九州大学学術情報リポジトリ
Kyushu University Institutional Repository
物理的細胞創傷時の遊走前過程における細胞内PKCα トランスロケーション
荒井, 雅貴
https://doi.org/10.15017/1931901
出版情報:Kyushu University, 2017, 博士(工学), 課程博士 バージョン:
権利関係:
博士論文
物理的細胞創傷時の遊走前過程における 細胞内 PKCα トランスロケーション
平成30年3月
九州大学大学院 工学府 機械工学専攻
荒井 雅貴
i
目次
第1章 緒言 ... 1
1.1 血管内皮細胞についての研究意義 ... 1
1.2 血管内皮細胞の機能解析への研究アプローチ ... 2
1.3 血管内皮細胞が受ける力学刺激と細胞応答 ... 4
1.3.1 力学刺激による細胞内・細胞間シグナル伝達 ... 4
1.3.2 力学刺激による細胞の形態変化 ... 4
1.3.3 力学刺激による遊走促進 ... 5
1.4 Protein Kinase Cα ... 6
1.5 細胞遊走開始時の細胞内タンパク質の動態とPKCα... 7
1.6 本研究の目的... 7
1.7 本論文の構成... 8
第2章 血管内皮細胞に関する基礎事項と従来研究 ... 9
2.1 緒言 ... 9
2.2 血管の基本構造 ... 9
2.3 血管内皮細胞 ... 9
2.3.1 細胞遊走・創傷治癒 ... 9
2.3.2 血管内皮細胞の機能 ... 11
2.4 細胞間情報伝達 ... 13
2.4.1 パラクライン・オートクライン ... 13
2.4.2 ギャップ結合 ... 14
2.5 Protein Kinase C ... 15
2.5.1 PKCαトランスロケーションの時間と部位 ... 16
2.6 細胞の力学応答に関する従来研究 ... 19
2.6.1 力学刺激による細胞応答 ... 19
2.6.2 細胞間接着による機械的なシグナル伝達 ... 21
2.6.3 GPCRの構造変化 ... 21
第3章 実験方法 ... 23
3.1 はじめに... 23
3.2 実験方法... 23
3.2.1 細胞培養 ... 23
3.2.2 遺伝子の調整(PKCα-GFP,PKCα-DronpaGreen) ... 23
3.2.3 遺伝子の大量培養・抽出 ... 27
3.2.4 リポフェクション法による遺伝子導入 ... 27
ii
3.2.5 Western Blotting法 ... 28
3.2.6 免疫染色方法 ... 29
3.2.7 性質確認 ... 29
3.2.8 細胞への創傷と微小変形負荷方法・観察系 ... 29
3.2.9 共焦点顕微鏡 ... 30
3.2.10 薬剤負荷方法 ... 31
3.2.10 蛍光解析方法 ... 34
3.2.11 統計処理 ... 34
第4章 隣接細胞創傷時の細胞内Ca2+濃度変化とPKCαトランスロケーション ... 35
4.1 緒言 ... 35
4.2 実験結果... 35
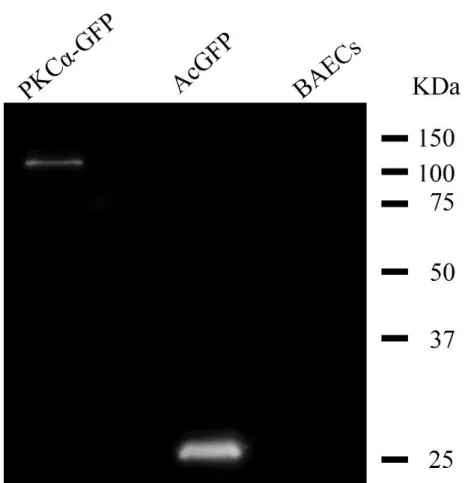
4.2.1 Western Blotting ... 35
4.2.2 PKCα - GFPの免疫染色 ... 37
4.2.3 PKCα - GFPの性質確認 ... 38
4.2.4 ATP負荷による細胞内Ca2+濃度変化とPKCα - GFPの蛍光変化 ... 39
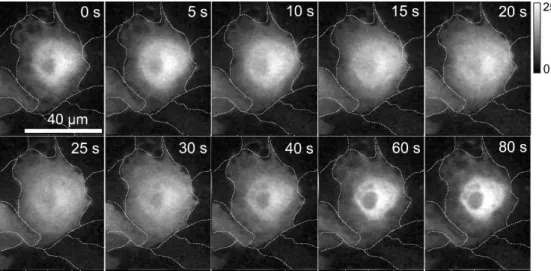
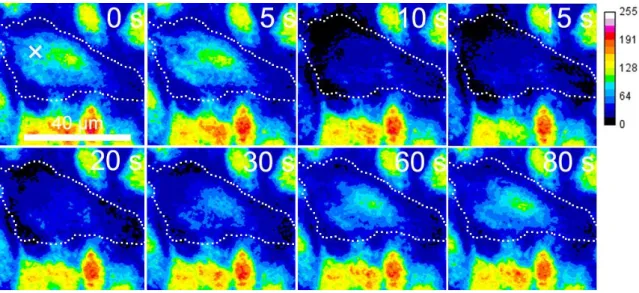
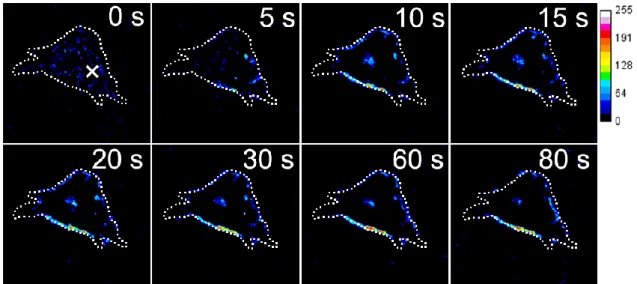
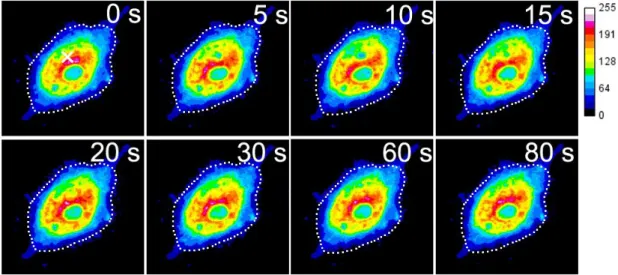
4.2.5 隣接細胞創傷時の細胞内Ca2+濃度変化とPKCα - GFPの蛍光変化 ... 41
4.2.6 共焦点顕微鏡を用いた隣接細胞創傷時のCa2+濃度変化とPKCα - GFPの蛍光変化 ... 44
4.2.7 創傷領域への細胞遊走 ... 45
4.3 考察 ... 48
4.3.1 薬剤刺激と隣接細胞創傷に伴う細胞内 Ca2+濃度変化と PKCα トランスロケーシ ョン ... 48
4.3.2 細胞内PKCαの二相性と細胞間情報伝達 ... 48
4.3.3 細胞内PKCαの二相性とメカノセンサーの関係 ... 49
4.4 結言 ... 49
第5章 局所力学刺激による細胞内Ca2+濃度上昇とPKCαのトランスロケーション .... 50
5.1 緒言 ... 50
5.2 実験結果... 51
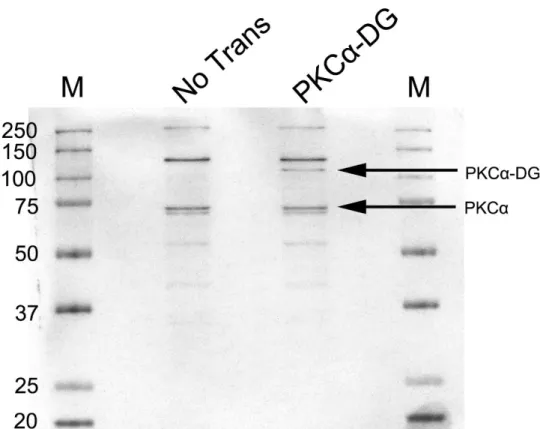
5.2.1 Western Blotting ... 51
5.2.2 PKCα – DGの免疫染色 ... 53
5.2.3 PKCα – DGの性質確認 ... 54
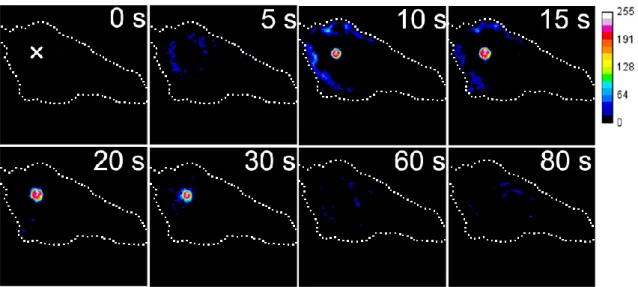
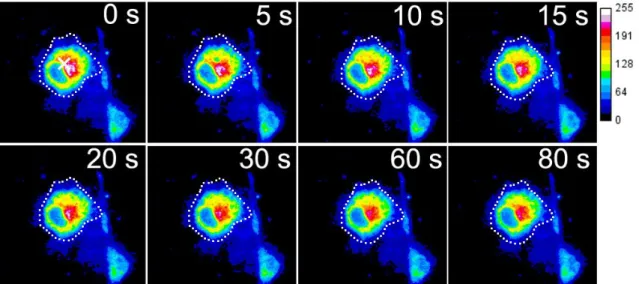
5.2.4 局所力学刺激によるCa2+濃度上昇とPKCαの二相性トランスロケーション .... 55
5.2.5 PKCα阻害剤負荷時の細胞内Ca2+とPKCα - DGの蛍光変化 ... 58
5.2.6 MSチャネル阻害時の細胞内Ca2+とPKCα – DGの蛍光変化 ... 62
5.2.7 細胞内・外Ca2+阻害時の細胞内Ca2+伝播とPKCα - DGの蛍光変化 ... 64
5.2.8 蛍光変化からのマイクロピペットの押し込み量と力の推定 ... 69
iii
5.2.9 局所力学刺激時のPKCα1次・2次反応まとめ ... 71
5.3 考察 ... 72
5.3.1 局所力学刺激によるPKCα の二相性トランスロケーションと細胞内・外 Ca2+の 関係 ... 72
5.3.2 PKCαの二相性トランスロケーションとCa2+・DAGの関係 ... 73
5.3.3 PKCαの二相性トランスロケーションとメカノセンサーの関係 ... 75
5.4 結言 ... 76
第6章 隣接細胞創傷時の二相性PKCαトランスロケーションと細胞間情報伝達 ... 77
6.1 緒言 ... 77
6.2 実験結果... 77
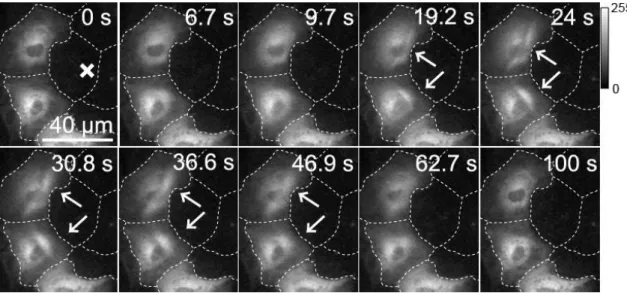
6.2.1 隣接細胞創傷時の細胞内Ca2+とPKCα – DGの蛍光変化 ... 77
6.2.2 PKCα阻害剤負荷時の細胞内Ca2+とPKCα – DGの蛍光変化 ... 80
6.2.3 細胞間情報伝達を阻害した時の細胞内Ca2+とPKCα – DGの蛍光変化 ... 84
6.2.4 細胞内・外Ca2+をそれぞれ阻害した時の細胞内Ca2+とPKCα – DGの蛍光変化 ... 91
6.2.5 PKCαの二相性トランスロケーションとDAGの関係 ... 98
6.2.6 創傷箇所への細胞遊走 ... 99
6.2.7 隣接細胞創傷時の細胞内PKCα1次・2次反応まとめ ... 107
6.3 考察 ... 108
6.3.1 細胞創傷によるPKCαの二相性トランスロケーションと細胞間情報伝達 ... 108
6.3.2 細胞創傷時の二相性トランスロケーションとCa2+・DAGの関係 ... 109
6.3.3 メカノセンサーと二相性トランスロケーション ... 110
6.3.4 細胞遊走とPKCαの二相性トランスロケーション ... 111
6.4 結言 ... 112
第7章 総括 ... 113
第8章 付録 ... 117
参考文献 ... 125
発表目録 ... 136
謝辞 ... 138
1
第1章 緒言
1.1 血管内皮細胞についての研究意義
血管は生命を維持するうえで必要不可欠な構造物である.血管はあらゆる臓器 に分布し,酸素や栄養素などの生命維持に必要な物質の供給を行う重要な役割 を担っている.それゆえ,血管の破綻や異常増殖が,がん,心疾患,脳血管障害 などの疾病と密接に関連する.1955 年以降,日本国内の死因上位にはがん,心 臓病,脳卒中が上げられる.これらは3大疾病と呼ばれ,治療法について研究が 進められている.3大疾病の治療法として血管内治療(カテーテル治療)[1]や血 管新生治療法[2]が注目されている.
血管はFig. 1-1のように内側から内膜(Intima),中膜(Media),外膜(Adventitia) の 3 層構造を持っており,血管の修復には血管の内膜を形成する血管内皮細胞 の生理機能が重要であることがわかっている.血管内皮細胞は血管の内側を単 層のシート状に基底膜と共に組織化しており,血管が創傷を受けると血管内皮 細胞は創傷領域に集団で遊走し修復する.この遊走機能は血栓形成を防ぐ血管 内皮細胞の重要な機能の1つと考えられており,細胞バイオメカニクスや細胞 組織工学の観点からも細胞遊走メカニズムを解明することは重要な意義がある と考える.
Fig. 1-1 血管の断面図
2
1.2 血管内皮細胞の機能解析への研究アプローチ
細胞の遊走や血管新生を誘発させる物質として,血管内皮増殖因子(Vascular Endothelial Growth Factor:VEGF)[3]や血小板由来成長因子(Platelet-derived growth
factor:PDGF)[4]などの血管増殖因子が重要な役割を果たしていることが報告さ
れている.そしてこれらの因子によって誘導される細胞内の複雑な代謝経路も 明らかになってきている[5].代謝経路の研究はWestern Blotting法などを用いた 研究によって明らかにされてきたが,免疫染色法によって細胞内構造の空間的 変化を観察する方法も用いられている.この方法は目的の分子を蛍光物質によ って標識するため,蛍光顕微鏡を用いて分子の分布を可視化することができる.
この方法によって内皮細胞は細胞内構造を変化させながら遊走することがわか っている[6].内皮細胞の遊走は細胞骨格であるアクチンが重要な働きをしてい る.アクチンはFig. 1-2 A-Cのように構造変化をしながら,Fig. 1-2 Dのように 動くことで進行方向に遊走する[6].近年では,緑色蛍光タンパク質(Green
Fluorescent Protein:GFP)を用いた研究によって生きた細胞内の時空間的な変化
の観察が可能になり,分子の運動や局在の観察が行われている.
3
Fig. 1-2 細胞遊走時の構造変化 [6]改編
(A)糸状仮足,(B)葉状仮足,(C)ストレスファイバー,(D)(ⅰ)糸状 仮足による刺激の感知,(ⅱ)葉状仮足を進行方向に伸ばす,(ⅲ)伸ばした仮 足の接着,(ⅳ)ストレスファイバーの収縮によって細胞内の構造を進行方向 に引き寄せる,(ⅴ)細胞後部を接着面から離し収縮する,(ⅵ)遊走関連分子 のリサイクル.
4
1.3 血管内皮細胞が受ける力学刺激と細胞応答
1.3.1 力学刺激による細胞内・細胞間シグナル伝達
血管内皮細胞は血液中の様々な化学的物質による刺激だけでなく,血流によ るせん断応力や血圧変動による張力などの力学刺激を受容するインターフェイ スとして働き,血管の恒常性を維持している.力学刺激を感知するメカノセンサ ーによって細胞外からの力学的な刺激を化学的なシグナルとして細胞内へ伝達 することで細胞遊走[7],分裂[8],成長[9]などの形態や機能の変化を伴う細胞応 答を起こすことがわかっている.力学刺激による細胞内の代表的な応答として 細胞内のCa2+濃度が上昇することが観察されている.この応答はせん断応力[10], 伸展刺激[11],マイクロピペットを用いたつつきによる局所的な力学刺激(微小 変形を加える)[12] [13],創傷領域を模擬するために細胞を削り取るようにスク ラッチしたとき[14],単一の細胞を創傷させることで発生することが観察されて いる[15].さらに,Ca2+濃度上昇は刺激を受けた細胞を中心に隣接細胞内のCa2+
濃度を上昇させ,周辺細胞に伝播していく現象(Ca2+伝播:約20~100 μm/s [16]) が知られている.この Ca2+伝播は細胞間の情報伝達手段の 1 つとして考えられ おり,細胞内のCa2+濃度が上昇することでAdenosin 5´ - triphosphate(ATP)など の分泌物が細胞外に放出され,その分泌物を隣接細胞が受け取ることで情報伝 達が行われるパラクライン作用と,細胞同士のヘミチャネルが結合することで 形成される Gap 結合を Ca2+や IP3が通ることで隣接細胞へ情報伝達する方法に よって引き起こされる.この Ca2+伝播は細胞の遊走やその後の細胞応答を誘発 させる要因であると考えられている.
1.3.2 力学刺激による細胞の形態変化
一般に内皮細胞は紡錘形あるいは多角形をしている.Naruse らは内皮細胞に 伸展刺激を加えると細胞内 Ca2+濃度が瞬時に上昇し,周期的な伸展刺激を 30 ~ 60 分加え続けると伸展方向とは垂直方向に細胞が配向することを示し,その過 程において細胞骨格であるアクチンが一旦分解され,細胞の配向と同じ向きに 再構築することを示した.伸展刺激による細胞内 Ca2+濃度上昇は Mechano-
Sensitiveチャネル(MSチャネル)の阻害と細胞外Ca2+の除去によって抑制され
たことから,伸展刺激によって MS チャネルが活性化されて細胞外 Ca2+が流入 したと考えられ,細胞の形態変化は MS チャネルがトリガーになっていると報 告している[17].Malekらは,内皮細胞にせん断応力を負荷すると 12 ~ 24 時間 後に細胞が流れ方向に沿って長軸を伸ばした紡錘形に配向し,アクチンも流れ
5
方向に沿って再構築することを報告している[18].このように力学刺激によって 細胞は細胞骨格を分解および再構築することで形態変化をすることが明らかに なっている.
1.3.3 力学刺激による遊走促進
創傷領域が発生すると,細胞は創傷領域に遊走し修復する.Sokabe らは,
Dimethylpolysiloxane(PDMS)薄膜上に細胞を 100%コンフルエントになるよう に培養し,人為的に創傷領域を模擬した後,薄膜に伸展刺激を加えると創傷領域 に隣接している細胞から Ca2+伝播が引き起こされると報告し,伸展刺激を加え ることで細胞の遊走が促進されると報告している.この細胞応答は創傷領域に 隣接している細胞のMS チャネルが伸展刺激を感知しATPを放出することで引 き起こされると報告している[11].Hsu らは細胞を 100%コンフルエントに培養 し,人為的に創傷領域を模擬した後,せん断応力を加えると細胞遊走は静置時よ りも促進されることを示し,流れの上流側にある細胞は下流側の細胞よりも遊 走が促進すると報告している.さらに,静置状態の細胞は 4 時間後に遊走を開 始したが,せん断負荷時の流れ上流側の細胞は 2 時間後に遊走を開始している ことを示した.これは流れの上流にある細胞はせん断応力によって細胞間接着 タンパク質である VE-Cadherin に結合している β-catenin の発現量が減少するこ とで細胞間接着が弱まり,遊走しやすい状態になっていると報告している[19]. 細胞が遊走するには細胞骨格がFig. 1-3のように空間的・時間的に変化する必要 があり,その調節を担っているのは低分子量 G タンパク質である.低分子量 G タンパク質であるRhoA[20],Rac[21],Cdc42[22]がそれぞれ細胞内の異なる部位 で働き,細胞骨格を分解・再構築している.RacとCdc42は細胞の進行方向先端 で活性化し細胞骨格の構築を促すことで進行方向への遊走を促進させる.一方,
RhoA は細胞後部で活性化することで細胞収縮および剥離をもたらすことが明 らかになっている.
このような従来研究によって細胞は力学刺激によって形態変化することがわ かっており,その形態変化には細胞内で異なるタンパク質が異なる部位で活性 化することで引き起こされているとわかっている.これらのことから,細胞機能 の解明には細胞内タンパク質の時空間的な分布を知ることが重要であると考え られる.
6
1.4 Protein Kinase Cα
細胞内タンパク質である Protein Kinase Cα(PKCα)は薬理学的な研究により 細胞の遊走や分化,増殖,アポトーシスなど様々な細胞応答に関与するセリン/ スレオニンリン酸化酵素である[23][24].PKCα は Ca2+感受性のタンパク質であ り,細胞内の Ca2+濃度上昇により細胞質から細胞膜へ局在を変化させる(トラ ンスロケーション)ことが報告されている[25].トランスロケーション後に,細 胞外刺激により活性化したG protein coupled receptor(GPCR)やReceptor tyrosine kinase(RTK)に含まれるPlatelet-derived growth factor receptor(PDGFR) が細胞
内のPhospholipase C(PLC)βとγをそれぞれ活性化し,細胞膜タンパク質であ
る Phosphatidylinoditol-4,5-bisphospate(PIP2)を加水分解することで産生される
Diacylglycerol(DAG)と結合することで PKCα は活性化することが報告されて
いる[26][27].これらの活性化はPKCαのN末端側にあるDAGが結合するC1ド
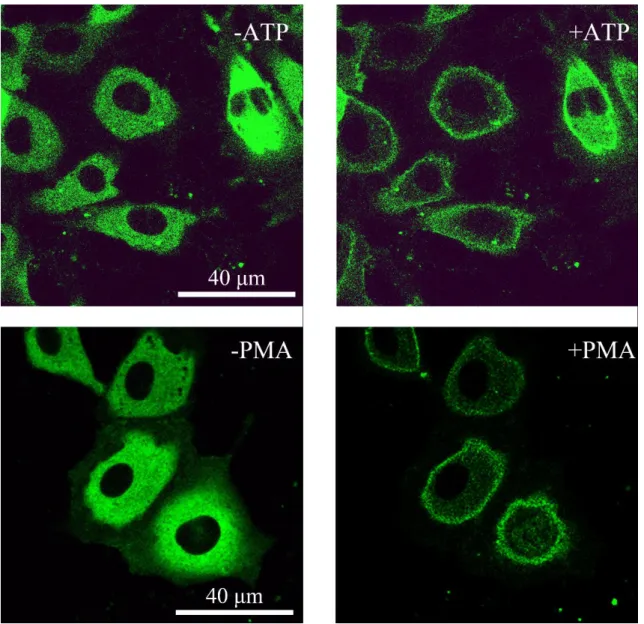
メインとカルシウムが結合する C2 ドメインによって制御されている.GFP を PKCαに融合させたタンパク質を用いた研究により,PKCαはATPによる生理的 な刺激により受容体を介して活性化された場合は,細胞質から細胞膜へすばや
く(15-30 秒)トランスロケーションした後に細胞質へ戻ってくる(1-2 分)一
過性のトランスロケーションであるのに対し,発がんプロモーターとして知ら れるPhorbol 12-myristate 13-acetate(PMA)による刺激では細胞質から細胞膜へ ゆっくり(3-5分)とトランスロケーションし,そのまま細胞膜に局在し続ける 不可逆的な反応を引き起こすことが明らかになっている.PKCαのトランスロケ ーション時間は刺激する試薬によって異なることから,PKCαのトランスロケー ションメカニズムには様々な経路が存在することが考えられ,PKCαトランスロ ケーションの時間的・空間的な変化が様々な細胞応答を制御している可能性が 考えられている.近年,GPCRやRTK,イオンチャンネル,カベオリンなどが力 学刺激に反応するメカノセンサーであるとの報告がされている[28][29].これら のメカノセンサーが外部から刺激されることでPKCをトランスロケーションさ せることが Western Blotting 法を用いた実験によって報告されている[28].しか
し,Western Blotting 法用いた実験では刺激直後のトランスロケーション反応を
時空間的に調査するのは難しい.
7
1.5 細胞遊走開始時の細胞内タンパク質の動態と PKCα
細胞の遊走はEpidermal Growth Factor(EGF)などの成長因子によって誘発さ れることがわかっている[3].細胞にEGFを負荷するとRac1とCdc42が1 分以 内に活性化を始め,遊走方向の先端に局在し,3 分後に活性化のピークに達しそ の後元に戻ると報告されている[30].その時に葉状仮足もEGF負荷後 1 分以内 に伸びはじめたと報告している.このことから,成長因子による細胞遊走の誘発 は 1 分程度で起こると考えられる.同じく細胞遊走を誘発すると考えられてい る Ca2+濃度上昇は数秒で起こるため,成長因子よりも早い段階で作用している と考えられる.
PKCαを阻害すると細胞遊走が抑制されることから,PKCαは細胞遊走に関与 するタンパク質だと考えられている[23].PKCαは活性化することで VE-cdherin を分解し細胞間接着を弱め[31],細胞骨格の制御に関与することで[32],細胞遊 走を促進する報告がされている[33].さらに,PKCαはCa2+濃度上昇と同時に活 性化することが薬剤を用いた研究によって報告されている[34].これらのことか ら細胞創傷に伴うCa2+濃度上昇によってもPKCαは活性化することが予想され,
PKCαがRac1やCdc42よりも早い段階で細胞遊走の誘発に関与している可能性
がある.しかし,細胞創傷に伴う Ca2+濃度上昇と PKCα トランスロケーション を調査した報告はなく,隣接細胞が創傷を受けた直後の細胞内の Ca2+濃度上昇 と PKCα トランスロケーションを観察することは,創傷領域を修復するための 細胞遊走の起点の解明に重要であると考えられる.
1.6 本研究の目的
細胞は,隣接細胞が物理的創傷を受けることで,創傷数秒後に細胞内 Ca2+濃 度が上昇する.そして,創傷数時間後に細胞骨格を再構成することで創傷領域へ 遊走を開始し修復する.その際,遊走細胞の前方部や後方部には特定のタンパク 質が局在し遊走を調節している.これらのことから,細胞内物質が時空間的に変 化することで細胞遊走を調節していると考えられるが,細胞遊走メカニズムの 詳細に関しては依然として不明な点が多い.本研究で着目する PKCα は Ca2+濃 度依存性のタンパク質である.そのため,隣接細胞の創傷による Ca2+濃度上昇 によっても同時に活性化されていることが予想される.また PKCα は刺激薬剤 によって局在する部位や時間が異なることや,PKCαの活性化は細胞骨格の形成 にも関与していることから,PKCαトランスロケーションの時空間的に異なる分 布が異なる細胞応答を引き起こしている可能性がある.そこで本研究では物理 的細胞創傷から細胞遊走初期に至る過程に PKCα トランスロケーションが深く
8
関与していると考え,PKCαトランスロケーションのライブイメージングする ことにより,細胞遊走初期過程における PKCα トランスロケーションのメカニ ズムについて検討をおこなった.
1.7 本論文の構成
第 1 章では本研究の背景および目的について述べた.第 2 章では血管内皮細 胞が物理的創傷を受けて,細胞遊走を開始する際の基礎事項および従来研究に ついて述べた.また,PKCαの基礎事項および従来研究について述べた.さらに,
細胞の力学応答および細胞間情報伝達機構についての基礎事項および従来研究 について述べた.第 3 章では本研究で行なった実験方法についてまとめた.本 研究では,内皮細胞に物理的創傷を人工的に与えるために,マイクロピペットに より物理的に細胞膜を破壊する方法を採用した.また,5章以降で検討する細胞 の力学応答を計測するために,マイクロピペットを用いて直接細胞を微小変形 させる方法を採用した.第 4 章では,隣接する細胞に物理的創傷を与えた際の 細胞内 Ca2+濃度上昇と PKCα トランスロケーションの動態を同時にリアルタイ ムで観察し,ATP 負荷時の動態と比較検討した.ATP を負荷すると Ca2+濃度上 昇に伴い細胞質に分布していた PKCα は細胞膜全体に一様に分布を変化させ,
従来の報告と一致した.マイクロイペットにより隣接細胞に物理的創傷を与え ると,ATP負荷と同様に PKCαはCa2+濃度上昇に伴って細胞膜全体でトランス ロケーションが引き起こされた.しかし,その後に創傷側でのみPKCαが局在し 続ける二相性のトランスロケーションが確認された.この遊走前過程における PKCαの二相性を引き起こすメカニズムとして,(1)隣接細胞の創傷による細胞 間張力の変化によって細胞のメカノセンサーが刺激され引き起こされる可能性 と,(2)パラクライン作用やGAP結合による細胞間情報伝達機構が重要な働き をしている可能性の2つが考えられた.そこで第 5 章では,(1)の力学刺激と PKCαの関連を調べるために,単一細胞に局所的な力学刺激(微小変形)を加え 細胞内 Ca2+濃度と PKCα の動態を観察した.局所的に微小変形を加えると刺激 位置近傍の細胞膜ではCa2+濃度上昇と同時にPKCαがトランスロケーションし,
刺激位置では PKCα が局在し続ける二相性が観察されたことから,PKCαは力 学刺激によってトランスロケーションすることが示された.第 6 章では 5 章の 結果を踏まえ隣接する細胞を創傷させた時の細胞内の Ca2+濃度と PKCα の動態 を計測した.(2)がPKCαの局在に関与しているか調べ,メカノセンサーの阻害 実験も行った.その結果,細胞間情報伝達機構を阻害してもPKCαの局在は確認 された.第7章は総括であり,各章の結言についてまとめを行い,本研究によっ て明らかになったことを記述する.
9
第2章
血管内皮細胞に関する基礎事項と従来研究 2.1 緒言本章では,本研究を理解する上で必要となる基礎事項と,内皮細胞の力学応答 に関する従来研究について説明する.
2.2 血管の基本構造[35]
血管は,心臓から拍出される血液を末梢に送り出す動脈,末梢から心臓に血液 を送り返す静脈,動脈と静脈の間にあり動脈血中の酸素と栄養素を各組織に供 給する毛細血管の 3 種類に大別できる.動脈と静脈の血管壁は内膜(Intima),
中膜(Media),外膜(Adventitia)の 3 層の構造からできている.内膜は血管内
皮細胞と血管基底膜,中膜は平滑筋細胞と弾性繊維や膠原線維,外膜は主に膠原 線維により構成されている.これらの膜は内弾性板と外弾性板によって仕切ら れており,血管に弾性を付与している.一方,毛細血管は中膜が無く,内皮細胞 と周皮細胞(Pericyte)とによって構築されており,血管基底膜に包まれて直接 接触している.Pericyte は収縮能力示し血流を調節することができる[36].この ような血管が毛細血管網を構成し,盛んに血管新生が起こる.
2.3 血管内皮細胞
2.3.1 細胞遊走・創傷治癒
成長した組織における細胞は遊走や増殖が抑制または行われない状態(静止 期)になっているが,必要に応じて活発に遊走・増殖する能力を有している[37]. 創傷を受けた場合は組織の恒常性を回復させるために様々な細胞内および細胞 間の経路を活性化し創傷治癒を行う[38][39].大血管における血管内皮単層が創 傷(剥離など)すると周囲の細胞が遊走して創傷箇所を修復する(再内皮化)こ とが観察されている.この過程は線維芽細胞増殖因子(Fibroblast growth factors: FGF)と呼ばれる成長因子によって促進される[40].この成長因子によって細胞 は創傷領域に向かい遊走する.創傷領域に隣接している細胞はリーダー細胞と して機能し後続の細胞を牽引する[40][11].この牽引力は後続の細胞に伝播し細 胞層として移動することによって創傷治癒が行われる.後続する細胞への伝播
は VE-cadherin や α-catenin といった細胞間を繋ぐ役目を果たしているものによ
って調整されている[41].
10
細胞はFig. 1-2のように形態を変化させながら遊走する.このとき細胞内では
酵素やタンパク質などが異なる部位に局在する極性化がみられる.走化性の実 験において,細胞の先端で PI3 キナーゼが活性化され,細胞前部の PIP3濃度が 上昇し,PIP3依存性グアニンヌクレオチド交換因子(Guanine nucleotide exchange
factor : GEF)を介しRhoファミリーである低分子量Gタンパク質が活性化する
ことで細胞前部のアクチン重合が誘導されることが報告されている[42].この時,
低分子量Gタンパク質であるCdc42は細胞先端で活性化され細胞骨格形成因子 に作用するWiskott-Aldrich syndrome protein(WASP)に結合し,糸状仮足を形成・
制御する[43],Rac は好中球において細胞前部で活性化され,WASP-family verprolin-homologous protein(WAVE)を活性化することで葉状仮足の形成・調節 をする[44].対照的に,RhoA は細胞後部側で活性化し,細胞後部を収縮させる と考えられている.しかし,近年ではRhoAが先端の突出に関与していることが 示唆されている[45].
細胞遊走に関与しているタンパク質などの局在を薬剤刺激によって観察して いる従来研究をTable. 2-1に記載する.
Table. 2-1 薬剤刺激による遊走因子の局在と時間
因子 反応開始 反応ピーク 局在箇所 観察方法 薬剤 細胞種,文献
PI3K 8 秒 - 細胞膜 FM* cAMP Dictyostelium [46]
IP3 - 約20 秒 細胞膜 FM* Bradykinin Endothelial [47]
Rac1 <1分 3分 葉状仮足 FRET EGF Cos1, A431 [30]
Cdc42 <1分 3分 葉状仮足 FRET EGF Cos1, A431 [30]
RhoA <1分 1.5分 葉状仮足 FM* Thrombin HUVEC [48]
*FM: Fluorescence microscope
11
2.3.2 血管内皮細胞の機能[49][37]
血管内皮細胞の生理機能として以下の4つに大別できる.
(1)物質透過性の調節:内皮細胞はお互いに接着し,血液が外に漏れないよ うにしている.内皮細胞間の接着としてタイトジャンクション(tight junction)・アドへレンスジャンクション(adherens junction)・ギャップ ジャンクション(gap junction)・シンデスモス(syndesmos)が知られて いる.一方で,特定の物質を血管内外で受け渡しする物質輸送の仲介を し,不必要に血液の成分が通過することを防ぐ選択的透過性を持つ.物 質輸送の経路としては,細胞間をすり抜ける経路と,いったん血管内皮 細胞に取り込まれるトランスサイトーシスとがある.
(2)血管緊張性の調節:血管内皮細胞は,血管拡張に作用する一酸化窒素
(NO),プロスタグランジン(PGI2),内皮細胞由来過分極因子(EDHF) と,血管収縮に作用するエンドセリン(EDCF)などの物質を生産・分 泌し,拡散により周囲を取り囲む平滑筋細胞に作用することで血管の 緊張性を調節している.NOはせん断応力などの刺激で産生が増加する ことが知られている.
(3)抗血栓性:血管内皮細胞は生理的状態では強い抗血栓性を発揮してい る.これは血管内皮細胞から生成される PGI2,ヘパラン硫酸プロテオ グリカンに結合したアンチトロンビンや組織因子経路インヒビター
(TFPI),血管内皮細胞膜タンパク質であるトロンボモジュリンなどに 依存している.
(4) 血管新生:血管内皮細胞の血管新生は細胞の遊走と増殖によって行わ
れる.Fig. 2-1に示すように,基底膜を局所的に分解し,内皮細胞が分
解箇所に遊走・増殖することで血管を形成する.
12
Fig. 2-1 血管新生の過程 [49]
①何らかの刺激が血管に加わる.②刺激を受けた内皮細胞は基底膜を分解し 遊走を始める.③他の細胞が追従するように遊走,増殖する.④内皮細胞の管腔 が形成される.
13
2.4 細胞間情報伝達
血管内皮細胞は各種の生物活性物質の産生,代謝,炎症・免疫への関与,血管 新生などの生理的機能を持っている.これらの機能は 1 つの内皮細胞の働きに よるものではなく,細胞同士が集団として統一した反応をすることで機能して いる.このためには細胞間での物質の交換,また 1 つの細胞で起きた現象を周 囲の細胞と共有することで反応を同期させる必要がある.従って細胞間での情 報伝達が重要な役割を担ってくる.シグナルの性質にかかわらず細胞は,外部や 細胞内からの刺激を受容体と呼ばれるタンパク質で受け取る.受容体はシグナ ル分子(細胞間および細胞内のシグナルに関わる分子)と特異的に結合すること で活性化し,細胞内に情報を伝達する.細胞間情報伝達の方法としては下記のも のが挙げられる.
2.4.1 パラクライン・オートクライン
シグナル細胞(signaling cell)が情報伝達物質を放出し,隣接した標的細胞
(target cell)に作用するパラクライン型(paracrin, 傍分泌)と,放出した物質を
再度自ら取り込むオートクライン型(autocrine, 自己分泌)がある(Fig. 2-2).
Fig. 2-2 パラクライン・オートクライン作用によるシグナル経路
14
2.4.2 ギャップ結合
ギャップ結合(gap junction)は隣接する細胞をつなぐ通路であり,細胞間で物 質を直接通過させることが出来る.1つの細胞の細胞膜にはギャップ結合の構成 要素であるヘミチャネル(hemichannel)が存在し,隣接する細胞のヘミチャネル が結合することでギャップ結合が形成される.ギャップ結合のヘミチャネルは,
膜貫通型タンパク質のコネキシン(connexin:Cx)の六量体で構成されている.
Cxはいくつかの種類が存在し,その組み合わせで結合の有効性が決まる.Huら [50]によると確認されている有効な組み合わせは,Cx43のみ,Cx40のみ,Cx37 とCx43,Cx37と Cx40の組み合わせである.またCxは内部に親水性の孔を持 っている.約分子量1000 Da(1 Da:水素原子1個の質量とほぼ等しい)以下の 無機イオンや情報伝達物質は,親水性の孔を通過することによって細胞間の情 報伝達を行っている(Fig. 2-3).
Fig. 2-3 Gap junctionによるシグナル経路
15
2.5 Protein Kinase C
プロテインキナーゼC(Protein Kinase C:PKC)は生体のほとんどの組織・細 胞に発現しているセリン-スレオニンリン酸化反応を触媒とする酵素で,ウシ小 脳に存在するサイクリックヌクレオチド非依存性プロテインキナーゼとして,
1977年に Nishizukaらにより発見された[51].一般にPKC は GPCRや RTKs な どの受容体刺激により細胞膜から産生されるDiacylglycerol(DAG)と,同時に 産生されるInositol trisphosphate(IP3)により誘導されるCa2+によって活性化さ れる.また PKC は,発ガンプロモーターである Phorbol 12-myristate 13-acetate
(PMA)によって特異的に活性化することが明らかになったことから,PMAを 用いた研究により増殖や分化,アポトーシスなど多くの細胞応答に関与してい ることが明らかにされてきた[52].
PKC は現在までに 10 種類のアイソフォームが同定されている.PKC の構造 と活性化の方法により大きく 3 つのサブファミリーに分類することができる.
通常型PKC(conventional PKC:cPKC)は,DAGとPMAが結合するC1ドメイ ン(C1AとC1B),Ca2+が結合するC2ドメインが存在し,DAG,Phosphatidylserine
(PS),Ca2+によって活性化する.新規型 PKC(novel PKC:nPKC)は,DAG, PS を必要とするが C2 ドメインが欠損しているために Ca2+不応答であり,非典 型PKC(atypical PKC:aPKC)はC2ドメインの他にC1ドメインが一つ欠損し ているため,Ca2+,DAG,PMA のいずれによっても活性化されない.これに対 して触媒ドメインは各アイソフォーム間で違いはみられず,C3 ドメインには ATP 結合部位,C4ドメインには基質認識部位が存在する(Table. 2-2, Fig. 2-4) [52][53].
Table. 2-2 PKCのアイソフォームと活性化因子[53]より改編
サブファミリー アイソフォーム 活性化に必要な因子
cPKC(conventional) α,β1,β2,γ DAG,PS, Ca2+
nPKC(novel) δ,ε,η,θ DAG,PS
aPKC(atypical) ι/λ,ζ PS
16
Fig. 2-4 PKCサブファミリーの構造
2.5.1 PKCα トランスロケーションの時間と部位
PKCαはcPKCに分類されるアイソフォームの1種であり,GPCRやRTKsな どの受容体刺激によって産生されるDAGと同時に産生されるIP3により誘導さ れるCa2+によって活性化される(Fig. 2-5).
Fig. 2-5 PKCαのシグナル経路
17
細胞内 Ca2+濃度が上昇すると Ca2+が C2 ドメインに結合し細胞膜へと移動す る(トランスロケーション).細胞膜へ移動後に DAG と結合することで偽基質 が触媒ドメインから外れることで構造が変化し活性化する[54][55].(Fig. 2-5).
PKCαトランスロケーションはGPCRやRTKを介した刺激の種類によって,
移動部位や移動時間が異なると報告されている(Table. 2-3)[56]. 従来研究に よってPKCαは細胞増殖やアポトーシス,分化,遊走および細胞接着など様々な 機能を制御すると報告されている(Fig. 2-6)[57][58].
Fig. 2-6 PKCαのトランスロケーションモデル[54]改編
18
Table. 2-3 様々な薬剤刺激によるトランスロケーションの時間-空間的変化
薬剤刺激 濃度 トランスロケーション 場所
トランスロケーシ ョン時間
Ionomycin 100 nM 細胞質,細胞膜 40 分
KCl 40 mM 細胞膜 30分
Bay K 8664 100 nM 細胞膜の特定の領域 2 ~3分
Thapsigargin 100 nM 細胞質,核周辺 6 ~7分
Ryanodine 100 mM
細胞膜の特定の領域(継代数0)
60 秒 核周辺(継代数6)
IP3 100 nM 細胞膜,細胞質の
特定の領域 10分
Thrombin 0.1 U/ml 細胞膜の特定の領域 2分
PDGF 1 ng/ml 細胞膜 8 ~ 9分
Fig. 2-7 PKCαの様々な生体内機能への関与 [59]改編
19
2.6 細胞の力学応答に関する従来研究
細胞の力学応答を観察するために,マイクロピペットなどで細胞をつつく方 法や流れによるせん断応力,薄膜上に細胞を播種し引っ張ることで伸展刺激を 加える方法などが行われている.
2.6.1 力学刺激による細胞応答
内皮細胞にガラスマイクロピペットを用いてつつくことで刺激を加えたとき に,細胞内のCa2+濃度が上昇し,周囲の細胞に伝播していく様子(Ca2+伝播)が 観察されている[12][13].この現象は GAP 結合を阻害すると伝播範囲が 50%程 度に抑制され,apyraseによって細胞外のATPを除去することでパラクライン作 用を阻害すると 伝播範囲が 15%程度にまで減少する[13].また,細胞内の小胞
体内 Ca2+を Thapsigargin によって枯渇させても Ca2+伝播は抑制されることが報
告されている[60].
血管内皮細胞に生体内の血流を模擬したせん断応力を加えると,直後に細胞 内のCa2+濃度上昇[10]やATPの放出[61]が見られ,長時間(4時間など)せん断 応力を加え続けると流れ方向に沿って形態が変化すること[62]などが報告され ている.このような細胞のせん断応力によるアクチンの再構築には細胞内 Ca2+
とチロシンキナーゼの活性化が重要だと報告されている[18].細胞の接着斑
(Focal adhesions : FAs)も細胞の形態変化に応じて局在を変化させることが報告
されており[63],接着班の構成物質であるIntegrinの局在変化も観察されている [64].Integrinの細胞質側ではIntegurin結合タンパク質として知られているFocal adhesion kinase(FAK)が結合しており,FAKのリン酸化によって下流にあるPI3K やRho,c-Jun NH2-terminal kinase(JNK),Extracellular Signal-regulated Kinase(ERK) などが活性化される.JNK は細胞の遊走を制御していると報告されており[65], ERKは細胞の運動性,細胞膜の突出を制御していると報告されている[66].
PDMS 膜上に細胞を培養して引っ張ることで伸展刺激を加えると,細胞内の Ca2+濃度の上昇[17]や,伸展刺激方向とは直角に細胞の配向が変化することが報 告されている[67].この反応はメカノセンサーが刺激を感知することで細胞が応 答していると考えられており,Mechanosensitive チャネル(MS チャネル)阻害 剤である Gadolinium Chloride Hexahydrate (Gd3+)を負荷すると抑制されること が報告されている[67].
20
遊走に関与しているタンパク質などの反応時間・局在を力学刺激によって観 察している従来研究をTable. 2-4に記載する.
Table. 2-4 力学刺激による遊走因子の局在と時間
因子 反応 開始
反応ピーク 局在箇所 観察方法 力学刺激 細胞種,文献
K+ channel 数秒 約10秒 細胞全体 Patch-Clamp Shear stress Endothelial [68]
Ca2+ 数秒 約15秒 細胞全体 FM* Shear stress HUVEC [69]
Ca2+ 数秒 細胞全体 FM* Stretch Endothelial [17]
PI3K - 1分 Peripheral
membrane
FM* Stretch Capillary
Endothelial [70]
IP3 - 5分 細胞全体 Chromate graphy
Shear stress Endothelial [47]
IP3 数秒 約10秒 細胞全体 FRET Poke HSY-EA1 [71]
PLC 数秒 約7秒 細胞全体 FRET Poke MDCK [72]
Rac1 5 分 30 分 - Western Shear stress BAECs [73]
Rac1 - 30 分 遊走方向先
端
FRET Shear stress BAECs [73]
Rac1 1分 5分 細胞全体 Western Stretch HASMCs [74]
Cdc42 5分 30分 遊走方向先
端
FRET Shear stress BAECs [22]
Cdc42 1分 5分 Membrane Western Shear stress BAECs [75]
RhoA 1分 - Membrane Western Shear stress BAECs [75]
RhoA - 10分 細胞全体 FRET Stretch NIH3T3 [76]
RhoA 1分 5分 細胞全体 Western Stretch HASMCs [74]
Integrin αvβ3
5分 - Apical
membrane
Immunostaining Shear stress Swiss 3T3 [64]
FAK 5分 1分 - Western Shear stress BAECs [77]
ERK 10分 - - Western Shear stress BAECs [77]
JNK 30分 - - Western Shear stress BAECs [77]
*FM: Fluorescence microscope