Nagoya City University Academic Repository
学 位 の 種 類 博士 (薬科学) 報 告 番 号 甲第1756号 学 位 記 番 号 第353号 氏 名 近藤 聡志 授 与 年 月 日 平成 31 年 3 月 31 日 学位論文の題名 ヒト iPS 細胞由来腸管幹細胞の維持培養方法の確立と腸管上皮細胞の粘膜 障害作用及び保護作用評価への利用 論文審査担当者 主査: 粂 和彦 副査: 松永 民秀, 山村 壽男, 尾関 哲也
名古屋市立大学学位論文
ヒト
iPS細胞由来腸管幹細胞の維持培養方法の確立と
腸管上皮細胞の粘膜傷害作用及び保護作用評価への利用
平成
30 年度 (2019 年 3 月)
名古屋市立大学大学院薬学研究科
臨床薬学分野
近藤
聡志
一.本論文は2019 年 3 月名古屋市立大学大学院薬学研究科において審査されたものであ る。 主査 粂 和彦 教授 副査 尾関 哲也 教授 山村 寿男 教授 松永 民秀 教授 二.本論文は、学術情報誌に掲載された次の報文を基礎とするものである。 【基礎となる報文】
1.
Satoshi Kondo, Shota Mizuno, Tadahiro Hashita, Takahiro Iwao, Tamihide Matsunaga. Using human iPS cell-derived enterocytes as novel in vitro model for the evaluation of human intestinal mucosal damageInflammation Research, 67, 975-984 (2018).
2.
Satoshi Kondo, Shota Mizuno, Tadahiro Hashita, Takahiro Iwao, Tamihide Matsunaga. Establishment of a novel culture method for maintaining intestinal stem cells derived from human induced pluripotent stem cellsBiology Open, 9 (1) (2020).
三.本論文の基礎となる研究は、松永民秀 教授の指導のもとに名古屋市立大学大学院薬 学研究科において行われた。
学位論文内容要旨 ヒトiPS細胞由来腸管幹細胞の維持培養方法の確立と腸管上皮細胞の 粘膜障害作用及び保護作用評価への利用 近藤 聡志 【序論】 小腸には多くの薬物代謝酵素や薬物トランスポーターが存在しており、肝臓と同様に薬 物の初回通過効果に関わる臓器として非常に重要である。そのため薬物動態に優れた医薬 品開発には、早期の段階から小腸における膜透過性や代謝安定性を評価することが重要で ある。現在、腸管上皮細胞のモデル細胞としてヒト結腸癌由来のCaco-2細胞が汎用されて いるが、薬物代謝酵素や薬物トランスポーターの発現パターンがヒト腸管上皮細胞と異な っていることから、腸管での薬物動態を正確に評価することは困難である。初代小腸上皮 細胞の利用が理想的ではあるが、その入手は難しい。そこで、多分化能とほぼ無限の増殖 能を有するヒト人工多能性幹細胞(induced pluripotent stem cells:iPS細胞)から作成される 腸管上皮細胞の利用が期待されている。本研究室では、これまでヒトiPS細胞由来腸管上皮 細胞の有用性を報告してきた。しかしながら、iPS細胞から腸管上皮細胞まで分化させるに は多くの時間とコストが必要であり、さらに、薬物動態試験で利用するためには、大量か つ安定的に供給する必要がある。そこで、第1章ではこれらの問題を解決するため、ヒト iPS細胞から腸管上皮細胞へ分化させる前段階である腸管幹細胞を維持培養する方法の検討 を行った。 また、第2章では、ヒトiPS細胞由来腸管上皮細胞の腸管ムチンの発現量を指標とした毒 性評価への利用として、薬物に起因する腸管粘膜障害作用及び粘膜保護作用の評価細胞と しての有用性について検討した。腸管ムチンは主に杯細胞から分泌される糖タンパク質で あり、粘膜上皮を保護すると共に細菌などの外的侵入から生体を守る重要な役割を果たし ている。腸管ムチンはMucin 2(MUC2)が主な成分であり、MUC2の発現量は薬物により 増減することが報告されている。例えば、抗炎症剤である非ステロイド性抗炎症薬(Non-Steroidal Anti-Inflammatory Drugs, NSAIDs)は胃粘膜だけでなく腸管粘膜の障害も引き起こ し、腸管粘膜のMUC2発現量を減少させることが報告されている。一方、レバミピドやイ ルソグラジンなどの粘膜保護薬(mucosal protective agents, MPAs)は胃粘膜だけでなく腸管 粘膜の保護作用も有しており、腸管粘膜のMUC2発現量を増加させることが報告されてい る。これらの評価は主に動物を用いたin vivo試験であり、in vitro試験で評価された報告は 少ない。そこで、ヒトiPS細胞由来腸管上皮細胞が薬物による粘膜障害や粘膜保護作用の評 価に有用であるか検討を行った。
ii 【本論】
第1 章 ヒト iPS 細胞由来腸管幹細胞の維持培養方法の確立 1.FBS及びKSRの腸管幹細胞性及び腸管細胞性への影響
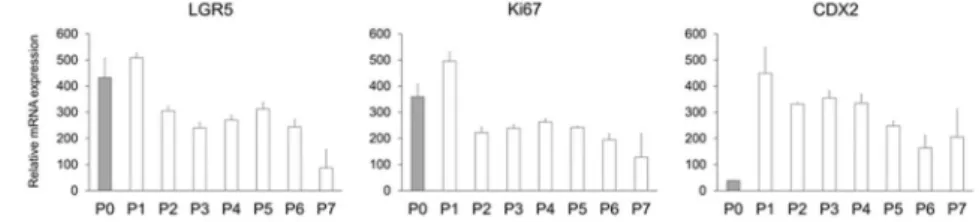
細胞培養で広く用いられるウシ胎児血清(Fetal bovine serum; FBS)には細胞増殖因子、分 化促進因子やホルモンなど未知の因子が含まれている可能性があり、幹細胞の維持培養に は適さないと考えられる。そこで、FBSの代替としてES細胞やiPS細胞の培養に用いられる KnockOut Serum Replacement(KSR)を培地に添加し、その影響を検討した。その結果、腸 管幹細胞マーカーとして知られるleucine rich repeat containing G protein coupled receptor 5 (LGR5)のmRNA発現量はFBS濃度の増加に伴って減少した。KSRについても濃度の増加に 伴うLGR5発現量の減少が見られたものの、その程度は小さかった。腸管細胞性マーカーで あるCDX2のmRNA発現量に対する影響には明らかな差はなかった。この結果から、KSRは FBSに比べて幹細胞性に与える影響は小さいと考えられ、KSRを代替として用いることで以 降の検討を進めた。 2.CHIR99021及びVPAの腸管幹細胞性及び腸管細胞性への影響 腸管幹細胞の維持にはWNT 及び Notch シグナルの両方が重要であることが報告されてい る。そこで、GSK-3β を阻害し WNT シグナル伝達を活性化させる CHIR99021、Notch シグ ナル伝達を活性化させるvalproic acid(VPA)の腸管幹細胞性及び腸管細胞性に対する影響 を培地に添加することで検討した。その結果、CHIR99021 は濃度依存的に LGR5 及び CDX2 のmRNA 発現量を増加させ、VPA も LGR5 の発現量を増加させた。CHIR99021 及び VPA を 併用した場合は、それぞれを単独で使用した場合と比べてLGR5 及び CDX2 の発現量を増 加させた。この結果より、CHIR99021 及び VPA が腸管幹細胞の維持に有効であることが示 唆された。したがって、これら化合物に加え、幹細胞性の維持に重要であることが既に報告 されているいくつかの因子を使用することで以降の検討を進めた。 3.継代を重ねた際の腸管幹細胞性及び腸管細胞マーカーの発現量評価 ヒトiPS 細胞を腸管幹細胞まで分化させ、様々な幹細胞性を促進する因子を添加した培地 を用いて継代を重ねた細胞における腸管幹細胞性(LGR5, Ki67)及び腸管細胞性(CDX2) マーカーのmRNA 発現量を評価した(Fig. 1)。その結果、LGR5 及び Ki67 は P7 までほぼ 同程度の発現量が維持されていた。CDX2 については P0 に比べ P1 から P7 で高く、発現量 は維持されていた。
Fig. 1. Effects of passage number on LGR5, Ki67, and CDX2 mRNA levels in ISCs derived from human iPS cells.
The expression levels of each gene were normalized to those of HPRT1 and are presented relative to levels in the human adult small intestine (set as 100). Data represent the mean ± standard deviation (n = 3).
4.継代数ごとの腸管上皮細胞における分化マーカーの発現量評価
複数回継代した腸管幹細胞が腸管上皮細胞への分化能を維持しているか評価するため、 分化誘導後いくつかの分化マーカーの mRNA 発現量を評価した(Fig. 2)。その結果、P-glycoprotein(P-gp)及び Breast cancer resistance protein(BCRP)はヒト小腸の発現量よりも 高く、P0 から P7 ではほぼ同程度であった。Villin、sucrase-isomaltase 及び MUC2 では、P0 からP3 までほぼ同等であった。CYP2C9、CYP2C19、CYP3A4、UDP グルクロン酸転移酵 素1A1(UGT1A1)、ペプチドトランスポーター1(PEPT1)、有機アニオントランスポーター 2B1(OATP2B1)及びグルコーストランスポーター1(SGLT1)は、継代を重ねるごとに発現 量の低下が認められた。
Fig. 2. mRNA levels of intestinal and pharmacokinetics markers in
enterocytes derived from human iPS cells (P0–P7). The expression levels of each gene were normalized to those of HPRT1 and are presented relative to levels in the human adult small intestine (set as 100). Data represent the mean ± standard deviation (n = 3). 5.継代を重ねた腸管幹細胞から分化した腸管上皮細胞の薬物代謝活性能及び薬物輸送活 性能評価 CYP3A4 代謝活性能評価 CYP3A4 は、腸管上皮細胞にもっとも多く発現している薬物代謝酵素であり、多くの薬物 が基質となることから、最も重要な代謝酵素である。そこで、CYP3A4 の代表的な基質であ るミダゾラムを用いてCYP3A4 の主要代謝物である 1'-ヒドロキシミダゾラムの生成量を定 量することで代謝活性能を評価した。その結果、1'-ヒドロキシミダゾラムの生成は P0 から P3 までの腸管上皮細胞で認められ、その生成量に大きな違いはなかった。 P-gp 及び BCRP の輸送活性能評価 P-gp 及び BCRP は、様々な薬物の腸からの吸収及び組織分布を制限する重要な役割を果 たす排出トランスポーターである。そこで、P-gp の基質として Rhodamin123、BCRP の基質 としてHoechst33342 を用いて P0 及び P3 の腸管上皮細胞における輸送活性能を評価した。 その結果、apical 側から basal 側及び basal 側から apical 側への見かけの膜透過係数(Papp) 値から算出したefflux ratio(ER)は P-gp では P0 及び P3 において 14.6 及び 7.2、BCRPP で
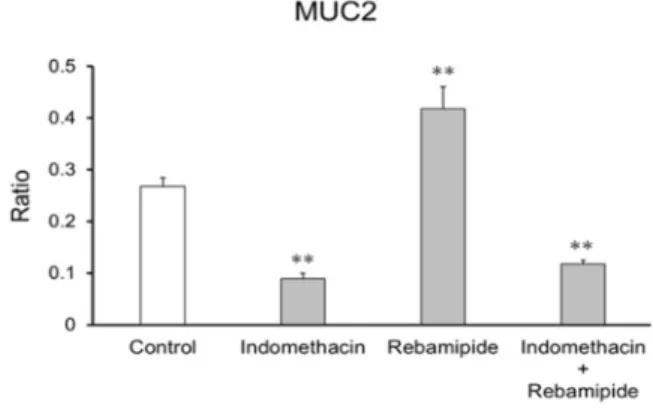
iv は11.6 及び 9.8 であり、輸送活性能を有することが示された(Fig. 3)。また、P-gp の阻害剤 としてベラパミル、BCRP の阻害剤として Ko143 を用いたところ、それぞれの P0 及び P3 でのER は 1 に近づいたことから輸送能が阻害されたことが示唆された。以上の結果より、 継代を重ねた腸管幹細胞から分化した腸管上皮細胞においてもP-gp 及び BCRP の輸送活性 能を有しており、排出トランスポーターの評価が可能であることが示唆された。 Fig. 3. Permeability of Rhodamine 123 and Hoechst 33342 across enterocyte monolayers. The ER for Rhodamine 123 or Hoechst 33342 was derived from the Papp values associated with basal-to-apical transport and apical-to-basal transport. Data represent the mean ± standard deviation (n = 3). 第2章 ヒトiPS細胞由来腸管上皮細胞の粘膜障害作用及び保護作用評価への利用 1.ヒトiPS細胞由来腸管上皮細胞のMUC2の発現量評価 ヒトiPS細胞から分化させた腸管上皮細胞におけるMUC2のmRNA発現量を評価した。そ の結果、腸管上皮細胞におけるMUC2の発現量はヒト小腸と同程度であった。一方、Caco-2細胞におけるMUC2の発現量はヒト小腸の100分の1以下であった。以上の結果より、腸管 上皮細胞は十分なMUC2の発現量を有していることが確認された。 2.NSAIDs及びMPAsのMUC2のmRNA発現量に与える影響 腸管上皮細胞の培地にNSAIDs(インドメタシン、ケトプロフェン、メロキシカム)、 MPAs(レバミピド、イルソグラジン)又は両薬物(インドメタシン+レバミピド)を添 加し、6日間培養後、MUC2のmRNA発現量を評価した。その結果、NSAIDs添加群ではコ ントロール群(媒体を添加)と比較してMUC2の発現量が減少し、MPAs添加群ではMUC2 の発現量が上昇した。また、両薬物を添加した群ではMUC2の発現量が低下した。 3.インドメタシン及びレバミピドのMUC2のタンパク発現量に与える影響 腸管上皮細胞の培地にインドメタシン、レバミピド又は両薬物を添加し、6日間培養し た細胞を用いて免疫蛍光染色を行い、MUC2陽性細胞率を評価した(Fig. 4)。その結果、
コントロール群と比較してレバミピド添加群では約1.6倍の陽性細胞率の上昇が認められ た。一方で、インドメタシン添加群では、その2分の1以下に減少した。また、両薬物を添 加した群においても2分の1以下の減少が認められた。この結果は、mRNA発現量の増減と 一致した。ラットを用いたin vivo試験おいてNSAIDsによる腸管粘膜でのMUC2発現量の減 少がMPAsにより抑制される結果が報告されているが、本検討ではインドメタシンによる MUC2発現量の減少に対するレバミピドの抑制作用は認められなかった。
Fig. 4. Ratio of MUC2-positive cells in enterocytes derived from human iPS cells.
The number of MUC2-positive cells was divided by the number of nuclei in enterocytes derived from human iPS cells. Enterocytes were treated with indomethacin, rebamipide, or indomethacin and rebamipide for 6 days from Day 21.
Data are presented as the mean ± S.D. (n = 3). Levels of statistical significance were compared with the control; **p < 0.01.
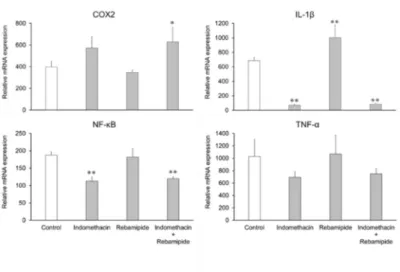
4.NSAIDs及びMPAsのCOX-2及びサイトカイン発現量に与える影響 Cyclooxygenase(COX)-2は炎症時に誘導されることが知られている生理活性酵素であ る。また、MUC2の発現は様々なサイトカインによって制御されていることが報告されて いる。そこで、腸管上皮細胞にインドメタシン、レバミピド、両薬物を添加し、6日間培 養した際のCOX-2及び炎症性サイトカイン(IL-1β、NF-κB及びTNF-α)のmRNA発現量を 評価した(Fig. 5)。その結果、COX-2の発現量はコントロール群と比べインドメタシン添 加群で上昇した。一方、レバミピド添加群ではコントロール群と比べ低い傾向を示した。 両薬物を添加した群はインドメタシン添加群と同様に上昇した。IL-1βの発現量はコントロ ール群と比較してインドメタシン添加群で大幅に低下し、レバミピド添加群では約1.4倍の 上昇が認められた。また,両薬物を添加した場合はインドメタシン添加群と同様に大幅に 低下した。NF-κB及びTNF-αの発現量は、コントロール群と比較してインドメタシン添加 群で低下が認められ、レバミピド添加群ではほぼ同等であった。また両薬物を添加した群 ではインドメタシン添加群と同様に低下が認められた。 ラットを用いたin vivo試験において、NSAIDsによる粘膜障害時にCOX-2発現量が上昇す ることが報告されており、本検討にいてもこれまでの報告に合致した結果を得ることがで きた。MUC2の発現量の増減については、IL-1β、NF-κB及びTNF-αが影響を与えている可 能性が考えられた。
vi
Fig. 5. Effects of indomethacin and rebamipide on COX-2, IL-1β, NF-κB, and TNF-α mRNA expression levels in enterocytes derived from human iPS cells.
Relative mRNA expression levels are presented relative to the level in the human adult small intestine (set as 100).
Data are presented as the mean ± S.D. (n = 3).
Levels of statistical significance were compared with the control; *p < 0.05, **p < 0.01.
5.NSAIDs及びMPAsのPappに与える影響
Transwell上に播種した腸管上皮細胞にインドメタシン、レバミピド又は両薬物を6日間添 加し、細胞間隙透過性のマーカー分子であるFITC-dextran (average molecular weight 4,000)の 膜透過性を評価した。その結果、コントロール群と比較してインドメタシン添加群の見か けの膜透過係数(Papp)は増大する傾向がみられ、レバミピド添加群では低下する傾向が みられた。両薬物を添加した群では、コントロールと同程度のPappであった。この結果か ら、インドメタシンによる粘膜障害作用、レバミピドによる粘膜保護作用が示唆され、レ バミピドはインドメタシンによって惹起される粘膜障害を緩和させる可能性が示唆され た。 【結論】 第一章では、腸管上皮細胞への分化能を維持した腸管幹細胞の培養方法を見出すことが できた。本方法を用いることで腸管幹細胞の増幅培養が可能となり、腸管上皮細胞を多量 かつ安定的に供給できると考えられる。また、第二章では腸管上皮細胞の粘膜障害作用及 び保護作用評価への利用の可能性が示唆された。医薬品開発での毒性評価や薬理スクリー ニングへの利用が期待される。 【基礎となる報文】
1. Satoshi Kondo, Shota Mizuno, Tadahiro Hashita, Takahiro Iwao, Tamihide Matsunaga. Using human iPS cell-derived enterocytes as novel in vitro model for the evaluation of human intestinal mucosal damage. Inflammation Research. 67, 975–984 (2018).
2. Satoshi Kondo, Shota Mizuno, Tadahiro Hashita, Takahiro Iwao, Tamihide Matsunaga. Establishment of a novel culture method for maintaining intestinal stem cells derived from human induced pluripotent stem cells. Biology Open. 9 (1) (2020).
略語一覧
BA bioavailability
BCRP breast cancer resistance protein BMP bone morphogenetic protein CDX caudal-related homeobox CYP cytochrome P450
DAPI 4',6-diamidino-2-phenylindole EGF epidermal growth factor ER efflux ratio
ES細胞 embryonic stem cells FBS fetal bovine serum
FD-4 fluorescein isothiocyanate-dextran with an average molecular weight of 4000 FGF fibroblast growth factors
HBSS Hanks’ balanced salt solution IBMX 3-isobutyl-1-methylxanthine iPS細胞 induced pluripotent stem cells KSR KnockOut™ Serum Replacement L-Glu L-gutamine solution
LGR5 luecine-rich orphan G-protein-coupled receptor MAPK mitogen-activated protein kinase
MEF mouse embryonic fibroblast MPAs mucosal protective agents MUC2 mucin 2
NSAIDs non-steroidal anti-inflammatory drugs OATP organic anion-transporting polypeptide OCT organic cation transporter
PAMPA parallel artificial membrane permeability assay Papp apparent permeability coefficient
PBS phosphate buffered salts PCR polymerase chain reaction PEPT1 peptide transporter 1 P-gp P-glycoprotein RT reverse transcription
SGLT1 sodium-dependent glucose cotransporter 1 TEER transepithelial electrical resistance
TGF-β transforming growth factor-β UGT UDP-glucuronosyltransferase
UPLC-MS/MS ultra performance liquid chromatography-tandem mass spectrometer VPA valproic acid
目次
序論 ··· 1 第Ⅰ章 ヒトiPS 細胞由来小腸幹細胞の維持培養法の検討 ··· 3 1. 実験材料及び実験方法 ··· 4 1-1 細胞 ··· 4 1-2 試薬・培地・剥離液・保存液 ··· 41-3 MEF(Murine Embryonic Fibroblasts:マウス胚性線維芽細胞)の培養 ··· 4
1-4 ヒト iPS 細胞の培養 ··· 5 1-5 ヒト iPS 細胞の腸管上皮細胞への分化 ··· 6 1-6 腸管幹細胞の幹細胞性及び腸管細胞としての性質に与える添加因子の影響 ··· 6 1-7 ヒト iPS 細胞由来腸管幹細胞の継代と培養 ··· 9 1-8 ヒト iPS 細胞由来腸管幹細胞の腸管上皮細胞への分化 ··· 10 1-9 Real-time RT-PCR 法による解析 ··· 10 1-10 免疫蛍光染色 ··· 11 1-11 透過型電子顕微鏡(TEM)観察 ··· 12 1-12 走査型電子顕微鏡(SEM)観察 ··· 12 1-13 CYP3A4 及び P-gp 誘導能評価 ··· 13 1-14 CYP3A4 活性評価 ··· 13 1-15 膜透過性試験 ··· 13 1-16 統計解析 ··· 14 2. 結果 ··· 14 2-1 ヒト iPS 細胞由来の腸管幹細胞における FBS 及び KSR の幹細胞性及び腸管細胞と しての性質に及ぼす影響 ··· 14
2-2 腸管幹細胞における LGR5 及び CDX2 mRNA レベルに対する CHIR99021 及び VPA の影響 ··· 15 2-3 CHIR99021 及び VPA が腸管幹細胞増殖に及ぼす影響 ··· 16 2-4 継代を重ねた腸管幹細胞における LGR5、Ki67、及び CDX2 の mRNA 及びタンパク 質発現量の評価 ··· 18 2-5 継代を重ねた腸管幹細胞から分化させた腸管上皮細胞における分化マーカーの mRNA 及びタンパク質発現量の評価 ··· 19 2-6 ヒト iPS 細胞由来腸管上皮細胞におけるリファンピシン及び 1α, 25-Dihydroxyvitamin D3(VD3)による CYP3A4 及び P-gp の誘導能評価 ··· 21
Ⅱ 2-7 腸管上皮細胞における CYP3A4 活性評価 ··· 22 2-8 腸管上皮細胞単層膜における Rhodamine 123 及び Hoechst 33342 の透過性 ··· 23 3. 考察 ··· 26 4. 小括 ··· 28 第Ⅱ章 ヒトiPS 細胞由来腸管上皮細胞の粘膜傷害作用及び粘膜保護作用評価への利用 · 29 1. 実験材料及び実験方法 ··· 30 1-1 細胞 ··· 30 1-2 試薬・培地・剥離液・保存液 ··· 30 1-3 MEF の培養、MMC 処理 ··· 30 1-4 ヒト iPS 細胞の培養 ··· 30 1-5 ヒト iPS 細胞の腸管上皮細胞への分化 ··· 30 1-6 NSAIDs 及び MPAs の腸管上皮細胞への影響 ··· 30 1-7 Real-time RT-PCR 法による解析 ··· 30 1-8 免疫蛍光染色及びタンパク質発現量の定量解析 ··· 30 1-9 膜透過性試験 ··· 31 1-10 統計解析 ··· 31 2. 結果 ··· 31 2-1 ヒト iPS 細胞由来腸管上皮細胞における MUC2 発現量の評価 ··· 31
2-2 NSAIDs 及び MPAs の MUC2 及び villin の mRNA 発現量に与える影響 ··· 32
2-3 インドメタシン及びレバミピドの MUC2 のタンパク質発現量に与える影響 ··· 33
2-4 NSAIDs 及び MPAs の COX-2 及びサイトカイン発現量に与える影響 ··· 35
2-5 NSAIDs 及び MPAs の Pappに与える影響 ··· 36
3. 考察 ··· 37
4. 小括 ··· 39
総括 ··· 40
謝辞 ··· 41
【序論】 医薬品には様々な剤形が存在するが、経口製剤は臨床で使用されている剤形の60%強を占 めていることから最も重要な投与剤形であると言える。経口投与された薬物は、主に小腸で 吸収され、門脈、肝臓を経て、全身循環系に入る。そのため、経口製剤には、初回通過効果 によるbioavailability(BA)の低下や、その個体間変動が、薬物療法上、極めて重要な問題と なる。経口製剤のBAが低い主な原因として、胃腸内液での薬物の溶解性や安定性、小腸粘 膜透過性など、薬物の物理化学的特性の問題、もしくは肝臓における初回通過効果によるも のが大きいと考えられている。しかし、これらの原因に加え、消化管におけるphaseI、Ⅱ代 謝や、小腸上皮細胞に発現する排出トランスポーターによる小腸管腔中への薬物の能動的 排出が、経口投与後の薬物のBAを左右する重要な因子や相互作用の因子となることが指摘 されている1-6。中でも、cytochrome P450 3A(CYP3A)と多剤排出トランスポーターである P-glycoprotein(P-gp)は、基質認識性が極めて広く、組織局在性も非常に類似していることか ら、薬物消化管吸収の機能的障壁として働くことでBAを制御していると考えられる。しか しながら、小腸での初回通過効果に与える両タンパク質の働きについては、対象となる薬物 や用いられる評価系により異なった見解が示されており、一定の評価を得るに至っていな いのが現状である。したがって、経口製剤投与後の薬物の小腸での薬物動態を把握すること は、効果的かつ安全な薬物療法を行う上で極めて重要である。 現在、薬物の消化管吸収を評価する方法として、メンブレンフィルターに塗布形成したリ ン脂質膜を生体膜の代替と考え、膜透過性を評価する人工膜透過性試験(PAMPA:Parallel Artificial Membrane Permeation Assay)、ヒト結腸腺癌由来細胞であるCaco-2細胞や動物組織 を用いたin vitro試験がある。PAMPAは受動拡散のみを評価する評価系であり、薬物代謝酵 素やトランスポーターの評価は不可能である。薬物の上皮細胞透過性をin vitro試験として評 価し、ヒト経口吸収率を予測する系として現在最も一般的に実施されているのがCaco-2 細 胞を用いた透過性試験である。Caco-2細胞は培養すると小腸の円柱上皮細胞に似た刷子縁や タイトジャンクションの形態学的特徴を示し、単層膜を形成するが、薬物代謝酵素の発現が ほとんど認められない。また、薬物トランスポーターの発現パターンはヒト小腸とは異なる といった問題がある。このように現在用いられている評価系には、薬物代謝酵素やトランス ポーターの発現量、活性等の種差を含めた多くの問題が存在し、ヒトの消化管吸収を正確に 予測することは困難である。小腸における薬物動態評価には初代小腸上皮細胞の利用が望 ましいと考えられるが、その入手には制限がある。
そこで、ヒト胚性幹細胞(embryonic stem cells:ES細胞)と同様の多分化能とほぼ無限の 増殖能を有するヒト人工多能性幹細胞(induced pluripotent stem cells:iPS細胞)から作製さ れる腸管上皮細胞の利用が期待されている。ヒトiPS細胞は、ウイルスベクターを用いて体 細胞に4つの遺伝子、OCT3/4、SOX2、KLF4、c-MYCを導入することによって樹立された7。
2 とほぼ無限の増殖能をもつ細胞である8。当研究室では、これまでヒトiPS細胞から分化させ た腸管上皮細胞の有用性を報告してきた9-12。しかしながら、ヒトiPS細胞から腸管上皮細胞 まで分化させるには多くの時間と費用が必要であることが課題として挙げられている。ま た、ヒトiPS細胞由来腸管上皮細胞を薬物動態試験へ利用するためには、大量かつ安定的に 供給することも必須である。 そこで、本研究の第I章では、ヒトiPS細胞から腸管上皮細胞へ分化させる途中の段階であ る腸管幹細胞を幹細胞の性質を維持させたまま長期培養可能な方法の検討を行った。さら に、維持培養した腸管幹細胞から分化させた腸管上皮細胞が薬物動態学的な機能を有して いるかについても検討を行った。 本研究の第Ⅱ章では、ヒトiPS細胞由来腸管上皮細胞の毒性又は薬理評価への利用として、 薬物に起因する腸管粘膜傷害作用及び粘膜保護作用を評価可能なモデル細胞としての有用 性を検討した。上述した様に腸管の主要な働きは摂取した食物や栄養素の消化・吸収・代謝・ 排泄である。しかしながら、腸管は免疫器官としても極めて重要な役割を担っている。食物 の摂取とともに外来微生物が通過し、膨大な数の微生物が共生する消化管の表層には、それ ら微生物から腸管組織を保護する、または常在する微生物群に対する過剰な免疫反応を回 避するために、腸管上皮によって形成される粘膜層といわれる障壁が存在する。その粘液層 は腸管上皮細胞の一つである杯細胞から産生される糖タンパク質であるムチンによって構 成され、腸管上皮を覆うことで物理的に腸管組織への微生物の侵入を防止している。臨床で はムチンの質や量の低下が、粘膜傷害、炎症性疾患、潰瘍性大腸炎等、様々な疾患において 認められている。腸管ムチンは杯細胞から分泌されるmucin 2(MUC2)が主な主成分であり 13,14、MUC2の発現量は薬物による影響においても増減することが報告されている。例えば、
動物を用いたin vivo試験では、抗炎症剤である非ステロイド性抗炎症薬(non-steroidal anti-inflammatory drugs: NSAIDs)は胃粘膜だけでなく腸管粘膜の傷害も引き起こし、その際、腸 管粘膜のMUC2の発現量が減少していたことが報告されている15,16。臨床においては、
NSAIDsのMUC2の発現量に関する報告はないが、動物と同様にNSAIDsは胃粘膜だけでなく 腸管粘膜の傷害を引き起こしていることが明らかとなっている17。潰瘍性大腸炎は杯細胞が
減少することが知られており、疾患の発症や進展の要因の一つである可能性が考えられて いる。また、クローン病においてはMUC2の発現量が減少することも報告されている。一方、 レバミピドやイルソグラジンなどの胃粘膜保護薬(mucosal protective agents: MPAs)は胃粘 膜だけでなく腸管粘膜の保護作用も有しており、MUC2の発現量を増加させることが報告さ れている16,18-20。以上の報告から、MUC2は粘膜傷害作用や粘膜保護作用を評価する際の指 標として有用であると考えられ、これまでMUC2を指標として粘膜傷害作用を評価した結果 はいくつか報告されている15,21。しかしながら、これらの評価は主に動物によるin vivo試験 であり、in vitro試験で評価された報告は少ない。そこで、本研究の第Ⅱ章ではヒトiPS細胞 由来の腸管上皮細胞が、薬物に起因する粘膜傷害や粘膜保護を評価できるモデル細胞とし て有用かを検討した。以下、2章にわたり得られた知見を論述する。
第I 章 ヒト iPS 細胞由来腸管幹細胞の維持培養法の検討 自己複製能と多分化能を有するヒト iPS 細胞から作製される腸管上皮細胞は医薬品開発 や再生医療などへの応用が期待されている。医薬品開発では、腸管での薬物動態を評価する 細胞としての利用が考えられ、当研究室で構築したヒトiPS 細胞由来腸管上皮細胞は、薬物 動態関連遺伝子の顕著な発現増加、CYP3A4 の発現誘導、CYP3A4 代謝活性、取り込み及び 排泄トランスポーターによる輸送活性を有しており、ヒトiPS 細胞由来腸管上皮細胞の有用 性が確認されている9-12。しかしながら、ヒトiPS 細胞から腸管上皮細胞への分化は、内胚 葉、腸管幹細胞を経由する分化誘導に3 週間以上の時間を要し、その供給量やコストが問題 となっている。さらに、スクリーニング系として利用するためには高品質で均質な細胞を大 量に供給する必要がある。これらの問題の解決方法の一つとして、腸管上皮細胞へ分化させ る前段階である腸管幹細胞の維持・培養方法の開発が考えられる。分化の途中である腸管幹 細胞を維持培養することが可能となれば、供給までの期間の短縮と大量供給が可能になる と考えられる。しかしながら、腸管幹細胞はその特性から維持培養を可能とした報告は少な い。その理由としては、腸管幹細胞は腸管上皮の陰窩底部に局在し、ニッチと呼ばれる陰窩 底部に位置する腸管幹細胞の機能を保つ微小環境でパネート細胞と隣接することで幹細胞 性が維持されている点にある。このような特殊な環境にある腸管幹細胞を単離して培養す ることは不可能であると考えられていた。しかしながら、佐藤らは、幹細胞性を維持するた めに必須ファクターとして、Wnt signal、epidermal growth factor(EGF)signal の agonist とし てそれぞれWnt 3a、EGF を、Wnt signal の増強因子として R-spondin 1 を、幹細胞の分化抑 制因子であるbone morphogenetic protein(BMP)antagonist の Noggin を培地に添加すること で、腸のクリプト構造を再現した形態で体外培養が可能であることを示し、腸管上皮オルガ ノイドと呼ばれる形態で腸管細胞の維持培養に成功したことを報告した22。これらの腸管上 皮オルガノイドは継代が可能であり、遺伝子変異などを生じずにもとの性質や形態を維持 することが可能である。これらの報告以外にも、近年、生体の腸管から腸管幹細胞を単離し、 体外で腸管上皮細胞を長期に維持培養する技術が多く報告されている 23,24。しかしながら、 報告されているほぼすべての培養手法は操作が煩雑で、薬物動態試験に供するには不向き であるマトリゲルなどを用いた 3 次元培養法であり、維持培養した後の細胞の薬物動態学 的評価もほとんど行われていない。さらに、これまでの報告は、ヒトや動物由来の腸管細胞 を使用した報告がほとんどである。そこで、本研究ではこれまでの報告を元にヒトiPS 細胞 由来腸管幹細胞を平面で維持培養する方法の検討を行った。さらに、維持した腸管幹細胞か ら分化させた腸管上皮細胞が薬物動態評価に利用可能かについても検討した。
4 1. 実験材料及び実験方法 1-1 細胞 ヒトiPS 細胞(#51:Windy)は、ヒト胎児肺線維芽細胞 MRC-5 にパントロピックレトロ ウイルスベクターを用いてOCT3/4、SOX2、KLF4、c-MYC を導入後、クローン化した細胞 であり、国立成育医療研究センターの梅澤明弘博士よりご供与いただいた。Caco-2 細胞は 理化学研究所バイオソースセンター(茨城)より提供をうけた。 1-2 試薬・培地・剥離液・保存液
Fibroblast growth factor (FGF)2 及び activin A は PeproTech(Rocky Hill,NJ,USA)より、 iMatrix-511 は Nippi(Tokyo,Japan)より、fetal bovine serum(FBS)は Biowest(Nuaille,France) より、CHIR99021 及び Y-27632 は Focus Biomolecules(Plymouth Meeting,PA,USA)より、 KnockOut™ Serum Replacement(KSR)、DMEM、DMEM/F-12、Advanced DMEM/F-12、N2 supplement 及び B27 serum-free supplement は Thermo Fisher Scientific(Waltham,MA,USA) より、R-spondin 1、Noggin 及び EGF は GenScript(Piscataway,NJ,USA)より、N-acetylcysteine はSigma-Aldrich(St. Louis,MO,USA)より、8-Br-cAMP は Enzo Life Sciences(New York, NY,USA)より、PD98059 及び A 83-01 は AdooQ BIOSCIENCE(Irvine,CA,USA)より、5-aza-2’-deoxycytidine は Chem-impex International(Wood Dale,IL,USA)より、Dulbecco’s modified Eagle’s medium(DMEM)(High Glucose)、L-Glu、minimum essential medium nonessential amino acid solution (NEAA)、penicillin-streptomycin solution、valproic acid(VPA)、nicotinamide、ベ
ラパミル
、リファンピシン及び
1α,25-dihydroxyvitamin D
3は和光純薬工業(株)(Osaka,Japan)より、total RNA from human small intestine samples (5 donors)は BioChain Institute(Newark, CA,USA)より、SB202190 は ChemScene(Monmouth Junction,NJ,USA)より、WNT3a は ATGen(Gyeonggi-do,South Korea)より、Dulbecco’s phosphate buffered saline without calcium、 magnesium(D-PBS(-))用錠剤は DS Pharma Biomedical(Osaka,Japan)より、CELLBANKER は十慈フィールド(Tokyo,Japan)より、Accutase は MS TECHONOSYSTEMS(Osaka,Japan) より購入した。その他の試薬はすべて市販の特級品又は同等の製品を用いた。
1-3 MEF(Murine Embryonic Fibroblasts:マウス胚性線維芽細胞)の培養 MEF 培地
10% FBS、2 mM L-Glu、0.1 mM NEAA、100 units/mL penicillin G、100 μg/mL streptomycin sulfate を含む DMEM(High Glucose)を用いた。また、培養にはゼラチンコーティングディ ッシュを使用した。
MEF の調製
妊娠14.5 日目の ICR マウスをセボフランにて吸入麻酔後、頸椎脱臼により安楽死させた。 子宮から胎仔を摘出し、D-PBS(-)で洗浄後、胎仔の頭部、手足、尾部、臓器を除去し、再び
D-PBS(-)で洗浄した後、細かく剪断した。0.25% trypsin-EDTA を加えて 37°Cの水浴で温め、 MEF 用培地を加えて 100 μm セルストレーナーで濾過した。160 × g で 5 分間遠心後、上清 を吸引除去した。MEF 用培地で懸濁し、CO2インキュベーター中で培養を開始した。 MEF の継代 培養液を吸引除去し、D-PBS(-)で洗浄した。0.05% Trypsin-EDTA で細胞を剝離後、培地で 細胞を回収し、1,000 rpm(160 × g)で 5 分間遠心した。上清を吸引除去して MEF 用培地で 懸濁した後、ゼラチンコーティングディッシュ上に播種し、CO2インキュベーター内で培養 した。 MEF の MitomycinC 処理
MEF に 10 μg/mL Mitomycin C(MMC)を含む MEF 用培地を加え、37°Cにて90 分間処理 した。その後MMC 含有 MEF 用培地を除去し、D-PBS(-)で洗浄し、MEF 用培地を加え、CO2
インキュベーター内で3 時間から一晩静置した。 MMC 処理 MEF の凍結保存 培養液を吸引除去し、D-PBS(-)で洗浄した。0.05% Trypsin-EDTA で細胞を剥離後、MEF 用 培地を加え、細胞懸濁液を回収した。1,000 rpm(160 × g)で 5 分間遠心後、上清を吸引除去 した。CELLBANKER で懸濁し、-80°Cで保存した。 1-4 ヒト iPS 細胞の培養 ヒトiPS 細胞の培地調製
最終濃度として20% KSR、1% NEAA、2 mM L-Glu、0.1 mM 2-mercaptoethanol 含む DMEM/F-12 を用いた。また、FGF2 を 5 ng/mL となるように用時添加した。
フィーダー細胞の播種
ゼラチンコーティングディッシュに、MEF の細胞数が 6 × 10 5 cells/10 cm dish となるよう
に播種した。播種後、3 時間以上 5% CO2/95% air 条件下 CO2インキュベーター中37°Cにて 培養したものをフィーダー細胞として用いた。 ヒトiPS 細胞の解凍 凍結したヒトiPS 細胞を iPS 細胞用培地で融解し、細胞懸濁液を 1,000 rpm(160 × g)で 5 分間遠心した。上清を吸引除去してヒト iPS 細胞用培地で懸濁した後、フィーダー細胞を 播種したディッシュ上に播種した。培地には5 ng/mL となるように FGF2 を添加し、さらに 10 µM となるように Y-27632 を添加した。CO2インキュベーター中で培養開始し、細胞播種 48 時間後に 5 ng/mL FGF2 を含むヒト iPS 細胞用培地に交換した。それ以降は毎日培地交換
来ることを見出している。これらの3 つの因子は胃、肝臓などの他の消化器上皮細胞由来の オルガノイド培養にも用いられ、消化器上皮に共通したニッチ因子であると考えられる。 EGF は幹細胞に限らず、全般的な上皮細胞に対する増殖因子であり、腸管上皮細胞の増殖 に必須である。Noggin は BMP の阻害タンパク質であり、その過剰発現による異所性の陰窩 形成作用が報告されている。R-spondin 1 は leucine-rich repeat-containing G-protein coupled receptor 5(LGR5)のリガンドであり、Wnt シグナルの効率的な活性を誘導する因子である。 ヒトの腸管上皮細胞では、これらの3 つの因子に加え Wnt 3a、ストレス応答 mitogen-activated protein kinase(MAPK)である p38 の阻害及び transforming growth factor(TGF)-β シグナル の抑制が重要であることが報告されていることから22、p38 MAPK 阻害薬である SB202190、
TGF-β 阻害薬である A 83-01 を添加因子としてこれらを用いることとした。さらに、腸管幹 細胞の幹細胞性の維持に有効であることが報告されているTable 1 に示す液性因子、低分子 化合物を添加因子として用いた。
Table 1
Constituents of intestinal stem culture media and their working mechanisms and
effects
Medium constituents
Concentrations Working mechanism in ISCs
Effect on ISCs and application
EGF 100 ng/mL
Activates
RAS/RAF/MEK/ERK signaling
Stimulates stem cell migration, proliferation, and inhibits apoptosis
Noggin 100 ng/mL Inhibits BMP signaling Stimulates crypt formation R-spondin 1 100 ng/mL Augments WNT/β‐
catenin signaling
Stimulates crypt cell proliferation and maintains stem cell state FGF2 30 ng/mL supports FGFR/Erk
signaling
Enhances intestinal stem cell survival
Wnt 3a 100 ng/mL Activates canonical Wnt
signaling
Ligand for members of the frizzled family
SB202190 10 μM Inhibits P38 MAPK
Inhibits secretory differentiation、 increases plating efficiency, and decreases degradation of the EGF receptor
A 83-01 1.5 μM Inhibits TGF-β signaling Inhibits differentiation Nicotinamide 10 mM Inhibits the activity of
8
Medium constituents
Concentrations Working mechanism in ISCs
Effect on ISCs and application
N-acetyl-systein 1 mM
Scavenges ROS and partially via ERK1/2 activation
Protects cells from in vitro stress N2 suplement 1X Serum-free supplement Maintains stem cell state B27 suplement 1X Serum-free supplement Maintains stem cell state
Y-27632 10 μM Inhibits caspase-3 Prevents anoikis after single‐cell dissociation
FBS 及び KSR が幹細胞性及び腸管細胞としての性質に与える影響
ヒトiPS 細胞から分化させた腸管幹細胞(Day 7)の培養液を吸引除去し、D-PBS(-)5 mL で2 回洗浄した。Accutase にて剥離し、Table 1 に示す因子を添加した Advanced DMEM/F-12 を用いて0.5 µg/cm2となるようにiMatrix-511でコーティングした24 ウェルプレート上に播
種し、異なる濃度のFBS(5%、10%、20%)または KSR(5%、10%、20%)で 3 日間培養し た。培養終了後、細胞を回収し、LGR5 及び caudal-related homeobox 2(CDX2)の mRNA 発 現量を測定した。
CHIR99021 及び VPA が幹細胞性、腸管細胞としての性質及び細胞増殖に与える影響 ヒトiPS 細胞から分化させた腸管幹細胞(Day 7)の培養液を吸引除去し、D-PBS(-)5 mL で2 回洗浄した。アクターゼにて剥離し、Table 1 に示す因子を添加した Advanced
DMEM/F-12 を用いて0.5 µg/cm2となるようにiMatrix-511 でコーティングした24 ウェルプレート上
に播種し、異なる濃度のCHIR99021(1、3、10、30、60、100 μM)又は VPA(0.1、0.3、1、 3、10 mM)で 3 日間培養した。培養終了後、細胞を回収し、LGR5 及び CDX2 の mRNA 発
現量を測定した。また、同様の方法でCHIR99021 及び VPA の影響を検討し、自動セルカウ
ンター(Countess,Thermo Fisher Scientific)を用いて細胞数を計数した。コントロールは溶
媒であるジメチルスルホキシドで処理した。なお、FBS 及び KSR が幹細胞性及び腸管細胞
としての性質に与える影響の検討結果から、以降の実験はKSR の濃度を 10%とした。
CHIR99021:GSK-3β 阻害薬の中でも非常に強力な選択的 GSK-3β 阻害薬である。GSK-3β が 阻害されることで,β カテニンが蓄積され,核内移行しやすくなることで Wnt シグナルが活性化される。 Valproic acid:ヒストン脱アセチル化酵素阻害剤である。Notch シグナルを活性化することが 報告されているが、作用機序は不明である。 1-7 ヒト iPS 細胞由来腸管幹細胞の継代と培養
ヒトiPS 細胞から分化させた腸管幹細胞(Day 7:passage 0(P0))の培養液を吸引除去し、 D-PBS(-)5 mL で 2 回洗浄した。アクターゼにて剥離し、細胞懸濁液を 15 mL 遠沈管に回 収した。1,000 rpm(160 × g)で 3 分間遠心後、上清を可能な限り吸引除去した。Table 2 に
示す因子を添加したAdvanced DMEM/F-12 を用いて0.5 µg/cm2となるようにiMatrix-511で
コーティングしたディッシュに継代した。継代後の最初の24 時間は Y-27632(10 μM)を添 加し、続いて2-3 日ごとに培地を交換した。その後の継代は、細胞が培養ディッシュに対し 占める割合が約80 から 90%になった状態で開始した。継代には継代前のディッシュ 1 枚当 たり約30%の細胞を使用し、約 10%の細胞は総リボ核酸(RNA)抽出に使用した。継代は 7 回繰り返し、RNA 抽出後、real-time RT-PCR 法による解析を行った。また、P 0 及び P 3 の 腸管幹細胞については、免疫蛍光染色を行った。
Table 2. Summary of compounds used in intestinal stem cell culture
Factor or Compounds Concentration
KSR 10% CHIR99021 10 μM VPA 1 mM EGF 100 ng/mL Noggin 100 ng/mL R-spondin 1 100 ng/mL FGF2 30 ng/mL Wnt3a 100 ng/mL SB202190 10 μM A 83-01 1.5 μM Nicotinamide 10 mM N-acetylcysteine 1 mM N-2 Supplement (100X) 1X B-27 Supplement (50X) 1X Y-27632 10 μM
Table 3. Sequences of primers for real-time PCR analysis
Gene name
Forward primer sequence (5'→3')
Reverse primer sequence (5'→3')
LGR5 AGAAGGCACCGTATATGACGAC CAGCGTTACAGACACACTGCTC
CDX2 AGCCAGATCACTGCTGAGGT TGGACAGGTGTTCCTCCTTC
Ki67 GACTTTGGGTGCGACTTGAC ACCCCGCTCCTTTTGATAGT
Villin GTGGCAATGAGGATGACTTGTTC TAGTGGTGGTCGGAGATTCGTA
P-gp GGCAGTCAGATCATCTTCTCG GCTGGTTATCTCTCAGCTCCAC
BCRP CTGGAAGCACGAATGACAGA CCTTCTGCTTGCAAATAGGC
CYP3A4 CTGTGTGTTTCCAAGAGAAGTTAC TGCATCAATTTCCTCCTGCAG
UGT1A1 CAGCAGAGGGGACATGAAAT ACGCTGCAGGAAAGAATCAT
MUC2 AGAAGGCACCGTATATGACGAC CAGCGTTACAGACACACTGCTC
OATP2B1 CTTCATCTCGGAGCCATACC GCTTGAGCAGTTGCCATTG
Sucrase–
isomaltase GGTAAGGAGAAACCGGGAAG GCACGTCGACCTATGGAAAT PEPT1 CACCTCCTTGAAGAAGATGGCA GGGAAGACTGGAAGAGTTTTATCG
CYP2C9 GACATGAACAACCCTCAGGACTTT TGCTTGTCGTCTCTGTCCCA
CYP2C19 GAACACCAAGAATCGATGGACA TCAGCAGGAGAAGGAGAGCATA
SGLT1 CAACATCGCCTATCCAACCT TAAACAACCTTCCGGCAATC HPRT1 CTTTGCTTTCCTTGGTCAGG TCAAGGGCATATCCTACAACA 1-10 免疫蛍光染色 細胞をPBS(1 mM CaCl2及び1 mM MgCl2を含む)で2回洗浄した。次に-20°Cに冷却した メタノールを加え、4°Cで5分間処理し、固定と膜透過処理を行った。PBSで3回洗浄し、 0.5% FBSを含むPBSを用いて室温、20分間ブロッキングした。その後、一次抗体を室温で 60分間反応させた。PBSで細胞を3回洗浄し、希釈倍率(1:200)のAlexa Fluor 488及び568 でラベルされた2次抗体を含むPBSで室温、60分処理した。さらにPBSで3回洗浄し1 μg/mL 4',6-diamidino-2-phenylindole (DAPI)を含むPBSで室温、5分間反応させ、PBSで1回洗浄 した。最後に細胞を封入剤(SlowFade® 褪色防止用封入剤、Thermo Fisher Scientific)を用 いてスライドガラスの上にマウントした。細胞の観察はECLIPSE Ti-S microscope(ニコ ン,Tokyo,Japan)を用いて行った。Table 4に使用した一次抗体及び二次抗体の詳細を示 した。
12
Table 4. Primary and secondary antibodies
Antibody name Source Catalog number Biological source Dilution
LGR5 ABGENT AP2745d Rabbit 1:40
CDX2 BioGenex MU392A-UC Mouse 1:50
Ki67 eBioscience 14-5699 Mouse 1:100
BCRP Abcam Ab3380 Mouse 1:100
Villin Santa Cruz
Biotechnology sc58897 Mouse 1:100
E-Cadherin BD Transduction
Laboratories™ 610181 Mouse 1:100
Occludin Thermo Fisher
Scientific 71-1500 Rabbit 1:100
Alexa Fluor® 488 donkey anti-Rabbit
IgG (H&L)
Thermo Fisher
Scientific A-21206 Rabbit 1:200
Alexa Fluor® 488 goat anti-Mouse
IgG (H&L)
Thermo Fisher
Scientific A-11001 Mouse 1:200
Alexa Fluor® 568 goat anti-Mouse
IgG (H&L)
Thermo Fisher
Scientific A-11004 Mouse 1:200
Alexa Fluor® 568 goat anti-Rabbit
IgG (H&L)
Thermo Fisher
Scientific A-11011 Rabbit 1:200
1-11 透過型電子顕微鏡(TEM)観察 P0 及び P3 の腸管幹細胞をカルチャーインサートに播種後、腸管上皮細胞まで分化させ た。0.1 M の PBS で洗浄後、2.5%グルタルアルデヒドで 4°Cにて一晩固定し、0.1 M の PBS で洗浄した。続いて、4°Cにて1%四酸化オスミウムで 2 時間固定した後、エタノールで脱 水し、樹脂に包埋した。次いで、埋め込まれた試料を0.1 μm 切片に切断し、酢酸ウラニル で染色し、H7600 透過型電子顕微鏡(Hitachi,Tokyo,Japan)を用いて観察した。 1-12 走査型電子顕微鏡(SEM)観察 P0 及び P3 の腸管幹細胞をカルチャーインサートに播種後、腸管上皮細胞まで分化させ、 TEM と同様の方法で固定・脱水した。続いて、臨界点乾燥を CPD300(Leica,Wetzlar,Germany) を用いて行った。次いで、オスミウムコーティングをプラズマコーター(OPC80AJ,Filgen, Nagoya,Japan)を用いて行い、S-4800 顕微鏡(Hitachi,Tokyo,Japan)を用いて観察した。
1-13 CYP3A4 及び P-gp 誘導能評価
P0 及び P3 の腸管幹細胞から分化させた腸管上皮細胞の培地に終濃度として 30 nM の 1α, 25-dihydroxyvitamin D3(VD3)または 30 μM のリファンピシン(それぞれ CYP3A4 及び
P-gp 誘導因子)を添加し、48 時間処理した。コントロール群はジメチルスルホキシドで処理 した。細胞を回収後、total RNA を抽出し、CYP3A4 及び P-gp の mRNA 発現量を定量する ことで誘導能を評価した。 1-14 CYP3A4 活性評価 P0 及び P3 の腸管幹細胞から分化させた腸管上皮細胞を 150 μM のミダゾラムを含むハン クス平衡塩溶液(HBSS)と共に 37°Cで 6 時間インキュベートした。インキュベーション 後、200 μL の反応培地を採取し、内部標準として 3.6 μM のクロロプロパミドを含有する 200 μL の氷冷アセトニトリルを添加した。細胞は 150 μL の 1M NaOH で溶解し、等量の 1M HCl を加えて中和した。次いで、Pierce BCA タンパク質アッセイキット(Thermo Fisher Scientific) を用いてタンパク質含有量を測定した。1’-ヒドロキシミダゾラムは XBridge BEH C18 カラ ム(2.1×50 mm,3.5 μm,Waters)を備えた UPLC-MS/MS システム(Waters,Milford,MA, USA)を用いて測定した。サンプルは遠心分離(20,600 g,5 分間,4°C)し、10 μL の上清 をUPLC-MS/MS システムにて測定した。移動相は、10 mM のギ酸アンモニウム(A)及び アセトニトリル(B)を含み、カラム及び試料温度は、それぞれ 40°C及び4°Cに設定した。 移動相のグラジエント条件は次のように設定した。0–0.5 min: 5% B、0.5–2.0 min: 5→95% B (linear)、2.1–3.5 min: 95% B、3.6–5.4 min: 95→5% B (linear)。流速は 0.25 mL/min(2.1~3.5 min; 0.55 mL/min を除く)。1’-ヒドロキシミダゾラム(m/z 342.03→168.40)の検出は Multiple Reaction Monitoring(多重反応モニタリング)モードを用いた。データ解析は、Mass Lynx 4.1 ソフトウェア(Waters)を用いて行った。 1-15 膜透過性試験 膜抵抗値の測定 セルカルチャ ーインサート上で培養した 腸管上皮細胞の膜抵抗値 をMillicell ERS-2 (Millipore,Bedford,MA,USA)を用いて測定した。 P-gp 及び BCRP の輸送機能評価 それぞれのトランスポーターの基質としてP-gpは1 μMのRhodamin123、BCRPは10 μMの Hoechst33342を用い、吸収方向(apical側からbasal側への透過)及び排泄方向(basal側から apical側への透過)の経細胞輸送試験を行った。また、P-gp及びBCRPの選択的阻害薬として、 それぞれ10 μMのベラパミル及び1 μMのKo143を用いた。 腸管幹細胞を0.5 µg/cm2となるようにiMatrix-511でコーティングしたカルチャーインサート に播種後、19日間培養した後、apical及びbasal側の培地を除去し、緩衝液(ハンクス平衡塩溶
14 液、pH 7.4) を加え、37°Cで20分間プレインキュベーションした。プレインキュベーション 後、ドナー側に基質を添加し、膜透過試験を開始した。試験開始後、30、60、90及び120分 の時点でレシーバー側からサンプリングを行い、これを測定用サンプルとした。採取後は 同量の緩衝液を受給側に加えた。測定用サンプルの蛍光強度は蛍光プレートリーダー (BioTek,Winooski,VT,USA)を用い、Rhodamin123は励起波長:505 nm、蛍光波長:534 nm、Hoechst33342は励起波長:352 nm、蛍光波長:461 nmで測定を行った。 見かけの膜透過係数の算出 輸送試験の見かけの膜透過係数(Papp)は次の式で算出した。 Papp = (1/A × C0) (dQ/dt) dQ/dt は単位時間あたりに透過された薬物総量、Aはセルカルチャーインサート膜の表面積 (0.3 cm2)、C 0は供与側に添加した基質の初濃度を示す。Efflux ratio(ER)はbasal側からapical
側へのPappをapical側からbasa側へのPappで除すことで算出した。
1-16 統計解析 統計的有意性は、分散分析を行った後、ダンカン法を用いた。その際の統計分析は、EXCEL 統計Ver.4.0(株式会社エスミ,Tokyo,Japan)を用いて行った。 2 結果 2-1 ヒト iPS 細胞由来の腸管幹細胞における FBS 及び KSR の幹細胞性及び腸管細胞とし ての性質に及ぼす影響 FBS は細胞を培養する際の必須因子である。しかしながら、FBS は増殖因子や分化促進 因子、ホルモンなど未知の因子を含んでいることから幹細胞の培養には不向きであると考 えられる。そこで、腸管幹細胞培養におけるFBS の代替として、ES 細胞や iPS 細胞の培養 で用いられている血清不含の無血清サプリメントであるKSR を検討した。 その結果、腸管幹細胞マーカーとして知られる LGR525の mRNA 発現量は継代で大きく 減少し、FBS 濃度の増加に伴って減少した(Fig. 3)。KSR の場合も LGR5 発現量は減少した が、同濃度で比較した場合、KSR 添加群はすべての濃度において FBS 添加群よりも LGR5 発現量が高かった。腸管細胞マーカーであるCDX2 の mRNA 発現量も継代で大きく減少し たが、FBS 及び KSR を添加した際の CDX2 発現量は同様の変動が認められ、FBS と KSR の 間に大きな違いは認められなかった。これらの結果から、KSR は FBS に比べて腸管幹細胞 性への影響が小さく、FBS の代替として有用であると考えられた。したがって、LGR5 発現 量が高かった10%の添加濃度で以降の検討を行うこととした。
Figure 4.
Effect of FBS and KSR on LGR5 and CDX2 mRNA levels in intestinal stem cells
derived from human iPS cells
LGR5 and CDX2 mRNA levels are normalized to those of HPRT1 and are relative to levels in the human adult small intestine (set as 100).
Data represent the mean ± standard deviation (n = 3).
2-2 腸管幹細胞における LGR5 及び CDX2 mRNA レベルに対する CHIR99021 及び VPA の 影響 腸管幹細胞の幹細胞性の維持には Wnt 及び Notch シグナルの両方が重要であることがこ れまで報告されている26-29。そこで、本研究ではGSK-3β 媒介性 β-カテニン分解を阻害し、 Wnt シグナル伝達を活性化させる CHIR99021、Notch シグナルを活性化させる VPA を培地 に添加し、ヒトiPS 細胞由来腸管幹細胞における腸管幹細胞性及び腸管細胞としての性質に 対する影響を評価した。
その結果、CHIR99021 は LGR5 及び CDX2 の mRNA 発現量を有意に増加させた(Fig. 4)。
VPA は LGR5 の発現量を増加させたが、CDX2 の発現量にはほとんど影響を与えなかった。 10 μM の CHIR99021 及び 1 mM の VPA を同時に添加した場合は CHIR99021 単独で添加し
16
Figure 5.
Effect of CHIR99021 and VPA on CDX2 and LGR5 mRNA levels in intestinal stem
cells derived from human iPS cells
Human iPS cells were differentiated into intestinal stem cells (day 7), passaged on iMatrix511-coated 24-well plates, and cultured in the presence of varying concentrations of CHIR99021 and/or VPA for 3 days.
CDX2 and LGR5 mRNA levels are normalized to those of HPRT1 and are presented relative to levels in the human adult small intestine (set as 100). Target gene expression was normalized to HPRT1 levels.
Data represent the mean ± standard deviation (n = 3).
C0V0, concentrations of CHIR99021 and VPA (0); C10V1, concentrations of CHIR99021 and VPA (10 µM and 1 mM, respectively).
Levels of statistical significance were compared with the C0V0; **p < 0.01.
2-3 CHIR99021 及び VPA が腸管幹細胞増殖に及ぼす影響 医薬品開発にヒト iPS 細胞由来腸管上皮細胞を利用する場合、大量に供給する必要があ る。そのためには腸管幹細胞を多量に培養できることが重要である。そこで、CHIR99021 及 びVPA が腸管幹細胞の増殖に及ぼす影響を検討した。その結果、コントロール群(C0V0) と比較して、CHIR99021 の濃度が 10 μM までは細胞数は有意に増加し、約 2 倍の細胞数が 得られた。30 μM においてもコントロール群と比較して有意に増加したが、その程度は減少 した。一方、60 μM 以上ではコントロール群と比較して細胞数は有意に減少した。VPA は、
0.1 mM では細胞数は有意に増加し、約 1.5 倍の細胞数が得られた。0.3 及び 1 mM において もコントロール群と比較して有意に増加したが、その程度は減少した。一方、3 mM では、 コントロール群と比較して有意に減少し、6 mM 以上では顕著に減少した。10 μM CHIR99021 と1 mM VPA を併用した場合は、コントロール群の約 2 倍の細胞数が得られた。
Figure 6.
Effect of CHIR99021 and VPA on the growth of intestinal stem cells derived from
human iPS cells
Human iPS cells were differentiated into intestinal stem cells, passaged on iMatrix511-coated 24-well plates, and cultured in the presence of varying concentrations of CHIR99021 and/or VPA for 3 days. Cell number was counted using an automated cell counter.
Data represent the mean ± standard deviation (n = 3).
C0V0, concentrations of CHIR99021 and VPA (0); C10V1, concentrations of CHIR99021 and VPA (10 µM and 1 mM, respectively).
Levels of statistical significance were compared with the C0V0; **p < 0.01.
本検討において、CHIR99021 は、腸管幹細胞の幹細胞性及び腸管細胞としての性質を促 進させる効果が確認された。一方、VPA は高濃度(10 mM)では腸管幹細胞の幹細胞性を促 進させる効果は得られたが、高濃度では腸管幹細胞の増殖性を著しく低下させた。しかしな がら、幹細胞性を維持するために Notch シグナルが重要であることを示す報告がこれまで が多くなされていること26-29、10 μM CHIR99021、1 mM VPA を併用することでそれぞれを 単独で使用した場合と比べ、僅かではあるが腸管幹細胞性及び腸管細胞としての性質を促 進させる効果が得られたこと、さらに、細胞増殖を促進させる効果が得られたことから 10 μM CHIR99021 と 1 mM VPA を併用することで以降の研究を行った。
18 2-4 継代を重ねた腸管幹細胞における LGR5、Ki67、及び CDX2 の mRNA 及びタンパク質 発現量の評価 複数回継代した腸管幹細胞が腸幹細胞性及び腸管細胞としての性質を維持しているか検 討するために、LGR5、Ki67(幹細胞及び細胞増殖マーカー)及び CDX2 の mRNA 及びタン パク質の発現量を評価した。LGR5 及び Ki67 の mRNA 発現量は継代を経るに従い、減少傾 向が認められたものの、ヒト小腸と同程度以上の発現量が維持されていた。CDX2 は P0 や ヒト小腸の発現量に比べP1–P7 で高い発現量が維持されていた(Fig. 7)。 免疫蛍光染色においては、P0 腸管幹細胞において LGR5 及び Ki67 は検出されたが、CDX2 は検出されなかった。P3 腸管幹細胞においては LGR5、Ki67 及び CDX2 は検出され、多く の細胞がLGR5 陽性、Ki67 陽性及び CDX2 陽性であった(Fig. 8)。以上より、腸管幹細胞 性及び腸管細胞としての性質がmRNA 及びタンパク質レベルで維持されていることが示唆 された。
Figure 7.
Effects of passage number on LGR5, Ki67, and CDX2 mRNA levels in intestinal
stem cells derived from human iPS cells
Total RNA from repeatedly passaged intestinal stem cells (P0–P7) was extracted, and gene-expression levels were measured by real-time RT-PCR.
The expression levels of each gene were normalized to those of HPRT1 and are presented relative to levels in the human adult small intestine (set as 100).
Figure 8.
Immunofluorescence analysis of LGR5, Ki67, and CDX2 in P0 and P3 intestinal
stem cells
Intestinal stem cells (P0 and P3) were stained with LGR5 (green), Ki67 (red), and CDX2 (green). Scale bar, 100 μm. 2-5 継代を重ねた腸管幹細胞から分化させた腸管上皮細胞における分化マーカーの mRNA 及びタンパク質発現量の評価 複数回継代した腸管幹細胞が腸管上皮細胞に分化する能力を有しているかを検討するた めに、いくつかの腸管上皮細胞マーカーのmRNA 発現量を評価した(Fig. 9)。その結果、 P-gp 及び BCRP の mRNA 発現量はヒト小腸と比べて発現量は高く、P-gp は P7 腸管上皮細 胞まで、BCRP は P6 腸管上皮細胞まで P0 腸管上皮細胞と同程度の発現が維持されていた。 Villin、sucrase-isomartase 及び peptide transporter 1(PEPT1)の発現量は、P3 腸管上皮細胞ま で P0 腸 管 上 皮 細 胞 と 同 程 度 で あ っ た 。 CYP2C9 、 CYP2C19 、 CYP3A4 、 UDP-glucuronosyltransferase 1A1(UGT1A1)、organic anion-transporting polypeptide 2B1(OATP2B1)、 及びsodium-dependent glucose cotransporter 1(SGLT1)、MUC2 は継代を繰り返すにつれて減 少傾向を示した。
次に、複数回継代した腸管幹細胞の腸管上皮細胞への分化能をタンパク質レベルで評価 するために、BCRP、villin、occludin、及び E-cadherin の免疫蛍光染色及び腸管上皮細胞の電 子顕微鏡観察を行った(Fig. 10)。その結果、P0 及び P3 腸管上皮細胞で BCRP、villin、occludin、 及びE-cadherin が検出され、SEM 及び TEM によって各腸管上皮細胞の表面に微小絨毛が存 在することが確認された。さらに、P0 の腸管上皮細胞ではタイトジャンクションが観察さ れた。
20
Figure 9.
mRNA levels of intestinal and pharmacokinetics markers in enterocytes derived
from human iPS cells (P0–P7)
Human iPS cells were induced to differentiate into intestinal stem cells, which were repeatedly passaged and cultured, with induction into enterocytes at each passage. Total RNA from enterocytes (P0–P7) was extracted, and the gene-expression levels of specific markers were measured by real-time RT-PCR.
The expression levels of each gene were normalized to those of HPRT1 and are presented relative to levels in the human adult small intestine (set as 100).
Figure 10.
Immunofluorescence staining of differentiation markers in enterocytes (P0 and P3)
and SEM and TEM analyses
Intestinal stem cells (P0 and P3) were stained for (a, f) BCRP, (b, g) villin, and (c, h) occludin/E-cadherin. Scale bar, 100 μm, (d, e, i, j) SEM and TEM analyses of microvilli and (e) tight junctions. Scale bar, 500 nm. (d, i) Surface morphology of the enterocytes was visualized by SEM. Microvilli were detected in P0 and P3 enterocytes. (e, j) TEM analysis of enterocytes derived from human iPS cells and displaying enterocyte-like differentiation features. Tight junctions (TJs) were observed in P0 enterocytes. 2-6 ヒト iPS 細胞由来腸管上皮細胞におけるリファンピシン及び 1α, 25-dihydroxyvitamin D3(VD3)による CYP3A4 及び P-gp の誘導能評価 CYP3A4 及び P-gp は腸管上皮細胞に高発現しており、消化管での薬物吸収や薬物相互作 用に関わる重要な因子である。P0 から P3 までの腸管上皮細胞における CYP3A4 及び P-gp の誘導能をCYP3A4 及び P-gp の誘導剤であるリファンピシン及び VD3 を用いて評価した
22
(Fig. 11)。P0 から P3 の腸管上皮細胞において、CYP3A4 の mRNA 発現量は、コントロー ルに比べリファンピシン及びVD3 添加群でそれぞれ 1.2 倍–2.5 倍及び 4.1 倍–41.5 倍高かっ た。P-gp の mRNA 発現量は、リファンピシン添加群では 1.3 倍–2.9 倍に増加したが、VD3 添加群ではコントロール群と同程度であった。
Figure 11.
Induction of CYP3A4 and P-gp expression in enterocytes derived from human iPS
cells
Cells were treated with 1α,25-dihydroxyvitamin D3 (VD3) or rifampicin for 48 h after differentiation,
and total RNA from enterocytes (P0 and P3) was extracted, followed by real-time RT-PCR. Target gene expression was normalized to HPRT1 levels.
Data represent the mean ± standard deviation (n = 3).
2-7 腸管上皮細胞における CYP3A4 活性評価 腸管上皮細胞の培地に終濃度として CYP3A4 の基質である 150 μM のミダゾラムを添加 し、6 時間インキュベートした際の CYP3A4 の特異的代謝産物である 1’-ヒドロキシミダゾ ラムの生成量を評価した(Fig. 12)。1’-ヒドロキシミダゾラムの生成は P0 から P3 の腸管幹 細胞から分化させた腸管上皮細胞において確認された。またその生成量はP0 から P3 の腸 管上皮細胞の間で大きな違いは認められなかった。
Figure 12.
Formation of 1’-hydroxy midazolam in enterocytes derived from human iPS cells
Enterocytes were incubated with 150 μM midazolam for 6 h, and LC-MS/MS was used to measure the formation of 1’-hydroxy midazolam as a specific metabolite generated by CYP3A4.Data represent the mean ± standard deviation (n = 3).
2-8 腸管上皮細胞単層膜における Rhodamine 123 及び Hoechst 33342 の透過性 BCRP は P-gp と同様、小腸の上皮細胞の先端膜において高度に発現し、様々な薬物の腸 からの吸収及び組織分布を制限する重要な役割を果たしている輸送担体である。そこで、イ ンサート上に腸管幹細胞を播種し、腸管上皮細胞へ分化後、それぞれの輸送活性を評価した (Fig. 13)。 P-gp の基質である Rhodamin123 の排出方向への Papp値は、P0 及び P3 の腸管上皮細胞の 吸収方向のPapp値よりも高かった。ER は、それぞれ P0 及び P3 で 14.6 及び 7.2 であった。 また、阻害薬であるベラパミルを添加した場合、排出方向の Papp値は減少し、吸収方向の Papp値が上昇し、ER はそれぞれ 0.7 及び 1.0 となった。 BCRP の基質である Hoechst 33342 の排出方向への Papp値はP0 及び P3 の腸管上皮細胞の 吸収方向のPappよりも高かった。ER は、それぞれ P0 及び P3 で 11.6 及び 9.8 であった。
BCRP 阻害薬である Ko143 を添加した場合、排出方向への Papp値は減少し、吸収方向のPapp
値は上昇し、ER はそれぞれ 1.5 及び 1.4 に増加した。これらの結果から P0 及び P3 の腸管 上皮細胞がP-gp 及び BCRP の輸送活性を有することが示された。また、膜抵抗値は P0 に 比べてP3 で低い結果が得られた(Fig. 14)。
24
Figure 13.
Permeability of Rhodamine 123 and Hoechst 33342 across enterocyte monolayers
Enterocytes were seeded on iMatrix511-coated cell-culture inserts and incubated with HBSS (pH 7.4) containing Rhodamine 123 (substrate for P-gp; 1 μM) or Hoechst 33342 (substrate for BCRP; 20 μM) for 120 min at 37°C and in the presence or absence of inhibitor (verapamil (P-gp inhibitor): 30 μM; Ko143 (BCRP inhibitor): 1 μM). The ER for Rhodamine 123 or Hoechst 33342 was derived from the Papp values associated with basal-to-apical transport and apical-to-basal transport.Figure 14.
Time-dependent changes of TEER values in the enterocyte-like cell layer
The enterocyte-like cells were seeded on iMatrix-511-coated cell culture inserts. TEER values were measured every three days from day 4 after seeding.