平成
29 年度
学位論文(博士)
意思決定に関わる
大脳基底核の神経基盤
平成
29 年 9 月 21 日提出
玉川大学大学院脳情報研究科
脳情報専攻
学籍番号
112715006
野々村 聡
2
要旨
ヒトや動物の思考,認知などのいわゆる高次脳機能は,脳でどのように実現 されているのであろうか.本論文では,多様な脳の高次機能の中でも「意思決 定 decision-making」に焦点を当て,霊長類(サル)とげっ歯類(ラット)を用 いた電気生理学的な研究を行うことで,線条体を中心とした皮質-基底核ループ で実現されている高次情報処理のしくみを明らかにすることを目的としている. 本論文は,霊長類の神経細胞活動記録とげっ歯類の膜電位記録の2つの実験 について2部の構成で記述する.霊長類の実験では,運動成分を含まない選択 肢に対して行う「対象の選択」に線条体がどのように関わるのかを調べるため に,運動要素を含まない対象の選択と,実際に行う運動の選択とを時間的に分 離した課題を考案し,課題遂行中のニホンザルの広範な線条体領域から単一神 経細胞活動の記録を行った.その結果,対象の選択に関わる情報表現は線条体 内でも吻側の尾状核に偏っていることが明らかになった.げっ歯類の実験では, 頭部固定下でレバー運動中のラットを用いて一次運動野と背外側線条体それぞ れでホールセル記録を行い,運動実行を行う際の膜電位変動とスパイクとの関 係を解析した.その結果,一次運動野・線条体どちらの領域においても,スパ イクは, 膜電位の揺らぎではなくむしろ膜電位そのものが安定的に変化するこ とによって生じる性質があることが明らかになった.これら神経細胞レベルで3 の意思決定研究で得た結果は,さまざまなレベルでの意思決定に関わる皮質-基 底核ループの役割を明らかにするための重要な基礎的所見であり,将来的には, 多様な症状を呈する大脳基底核疾患の神経メカニズムの解明にもつながると考 えられる.
4
目次
第 1 章 序論 1.1. はじめに ... 6 1.2. 高次脳機能研究と単一神経細胞活動記録法 ... 6 1.3. 大脳基底核の解剖と機能 ... 7 1.4. 線条体と価値に基づく意思決定 ... 11 1.5. 中央実行系とそれに関連する脳領域 ... 12 1.6. 本論文の目的 ... 13第 2 章 Monkey study : Neuronal activities in striatum during object-decision-making 2.1. 導入 ... 15 2.2. 対象と方法 ... 18 2.2.1. 被検体および実験装置 ... 18 2.2.2. 行動課題 ... 19 2.2.3. 課題訓練と手術 ... 21 2.2.4. 細胞活動の記録とデータ収集 ... 21 2.2.1. データ解析 ... 23 2.3. 結果 ... 26 2.3.1. 行動選択および反応時間 ... 26 2.3.2. 線条体の課題関連細胞活動のデータベース ... 32 2.3.3. 対象の選択をしていることを示す集団細胞活動 ... 34 2.3.4. 対象の選択時期における各細胞の情報表現 ... 37
2.3.5. 対象の選択時期における Offered stimulus と Chosen stimulus の情報 表現の線条体内分布 ... 41
5
第 3 章 Rat study:Continuous membrane potential fluctuations in motor cortex and striatum neurons during voluntary forelimb movements and pauses
3.1. 導入 ... 54 3.2. 対象と方法 ... 57 3.3. 結果 ... 60 3.4. 考察 ... 70 第 4 章 考察 4.1. 線条体特有の情報処理の可能性 ... 71 4.2. 結論 ... 79 謝辞 ... 81 引用文献 ... 82 研究実績 ... 94
6
第
1 章
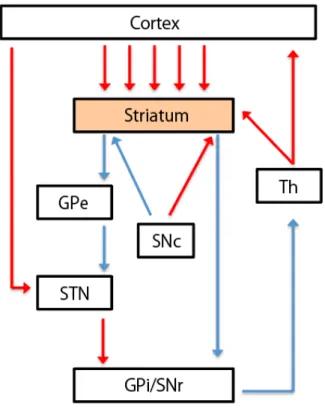
1.1. はじめに 変動する環境の中で最適な行動を選択し実行することは,我々が生き残るための 本質的な脳機能である.心理学,経済学,神経科学分野では,行動選択を,「脳が, 始めに複数ある選択肢の価値を計算し,その後に最も望ましい価値を選択する」一連 の過程を指す概念として提唱している1, 2, 3.神経科学研究においては,前世紀末に開 発された単一神経細胞活動記録によって 4,近年までに,価値に基づく意思決定に関 わる脳内信号を検出した知見が数多く積み重ねられてきている.こうした知見は,大脳 基底核が価値に基づく意思決定に非常に重要な領域であることを示している.また, 大脳基底核は,皮質-基底核ループを介して、前頭前野を含む前頭葉と相互作用す ることによって、意思決定に関連した情報処理を行っている可能性があることも示唆さ れている 9.本論文では,価値に基づく意思決定に関わる,線条体を中心とした皮質— 基底核ループの情報処理のしくみを調べるために,サルまたはラットを被検体とし,行 動課題遂行中の細胞活動を記録することで,神経細胞レベルでの情報処理のしくみ に迫った. 1.2. 高次脳機能研究と単一神経細胞活動記録法 高次脳機能の神経メカニズムの研究は,1968 年に発表された Edward Evarts らによる霊長類動物(サル)を使った研究に端を発する4.この研究では,頭部7 固定下で,予め訓練された運動課題を遂行中のサルの一次運動野から,単一神 経細胞活動の記録を行うというものであった.この記録法は,電極を直接目的 の脳領域に刺入することによって行動中の動物の脳活動を高い時間分解能で捉 えることができ,進化上ヒトに近い霊長類 (サル)を被験体として使うことで, 運動中の細胞活動のみならず,運動出力以前の思考や認知などの高次脳機能に 関わる脳活動を神経細胞レベルで捉えることを可能にした. 1.3. 大脳基底核の解剖と機能 大脳基底核は,大脳の内部深くに存在する神経核の集まりである.大きく分 けると,入力核である線条体(尾状核+被核)と,出力核である淡蒼球内節と 黒質網様部,中継核である淡蒼球外節,視床下核に分けることができる(図1.1A). その他に,黒質緻密部があり,ここにはドーパミンを神経伝達物質として放出 する細胞が多く存在する.入力部である線条体は,広範な大脳皮質領域と黒質 緻密部からの神経投射を受けており,認知から運動まで広範な機能に関わって いると言われている5, 6. 大脳基底核が障害されると重篤な運動障害を呈することが多い.たとえば, 黒質緻密部のドーパミン細胞の変性・脱落によって引き起こされるパーキンソ ン病では振戦・無動・固縮・姿勢反射障害といった4大症状といわれる運動機 能障害が,線条体の神経細胞の変性・脱落に起因するハンチントン病では舞踏
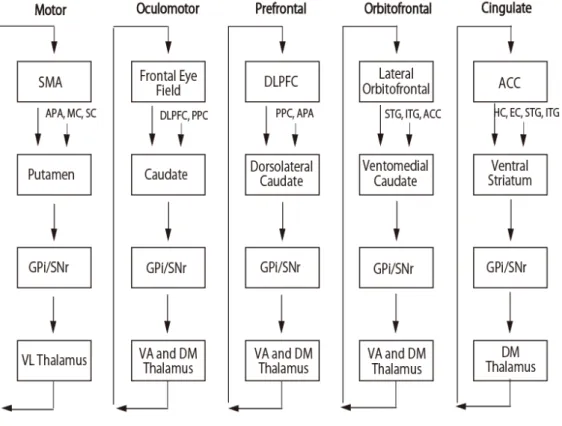
8 用運動と言われる不随意運動が生じることが知られている.このように大脳基 底核を中心とする疾患では運動の発現やその制御が障害されることから,その 機能は随意運動に関わっていると広く認識されている.しかし,実際は注意や 学習など高次脳機能障害も呈することが報告されている7, 8.解剖学的にも線条 体は,運動野のみならず前頭前野を含む広範な皮質領域から線維投射を受けて おり,線条体の小区分毎に比較的独立したいくつかのループ(皮質-基底核ルー プ)を形成していることが知られている(図1.2)9 10.さらに最近では,パーキ ンソン病の患者の意思決定能力は眼窩前頭皮質の体積に関係があることを示し た報告もある 11.こうした解剖学的な知見と臨床知見は,線条体は,皮質-基底 核ループを介して,運動のみならず,意思決定などの高次機能にも関与してい ることを意味している.
9
図 1.1 大脳基底核を構成する神経核群
GPe: external segment of globus pallidus, GPi: internal segment of globus pallidus, SNr: substantia nigra pars reticulata ,SNC: substantia nigra pars
10
図 1.2 大脳皮質—基底核ループ
SMA:supplementary motor area, GPi: internal segment of globus pallidus,
SNr: substantia nigra pars reticulata, VL: ventral lateral, VA: ventral
anterior ,DM: Dorsal medial, DLPFC: dordolateral prefrontal cortex, ACC:
anterior cingulate cortex, APA: arcuate premotor area, MC: pre-motor
cortex,SC: somatosensory cortex , PPC: posterior parietal cortex, STG:
superior temporal gyrus, ITG: inferior temporal gyrus, HC: hippocampal
11 1.4. 線条体と価値に基づく意思決定 神経科学的に,意思決定は,報酬あるいは価値に基づく行動選択として調べ られてきた.このような意思決定は,複数ある選択肢の中から,それらの価値 を比較して望しいものを選ぶという情報処理過程である.価値に基づく行動選 択の学習の枠組みとして,将来的な価値(得られる報酬量の総和)の見積もり が,試行錯誤によって徐々に正確になっていく過程を想定している強化学習モ デルが提案されている.各時間ステップで得られた実際の報酬が,そのとき見 積もられていた価値と比較してどれほど高かったか,あるいは逆にどれほど低 かったかという差分(実際に得られた報酬 – 見積もられた結果の価値)が報酬 予測誤差であり,強化学習モデルとは,報酬予測誤差が小さくなるように行動 選択を繰り返し行うことで将来得られる報酬量が最大となるような最適行動を 学習するモデルのことを言う.これまでサルを用いた行動学的・生理学的研究 では,黒質緻密部のドーパミン細胞が報酬予測誤差と同様の活動を示すこと12, その投射先である線条体が報酬期待を表象すること13,ドーパミンの拮抗薬を線 条体に注入すると報酬と刺激の連合の学習が障害されること 14などから大脳基 底核の解く問題は,ドーパミン信号を基にした強化学習として定式化できると 考えられている.すなわち,黒質緻密部のドーパミン細胞の報酬予測誤差信号 が線条体に伝えられることで,報酬を最大化するような行動が学習されるとい う仮説である.実際,線条体には,強化学習で用いられる行動価値パラメータ
12 に相関する細胞活動が多数存在していることが明らかにされている15, 16, 17. こ うした結果は,大脳基底核の機能が,価値に基づく意思決定に強く関与してい ることを示唆している. 1.5. 中央実行系とそれに関連する脳領域 認知心理学分野では,意思決定を含む認知システムを理解する概念として,
中央実行系(central executive system)が提案された18, 19.これは,中央実行系
は,状況に応じて必要な情報を更新するなどの認知制御を行うことで目的指向 行動を実現しているとされ,運動制御とは独立したその機能は,前頭前野に帰 せられることが示唆されている20, 21.さらに近年の神経科学分野においても, この考えを支持する概念モデルが提案されている22.こうしたモデルは,はじめ に運動とは独立した空間で意思決定に関わる様々な情報を統合することで対象 の価値を算出・選択をし,その後,その決定に基づいた行動を計画し実行する という意思決定過程を想定している.このモデルの神経基盤としては,前者の 意思決定には眼窩前頭皮質が,後者の行動の計画・実行には高次運動野・一次 運動野が関わっていると想定されている.しかし線条体については,運動の制 御と位置付けているだけである.実際にサルを被験体とした電気生理学的実験 によって,このモデルを支持する前頭前野の細胞活動が得られている23, 24.こ うしたモデルは,意思決定の中枢が前頭前野にあることを意味している. Samejima らは,前頭前野のみならず大脳基底核も含めた意思決定モデルを提唱
13 している25.このモデルでは,上述のように意思決定過程を一つのストリームに するのではなく,意思決定の対象が異なれば,異なる皮質—基底核ループが使わ れることを想定している.具体的には,運動要素を含まない価値に基づく意思 決定は前頭前野と吻側線条体ループが,実際の運動を伴う運動選択には高次運 動野/一次運動野-尾側線条体ループが関与していることを主張している.しかし ながら,大脳基底核においてこのモデルを確かめるような実験はこれまでに行 われていない. 1.6. 本論文の目的 本論文では,運動の制御とは独立した空間で行う選択と,実際に行う運動の空 間で行う選択に,複数の大脳皮質-基底核ループがどのように関わっているのか を調べることを最終目標として,サルを用いた実験(第2 章 Monkey study)と, ラットを用いた実験を行った26(第3 章 Rat study).サルを用いた実験では,運 動の制御とは独立した空間で行う対象の選択を,実際に行う運動の選択とは時 間的に分離した課題をサルに訓練し,課題遂行中の線条体から単一神経細胞活 動記録を行うことで,対象の選択に関わる線条体の情報表現を調べた.ラット を用いた実験では,意思決定の最終過程,すなわち運動実行を行う際に,一次 運動野とその出力先である背外側線条体のスパイク情報の性質を,頭部固定下 でレバー運動中のラットを用いてホールセル記録を行い,その時の膜電位変動
14
とスパイクとの関係を解析することで,課題遂行中における皮質と基底核のス
15
第
2 章
Monkey study:Neuronal activities in striatum
during object-decision-making
2.1. 導入
変動する環境の中で最適な行動を選択し実行することは,我々が生き残るための本 質的な脳機能である.心理学,経済学,神経科学分野では,行動選択を,「脳が,始 めに複数ある選択肢の価値を計算し,その後に最も望ましい価値を選択する」一連の 過程を指す概念として提唱している1, 2, 3, 4. 大脳基底核の線条体は,行動選択に関わる重要な脳領域として考えられている.解 剖学的に線条体は前頭前野や高次運動野,一次運動野などを含む大脳皮質領域と の間に皮質—基底核ループと呼ばれる神経連絡を有しており,連絡する皮質領域毎 に比較的独立したループを形成していることが知られている 5.これは,各々のループ によって認知から運動に至るまでの様々な情報が処理されていることを示唆している. 実際に,大脳基底核疾患であるパーキンソン病やハンチントン病,強迫性神経障害で は,行動の選択や運動制御の障害といった多様な症状を呈することから 6, 7, 8,線条体 が多岐にわたる情報処理をしている可能性を支持している.また近年では,皮質—線 条体結合は,線条体上での投射元の重複9, 10, 11が報酬や認知,運動の情報を統合す16 るために重要であると考えられ始めている 12.こうした解剖学的・臨床・神経生理学的 知見は,線条体が,皮質からの神経投射に基づいた機能的な神経回路を介して行動 選択の一連の過程に関わっている可能性を示唆している. 行動選択は,選択肢そのものに運動成分を含む選択と,含まない選択の2つの場合 が考えられる13, 14.前者は,実際に行う運動空間での選択であるのに対し,後者は,実 際に行う運動を選択する以前に決める抽象的な空間での選択であり,両者は段階的 な選択過程と考えられる.多くの神経科学的研究によって,外側頭頂葉 15,頭頂間溝 16,運動前野17,前頭前野18,線条体19で運動成分を含む選択に関わる信号が表象さ れていることが明らかにされている.とくに線条体は,強化学習の枠組みの中で運動 成分を含む選択に関わることが強く示唆されている 20, 21,22, 23.運動成分を含まない選 択に関しては,主に内側前頭前野がその選択肢や,実際に選択した選択肢の信号を 表象していることが明らかにされている24, 25, 26.大脳基底核・線条体においては,報酬 を予告する運動成分を含まない視覚刺激(=報酬期待)に応答する活動が見つけら れているが 27, 28,こうした研究で用いられた課題では,提示される視覚刺激が一つし か用いられていないため,実際に,線条体が運動成分を含まない選択にどのように関 わるのかは調べられていなかった. 線条体における大脳皮質の解剖学的な連絡 6, 7, 8, 9, 10, 11や,運動とは関係のない報 酬期待に相当する情報表現 27, 28 を考えると,線条体は,選択肢の対象が運動成分を
17 含む選択のみならず,内側前頭皮質を含む神経回路網の一部として,実際に行う運 動とは独立した運動要素を含まない選択においても重要な役割を担っている可能 性が考えられる.しかしながら,これまでに運動成分を含まない選択に線条体が関わ っていることを調べた神経生理学的な研究はない.その理由として,線条体における 行動学的・生理学的研究で使用されてきた行動課題が,運動要素を含まない選択を 調べるのに適していなかったことが挙げられる.たとえば,行動価値に相当する信号が 線条体に表象されていることを明らかにした Samejima らの研究13では,報酬を予測さ せる 2 つの選択肢は,直接ジョイスティックを倒す方向を指示する選択肢となっていた ため,選択肢そのものに運動要素が含まれていたことになる.そのためこの課題では, 運動要素を含む選択に関わる神経メカニズムを調べることにはできるが,運動要素を 含まない選択に関わる神経メカニズムを調べることには適していなかった.線条体が 運動要素を含まない選択に関わっていることを調べるためには,まず運動要素を含ま ない選択肢と運動要素を含む選択肢が時間的に分離されて提示される課題を使用す る必要があった. そこで本研究では,提示される2つの選択肢から1つを選択する任意選択課題を用 い,さらに運動要素を含まない対象の選択と,実際に行う運動の選択とを時間的 に分離した課題を考案した(図2.1A).課題遂行中のニホンザルの広範な線条体 領域から単一神経細胞活動の記録を行い,記録した細胞活動を用いて相互情報
18 量解析を行ったところ,行動の結果からだけでは見出すことのできない対象の 選択を行っている証拠,すなわち,選択する選択肢= Chosen stimulus,の情報表 現を線条体の細胞集団活動から見出すことができた(図 2.4).さらに,個々の 細胞に対して,対象の選択期間中の情報表現を解析したところ,Chosen stimulus を表現する細胞に加えて,提示された選択肢の情報= Offered stimulus を表現して いる細胞もいることがわかった(図 2.5).さらに,それら意思決定前後の情報 表現をもつ細胞は,線条体の中でも,吻側尾状核に偏って存在していることが 明らかになった(図 2.6, 図 2.7).これらの結果は,吻側尾状核が前頭前野を含 む神経サーキットの一端として,対象の選択,すなわち運動要素を含まない選 択に重要な役割を担っていることを示唆している.
2.2. 対象と方法
サルの飼育・管理・使用は,玉川大学動物実験倫理委員会(H21/27–14)の規定 に基づいて行われ,アメリカ国立衛生研究所のガイドラインおよび玉川大学の 規定に基づいて従って最小限の苦痛に収まるように努めた. 2.2.1. 被験体および実験装置 2頭のオスのニホンザルを被験体とした(monkey1:8.5kg,monkey2:8.0kg). 実験の最中は,サルをモンキーチェアーに座らせ,頭部を固定した.Monkey1 は左腕を,monkey2は両腕の動きを制限した.モンキーチェアーの右側前方の腰19
の高さには,課題を遂行するための押しボタンを設置した.サルの眼前には, 19
インチ液晶モニターを設置した.LCDモニターの背面に音刺激を提示するため
のスピーカーを設置した.眼位は, 赤外線カメラによる眼球追跡システム
(EYETRAC6000,Applied Science Laboratories)を使用し240 Hzのサンプリング
レートで測定した.サルの口元には、報酬としてジュースを与えるためのパイ
プを設置した.ジュースの量は,TEMPO systemからの制御信号を用いて電磁バ
ルブの開閉によって制御された.行動課題の制御は,TEMPO system(Reflective
Computing,Olympia)を用い,LED display による視覚刺激提示, 報酬量とタイ
ミング,報酬の予告刺激として音声刺激提示を行った.また,刺激順序の制御
はMATLAB(Math Works)によるCustom codeを用いた.
2.2.2. 行動課題
課題は,2頭のニホンザルに2つの選択肢のうちから1つを任意に選ぶ任意
選択課題(Free choice task)を用いた.課題は, サルをモンキーチェアーに乗せ,
サルの眼前に設置されたモニター上に提示される視覚刺激に従って手元のボタ
ンを押したり離したりすることで遂行された.課題遂行に成功すると,口元の
パイプから決められた量のジュースと,成功を知らせる報酬音が与えられた.
任意選択課題の1試行の流れを図2.1aに示す.サルがボタン押すと,まず注視
20 された.選択肢図形は,4種類の色(赤・青・黄・緑),4種類の形(○・△・ ◇・+)の組み合わせで構成される16種類の図形からランダムに選ばれた.Cue 刺激消失後0.8~1.2秒の遅延期間(一様分布によるjitter)の後に,2つの選択肢の うちいずれか1つが提示された(1st target).サルがこのままボタンを押し続け ると,1st target 消失に続いてもう1つの選択肢が提示された(2nd target).選 択は,各target提示中にボタンから手を離すことによって行われた.報酬の量は, 選択したtargetの属性(色または形のいずれか)に対して4段階に割り当てられ ていた.属性と報酬量の関係は144試行のブロック毎に変化した(図2.1C).選 択は必ず異なる色,異なる形の図形で行われた,すなわち,各試行において必 ず2つの異なる報酬量が提示された.この課題では,サルが図形と報酬の関係 を学習できていたとすると,選択肢図形の提示開始から1st target提示までの間, すなわち実際の運動(ボタンから手を離すか否か)を決める以前に,より多い 報酬を得るためにどちらの図形を選択するのかを決定できるところが特徴であ った.本課題にでは,運動の選択とは時間的に分離した時点で行う図形選択の ことを対象の選択(Object choice)と定義した.
対比課題は,選択を行わない強制選択課題(Forced choice task)が用いられた
(図2.1b).課題は,Cue刺激として1種類の図形のみが提示され,遅延期間の
21 異なる図形でボタンから手を離した場合は,報酬音および報酬は与えられず失 敗試行となった.課題中の眼球運動は固視点付近にあるように制限し,もし固 視点付近から大きく視点をずらした場合は失敗試行とした.任意選択課題,強 制選択課題の両方において,失敗試行の後は同じ刺激が再び提示されるコレク ションメソッドを用いた.各ブロックは,強制選択課題12試行・任意選択課題 24試行を1サブブロックとして、4サブブロック,計144試行で構成した(図2.1c). 2.2.3. 課題訓練と手術 課題学習が完了した時点で,外科的手術を行った.ケタミン,ペントバルビ タールの麻酔下で,頭部固定用装置と記録用チャンバーを装着した.手術から 回復後,頭部を固定して再び課題訓練を行った.このとき,サルに固視を行わ せるようにした.また,MRI装置で脳画像を撮影し,記録用チャンバーと脳部位 との関係が明らかにした. 2.2.4. 細胞活動の記録とデータ収集 課題成績が手術前の成績まで戻った後,電気生理学的手法(細胞外記録)を 用いて細胞活動の記録が開始した.細胞活動は,インピーダンス1.5MΩ(at 1
kHz)のガラス被覆のタングステン電極(Alpha Omega Engineering)によって記
録した.電極は,固定装置用マニピュレータ(SM-15, 成茂科学器械研究所),
22
ャンバー上にグリッドを設置し,グリッドを通して電極は挿入した.電極の挿
入は,トップスパイナル針23G,89mm/70mm(SN-2370/239,トップ)をガイド
チューブとして使用し硬膜を貫通させた後,ガイドチューブの中を通して挿入
した.細胞活動の検出とオンラインソーティングはMultichannel Acquisition
Processor ( MAP/16 , Plexon ) を 用 い て 行 っ た . 信 号 は ヘ ッ ド ス テ ー ジ
(HST/8o50-G20)とフィルター付きプレアンプ(PBX2/16wb-G50,Plexon)に
よって増幅とフィルタリングを行った(final gain, 500; band-pass filter 0.1–8 kHz).
行動課題の制御は,TEMPO system(Reflective Computing,Olympia)とMATLAB
(Math Works)によって行い,行動課題制御信号はTEMPOシステムからMAPシ
ステムによって神経信号との統合記録が行われた.活動電位のスパイクソート
は,Offline Sorter(ver3,Plexon)によってオフラインで行った.細胞活動,行
動データはともにMultichannel Acquisition Processorに記録した.ソートされた活
動電位データおよび行動データの解析はオフラインでMATABによって行った.
記録部位は,左半球の線条体から行った(AP: 21-30mm,L: 18-27mm (monkey1),
AP22-30mm,L18-28mm (monkey2) ).記録用チェンバーは,ステレオタクティ
ックフレームによって基準位置から前方の左脳へアプローチするように垂直方
向から側方へ35度傾けて設置した.記録用チェンバー設置後にも記録領域確認
23
目的の脳領域に電極を刺入後に,投射細胞と推定される(低い発火頻度)細胞
活動の記録を行った29, 30.
2.2.5. データ解析
課題関連細胞活動を調べるために,記録した個々の単一神経細胞の活動を
Choice cue提示時点,1st target提示時点,2nd target提示時点,1st release 時点,2nd
release時点に時間を揃えて,20ミリ秒のタイムビンを用いて最大発火率を示すイ
ベントのタイミングを調べた(図2.3A). 最大発火頻度が,Choice cue提示前0.8
秒(Pre cue期間),課題中のCue刺激の提示期間(0.8 秒)(Cue提示期間),Cue
刺激消失後1st targetが提示されるまでの遅延期間(0.8 秒)(Cue遅延時間),target 提示期間中(1st target提示期間),ボタンホールド期間中(2nd target提示期間),ボ タンをリリースする時点(Release運動期間),1sttarget後にボタンを保持し続ける 期間(Hold期間),報酬音・報酬が与えられた期間(Feedback期間),その他の試 行間時間(ITI)に応じて,記録した神経細胞を分類した(図2.3.b).また,細胞 活動様式を示すために50ミリ秒 のbinを用いて時間をCue提示時点に揃えた
pre-stimulus time histogram(PSTH)を作成した(図2.4A, 2.5A, 2.5A).
これ以降,サルが実際に対象の選択を行っているか否かを示すための細胞活
動の情報表現に焦点を当てるため,細胞活動の解析は学習による影響がない図
24
用いて行った.対象の選択に関わる神経細胞活動の情報表現を調べるために,
記録した個々の細胞活動に対して相互情報量解析を行った34, 35.①提示された2
つのCue刺激の図形の色の組合せ6種類,または形の組合せ6種類(= Offered
stimulus),②最終的に動物が選択した図形の色または形(= Chosen stimulus)の
課題条件を用いたアプリオリな情報量と,細胞の発火数の中央値を取り①—②の 条件それぞれで中央値よりも発火数が高かった試行と低かった試行を用いて計 算した情報量の平均との差分により相互情報量を求めた.以下の式で与えられ る. I (S; R) = H (S) – H (S|R) = −p (s) log p (s) − < −p (s|r) log p(s|r)>r ここでSは課題条件{s1, s2 …}のセット,Rは観測された神経活動{r1, r2…}のセッ ト,H (S) は課題条件Sについてのアプリリな情報エントロピー,H (S|R) は神経 活動R が観測されて条件づけられた課題条件Sについての情報エントロピー,<
>r は全神経活動平均を表している.また,Offered stimulusとChosen stimulusに
関しては色と形それぞれで相互情報量を計算した.
対象の選択期間中に,線条体全体で表現されている Chosen stimulus の相互情報
量が,Offered stimulus から期待される Chosen stimulus の相互情報量より,実際
s
∑
s
∑
25
に選ばれた stimulus の情報を有意に表現しているのかを調べた.同一の Offered
stimulus の組み合わせが提示された試行を集め,試行集団ごとに各試行での実際
に選択した stimulus をシャッフルすることで Chosen stimulus の情報をランダム
化したサロゲートデータを作成する.そのサロゲートデータから求められる Chosen stimulus の相互情報量は,活動電位の試行毎の自然なばらつきによって求 められる情報量となる.記録した全ての細胞に対してサロゲートデータ情報量 を計算・加算する手続きを 10,000 回繰り返すことで,サロゲート情報量分布を 作成した(図2.4B).サロゲート情報量分布の上位 5% を有意水準と定め,上位 5%に相当する情報量を超えていた場合に,その Chosen stimulus の加算相互情報 量は有意であるとした(すなわち p < 0.05).Choice cue 提示後から 1.6 秒間を 0.4 秒のタイムウィンドウ毎(0- 0.4 秒, 0.4- 0.8 秒, 0.8- 1.2 秒, 1.2- 1.6 秒)に4つの 区間に分け,情報量の有意差検定を行った.図2.4 に示してある加算相互情報量 は,全ての細胞に対して Choice cue 提示開始から 0.4 秒幅のウィンドウを 0.05 秒ステップでスライドさせて相互情報量解析を行い,個々のウィンドウでの相 互情報量を加算している. 個々の細胞に対しても Chosen stimulus の情報を有意に表現しているかを調 べた.この場合の有意性検定は,個々の細胞で情報をランダム化したサロゲー ト分布(シャッフル100 回分)を作成し,実際の相互情報量が上位 5%を超えて
26
いた場合に有意に情報を表現している細胞(Chosen stimulus type neuron)とした
(図2.5A).また,提示された図形の組み合わせの情報(Offered stimulus)につ
いても有意性検定を行った.同じstimulus を選択した試行集団の中で,提示され
たstimulus の組み合わせをシャッフルして Offered stimulus の情報をランダム化
したサロゲート情報量分布を作成し,実際に計算された相互情報量の値がその
分布の上位5%を超えていた場合に有意とした(Chosen stimulus type neuron:図
2.5B).
2.3. 結果
2.3.1. 行動選択および反応時間 本研究で用いた課題は,実際に行う運動選択とは独立した時点で,対象の選 択を行うことができるという特徴を持った課題であった(図2.1A,2.2.2 行動課 題参照).属性(色または形のいずれか)と報酬量の関係は試行のブロック毎(144 試行)に変化するため,サルはブロック毎に属性と報酬量の関係を学ぶ必要が あった(図2.1C).図 2.2A-C に monkey1 の 1 日の行動結果の例を示す.サルが, 属性と報酬量の関係を学習でき,より大きい報酬量が割り当てられている図形 を正しく選択することができているのかを調べるために,1 ブロック内の任意選 択課題で,正しく選択を行った割合の推移と,1 ブロック前の正しい選択行動の 割合の推移を求めた(図2.2A).ブロックが切り替ってから2サブブロックが経27
過した時点で,色もしくは形の属性のどちらに対しても正しい選択行動が上昇
(54.2%(1 試行)→86.9%(96 試行)color, 55.5%(1 試行)→87.9%(96 試行)shape)
し,反対に前ブロックで正しかった選択行動の割合は減少した(78.8%→54.9%
color , 78.8%→54.89% shape).また, target が提示されてからボタンを離すまで
の時間(反応時間)は,target が予告する報酬量が大きいほど早く,それはブロ ックの切り替わりからサブブロックの進行に伴ってより明確になっていた(図 2.2B).さらに,1st と 2nd target の予告する報酬量の組合せに対する 1st target の 選択率を求めたところ,どの報酬量の組合せにおいても1st target の予告する報 酬量が高いときは1st target の選択率が高くたっており,それは,ブロックの切 り替わり後からサブブロックの進行に伴ってより明確になっていた(図 2.2C). この結果は,monkey1 は属性と報酬量の関係を毎ブロックごとに学習しており, どの報酬量の組合せが提示されたとしても正しくより大きい報酬量が割り当て られている図形を選択していることを示している.そこで,2 頭のサルがより大 きい報酬量が割り当てられている図形を正しく選択できているのかを定量的に 解析するために,実際に細胞活動を記録していたセッションのデータ(monkey1: 201 日,monkey2: 174 日)を使用して,提示された報酬量の差分(1st – 2ndtarget) に対する1st target 選択率(図 2.2D と F)と平均反応時間(図 2.2E と G)を調べ た.また,この際に使用したデータは,属性と報酬量の関係の学習が完了して
28
いるブロックの後半試行(3-4 サブブロック)を用いた.2頭のサルともに,1st
target 選択率に対しては,シグモイド関数でよくフィッティングでき(monkey1:
R2 = 0.824, monkey2: R2 = 0.802),反応時間に対しては報酬量の差分による効
果が有意であった(one-way ANOVA, monkey1: F = 9.99, p = 0.000054,monkey2:
9.16,p = 0.0001).これらの選択行動は2頭のサルで有意な差はなかった(図
2.2D :one-way ANOVA, F = 2.17, p = 0.52 ).こうした行動の結果は2頭のサル
ともに属性と報酬量の関係を毎ブロックごとに学習し,より大きな報酬を短期
29
図 2.1 認知選択課題と報酬スケジュール
A. 認知選択課題.B. 強制選択課題.C 属性(色または形)と報酬量 4 段
31 図 2.2 行動選択と反応時間 A. 任意選択課題における,ブロックの切り替わり後の正しい図形の選択率の推 移(1日分のデータ).シアンと赤はそれぞれ現在の色または形ブロックでの, 緑と赤は1つ前の色または形ブロックにおける正しい図形の選択率を表してい る.B. サブブロック(1st, 2nd, 3rd, 4th)の進行に伴う平均反応時間の推移(1 日分のデータ).それぞれ青,緑,赤,シアンは,報酬量1, 2, 3, 4 を選んだ時の 反応時間を示している.C.提示された2つの図形に対して連合している報酬量 の組み合わせに対する1st target の選択率(1 日分のデータ).横軸・縦軸にそれ ぞれ1st もしくは 2nd target に対して割り当てられていた報酬量4段階を示し, 寒暖色によって1st target 選択率を示した.D. monkey1 における,提示された報
酬量の差分(1st – 2nd target)に対して 1st target の選択率を示した(n = 201 session).
赤線はシグモイド関数でフィッティングをした結果を示している.E.monkey1
における,提示された報酬量の差分(1st – 2nd target)に対しての 1st target 提示
時の平均反応時間(n = 201 session). Rank sum test with Bonfeloni collection によ
って統計は行った.F.D と同様で,monkey2, n = 174 session のデータを使用.G.
E と同様で monkey2, n = 174 session のデータを使用.それぞれのバーは s.e.m を
32
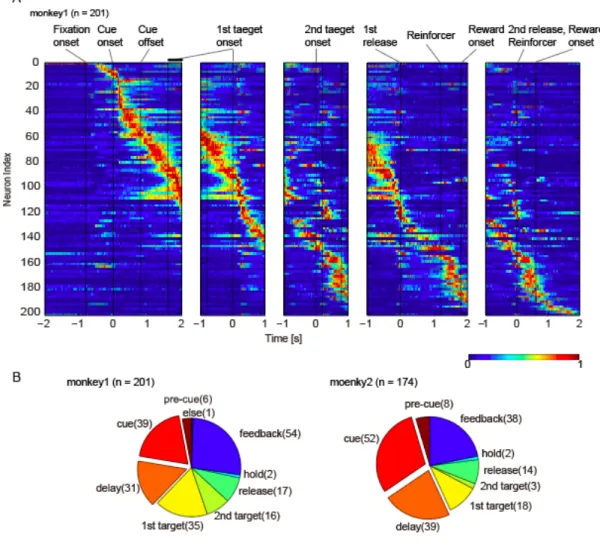
2.3.2. 線条体の課題関連細胞活動のデータベース
課題遂行中の 2 頭のサルの線条体から 375 個の細胞活動を記録した.課題中
の最大発火頻度を示す期間によって(図2.3.A,2.2.5. データ解析参照), Pre-cue
関連細胞,Cue 関連細胞,Cue 遅延時間関連細胞,1st target 関連細胞,2nd target
関連細胞, Release 関連細胞,Hold ド関連細胞,Feedback 関連細胞とした.これ
らの課題関連細胞活動のうち,monkey1(n = 201)では Pre cue 関連細胞が 2%
(n = 6),Cue 関連細胞が 19.4% (n = 39),Cue 遅延関連細胞が 15.4% (n = 31),1st
と2nd target 関連細胞が合計 25.4% (n = 51),Release 関連細胞が 8.5% (n = 17),
Hold 関連細胞が 1%(n = 2),Feedback 関連細胞が 26.9% (n = 54)であり(図 2.2B
左),monkey2(n = 174)では Pre cue 関連細胞が 4.6%(n = 8),Cue 関連細胞が
29.9% (n = 52),Cue 遅延関連細胞が 22.4% (n = 39),1st と 2nd target 関連細胞の
合計が12% (n = 21),Release 関連細胞が 8.1% (n = 14),Hold 関連細胞が 1.2%(n
= 2),Feedback 関連細胞が 21.8% (n = 38)であることがわかった(図 2.2B 右).
これらの記録された細胞活動のうち, Choice cue 提示開始から 1st target が提示
されるまでの期間,すなわち対象の選択期間に最大応答を示す細胞(Cue、Cue
遅延関連細胞)の割合は,monkey KD で 34.8%,monkey UT で 45.3%であった(図
33
図 2.3 線条体の課題関連細胞活動
A. monkey1 から記録した 201 個の細胞を使用して,課題イベント(Choice cue
onset,1st target onset,2nd target onset, 1st release 2nd release)に対して時間を揃え,
各時間帯の発火頻度を最大発火頻度で標準化し,寒暖色によって0-1 を示してい
る.B. 最大発火頻度のある時間帯に応じて分類した課題関連細胞活動の割合を
34
2.3.3 対象の選択をしていることを示す集団細胞活動
本課題の特徴は,Choice cue 提示開始から 1st target 提示までの間により多い報
酬を得るためにどちらの図形を選択するのか(対象の選択)を決定できるとこ ろであるが,サルは必ずしもこの期間に対象の選択を行う必要はない.たとえ ば,サルは, 1st target が提示された時点で実際に行う運動とともに図形の選択 を行うことで本課題を遂行することができる.そこで,実際にサルが対象の選 択を行っているのか否かを調べるために,記録した全ての細胞に対して Choice cue 提示開始から 1st target 提示までの間の細胞活動に対象の選択後の情報,すな わち最終的にボタンをリリースすることによって選択する図形(= Chosen stimulus)の情報が有意に表現されているか否かを調べた(2.2.5 データ解析参照).
実際に記録した全ての細胞に対して,Choice cue 提示開始から 1st target が提示
されるまでの1.6 秒間を 4 つのウィンドウに区切り,それぞれのウィンドウで相
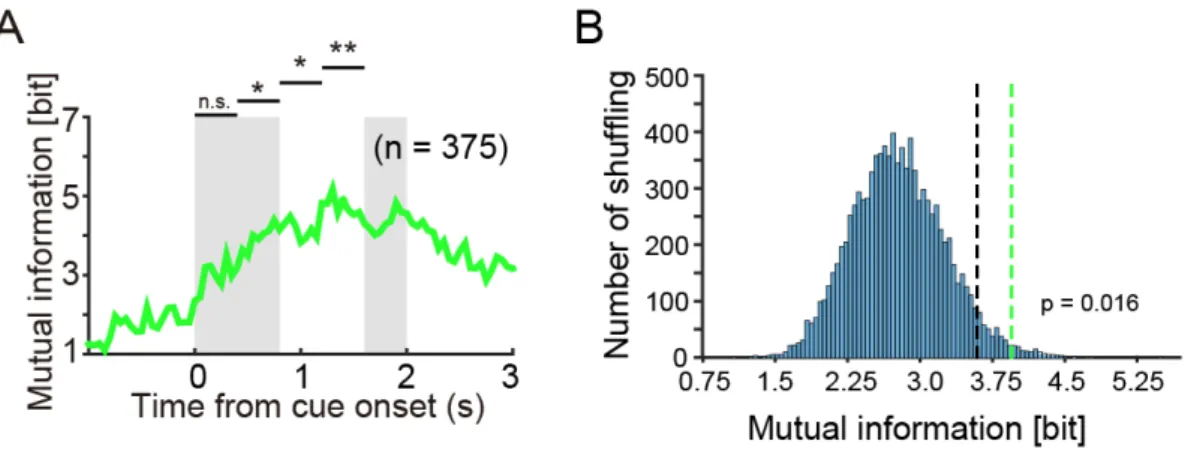
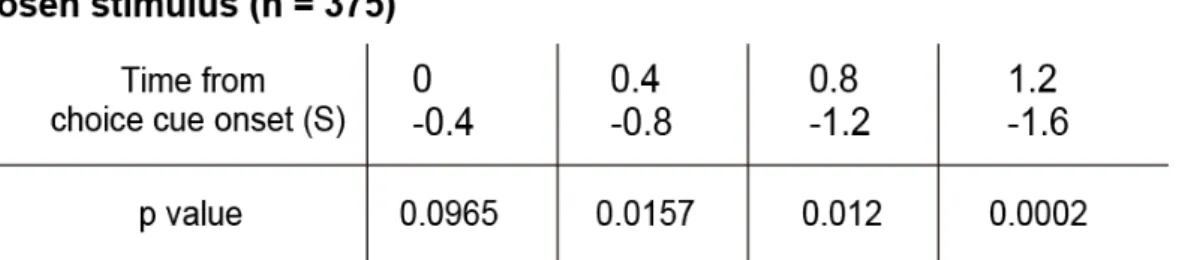
互情報量を計算・加算すると,choice cue 提示開始後 0.4 秒以降の 3 つのウィン
ドウでChosen stimulus の相互情報量が Chosen stimulus の情報をランダム化した
サロゲート分布の上位 5%に相当する情報量よりも大きいことがわかった(図
2.4A, B, 表 1).すなわち,線条体の集団細胞活動は,Choice cue 提示開始から
1st target が提示されるまでの間に,サルが最終的に運動によって選択する図形
の情報を有意に表現しており,これはサルが実際にこの期間に対象の選択を行
35
図 2.4 対象の選択をしていることを示す集団細胞活動
A. 線条体の細胞群が表現する選択した図形(Chosen stimulus)の相互情報量.
時間はChoice cue 提示開始に揃えている.灰色の影は,左側が Choice cue 提示
期間中,右側が1st target の提示開始時点を示している.図上の 4 本のバーは,
Choice cue 提示開始から 1st target が提示されるまでの期間を 4 つの期間に分け,
それぞれの期間における相互情報量の有意性を示している.B. Chosen stimulus
の情報をランダム化したサロゲートデータ(シャッフル 10,000 回分)によって
作られた相互情報量分布.図中の黒の点線は,サロゲート分布の 95%に相当す
る情報量(3.6bit),緑の点線は実際の Chosen stimulus の相互情報量(4.16bit)を
36
表 1 線条体の細胞群における Chosen stimulus の情報表現の有意性検定.
Choice cue 提示開始から 1st target 開始時点までを 0.4 秒ずつの 4 つのウィンドウ
に分け,それぞれのウィンドウ内における実際の情報量と,サロゲート情報量
37
2.3.4 対象の選択時期における各細胞の情報表現
サルが対象の選択を行っていることがわかったので,つづいて,個々の細胞
に対して,対象の選択が行われている期間(Choice cue 提示開始から 1st target
提示開始までの間)の情報表現を調べた(図 2.5).ここでは,提示された選択
肢図形の組み合わせ(offered stimulus)の情報と最終的にサルが運動によって選
択した図形(Chosen stimulus)の情報,すなわち対象の選択前後の情報表現につ
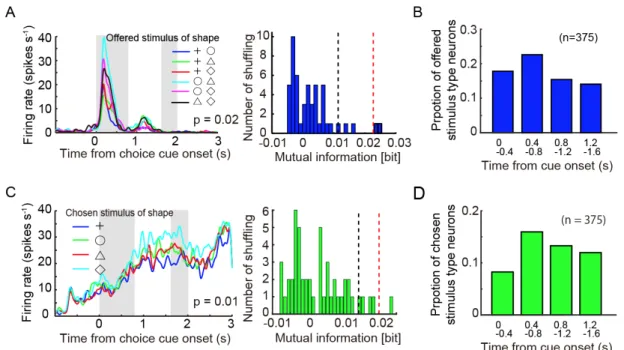
いて調べた.図2.4A 左は,Choice cue 提示開始時点に揃えたある一つの細胞の
発火頻度をOffered stimulus でソートした例で,Choice cue 提示期間中に Offered
stimulus を有意に表現していた(図 2.4A 右,p = 0.02,2.2.5 データ解析参照).
ここでは,この細胞をOffered stimulus type と定義した. 0.4 秒タイムウィンド
ウをChoice cue 提示後から4つ並べて(0- 0.4 秒, 0.4- 0.8 秒, 0.8- 1.2 秒, 1.2- 1.6
秒),記録した375 個の細胞活動に対して Offered stimulus type neuron の割合を調
べたところ(P < 0.05),全ての期間において Offered stimulus type neuron が有意
に存在していることが明らかになった(図2.5B, p < 0.05).図 2.5A は,Choice cue
提示開始時点に揃えたある一つの細胞の発火頻度をChosen stimulus でソートし
た例である.この細胞活動は, Choice cue delay 期間中に Chosen stimulus の情報
を有意に表現していた(図2.5 C,p = 0.01: Chosen stimulus type neuron).Chosen
stimulus type neurons(P < 0.05)の割合は,全ての期間において有意に存在して
38
選択期間中に,選択前後の情報を表現するOffered stimulus type neurons と Chosen
39
図 2.5 対象の選択時期における各細胞の情報表現
A. 提示された図形の組み合わせの情報(offered stimulus)を表現する細胞例.
時間をChoice cue onset に揃え,提示された図形の形の組み合わせによってソー
トした発火頻度.それぞれの色は,提示された形の組み合わせを示している
(青:プラス v.s. 丸,緑:プラス v.s. 三角,赤:プラス v.s. ダイヤ,シアン:
丸 v.s.三角,紫:丸 v.s. ダイヤ,黒:三角 v.s. ダイヤ).灰色の影は,左
側がChoice cue 提示期間中,右側が 1st target の提示開始時点を示している.B.
A の細胞を使用し,Choice cue 提示開始から 0- 0.4 秒のウィンドウで,offered
stimulus をランダム化して計算したサロゲートデータ相互情報量の分布(n shuffle
=100)と実際の offered stimulus の相互情報量を示している.黒の点線は,この
40
相互情報量を示している. C. 選択した図形の情報(chosen stimulus)を表現す
る細胞例.時間をChoice cue onset に揃え,サルがその試行で最終的に選択をし
た図形の形によってソートした発火頻度.それぞれの色は,選択した形を示し
ている(青:プラス,緑:丸,赤:三角,シアン:ダイヤ).灰色の影は,左側
がChoice cue 提示期間中,右側が 1st target の提示開始時点を示している.D. C
の細胞を使用し,Choice cue 提示開始から 1.2- 1.6 秒のウィンドウで,chosen
stimulus をランダム化して計算したサロゲートデータ相互情報量の分布(n shuffle
=100)と実際の chosen stimulus の相互情報量を示している.黒の点線は,この
分布の上位5%相当の相互情報量を,赤の点線は実際に A の細胞が表現している
41
2.3.5 対象の選択時期における Offered stimulus と Chosen stimulus の情報表
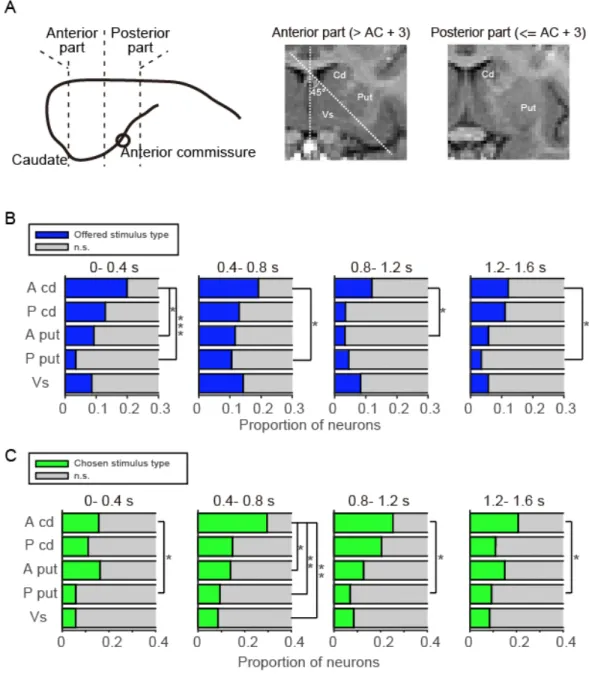
現の線条体内分布 線条体は皮質からの神経投射に基づいた複数の機能的なサーキットを介して行動 選択の一連の過程に関わっている可能性がある 5, 12.そこで,線条体内で Offered stimulus,Chosen stimulus の情報表現に偏りがあるのかを調べることで,線条体 内で対象の選択に関わる領域,ないしはサーキットの一端の特定を試みた.ま ず,記録した線条体を 5 つのサブ領域に分けた 31, 32(図 2.6A);吻側尾状核
(Anterior part of caudate: A cd),吻側被核(Anterior part of putamen: A put),腹側
線条体(Ventral striatum: vs),尾側尾状核(Posterior part of caudate: P cd),尾側
被核(Posterior part of putamen: P put).つづいて,Choice cue 提示後から4つ並
べたウィンドウを用いて(0- 0.4 秒, 0.4- 0.8 秒, 0.8- 1.2 秒, 1.2- 1.6 秒),各領域に
おいてOffered stimulus type または Chosen stimulus type に分類された細胞の割
合を調べた(図2.6B と C, chi-square test with Benjamini-Hochberg Procedure (q =
0.05)).その結果,吻側尾状核の Offered stimulus type の細胞の割合が,Choice cue
開始直後に他の領域(吻・尾側被核)と比べると有意に多くなっていた(図 2.
8A).Chosen stimulus type の細胞においても,吻側尾状核での割合が Choice cue
提示から 0.8 秒後の時点で他の領域に比べ最も多くなっていることがわかった
(図2.6C).そこで,吻側尾状核に着目をして,Offered stimulus,Chosen stimulus
42
Offered stimulus,Chosen stimulus のどちら情報表現においても,Choice cue 提示
後0.4 秒後以降に有意であった(図 2.7A, B, 表 2).これらの結果は,対象の選
択の期間中に,Offered stimulus,Chosen stimulus の情報表現が吻側尾状核に偏っ
た情報が表現されていることを示唆している.
つづいて,線条体で表現されているOffered stimulus と Chosen stimulus の情報
は,別々の細胞で表現されているのか,または,同一細胞においてその表現が
遷移するのかを調べた. Choice cue 提示後から4つ並べたウィンドウを用いて
(0- 0.4 秒, 0.4- 0.8 秒, 0.8- 1.2 秒, 1.2- 1.6 秒)同定された Offered stimulus type の
細胞に対して,同定されたウィンドウ以降で Offered stimulus type→Chosen
stimulus type へと表現する情報が遷移した細胞の割合を各5つのサブ領域で解
析したところ,5つのサブ領域全てにおいて,Offered stimulus の情報と Chosen
stimulus の情報表現は,異なる細胞によってこれらの情報が表現されていること
がわかった(図2.8, chi-square test, chi2= 3.3356, p = 0.6786).最後に,線条体のサ
ブ領域における相互情報量の表現の違いが領域間における発火頻度の違いに依
存している可能性を調べるために,Choice cue 提示後から4つ並べたウィンド
ウを用いて(0- 0.4 秒, 0.4- 0.8 秒, 0.8- 1.2 秒, 1.2- 1.6 秒),各領域間の発火頻度を
比べた.その結果,2頭のサルで全ての期間において,全ての領域間での発火
43
報表現(Offered stimulus と Chosen stimulus)は,線条体の中においても吻側尾状
核に偏って表現されており,さらにそれらの情報は異なる細胞によって別々に
44
図 2.6 Offered stimulus と chosen stimulus の情報表現の線条体内分布
A. 線条体の各領域.左は側面からみた尾状核を意味したイラスト.前交連
(Anterior commissure: AC)を基準に,AC から 3mm より前方を吻側部,後方を
尾側部とした.右は,monkey1 の MRI 画像前額断面. 尾状核,被殻,腹側線条
体を吻側部,尾側部に分けて示した.尾状核と被殻は内包によって分け,吻側
45
領域における,Choice cue 提示開始から 4 つの期間(0-0.4 秒, 0.4-0.8 秒, 0.8-1.2
秒, 1.2-1.6 秒)で検出された offered stimulus type neuron の割合.統計は chi-square
test with Benjamini-Hochberg Procedure (q = 0.05) を用いて行なった.C. B と同様
の条件で,Chosen stimulus type neuron の割合を示した.D. 吻側尾状核の細胞群
を使用して計算したOffered stimulus と Chosen stimulus の加算相互情報量.青,
緑がそれぞれOffered stimulus,Chosen stimulus の加算相互情報量を示している.
灰色の影は,左側がChoice cue 提示期間中,右側が 1st target の提示開始時点を
示している.*: 0.05 < P, **:0.01 < P, ***:0.001 < P.A cd: 吻側尾状核,P cd: 尾側
46
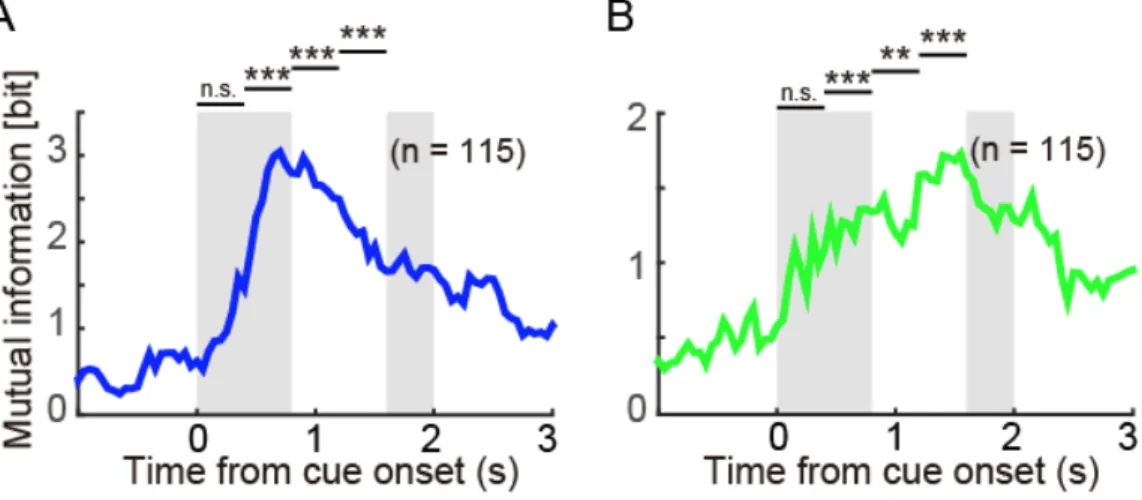
図 2.7 吻側尾状核における Offered stimulus と chosen stimulus の情報表現
A.吻側尾状核の細胞群が表現する提示された選択肢(Offered stimulus)の相互
情報量.時間はChoice cue 提示開始に揃えている.灰色の影は,左側が Choice cue
提示期間中,右側が1st target の提示開始時点を示している.図上の 4 本のバー
は,Choice cue 提示開始から 1st target が提示されるまでの期間を 4 つの期間に分
け,それぞれの期間における相互情報量の有意性を示している.B. 吻側尾状核
の細胞群が表現する選択した図形(Chosen stimulus)の相互情報量. *: 0.05 < P,
47
表2 吻側尾状核の細胞群におけるOffered stimulus と Chosen stimulus の情
報表現の有意性検定.
Choice cue 提示開始から 1st target 開始時点までを 0.4 秒ずつの 4 つのウィンド
ウに分け,それぞれのウィンドウ内における実際の情報量と,サロゲート情報
48
図 2.8 同一細胞における Offered stimulus type neurons と Chosen stimulus type neurons の情報表現
線条体内の5つのサブ領域において,offered stimulus type の細胞から Chosen
stimulus type へ遷移した細胞の割合.Choice cue 提示開始から 4 つの期間(0-0.4
秒, 0.4-0.8 秒, 0.8-1.2 秒, 1.2-1.6 秒)を用いて Offered stimulus type の細胞を検出
し,いずれかの期間で Chosen stimulus type の細胞に遷移があったものの割合を
49
2.4. 考察
運動の要素を含まない対象の選択に関わる線条体の機能を調べるために,ボ タンを離す運動を行うか否かの運動の選択とは明示的に分離した時点で,異な る報酬量が割り当てられた2つのChoice cue 刺激からより多い報酬を得るため の図形の選択,すなわち,対象の選択を行う課題を考案し,2 頭のサルに訓練を した(図2.1,図 2.2).課題遂行中の線条体から単一神経細胞活動を記録した結 果,2 頭のサルで合計 375 個の細胞活動を記録することができ, 161 個(42.9%) の細胞活動が対象の選択期間に最大の発火頻度を示していた(図2.3).375 個の 細胞群に対し,細胞活動と課題条件(サルが最終的に選択した図形:Chosen stimulus)から相互情報量解析を行ったところ,線条体の細胞群は,対象の選択 期間にChosen stimulus の情報を有意に表現していることがわかった(図 2.4, 表 1).個々の細胞に対しての情報表現を調べたところ,Choice cue として提示された図形の組み合わせ(Offered stimulus),Chosen stimulus の課題条件を用いた相
互情報量解析によってOffered stimulus type,Chosen stimulus type の細胞が見つ
かった(図2.5A-D).さらに,線条体を5つの領域(吻側尾状核: A cd, 吻側被
核: A put,腹側線条体: Vs,尾側尾状核: P cd,尾側被核: P put))に分けて,対象
の選択期間中のOffered stimulus type と Chosen stimulus type の細胞の分布を調べ
50
に,その割合が他の領域と比べて多く存在していた(図 2.6).それらの細胞集
団としての情報表現は,線条体全領域の情報表現と同様に,Choice cue 提示後に
Offered stimulus,Chosen stimulus の情報がともに有意に表現されていた(図 2.7,
表2).対象の選択期間中における Offered stimulus と Chosen stimulus の情報は,
それぞれ別々の細胞によって表現されていた(図2.8). 本研究で用いた行動課題は,実際に行う運動の選択とは時間的に独立した時 点で図形の選択を行うことができる独創的な課題である.さらに,Choice cue と して提示される 2 つの図形は,運動の方向(e.g. 眼球運動方向やリーチング方 向)を示さない上に,2×2 の形式(左右上下にそれぞれが提示)で提示される ため,空間情報も 2 つの Choice cue で等しくなっている.これまでは,運動選 択の要素が含まれた課題や運動の方向を示す課題を用いることで,線条体が行 動選択に関わっていることが主張されてきた19, 21, 22, 33.いくつかの研究では,線 条体が実際に行う運動とは関係のない報酬情報をコードしていることを報告し ているが27, 28,これらの研究では,2つ以上の選択肢の価値を比較して望ましい 1つを選ぶという任意選択の要素が含まれていなかったため,実際に線条体が 運動要素を含まない選択行動に関わっているのかは明らかにできていない.そ のため本行動課題は,運動要素を含まない行動選択の時間帯を巧みに抽出した
51 stimulus の情報が線条体で有意に表現されていた結果は,線条体が運動要素とは 関係のない行動の選択に関わっていることを十分に示唆している. 線条体は,解剖学的に比較的独立しているいくつかの皮質—基底核ループによ って広範な皮質領域と神経連絡を持ち,それぞれのループは,運動ループ,前 頭前野ループ,辺縁系ループ,眼球運動ループといわれている5.それぞれのル ープは異なる機能を持ち,運動ループでは行動価値に基づく運動選択が行われ, 前頭前野ループでは運動要素とは関係のない価値に基づいた対象の選択が行わ れることを示したモデルが提唱されている34.また,皮質から線条体への投射は, 基本的にはトポロジーであるとされているが,線条体上で異なる皮質領域から 投射を受ける領域もあることが知られており 9,10,11,この投射の重複が皮質から の情報を統合するのに重要であるという考え方もある 12.本研究における
Offered stimulus,Chosen stimulus の情報が強く表現されていた吻側尾状核は,背
外側前頭前野(DLPFC)や,眼窩前頭皮質(OFC),前帯状運動皮質(ACC)か らの投射を受けており5, 10, 10, 12,前頭前野ループに位置付けることができる.多 くのサルの電気生理実験やヒトのイメージング研究では,前頭前野,とくに眼 窩前頭皮質は運動要素とは関係のない価値に基づく行動選択に関わっていると 示唆されている24, 25, 26.また,前頭前野における色・形に反応する非空間的な視 覚情報は,吻側の尾状核へ入力されていることを示唆する研究もある35, 36, 37, 38.
52 一方で,運動要素を含む選択においては,皮質領域では,高次運動野27, 39, 40, 41 が 重要であることを示唆する研究があり,こうした知見は運動要素を含む選択に は,高次運動野を含むループの関与が考えられる.吻側線条体と前頭前野との 解剖学的な結合と前頭前野における情報表現を考慮すると,吻側尾状核は,前 頭前野ループ内における眼窩前頭皮質や背外側前頭前野,前帯状皮質 27などの 領域と運動要素とは関係のない価値や刺激の属性の情報をやり取りすることで 対象の選択に関わっている可能性がある. 吻側尾状核において,これまでに運動の要素とは関係のない報酬期待や,価 値(Flexible value)に相当する細胞活動があることが知られている27, 28, 42.また, 運動要素は含んでいるが,線条体の細胞が価値を表現している報告もある21, 22. しかしながら本研究では,二種類の属性に報酬を割り当てており,それぞれ色 と形で少なくとも一種類の報酬量パターンの連合における神経活動のみしか観 測できなかった.すなわち,細胞活動が提示された図形の色と形と連合した価 値情報を表現しているのか,色や形そのものであるのかを区別することはでき なかった.本研究における運動要素がない条件での対象の価値情報が線条体に おいて,選択の前後でどのように表現され,選択に直接的に関わるのかは今後 の問題のひとつである. 本研究における吻側尾状核でのOffered stimulus の情報表現は,同一細胞上で
53
その情報を遷移させる(Chosen stimulus type に変わる)ことがなかった(図 2.8).
これは,Offered stimulus の情報が同一細胞上で遷移するのではなく,吻側尾状 核内の異なる細胞に変換された情報が送られていることを示唆している.この 結果の可能性の一つとしては,単純に皮質において 2 つの情報表現が存在し, その領域からの異なる情報が異なる細胞に投射されているだけかもしれない. もう一つの可能性は,Offered stimulus の情報がループを介して異なる細胞に少 しずつ変換されながら受け渡されているという可能性である.前頭前野ループ 内にとどまらず,皮質—基底核ループ間で情報の遷移がどのように起きているか を調べることは大きな課題の一つである. 本研究結果は、前頭前野—皮質基底核ループにおいて運動の要素を含まない対 象の選択が起きているという仮説47を支持しており,今後,皮質—基底核ループ において意思決定過程を調べる神経科学研究(Decision-neuroscience)において 吻側尾状核の位置づけを示す重要な研究であると言える.
54
第
3 章
Rat study : Continuous membrane potential
fluctuations in motor cortex and striatum
neurons during voluntary forelimb movements
and pauses
3.1. 導入
霊長類は,一次運動野の神経活動の制御によって頭や体,腕などの分離運動を 自発的に巧みに行っている4, 7.げっ歯類も同様に四肢の分離運動を行うことが でき,それは皮質下のパターンジェネレータが駆動するような,前肢における 自動反復運動(ロコモーションやウィスキング)などとは区別されている17, 27, 6. 近年では,頭部固定下で課題遂行中のラットの一次運動野(M1)の細胞が分離運動を行っている間や(e.g. push/pulling a lever ; Movement-type neurons ),止ま
っている間(e.g. keeping a lever in the hold position; Hold-type neurons)に活動を上
昇させることが明らかにされている10, 11, 24.Movement type neuron は浅層と深層
に,Hold type neuron はそのほとんどが深層に分布している 9.Movement type
neurons と Hold type neurons の細胞活動は,M1 からの神経投射を受ける背外側
55 シータ,ベータ,ガンマ帯域を含んだ局所フィールド電位(LFP)や皮質脳波 (ECoG)の同期が自発的な四肢の運動に依存して生じることも明らかにされて いる9, 1, 29.基本的に,アクティブな運動は運動野におけるベータや他の遅い揺 らぎを減少させ,ガンマ帯域の揺らぎを大きくする.運動情報は実際に、ガン マ帯域の活動からデコーディングをすることができる14 (>30 Hz).運動野の細 胞活動は,行動中に,位相にロックしたガンマやシータの揺らぎがあり,それ らは運動野内の細胞間で同期している16. では,それらの細胞はどのようにして異なる行動をしている状況に依存した 機能的な細胞活動を引き起こしているのであろうか.これまで,in vivo におけ る細胞膜電位の解析は,機能的な情報を適切に運ぶために個々の細胞で細胞活 動が発生することによる細胞内メカニズムを解明することをゴールにして研究 されてきている.重要なこととして,行動中の皮質細胞の膜電位の揺らぎは, 細胞活動を発生させる閾値以下で生じ20, 3,そしてそれは行動の状況に依存して ECoG の活動と同期や非同期をしていたことである.理論的なシミュレーション において,膜電位の安定的な変化(脱分極/過分極)に伴った膜電位の揺らぎの ゲイン変化は,シナプス入力に対する細胞の反応性を最適化する可能性がある ことを示している 8, 2, 21. げっ歯類では,体性感覚の皮質細胞が自発的なヒゲの 運動をしている間に,ヒゲ運動をせずに静かにしているときと比べて,膜電位
56 を脱分極させ、その揺らぎを減弱させていることが知られている23, 30.同様に, 運動野の細胞活動も静かに座っているときと比べると,走ったり,歩いたりし ているときに膜電位の揺らぎを減らしている25.また,いくつかの運動野細胞は リッキング反応に先立ってその揺らぎと細胞活動を減少させていることも報告 されている31.つまり,皮質細胞は,ロコモーションやリッキング,ウィスキン グのような反復運動の間は,安定的な脱分極または過分極を伴って膜電位の揺 らぎを減少させていることがわかる.線条体の細胞も,リッキング運動に伴っ て膜電位を減少させていることも知られている26.それゆえ,個々の細胞におけ る運動情報は膜電位の揺らぎのパワーによって直接コードされている可能性が ある.しかしながら,行動実験や電気生理実験の技術的な制約により,運動野 と線条体における膜電位の揺らぎが,運動を行なっていないときと比較して, 前肢の分離運動を行なっている間に機能的な変化が見られるか否かは知られて いない. この可能性を確かめるために,ここでは,分離運動をしている間と静止して いる間の運動野と線条体の細胞の膜電位の揺らぎを調べるために in vivo
whole-cell recording を行なった(図 3.1A; それぞれレバープル とレバーホール
57
3.2. 対象と方法
Long-Evans ラット(N = 32, 150–243 g, male; Institute for Animal Reproduction,
Japan)に対し,2.0-2.5%のイソフルランの麻酔下で,頭部固定具をアンカーネ
ジとデンタルレジンセメント(uper-Bond C & B, Sun Medical,Japan; Panavia F2.0,
Kuraray Medical, Japan; Unifast II, GC Corporation, Japan)を用いて頭蓋に取り付け
た(Univentor 400 anesthesia unit, Univentor, Malta).手術中は,体温は 37.0 ℃に
保 持 し た (BWT-100, Bio Research Center, Japan) . 2 本 の 銀 線 ワ イ ヤ ー
(Teflon-coated, 180 µm in diameter, A-Msystems, USA)を,小脳上にリファレン
ス電極とグランドしてそれぞれ設置した.手術から回復後,ラットに給水制限
を開始した.ただし課題をこなすことによって十分な量の水を報酬として与え
た.必要に応じてアガーブロック(水15ml 含有)を与え,給水制限をかける前
の体重から 80%を下回らないように制御された.自動課題制御装置システム
(TaskForcer, O'hara & Co., Ltd., Japan)の中で前肢運動課題をラットに3日連続
(2−5時間/日)で訓練した.この課題では,ラットは頭部固定下で,右前肢で
スパウトレバーをプッシュすることによって課題を自発的に始めることができ,
レバーを0.5 秒以上ホールドした.ホールド期間の後,スパウトレバーをプルす
ると0.2-0.8 s の遅延後,報酬(0.1%サッカリン水,10µl)が与えたれた.