oooooooooo0 ..t mo-as(MCO^l^サtSOii-i- ℃ 14.0 lightly cloudy 17.6 lightly cloudy 22.5 lightly cloudy cloudy cloudy cloudy cloudy cloudy cloudy cloudy cloudy H I D N O O N C D ^ O O C 」 > L O I > - t D O O O O O O < L O e ^ C O 日日 1 1 1 O N CO !N 10 5D ^ 05 a) (D (M (N i -i O ^ L O C O C S I O O ^ C O O ' -I i -I O ^ ^ ^ " ^ ^ ^ . ^ J * " * -r H ^ H ^ * May 17, 1978
Number of flowers observed
Time Air
temperature Weather
Normal Recurvate Upright
style style style type type type
oO CDOoooooo ooooooocti ・4-3 LDNO5NCO^LONOii-i- ( D C D C D C D C D C D C D C D C D C D C D G G C I S a c J c l c i c G C ・l ・l ・l ・l ・l ・l ・l ・l ・l ・l ・1 ー ー ー ー ー ー f ー ー ー ー 1 1 O O CO LO OO l>- 0 00 *-I *-H O CO -t co oo r-i -h ct> -h
・- I l - I l - I '- I
: : ‖ コ
o o ^f Ln i^ oo o t>- a> a> o o
N tO Ln O OO tD in iS LD N N o o o o o o o c o o o o o o o o o o o o o o o l 1 1 1 1 1 1 1 1 1 1 1982年5月1日から∼6月4日までの気温と開筋時刻との関係を第5表に示した。この期間 における接近型及び直立型の平均の開所時刻は午前7時57分± 1時間29分で,両花型間に差はみ られなかった。開花前日の午後,とくに, 12時,午後4時,開花当日の4時, 6時の気温と関前 時刻との間には負の相関が認められ,前日の気温及び当日朝の気温が高いほど関前の時刻は早まる 傾向がみられた(第6表)。
s j 9 M o n j o j a q u i n j s j 22 25 April 15 20 25 30 0 0 0 0 8 6 1 0 0 4 2 0 s a d X ; j a M o j i i p e a l 0 9 S b ; u 9 d j o c j Date 第7図 実験地における各花型花の着生率の推移1979).
Fig. 7. Number of flowers and percentage of each flower type in the field, 1979.
j o ) a j n ; p j a d u i a T 22 25 April 30 10 15 20 25 30 May Date 第8図 実験期間中の実験地の気温.
第4表 パッションフルーツの3花型花の形態的特性
Table 4. Morphological characters of the three flower types in purple passion fruit
Cross Type of flower diameter
of flower Length Width Length Width
Normal style type Recurvate style type Upright style type
Cm mm mm 6.8 13.9 11.4 6.7 15.1 11.6 6.1 13.7 11.6 耽 mm 27.9 11.5 28.2 11.6 24.9 11.7 Type of flower Length Width Length of corona Ovary Length of
Length Diameter style Normal style type
Recurvate style type Upright style type
訊 mm 23.7 7.2 22.8 7.7 21.4 6.8 mm mm 18.5 7.0 20.3 7.1 18.6 5.9 耽 mm 4.9 13.5 4.9 13.7 4.2 12.0
Stigma Filament Anther
Type of flower
Length Width Length Width Length Width
Normal style type Recurvate style type Upright style type
mm 耽7n 5.4 3.7 5.6 3.6 4.4 3.0 n7n 耽7n 7.1 1.6 7.4 1.6 7.6 1.6 耽 mm 9.3 3.9 9.0 4.0 9.5 3.7 第5表 パッションフルーツの開花前日の気温と関前時刻との相関(1982年5月1日から6月 4日)
Table 5. Correlation coefficients between time of the anther dehiscence(2) and air
temper-ature of the day before flowering from May 1 to June 4, 1982, in purple
passion fruit Time of the day Correlation.(, lemperaturerr..JRegressionequation^ coefficients ℃ o o o o o o o o o o o o o o o o o o o o ●● N ^ ^ O O O N ^ N ^ t O C S I C M < X l 23.0±1.3 22.6 ±3.8 23.2 ±2.8 21.4±2.8 19.9±2.7 19.1 ±2.7 18.4 ±3.2 15.9±4.7 17.3±3.2 17.2±3.0 -0.71s1 -0.34 -0.62* -0.39 -0.46 -0.39 -0.45 -0.19 -0.48* -0.58* Y- -0.34X+ 15.70 Y- -0.13X+10.99 Y- -0.33X+15.62 Y- -0.20X+ 12.32 Y- -0.25X+ 12.98 Y- -0.21X+ll.94 Yニー0.21X+ll.79 Y--0.06X+ 8.86 Y--0.23X+Hi Y- -0.29X+12.96
(i) Y : Time, X : Temperature.
(2) Mean time of anther dehiscence was at 7:51 am. ** : Significant at 1% level.
第6表 パッションフルーツの開花前日の気温が開花日の開所時刻に及ぼす影響1982年5月 1日から6月4日)
Table 6. Effects of air temperatures of the day before flowering on anther dehiscence
from May 1 to June 4, 1982, in purple passion fruit
Air temperature Time of anthesis in the field at flowering day
The day before flowering
Max. Mm.
Early morning of Max. flowering day Min.
26.3± 1.4 21.5±1.7 18.3±2.0 13.6±1.5 a.m. 7:11±0:34 9:06±1:40 7:26±1:06 9:00±1:52 考 察 パッションフルーツでは雄蕊器官は全ての花において正常に発達しほぼ類似した形態を示すが, 雌蕊器官は花によって形態的に大きな差異がみられた。そこで,花柱の形態によって接触型,接近 型及び直立型の3花型に類別したが,前2型は開花直後花柱が外側-琴曲し,やがて水平になり, 接触型は柱頭と約が接触する。しかし,接近型には開花中の時間の推移で接触型となる花も含まれ ており,接触型花は接近型花の1つのタイプと見なされた。一方,直立型花の花柱は開花直後から 夕刻∼夜半の開花に至るまで直立の形態を保ったままで変化しなかった。 1979年4月22日から5月31日の期間における3花型の着生割合は,第7図に見られるとおり 経日と共にわずかずつ変り, 5月20日以後の開花期後期には,直立型は10-22 に増加し,接近 型も漸増したが,接触型は漸減した。 パッションフルーツは熱帯の低地では生育しにくく7,19,41,75,98,163)標高500--3,000mの高地が適地 とされているが19・41,75,135)本調査の後期において接触型が漸減し直立型が増加した要因として花 芽の発育に及ぼす温度の影響が推察される。 1979年4月下旬の開花期初期における平均最高気温 は24.2℃,平均最低気温13.5℃であったが,開花後期の5月下旬には28.4℃, 14℃と最高気温が かなり高温となり(第8図),新梢の生長もやや鈍った。このようなことから,花芽発育後期及び 開花期の気温が花柱の形態に影響を及ぼすものと考えられる。
GACHANJA & GURNAH38)によれば,ケニア高地(0-59′S, 37-4′E,標高1,548m)におけるパッ ションフルーツ(開花期4月)の開花時刻は晴天で暖い日は7時∼7時45分,曇天でやや気温の 低い日は8時∼9時とされているが,桔宿市(31 15′N, 130038′E,標高5m)では4月は日の出 直後の5時30分から6時頃までに開花した。開花期中期(5月)の平均日最高気温;26.3±1.4℃, 平均日最低気温; 13.6±1.5℃では,関前時刻は6時30分から10時52分の間で,温度が高い日ほ ど早い時刻に関前がみられるようである。キイロトケイソウでは正午頃から開花が始まって夕方か ら朝にかけて閉花するとされているので1),パッションフルーツは開花・開所から閉花までの時間 がキイロトケイソウより長く,日中活動する訪花昆虫による花粉媒介には好都合であると思われる。
第3節 花型と花粉の発芽 材料と方法 パッションフルーツの花粉は一般に実験に供されるショ糖を加えた寒天培地では発芽が認められ なかったと報告されているので54)本実験では表面に雌蕊組織の圧搾汁を塗布した人工培地を供し た。 培地は寒天2%,ショ糖30%,硝酸カルシウム0.1%,ホウ酸0.02%の混合液をクエン酸でpH 6.2に調整し,オートクレーブ(l.2ka/cd)で5分間加圧滅菌した後,厚さ1耽7nになるようにシャー レ内のグラス板上に薄く拡げて作成した。凝固した後の培地を硬質スライドグラス上に貼付し,開 花当日の直立型花の柱頭,花柱の上部,花柱の下部及び子房の各圧搾汁を別々の培地表面上に塗布 して発芽床とした。湿度を十分保持したシャーレ内に置き,室温25-26℃で花粉の発芽試験に供 した。なお,対照区として接触型花の柱頭,花柱及び子房の圧搾汁を塗布した培地を用いた。花粉 の発芽試験は4回反復で,培地上にそれぞれ異なる花型の花から採取した花粉を置床し, 24時間 後に発芽率の調査を行った。 結 果 第7表に示したように,接触型花の柱頭と花柱の圧搾汁を塗布した培地上では直立型花の花粉 の発芽率は16.2%であった。一方,直立型花の柱頭,花柱の上部,花柱の下部及び子房の圧搾汁を 第7表 人工培地における直立型花の雌ずい各部位の圧搾汁が各花型花の花粉発芽に及ぼす影 響
Table 7. Effects of exudates collected from different parts of pistils of upright style type flowers on pollen germination in vitro
Pollen Exudate collecting organ Number of pollen grains observed Number of Percentage germinated of pollen pollen germination Upright Stigma(1) 1 ,046 style type Upper part of style 759 flower Lower part of style 971 Ovary(l) 1,172 a> oo o oa 0.9 0.4 0.0 0.2 Normal Stigma(1) 1, 188 style type Upper part of style(i) 1,040 flower Lower part of style 902 Ovary(l) 1 ,006 Oi O O ^f 00 0 0 ^ ● O O O O Upright
style type Stigma and stylel } 1,016 flower
(1) : Upright style type flower. (2) : Normal style type flower.
塗布した培地上では直立型花の花粉の発芽率はそれぞれ0.9%, 0.4%, 0%及び0.2%,接触型花の 花粉の発芽率は0.8%, 0%, 0%及び0.4%であった。このように,直立型花の花柱上部圧搾汁を 塗布した培地上では,直立型花の花粉は0.4%発芽したが,接触型花の花粉の発芽は認められず, 花柱下部の圧搾汁を塗布した培地上では直立型花,接触型花ともに花粉の発芽は認められなかった。 考 察 GILMARTIN39)は,株間で不和合性を示すキイロトケイソウでは,柱頭上の花粉は殆んど未発芽 の状態に留っているが,まれに発芽しても花粉管は80urn以上には伸びないと報告している。 本実験では,接触型花の柱頭と花柱の圧搾汁液を塗布した人工培地上では,直立型花の花粉は発 芽率が16.2%であったのに対し,直立型花の柱頭,花柱及び子房の圧搾汁を塗布した人工培地上 では,直立型及び接触型花の花粉の発芽率は1 %以下であった。とくに,直立型花の花柱下部の圧 搾汁を塗布した培地上では直立型花及び接触型花の花粉とも全く発芽は認められなかった。この結 果は,直立型花の柱頭や花柱には花粉の発芽及び花粉管の伸長に対して抑制的に作用する物質が存 在し,その抑制物質は花柱部により多く含まれていることを示唆している。 以上の結果から,パッションフルーツにおいて,結果率が低い原因の一つは比率は低いものの直 立型花の増加であろうと考えられる。 第4節 花型と結果 材料と方法 指宿市十二町で合掌仕立てで栽培している実生3-5年生株を用い, 1979年5月21日∼30日 の間に,交配実験を行った。開花前日に菅に袋をかけ,開花当日に花型を確認しながら自家受粉を 行った。また開花前日に除雄して袋をかけ,開花当日に花型を確認しながら異花型間の人工受粉を 行った。 さらに, 1979年5月24日, 25日, 28日には無袋条件下で直立型×直立型,直立型×接触型の 人工受粉を行った。受粉30日後に結果率を調査した。 結 果 第8表に示したとおり,各花型の自家受粉に供試した花数は接触型:191 73.5% ,接近型: 38 (14.6%),直立型:31 (ll.9% であったが,結果率は接触型では94.2%,接近型では89.5% でほぼ同程度,直立型では0%であった。 次に,第9表に示したように直立型の花粉を人工受粉した花数は接触型:240 (60%),接近型: Ill (27.8% 及び直立型; 16 (4.0%)であった。結果率は接触型花は84.2%,接近型花は60.4%, 直立型花は0%であった。一方,異型花の花粉を受粉した直立型花の結果率は0%であった。なお, 直立型花を開花前日に確認することはきわめて困難であったので,自然条件下で開花した花につい て,直立型花相互間の他家受粉を122花,直立型×接触型の組合せで170花を受粉したが,これら はすべて結果しなかった(第10表)。
第8表 パッションフルーツの各花型花の自家受粉による結果率
Table 8. Percentage of fruit set in different types of flower self polli-nated artificially in purple passion fruit
● Type of flower Number of flowers observed Number of Percentage of
fruit set fruit set
Normal style type Recurvate style type Upright style type
第9表 圃場における各花型花相互の人工受粉(紙袋掛け)による結果率
Table 9. Effect of artificial crosses on fruit bearing of flower covered with paper bag under field condition
Cross combination
Number of flowers crossed
Number of Percentage of
fruit set fruit set
Normal style type X Upright style type Recurvate style type
X Upright style type Upright style type
X Upright style type Upright style type
X Normal style type Upright style type
X Recurvate style type
240 円Ill 16 17 202 67 84.2 60.4 0.0 0.0 第10表 圃場における各花型花相互の人工受粉(紙袋掛け無し)による結果率
Table 10. Fruit set m cross pollinated flowers without bagging under field condition Cross combination Number of flowers observed Number of Percentage of
fruit set fruit set
Upright style type X Upright style type Upright style type
X Normal style type Normal style type
X Upright style type
122
170
0.0'
考 察
パッションフルーツは自家稔性とされているが,放任条件下では結果率が低く,特に雨天時の 花は結果率が著しく低下することが知られている。人工受粉によって結果率を高めることが可能 とされているが,その実施には仕立て方や受粉作業などに多大の労力を要するので問題がある。 一般に,トケイソウ属の花では蜜の分泌量が多いため29)サ これを求めて多くの昆虫が訪花するが29)
ハワイ,フロリダ及びアフリカ等では有効な花粉媒介昆虫,とくにcarpenter bees (Xylocopa varipuncta Patlon, X.spp.)やhoney beesあるいはwasps (Polistes spp.)などが報告されて いる4,5,39,42,87,119,135。しかし,これらの昆虫は南九州には生息しておらず,時々,ミツパテ(Apis indica japonica Radoszkowski)の訪花はあるが,結果率の向上に対して人工受粉ほどの効果は 得られていないようである。 各種の花粉媒介生物が生息する熱帯地方では,トケイソウ属内の種間雑種がかなり多くみら れ7,8,98)種間の交雑和合性がかなり高いことがうかがわれるが,パッションフルーツには人工受 粉を行っても結果しない直立型花が着生する。しかし,直立型花の花粉を接触型花や接近型花へ受 粉した場合は結果することから,直立型花の花粉は受精能力を有することが確認された。 Akamine & GIROLAMI5)はキイロトケイソウは系統によって直立型花が0-51.0%の変異幅で発生するが, これらの花には生理的な障害があり,結果は得られないと報告している。これらのことからトケイ ソウ属の直立型花の雌蕊組織には花粉発芽や花粉管伸長を抑制する物質の存在が考えられる。 第5節 摘 要 パッションフルーツの花型の類別と開花及び関前状況,花型別の花粉発芽特性,花型別の自家受 粉及び異花型間の人工受粉による結果性について調査を行った。 1.花は早朝に開花して,夕刻より夜半に閉花し,翌日再び開花することはなかった。 2.開花時における花柱の形態ならびに柱頭と約の位置関係から,接触型(正常花) (normal style type),接近型(recurvate style type)及び直立型(upright style type)の3花型に類別され た。前二者は形態的にはほぼ類似しており,接触型は開花後に接近型が変化して出現することが 明らかになった。直立型は花の直径,琴片,花弁長,花柱長,柱頭の長径及び短径,さらに子房 の長径が接触型及び接近型より明らかに小さかった。 3.接触型の出現率が最も高い午後1-3時の調査結果によれば,総開花数6,053のうち接触型が 76%,接近型が17%及び直立型が7%であった。 4.最高気温:26.3±1.4℃,最低気温: 13.6±1.5℃の期間に開花した花においては開所時刻が6 時30分∼ 10時52分であったが,この気温の範囲内では気温が高いほど開所時刻は早かった。 5.寒天2%,ショ糖30%,硝酸カルシウム0.1%,ホウ酸0.02%の混合液をクエン酸でpH6.2に 調整して作成した人工培地に,直立型花の柱頭,花柱上部及び下部,子房の圧搾汁をそれぞれ塗 布して行った接触型花及び直立型花の花粉の発芽率は1%以下であった。接触型花の柱頭及び花 柱の圧搾汁を塗布した培地上では直立型花の花粉の発芽率は16.2%であった。 6.人工受粉を行った自家受粉花の結果率は接触型花では94.2%,接近型では89.5%,直立型花 では0%であった。 7.直立型花の花粉を受粉した接触型花は84.2%,接近型花は60.4%の結果率を示したが,異花 型の花粉を受粉した直立型花の結果率はいずれの場合も0%であった。
第4章 受粉及び受精 第1節 緒 看 一般にキイロトケイソウは自家不稔性であるが3,4)本実験に用いたパッションフルーツは自家 稔性とされている3,4,61,62,75,84,85,112,145)。しかしながら,パッションフルーツには自家不稔性花が出 現することもあって,放任条件下で結果率は低く58,59,64,180)果実生産上問題となっており,結果率 向上対策の改善が切望されている。一方,ハワイ及びオーストラリアでは放任条件下で高い結果率 が得られているが,これには受粉媒介昆虫carpenter-beesの存在が大きく貢献していると報告され ている4,42,75,120)。我が国には同種は生息していないため,ミツパテなどの訪花はみられているが, 昆虫による十分な受粉媒介効果はあがっていない現状である。 パッションフルーツでは果汁量及び果汁重は果実に含有されている種子数と相関が高いことか ら63)受粉の効果がとくに大きいことが推察される。一方,受粉の良否は開花時の天候にも大きく 左右されることが報告されている67)。しかし,受粉に及ぼす気温の影響に関する報告はみられない。 パッションフルーツの自家不稔性または交雑不和合性の究明の方法として,また,受粉用花粉の 確保のうえからも花粉の発芽特性を明らかにすることは重要である。パッションフルーツの柱頭上 での花粉発芽についてはGILMARTIN39)の報告があるが,人工発芽床の培地組成や発芽条件に関し て詳細に検討した報告は見当らない。
AKAMINE & GIROLAMI5)はキイロトケイソウの花粉発芽用の人工培地の条件として浸透圧の問 題やインドール酢酸様発芽促進物質の必要性を挙げているが,花粉発芽に関するデータは見当らな い。 本章ではパッションフルーツの花粉発芽に好適な人工培地の成分組成, pH及び培養温度につい て検討するとともに,自家または他家受粉しても結果しない直立型花の花粉の発芽特性を人工培地 上で実験し,花粉の利用性について検討した。 さらに,雌蕊組織内における花粉管の伸長と受精にいたる過程を究明するため,受粉後経時的に 解剖形態学的観察を行った。トケイソウ属の雌蕊は分岐した3本の花柱, 3心皮1室,側膜胎座か らなっている23,39,112,121)。柱頭に受粉され,発芽した花粉管はそれぞれの心皮組織につながる柱頭一 花桂一胎座へ伸長し,受精するものと思われるので,とくに,柱頭一花柱と子房内胎座との組織的 な関係,及び柱頭上で発芽した花粉管が3心皮の胎座へどのように伸長していくかについて着目し, 受精にいたる経過を究明した。 第2節 人工培地上における花粉の発芽 材料と方法 指宿植物試験場の圃場で栽培中のパッションフルーツ実生の3-5年生株の花を供試した。花粉 の発芽床には,滅菌した硬質スライドグラス上に貼付した各種組成からなる寒天培地の薄片(1×1 cm)を用いた。実験に際しては,開花前日に除雄して袋掛けしておいた接触型花の柱頭の圧搾汁を 寒天培地の表面に薄く拡がる程度に塗布し,圧搾汁を塗布しない培地を対照とした。これらの培地 (発芽床)は,底にろ紙(Toyo, No.2)を敷き,給水してほぼ100%の湿室状態を保ったシャーレ
内(直径15cm)に静置した。なお,柱頭の圧搾汁は開花開始時に切り取った柱頭をポリエチレン 製の網でつつみ,乳鉢の中でガラス棒を押しつけて搾汁し,その汁液を供用した。特記する場合の ほかは花粉は荊裂開直後の接触型花から採取し,発芽床に振り落し,塊状になった花粉粒は表面に 薄く拡げる方法を用いた。発芽床1個当りの花粉数は500-900粒であった。発芽試験はそれぞれ 3-6回反復し,置床24時間後酢酸カーミンで染色,固定して鏡検した。 なお,本実験に先だって予備実験を行い,次のような知見が得られた。すなわち, i :ショ糖濃 度1-25 の水溶液中に入れた花粉は5-20秒の間に破裂した。 ii :寒天濃度1-5 とショ糖濃 度0-60%の組合せによる培地上では,花粉の発芽は認められなかった。 Ill 寒天濃度: 1%及び 2%とショ糖濃度:30%及び40%を組合せた組成に,さらにホウ酸:0.01%を加えて作成した培地 上に接触型花の柱頭圧搾汁を塗布した場合,わずかながら花粉の発芽が見られた。これらの場合, 培地のpHはほぼ6.4が適当であった。 実験Ⅰ.花粉の発芽に及ぼす培地の寒天,ショ糖,ホウ酸濃度の影響 (1)寒天濃度:1%または2%,ショ糖濃度:30%, 40%または50%,ホウ酸濃度:0.002%, 0.005%, 0.01%, 0.015%または0.02%を組み合せた培養液を,クエン酸でpH6.4に調整した後, オートクレーブで滅菌して培地を作成した。そして,その培地表面に柱頭圧搾汁を塗布した区と塗 布しない区(対照)を設けた。発芽床は20-25℃の室内に静置した。 (2)寒天濃度:1%または2%,シヨ糖濃度:20%, 30%または40%,ホウ酸濃度:0%, 0.01%, 0.02%, 0.03%, 0.04%または0.05%を組合せて作成した培地表面に柱頭圧搾汁を塗布した区と, 塗布しない区を設けた。発芽床は20-25℃の室内に静置した。 実験Ⅱ.花粉の発芽に及ぼす培地のpH値及び温度の影響 実験Ⅰの結果から寒天濃度:2%,ショ糖濃度:30%,ホウ酸濃度:0.03%の培養液をクエン酸 でpH:5.4, 5.6, 5.8, 6.0, 6.2, 6.4及び7.0に調整して培地を作成し,表面に柱頭圧搾汁を塗布 した。花粉置床後15℃, 20℃, 25℃, 30℃及び35℃の恒温器内に静置した。 実験Ⅲ.花粉の発芽に対するホウ酸及び硝酸カルシウム濃度の影響 実験Ⅱにおいて, pH6.2,温度25℃の場合に比較的高い発芽率が得られたので,寒天濃度: 2%, ショ糖濃度:30%を基本に,ホウ酸濃度:0.02%, 0.03%及び0.04%,硝酸カルシウム濃度:0%, 0.04%, 0.06%, 0.08%, 0.10%及び0.12%を組合せ, pH値は6.2に調整した培地を作成し,柱頭 圧搾汁を表面に塗布して25℃の恒温室内で実験を行った。 実験Ⅳ.花粉の発芽に及ぼす培地pHの影響 培地の組成は実験で高い発芽率が得られた培地と同じ培地(寒天濃度: 2%,シヨ糖濃度: 30%, ホウ酸濃度:0.02%,硝酸カルシウム濃度:0.1% を基本培地とし, pHを3.0, 3.5, 4.0, 4.5, 5.0, 5.5, 6.0及び7.0に調整して培地を作成した後,柱頭圧搾汁を培地の表面に塗布した。発芽実 験は25℃の恒温室内で行った。 実験V.花粉発芽に及ぼす糖の種類と濃度の影響 実験Ⅳで高い発芽率が得られた寒天濃度:2%,ホウ酸濃度:0.02%,硝酸カルシウム濃度: 0.1%, pH:4.0及び4.5の培養液に糖としてブドウ糖及び果糖濃度を1%, 5%, 10%, 15%, 20 i, 30%及び40%になるように添加して培地を作成した後,柱頭圧搾汁を培地表面に塗布した。 他の発芽条件は実験Ⅳと同じであった。 実験Ⅵ.花粉の発芽と花型との関係 晴天及び雨天の日における接触型花及び直立型花の花粉発芽を,実験Ⅳで高い発芽率が得られた
培地(寒天濃度:2%,ホウ酸濃度:0.03%,硝酸カルシウム濃度:0.1%,ショ糖濃度:30%, pH: 4.0及び4.5 で調査した。雨天の日の花は雨水で濡れていない薪と濡れた荷を供試した。他の発 芽条件は実験Ⅳと同じとした。 結 果 実験Ⅰ∼Ⅵを通じて,接触型花の柱頭の圧搾汁を培地(発芽床)表面に塗布しなかった対照区に おいては,花粉の発芽は全く認められなかった。 \ 実験Ⅰ.花粉の発芽に及ぼす培地の寒天,ショ糖及びホウ酸濃度の影響 まず,寒天,ショ糖及びホウ酸濃度の花粉発芽への影響を第9図に示した。供試した各培地に おいて発芽率は低かったが,ショ糖濃度:30%,ホウ酸濃度:0.02%,寒天濃度:1%及び2%の 培地上における花粉の発芽率は2.95%及び2.60%を示し,その他の濃度組成の培地よりやや高率 であった。発芽率は総体的に低かったが,ホウ酸の濃度が高くなるにつれてやや高くなる傾向が認 められた。 次に,ホウ酸濃度を0.01%から0.05%にまで高めた場合の花粉発芽への影響を第10図に示し た。花粉の発芽率は寒天濃度:2%,ショ糖濃 皮:30%,ホウ酸濃度:0.03%の培地で6.5% と最高を示し,寒天濃度:1%,ショ糖濃度: 30%,ホウ酸濃度: 0.01%の培地では3.84%を 示した。これらの結果から,寒天濃度が1-2 %,ショ糖濃度が30%の培地ではホウ酸の適 正濃度は0.01-0.03 の範囲にあるものと推 測される。 U O l } B U I U I J 9 S J O 9 S h ; U 9 D J 9 J 0 0.002 0.005 0.010 0.015 0.020
Concentration of boric acid {%) 第9図 花粉発芽に及ぼす寒天,ショ糖及び
ホウ酸濃度の影響.
Fig. 9. Effect of concentrations of agar, sucrose and boric acid on pollen germination. The media were add-ed with juice pressadd-ed out of the stigma and pH was adjusted to 6.2. The germination beds were maintained at 20 to 25oC.
L O ^ t c O O O , -I U O I } B U I U I i a g l 0 9 S h ; U 9 3 J 9 J 0.01 0.02 0.03 0.04 0.05
Concentration of bork acid {%) 第10図 花粉発芽に及ぼす寒天,ショ糖及び
ホウ酸濃度の影響.
Fig. 10. Effect of concentrations of agar, sucrose and boric acid on pollen germination. The media were added with juice pressed out of the stigma and pH was adjusted to 6.4. The germination beds were maintained at 20- to 25C.
実験Ⅱ.花粉の発芽に対する培地のpH値及び温度の影響 第11表及び第12表にみられるように,寒天濃度:2%,ショ糖濃度:30%,ホウ酸濃度: 0.02%の培地では,培地のpH値が5.4-7.0の間では発芽率に有意差は認められなかった。しかし, 平均花粉発芽率はpH6.2で最高を示し4.65%,次いで6.0で2.25%, 6.4で2.12%の順であった。 そこで,各pHごとに最も高い発芽率を示した温度についてみると, pH6.2では30℃で13.80%, 6.0では25℃で7.60%, 6.4では25℃で5.06%と, pHの違いによって最も高い発芽率が得られた 温度は異なっていた。なお,花粉の平均発芽率と温度との間には1%水準で有意差が認められ,平 均発芽率は25℃で最高の5.35%,次いで30℃の3.75%であったが,その他の温度区では非常に低 かった。とくに, 15℃ではpH5.4で0.50%の発芽がみられたにすぎず, 20℃では0-1.60%, 35 ℃では0-2.96%と非常に低い発芽率であった。 第11表 パッションフルーツの花粉発芽に及ぼす人工培地のpHと温度の影響 培地の組成は寒天2%,ショ糖30%,ホウ酸0.02%及び硝酸カルシウム0.10%.培 地上に柱頭圧搾汁を塗布した
Table ll. Effects of pH of the culture media and temperatures on pollen germination of purple passion fruit. The media were prepared with the combinations of 2% agar, 30% sucrose, 0.02% boric acid and 0.10% calcium nitrate, and
add-ed juice pressadd-ed out of the stigma on the surface of madd-edia
Temperature ℃) 25 30 Average % % % % 5.4 5.6 5.8 6.0 6.2 6.4 7.0 Average O O O O O O O N L O O O O O O O O o o o o o o o o O O C D C O O O O C D o o m c o o < r > o c o O O O O O -H O O <」> 00 CO O ) N -I C O L O O O C O ^f LO ^ l^- CO LO ^ LO 1.13 1.33 1.50 2.93 13.80 3.56 2.00 3.75 O O O C D O c o O C O C O o t>- -i ^ o^ ^H oo as ●● O O -I O C S I O O O L O L O C N I co ^ ^ Csi cD -H Co o ● c^ ^h <>a 第12表 第11表の分散分析
Table 12. Analysis of variance of data shown in Table ll SumofD Factorsquaresir≡:dr;eofTr.'Variance--1#1. Variance.Probability >mratio pH 42.47 Temperature 152.94 Error 97.99 24 Total 293.41 34 7.08 1.73 38.26 9.36 4.08 ** : Significant at 1% level.