1
論文内容の要旨
学位論文題名
転写活性を指標とした内因性・外因性リガンドの作用予測に関する検討
Prediction for pharmacological actions of ligands based on transcriptional activities.学位申請者 二若 久美 ㊞
序章
内分泌・代謝系は、生体における恒常性維持に欠かせないシステムとして、神経系や免疫系と協調して機 能を発揮する。内分泌・代謝系の作用は、受容体に対するリガンドの結合によって制御されることが多い。 リガンドは作用する状況下によって、ホルモン、神経伝達物質、サイトカイン、増殖因子などの名称で分類 される。内因性リガンドの一つであるホルモンの古典的な定義は、標的細胞から離れた場所でごく微量分泌 されて血流によって運ばれ、標的細胞に存在する受容体に高い親和性を持って特異的に結合する物質のこと を言う。今日までに数多くのホルモンが同定され、その生理作用が明らかになってきた。しかしながら、古 典的とされるホルモンにおいても新たなシグナル伝達経路や生理作用が発見されることは少なくなく、未だ に数多くのホルモンについて未解明の点が多い。また、医薬品として利用されている外因性リガンドにおい ても、近年になって新たな生理機能が明らかになり、多面的作用として効果が追加されることも珍しくない。 同様に、主にアジア圏で古くからハーブや漢方の構成成分として利用される植物には、脂質、糖質代謝改善 効果を持つことが経験的に知られているものが存在する。しかし、その有効成分と作用メカニズムが明らか になっているものは僅かである。 申請者が所属する研究室では、内分泌・代謝領域において、特にホルモンやホルモン受容体に対するリガ ンドに注目し、未知の生理的機能や病態形成、及びその改善作用について検討を行ってきた。本論文は、既 存のリガンドが持つ未知の生理的機能や疾患に対する改善効果を、転写という視点で検討しまとめたもので ある。第一章では、医薬品としても利用されている内因性リガンドである成長ホルモン (growth hormone; GH) の生理作用について、新たなシグナル伝達経路の解明と生理作用の可能性について詳述する。第二章では、 外因性リガンドとして、医薬品として汎用されているビタミン D3製剤について、最適な薬物治療を目指して 比較検討を行った。具体的には、活性型ビタミン D3に対するビタミン D 依存性クル病 (vitamin D dependent rickets type II; VDDRII) の治療効果を、転写活性という視点からの予測を試みた。第三章では、アジア圏の薬 用資源であり、古来からよく利用される漢方の一つである桂皮 (Cinnamomum cassia) の主要成分である cinnamaldehyde (CA) が、外因性リガンドとして内分泌・代謝領域に及ぼす効果、特に CA のもつ抗糖尿病効 果の作用メカニズムについて、転写活性を軸に検討を行った。2
第一章
転写活性に基づく GH の内分泌代謝領域における生理作用の予測
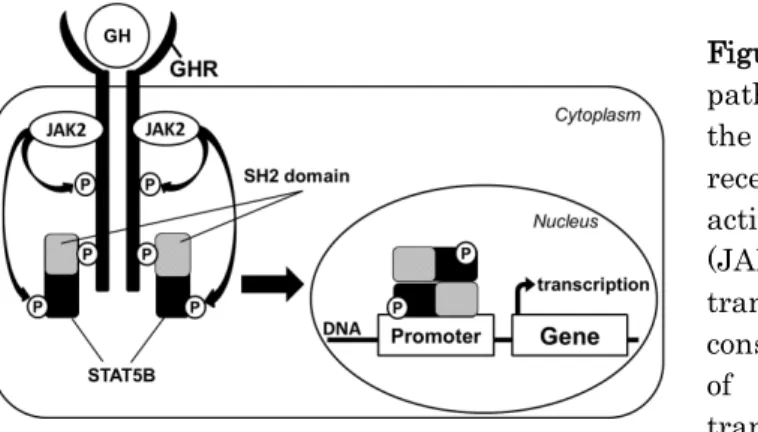
序論GH は、脳下垂体前葉の GH 分泌細胞 (somatotrophs) から分泌されるホルモンである。GH が肝細胞膜上の GH 受容体 (GH receptor; GHR) に結合すると、janus activation kinase 2/signal transduction and activator of transcription (JAK2/STAT) 経路が活性化され、insulin-like growth factor-1 (IGF-1) 産生を促進する。産生された IGF-1 は、肝臓 由来の成長因子として骨の成長などに重要な役割を果たしている [1]。このように GH の作用は、その大部分が IGF-1 による間接的作用であると考えられてきたが、近年、GH が直接的作用を有することがわかってきた [2,3]。GH の直接 的作用は全身に発現する GHR を介したものであり、筋組織や脂肪組織等に作用し、エネルギー代謝を促すことが報 告されている [2,3]。しかしながら、GH のエネルギー代謝に関する直接的作用がどのようなシグナル伝達経路を介し て標的遺伝子に関わっているのか、未解明の部分が少なくない。 GHR を介したシグナル伝達経路では、まず GHR への GH の結合により、JAK2 がリン酸化される。それに引き続き、 STAT5、mitogen-activated protein kinase (MAPK)、及び insulin receptor substrate (IRS) を介して、
phosphatidylinositol-3 kinase (PI3K) 経路などが活性化される [4]。GH で活性化される転写因子の一つである STAT5 には、STAT5A、STAT5B の 2 つの相同体が存在し、アミノ酸レベルで 90%以上の相同性を持つ[5,6]。これら は共通のコンセンサス配列 (TTCnnnGA) に結合し機能的冗長性をもつが、それぞれのノックアウトマウスは異なる表 現型を示す。STAT5A ノックアウトマウスはプロラクチン感受性欠失により雌で乳腺発達不全を呈するが [7,8]、 STAT5B ノックアウトマウスは GH 感受性欠失による成長遅延を示す [8,9]。これらの知見から、GH によって活性化さ れる転写調節因子は主に STAT5B であると考えられている。成人において、GHR はあらゆる組織に広く発現しており、 上記のシグナル伝達経路を介した GH の作用は多岐にわたることが報告されている。上述の GHR/JAK2/STAT 経路 により、脂肪前駆細胞における脂肪産生は促進されるが [10]、一方で、成熟脂肪細胞において脂肪合成を抑制し、 ホルモン感受性リパーゼを活性化することで脂肪分解を促進することがわかっている [11]。また、臨床的には、小児 期に GH 補充療法を受けて標的身長となった成人患者において、GH 投薬を終了した後では、内臓脂肪が増加や脂 質代謝異常症が高頻度に合併することが報告されている (成人成長ホルモン欠乏症、adult GH deficiency; aGHD) [12–14]。
既報における KK-Ay 糖尿病モデルマウスへの GH 投与実験では、骨格筋組織などにおいて、ミトコンドリア脱共役 タンパク質 (uncoupling protein; UCP) の発現量が増加したと報告されている [15]。このことから、GH 下流のシグナ ル伝達経路により UCP 発現が促進されたことが推察されるが、この作用が GH の直接的作用なのか、IGF-1 を介した 間接的作用なのかは明確ではない。そこで本研究では、in vitro において GH/JAK2/STAT 経路の下流に存在するシ グナル伝達経路を明らかにし、GH によるエネルギー代謝への直接的関与の有無と、関与があればそのメカニズムを 明らかにすることを目的として検討を行った (Figure 1)。
3
Figure 1. GH signal transduction pathway through STAT5B. GH binds to the transmembrane growth hormone receptor (GHR) resulting in the activation of janus-activated kinase2 (JAK2) and STAT5B. STAT5B then translocates to the nucleus, binds to the consensus sequences on the promoters of target genes, and regulates transcription.
第一節 UCPs プロモーター上の STAT5B コンセンサス配列の探索とプロモーターアッセイ 【目的】
恒常活性型 STAT5B (activated STAT5B; aSTAT5B) を用いて、エネルギー代謝関連遺伝子 UCPs の発現調節に ついて、プロモーターアッセイで検討を行った。
【材料及び方法】
UCP2promoter-luciferase (UCP2pro-Luc) 及び UCP3promoter-luciferase (UCP3pro-Luc)は、ヒト UCP2 及び UCP3 プロモーター領域をヒトゲノム DNA より増幅し、ホタルルシフェラーゼ発現ベクターにクローニングして構築した。さら に、UCP2 及び UCP3 プロモーター上の STAT5B コンセンサス配列 (TTCnnnGA) を Basic Local Alignment Search Tool (National Center for Biotechnology Information) によって確認し、様々な長さの UCPs プロモーターコンストラクト を構築した。リン酸化を経ずに活性化されている aSTAT5B 発現ベクターとして、299 番目のヒスチジンをアルギニンに、 711 番目のセリンをフェニルアラニンにそれぞれ置換した pCMX-aSTAT5B (aSTAT5B) を用いた [16]。また、内在性 コントロールプラスミドとして pGL4.70 (Promega Corporation) を使用した。ヒト胎児腎細胞株 HEK293 由来 TSA201 細 胞 (JCRB 細胞バンク) は、ダルベッコ変法イーグル培地 (Dulbecco’s modified eagle’s medium; DMEM) (日水製薬 株式会社) [10% ウシ胎児血清 (fetal bovine serum; FBS)、100 U/ml ペニシリン G、100 μg/ml ストレプトマイシン硫酸 塩、2 mM L-グルタミン含有] で、37°C、5%CO2条件下で継代培養を行った [17]。12 well plate に播種した TSA201 細胞に、UCP2pro-Luc もしくは UCP3pro-Luc を 100 ng/well、aSTAT5B もしくは pCMX (mock) を 200 ng/well、 pGL4.70 を 20 ng/well をリン酸カルシウム法で同時導入し、6 時間培養後にフェノールレッド不含、活性炭処理済 10% FBS 含有 DMEM に培地交換した。さらに 20 時間培養後、細胞を回収して処理し、ルシフェラーゼ活性を測定した。 データは平均値 ± S.D.で表す。統計解析は、SPSS 17.0 software (IBM) を使用し、群間比較は一元配置分散分析 の後、Bonferroni 法で多重比較検定を行った。
【結果】
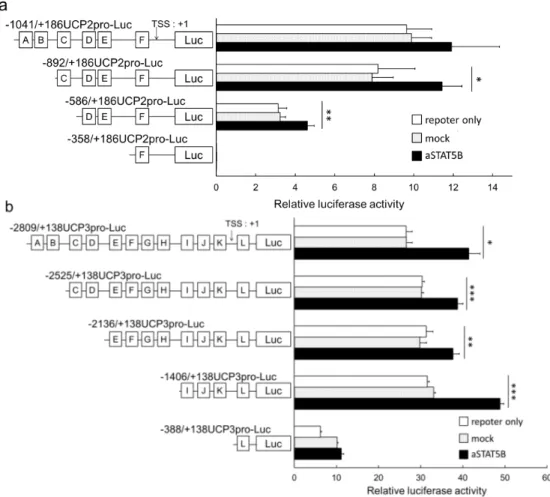
aSTAT5B による UCP 遺伝子の転写活性について、UCP2pro-Luc、UCP3pro-Luc 及びそれぞれのコンストラクトを 用いて検討した結果、-358/+186UCP2pro-Luc で基本転写活性が消失した (Figure 2a)。また、
-388/+138UCP3pro-Luc で、aSTAT5B による転写活性の増加が認められなかった (Figure 2b)。これらのことから、 UCP2 プロモーターにおいては、-586/+186UCP2pro-Luc に含まれる候補配列 D、E が、UCP3 プロモーターにおいて は、-1406/+138UCP3pro-Luc に含まれる候補配列 I、J、K が、STAT5B コンセンサス配列として機能していると推察し た。
4
Figure 2. The effects of active STAT5B proteins on the transcriptional activity of the (a) UCP2 promoter constructs and (b) UCP3 promoter constructs. The squares indicate the predicted STAT5 consensus sequences. The data are represented as relative values that indicate the basal transcriptional activity of each reporter plasmid. RLU, Relative luciferase unit.*, P < 0.05; **, P < 0.01; ***, P < 0.001.
第二節 UCPs プロモーター上の機能的 STAT5B コンセンサス配列の検討 【目的】
前節の結果より、UCPs プロモーター上の STAT5B コンセンサス配列を特定するため、候補 STAT5B コンセンサス 配列を欠失させた UCPs プロモータープラスミドを構築し、レポーターアッセイを行った。
【材料及び方法】
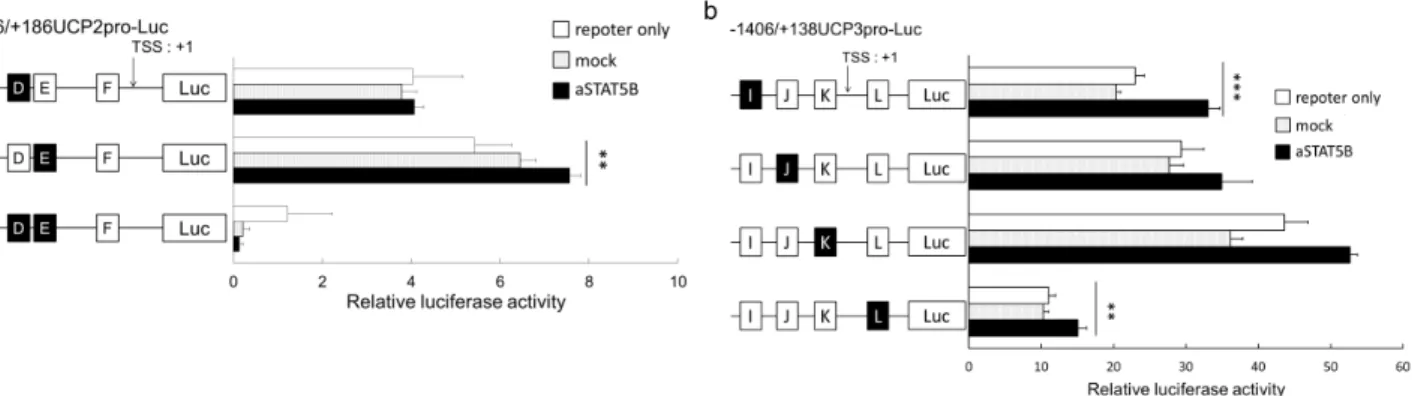
PrimeSTAR Mutagenesis Basal Kit (TakaraBio) を用いて、推定 STAT5B コンセンサス配列を欠失させた UCPs プロ モーターコンストラクトを構築した。レポーターアッセイ及び統計解析は前節と同様の方法で行った。 【結果】 候補 STAT5B コンセンサス配列を欠失させたコンストラクトの転写活性を検討した結果、UCP2 プロモーターにおいて は候補配列 D を欠失させたコンストラクトで、UCP3 プロモーターにおいては候補配列 J、K を欠失させたコンスト ラクトで、それぞれ aSTAT5B 導入によって転写活性が増加しなかった。以上のことから、これらの配列が主 たる STAT5B コンセンサス配列として機能している可能性が示唆された (Figure 3)
5
Figure 3. The effects of active STAT5B proteins on the transcriptional activity of the (a) UCP2 promoter constructs and (b) UCP3 promoter constructs with deleted STAT5B consensus sequences. The white squares indicate existing consensus sequences, while the black squares indicate deleted consensus sequences. **, P < 0.01; ***, P < 0.001.
第三節 推定 STAT5B コンセンサス配列と STAT5B の分子間相互作用の検討 【目的】
UCP2 プロモーター上の機能的 STAT5B コンセンサス配列をプローブとしたゲルシフトアッセイを行い、STAT5B タン パク質との相互作用を検討した。
【材料及び方法】
10 cm dish に播種し一晩培養した TSA201 細胞に、aSTAT5B 10 μg をリン酸カルシウム法で導入し、さらに 24 時間 培養後、細胞溶解液 [50 mM トリス塩酸バッファー、0.15 M 塩化ナトリウム、0.1% SDS、1% Triton X-100、10% コー ル酸ナトリウム、Halt Protease Inhibitor Cocktail, EDTA-Free (Thermo Fisher Scientific)] を用いてタンパク質を抽出した。 タンパク質抽出液は、ビオチンラベルした推定 STAT5B コンセンサス配列オリゴヌクレオチドプローブと共に、LightShift Chemiluminescent EMSA kit (Thermo Fisher Scientific) を用いてゲルシフトアッセイ (Electrophoretic mobility shift assay; EMSA) に供した。 【結果】 UCP2 プロモーター上の推定 STAT5 コンセン サス配列と STAT5B の相互作用について EMSA により検討を行った結果、STAT5B タンパク質量 依存的にシフトバンドのシグナルが強くなった ことから (Figure 4)、UCP2 及び UCP3 プロモータ ー上で STAT5 コンセンサス配列として機能して いる可能性が高いと推察された。
Figure 4. Electrophoretic mobility shift assay (EMSA) was performed using of whole-cell extracts of transfected TSA201 cells. DNA binding of in vitro-translated, active STAT5B was analyzed with biotin-labeled
oligonucleotides of the predicted STAT5 consensus sequence. The protein amounts shown represent consumption from the same extracted sample.
6
第四節 STAT5B 導入による筋細胞における UCP2 及び UCP3 発現の検討 【目的】
筋細胞内で過剰発現させた aSTAT5B が、UCP2 及び UCP3 発現量を増加させるかどうか検討を行った。 【材料及び方法】
ラット筋芽細胞 L6 (JCRB 細胞バンク) は、TSA201 細胞と同様の培地及び培養条件下で継代培養を行った。筋細 胞への分化誘導のため、6 well plate に播種して一晩培養後、分化誘導培地 [DMEM、2%ウマ血清、100 U/ml ペニ シリン G、100 μg/ml ストレプトマイシン硫酸塩、2 mM L-グルタミン含有] に培地交換した。分化誘導培地は 2 日毎に 交換し、10 日間培養して筋細胞に分化させ、実験に供した。Lipofectamine 3000 reagent (Thermo Fisher Scientific) を 用いて aSTAT5B もしくは pCMX (mock) をそれぞれ 2 μg 導入し 48 時間培養後に、RiboZol kit (AMRESCO) を用い て total RNA 及びタンパク質を抽出 した。抽出した total RNA 1 μg を用 いて cDNA に逆転写し、リアルタ イム qPCR に供した。使用した遺 伝子特異的プライマーを Table 1 に示す。 また、抽出したタンパク質は、10%ポリアクリルアミドゲルによる SDS-PAGE によって分離し、PVDF 膜に転写後、ウエ スタンブロッティングに供した。一次抗体として抗 UCP2 ポリクローナル抗体 (1:100、Santa Cruz Biotechnology、 sc-390189) もしくは抗 UCP3 ポリクローナル抗体 (1:500、Bioworld Technology、BS6757) を用いた。二次抗体として HRP 標識抗マウス IgG 抗体 (1:5000、Santa Cruz Biotechnology) で反応させてタンパク質を検出した。同一転写膜に 対してβ-actin の発現量を調べ、ローディングコントロールとした。検出されたバンドから、タンパク質量を ImageJ で解析 した。データは平均値 ± S.D.で表し、統計解析は、SPSS 17.0 software (IBM) を使用し、independent student-t 検定で 群間比較を行った。
【結果】
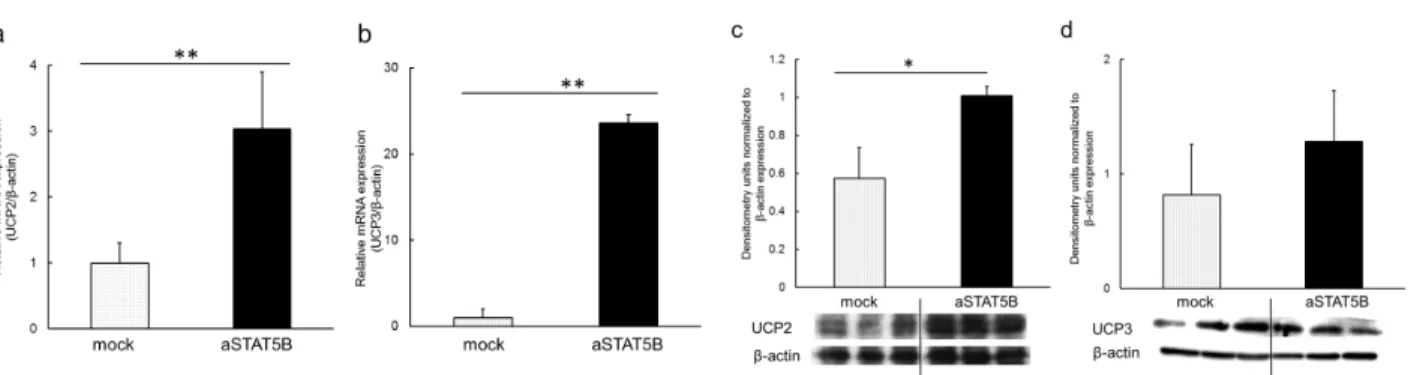
筋細胞への aSTAT5B 導入による UCPs mRNA 発現量は、mock 導入時と比較して有意に増加した (Figure 5a, b: UCP2; 3.6 倍、UCP3; 23.6 倍)。また、UCP3 タンパク質発現量の増加は有意ではなかったが (Figure 5d)、UCP2 タン パク質発現量は mock 導入時と比較して 1.8 倍有意に増加した (Figure 5c)。
Figure 5. UCP2 and UCP3 expression in differentiated L6 cells transfected with pCMX-aSTAT5B. Cells were transfected with 2 μg pCMX (mock) or 2 μg pCMX-aSTAT5B (aSTAT5B). Forty-eight hours later, cells were collected and analyzed for UCP2 and UCP3 mRNA expression by quantitative PCR (a, b) and western blotting (c, d). *, P < 0.05; **, P < 0.01.
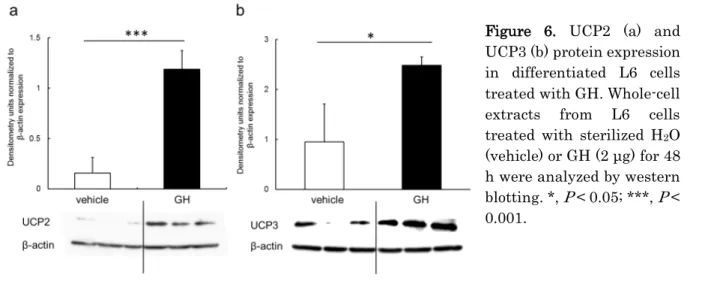
7 第五節 筋細胞に対する GH 刺激による UCP2、UCP3 タンパク質発現の検討 【目的】 前節までの結果より、GH 添加による UCP2 ならびに UCP3 の発現の増加について検討を行った。 【材料及び方法】 GH (成長ホルモン、ヒト組換え体、#071-04591) は、和光純薬工業より購入し、滅菌水に 1 mg/mL の濃度で溶解して 使用した。前節と同様の方法で L6 細胞を 6 well plate で培養し、筋細胞への分化誘導を行った後、GH (2 μg/well) もし くは滅菌水 (vehicle) を添加した。48 時間培養後に前節と同様の方法でタンパク質を抽出した。統計解析は前節と同様 の方法で行った。
【結果】
筋細胞への GH 添加により、vehicle 添加時と比較して UCP2 タンパク質発現量は 2.7 倍、UCP3 タンパク発現量は 2.6 倍増加した (Figure 6)。
Figure 6. UCP2 (a) and UCP3 (b) protein expression in differentiated L6 cells treated with GH. Whole-cell extracts from L6 cells treated with sterilized H2O (vehicle) or GH (2 μg) for 48 h were analyzed by western blotting. *, P < 0.05; ***, P < 0.001.
第一章 考察
本研究において申請者は、GH によるエネルギー代謝作用に特に注目し、これに関与するシグナル伝達機構として、 GHR 結合以降の JAK2/STAT5 経路を介した UCP2 及び UCP3 の発現調整について検討を行った。
UCP は、脂肪酸やグルコースなどの化学的エネルギーを直接熱へと変換させる脱共役タンパク質である。その内 UCP2 は筋組織や脂肪組織を含む全身に、UCP3 は主に骨格筋に発現している。本研究の結果、UCP2 プロモーター上 に既知配列とは異なる配列が、STAT5B コンセンサス配列として機能している可能性が示唆された。さらに骨格筋由来細 胞への GH 添加によって、UCP2 及び UCP3 タンパク質発現がいずれも有意に増加した。STAT5B タンパク質は、筋肉や 肝臓で顕著な発現が認められ、GH をはじめとした様々なサイトカインによって活性化される [18–20]。本研究の結果から、 骨格筋由来細胞内において、STAT5B が UCP 発現を促進する可能性が示唆された。 本研究により GH は、IGF-1 を介した間接的作用ではなく、その直接的作用として UCP 発現量を増大させることが明ら かになった。これは、aGHD 患者に対する GH 投与により、IGF-1 が上昇する前より体脂肪や内臓脂肪が減少し始めると いう臨床現場における経験知と良く一致する [21]。さらに、小児期より GH 治療を受けてきた aGHD 患者以外にも、成人 期になって相対的な GH 分泌不全に陥っている aGHD の潜在患者は多数いると推測されている。しかしながら、GH は高 価な注射製剤である上、GH 下流には細胞周期に関与するシグナル伝達経路が存在することから、臨床応用は慎重に
8 検討する必要がある。STAT5B を特異的に制御する他の薬剤、例として a3-Methylcholanthrene [22] 等をシーズとして開 発することができれば、高価な GH に依らない aGHD への治療の補助として有益であると考えられる。
第二章
転写活性に基づく活性型ビタミン D
3の内分泌・代謝領域における治療効果の予測
序論活性型ビタミン D3 (activated vitamin D3; aVD3) は、核内受容体スーパーファミリークラス 2 に属するビタミン D 受容体 (vitamin D receptor; VDR) の正統なリガンドであり、標的遺伝子の転写制御を行う脂溶性ビタミンである。aVD3の生合成 経路では、まず皮膚におけるプロビタミン D3が紫外線と熱によって VD3に変換される。皮膚で合成された VD3は、血中を 介して肝臓で CYP2R1 水酸化酵素による 25 位の水酸化を受け、25-ヒドロキシビタミン D3 (25(OH)D3) となる [23]。その 後、腎臓で CYP27B1 水酸化酵素による 1α 位の水酸化を受けて aVD3である 1α,25-ジヒロドキシビタミン D3 (1α,25(OH)2D3) となる [24]。現在市販されている VD3製剤は、骨粗鬆症、慢性腎不全、副甲状腺機能低下症、クル病、 骨軟化症など、様々な疾患に臨床使用されている [25–29] 。 aVD3は、骨、腎臓、小腸、肝臓から神経系等、全身に発現する VDR を介して様々な生理作用を発揮する [30]。VDR の領域構造として、タンパク質 N 末端側には、標的遺伝子 DNA 配列に直接結合する DNA 結合ドメイン (DNA binding domain; DBD) が、タンパク質 C 末端側にはリガンド結合ドメイン (ligand binding domain; LBD) が、両者の間にはヒンジ ドメインが存在する。 VDR 遺伝子突然変異がもたらす疾患として、VDDRII が知られている [31]。VDDRII はこれまでに数多くの変異が報 告されており、そのほとんどの症例で、生後数週間の内に重篤なクル病を発症する疾患である [32]。本疾患の内、DBD に変異を有する症例では、VDR への aVD3の結合は認めるものの、DNA への結合性が欠失するため、VDR 標的遺伝子 の転写調節が行われなくなることにより発症する。一方、LBD に変異を有する症例では、VDR への aVD3の結合が不全と なることから、VDR による標的遺伝子の転写活性化が十分達成されないため、同様に VDDRII の症状を呈する。 VDDRII に対する治療法として、カルシトリオールを高濃度で使用することで多くの場合改善が可能である。しかしながら、 変異 VDR (mutation VDR; mVDR) による転写活性の低下の程度は症例により差が大きいため、カルシトリオールの必 要量とその効果は各症例により大きく異なる。 現在我が国の臨床現場で用いられている VD3 製剤は、前述の通りカルシトリオール以外にも 6 種存在するが、各 VD3 製剤間の VDDRII 治療効果に対する比較検討はこれまでなされていない。そこで本研究では、保険適応のあるカルシト リオール及び VD3製剤 6 種を、転写活性を評価指標とし、カルシトリオールを標品として比較検討した。さらに、それらの 結果を踏まえ、より臨床の適応として相応しい疾患とより効果的な VD3製剤の選択について検討した。 第一節 7 種の VD3製剤の野生型 VDR に対する転写活性の検討 【目的】 医療用の適応を有する VD3製剤 7 種について、野生型 VDR に対する転写活性を比較検討する。 【材料及び方法】
VD3製剤として、カルシトリオール (Cayman Chemical)、アルファカルシドール (LKT Laboratories)、カルシポトリオー ル (Toronto research)、エルデカルシトール (中外製薬株式会社)、ファレカルシトリオール (塩野フィネス株式会社) 、タ
9
カルシトール及びマキサカルシトール (Santa Cruz Biotechnology) を使用した。野生型 VDR (wild type VDR ;
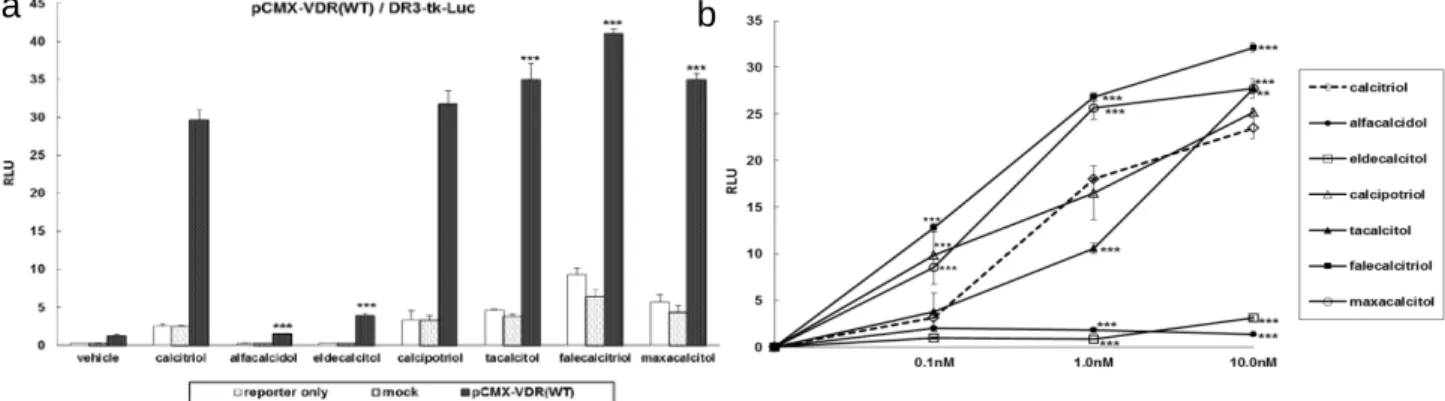
WT-VDR) 発現プラスミド (pCMX-VDR) は、Dr. R. M. Evans より譲渡された [33,34]。VDR 応答配列 (VDR response element; VDRE) レポータープラスミドは、マウスオステオポンチンプロモーター上の VDRE を、pGL3 ベクター (Promega Corporation) にクローニングし作製した。これらのプラスミドを 12 well plate に播種した TSA201 細胞に、VDRE-tk-Luc を 50 ng/well、pCMX-VDR を 20 ng/well、pGL4.70 を 5 ng/well、リン酸カルシウム法で同時導入し、6 時間培養後に VD3 製剤を添加した。十分な薬理作用量で比較検討するために、VD3製剤の添加量は 10.0 nM とした。その後、前章の方法 と同様にレポーターアッセイを行った。 【結果】 各 VD3製剤 10.0 nM で刺激したところ、WT-VDR の転写活性は、タカルシトール、ファレカルシトリオール、マキサカ ルシトール刺激時に、カルシトリオールと比較して有意に高かった (それぞれ 1.2 倍、1.4 倍、1.2 倍、Figure 7a)。一方 で、アルファカルシドール及びエルデカルシトールによる転写活性はカルシトリオールと比較して有意に低かった (Figure 7a)。また、各 VD3製剤による濃度依存的な転写活性について検討したところ、ファレカルシトリオール、マキサカ ルシトールは全ての濃度で同濃度のカルシトリオールよりも転写活性が高かった (Figure 7b)。
Figure 7. The effect of active synthetic VD3 analogs on the transcriptional activity of the WT-VDR. a) The effects of 10.0 nM of VD3 analogs on WT-VDR transcriptional activity. b) The effects of different doses of VD3 analogs on WT-VDR transcriptional activity. RLU, relative luciferase units. **, P < 0.01; ***, P < 0.001 vs. calcitriol. 第二節 VD3製剤が mVDR の転写活性に及ぼす影響 第一項 mVDRs の発現解析と基本転写活性の検討 【目的】 LBD に変異を持つ VDDRII の既報症例に基づき、mVDR 発現プラスミドを構築して各 VD3製剤の転写活性を検討 した。 【材料及び方法】
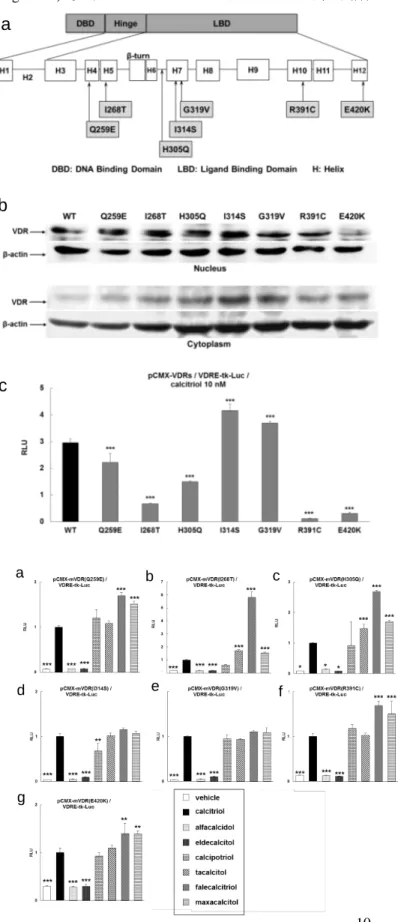
mVDR 発現プラスミド (pCMX-mVDRs) は、pCMX-VDR をテンプレートとして、PrimeSTAR Mutagenesis Basal Kit (TakaraBio) を用いて変異を導入した。変異箇所は Figure 8a に示す通りである。mVDR の発現と細胞内局在を確認す るために、TSA201 細胞に構築した pCMX-mVDRs を導入し、カルシトリオールを 10.0 nM 添加して 24 時間培養した。 Nuclear Extract Kit (Active Motif) を用いて細胞質分画及び核分画を抽出し、ウエスタンブロッティングに供した。一次 抗体は抗 VDR ポリクローナル抗体、二次抗体は HRP 標識抗ウサギ IgG 抗体を用いた。レポーターアッセイ及び統計解 析は、前節と同様の方法で行った。
【結果】
10
mVDR 発現解析の結果、WT-VDR と同様に主として核内に移行していることが確認された (Figure 8b)。カルシトリオ ール 10.0 nM による転写活性を確認したところ、WT-VDR の転写活性と比較して、I314S 及び G319V は有意に転写活 性が高く、その他の mVDRs は有意に転写活性が低かった (Figure 8c)。mVDRs に対する各 VD3製剤の転写活性を比 較検討したところ、ファレカルシトリオールとマキサカルシトールによる転写活性は、I314S (Figure 9d) 及び G319V (Figure 9e) 以外の mVDR でカルシトリオールによる転写活性よりも有意に高かった (Figure 9a, b, c, f, g)。
Figure 8. Introduction of point mutations in the LBD of VDRs and protein expression of mutant receptors. a) Schematic diagram of the VDR and the location of mutations identified in clinical cases. b) Expression of recombinant VDRs in TSA201 cells. Nuclear and
cytoplasmic extracts from transfected TSA201 cells treated with calcitriol (10.0 nM) were analyzed by western blotting using an anti-VDR antibody. c) The effects of 10.0 nM calcitriol on the transcriptional activity of the WT and mutant receptors. RLU, Relative luciferase unit. ***, P < 0.001 vs. WT.
Figure 9a-g. The effects of VD3 analogs on the transcriptional activity of the mutant (m) VDRs. Transcriptional activation by VD3 analogs (10.0 nM) was normalized to that by calcitriol. RLU, Relative luciferase unit. *, P < 0.05; **, P<0.01; ***, P<0.001 vs. calcitriol.
a
b
c
a b c d e f g11 第二項 mVDRs に対するファレカルシトリオール及びマキサカルシトールによる転写活性の検討 【目的】 ファレカルシトリオール及びマキサカルシトールによって、カルシトリオールよりも高い転写活性を示した Q259E、 I268T、H305Q、R391C、E420K の 5 変異について、3 剤に濃度勾配を配し、その転写活性を比較検討した。 【材料及び方法】 細胞培養、レポーターアッセイ及び統計解析は、前節と同様の方法で行った。 【結果】 Q259E、I268T、H305Q、R391C、E420K の 5 変異について、カルシトリオール、ファレカルシトリオール、マキサカル シトールに濃度勾配を配して転写活性を検討した結果、マキサカルシトールによる転写活性は、R391C を除く mVDRs で濃度依存的に増加した (Figure 10)。ファレカル シトリオールによる転写活性は、用いた全ての mVDRs で濃度依存的に増加した。また、1.0 nM 及び 10.0 nM 添加時の転写活性は、全ての mVDRs において、カルシトリオールよりもファレカ ルシトリオールで有意に高かった。
Figure 10a-f. The effects of different doses of falecalcitriol and maxacalcitriol on the transcriptional activity of VDRs. TSA201 cells were co-transfected with the expression plasmids (20 ng) carrying the WT or indicated mVDR, VDRE-tk-Luc (50 ng), and pGL4.70 (5 ng) in the absence or presence of increasing doses of VD3 analogs (0.1 nM – 10.0 nM). RLU, Relative luciferase unit. **, P < 0.01; ***, P < 0.001.
第三節 ファレカルシトリオール及びマキサカルシトールが VDR と転写共役因子との相互作用に及ぼす影響 【目的】
カルシトリオールよりも高い転写活性を有するファレカルシトリオール及びマキサカルシトールについて、VDR と転写 共役因子 (コファクター) との相互作用にどのような影響を及ぼすか、Mammalian two-hybrid assay を用いて検討を行 った。
【材料及び方法】
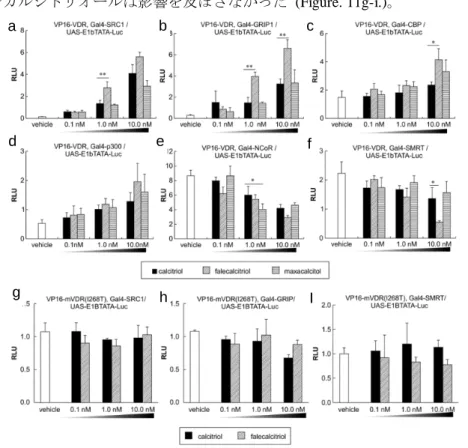
転写活性化に関わる転写共役因子の関与について、Mammalian two-hybrid assay を行い確認した。活性化補助因子 (コアクチベーター) として steroid receptor coactivator-1 (SRC-1)、glucocorticoid receptor interacting protein 1 (GRIP1)、 cAMP response element binding protein-binding protein (CBP)、p300 を用いた。また、抑制補助因子 (コリプレッサー) として nuclear receptor co-repressor 1 (NCoR)、scilencing mediator of retinoic acid and thyroid hormone receptor (SMRT) 用いた。これらを、DNA 結合ドメインである Gal4 DBD との融合タンパク質発現プラスミドとして構築した、 Gal4-SRC-1、Gal4-GRIP1、Gal4-CBP、Gal4-p300、Gal4-NCoR、Gal4-SMRT を使用した [35,36]。また、転写活性化ド メインである VP16 AD と WT-VDR 及び mVDR との融合タンパク質発現プラスミドとして、VP16-VDRs を使用した a b c d e f
12 [33,37]。さらに、Gal4 DBD に対する応答配列レポータープラスミドとして、UAS-E1b-TATA-Luc を使用した [36]。レポ ーターアッセイ及び統計解析は、前節までと同様の方法で行った。 【結果】 ファレカルシトリオールとマキサカルシトールは、コアクチベーターである SRC1、GRIP1、CBP1、p300 のリクルートを濃度依存的に促進し (Figure 11a-d)、さらにファレカルシトリオールは、コリプレッサーであ る NCoR、SMRT の解離を促進した (Figure 11e,f)。また、ファレカルシトリオールはカルシトリオールよりも 有意に SRC1 (Figure 11a、1.0 nM)、GRIP1 (Figure 11b、10.0 nM)、CBP (Figure 11c、10.0 nM) のリクルートを促 進し、SMRT (Figure 11f、10.0 nM) の解離を促進した。一方で、I268T とコファクターとの相互作用に、ファ レカルシトリオールは影響を及ぼさなかった (Figure. 11g-i.)。
Figure 11a-i. Interactions of cofactors with the wild-type and mutant VDRs in the
mammalian two-hybrid assay. (a–f) TSA201 cells were co-transfected with the
VP16-VDR expression plasmid , UAS-E1b-TATA-Luc, and Gal4-cofactors in the absence or presence of VD3 analogs. (g–i) TSA201 cells were
co-transfected with the
VP16-mVDR (I268T) expression plasmid, UAS-E1b-TATA-Luc, and Gal4-cofactors in the absence or presence of VD3 analogs. RLU, Relative
luciferase unit. *, P < 0.05; **, P < 0.01. 第四節 VDR 標的遺伝子の発現調節に対する VD3製剤の影響 【目的】 前節までの結果から、転写活性が高いと推察されたファレカルシトリオールが、VDR 標的遺伝子のプロモーター活 性と mRNA 発現に及ぼす影響について検討を行った。 【材料及び方法】 マウス骨芽細胞前駆細胞 MC3T3-E1 (理研バイオリソースセンター) は、TSA201 細胞と同様の培地、培養条件で継 代培養を行った。6 well plate に播種後、骨芽細胞分化試薬 (TakaraBio) を添加した培地を用いて 10 日間培養し、骨 芽細胞への分化誘導を行った。VDR 標的遺伝子に対するプロモーター活性を検討するために、aVD3によって発現が 抑制される CYP27B1 遺伝子のプロモーターをルシフェラーゼ発現ベクターにクローニングした CYP27B1promoter-Luc を用いた。前節までと同様の方法で、pCMX-VDR 及び pGL4.70 と共に TSA201 細胞に導入し、 レポーターアッセイを実施した。 a b c d e f g h I
13
また、TSA201 細胞及び骨芽細胞に分化した MC3T3-E1 細胞に、vehicle、カルシトリオール 10.0 nM、ファレカルシト リオール 10.0 nM を添加し 48 時間培養後、前節までと同様の方法で total RNA を抽出し、リアルタイム qPCR を行った。 発現解析遺伝子として、cytochrome P450 family 24 subfamily A member 1 (CYP24A1)、alkaline phosphatase (ALP)、 osteopontin (OPN)、collagen type I
(COL1)、receptor activator of nuclear factor kappa-B ligand (RANKL) を用 い、内在性コントロール遺伝子として、 β-actin を用いた。使用した遺伝子特 異的プライマーを Table 2 に示す。 【結果】 CYP27B1 プロモーターを用いたレポーターアッセイの結果、WT-VDR において、カルシトリオール及びファレカルシ トリオール濃度依存的に転写活性が低下したが、2 剤間の差は認められなかった。一方、mVDR (I268T) において、 10.0 nM 刺激時、ファレカルシトリオールはカルシトリオールよりも有意に CYP27B1 プロモーター活性を抑制した。 リアルタイム qPCR により VDR 標的遺伝子 Mrna の発現を検討した結果、ファレカルシトリオール刺激によって、VDR の標的遺伝子である CYP24A1、ALP、 OPN、COL1、RANKL の mRNA 発現 は、カルシトリオールと比較して、それ ぞれ、2.3 倍、2.6 倍、1.5 倍、1.9 倍、 3.5 倍と有意に高かった (Figure 12)。
Figure 12. Transcriptional control of the VDR target genes by calcitriol and falecalcitriol. (a, b) Functional analysis of calcitriol and falecalcitriol in the CYP27B1 promoter activation assay. *, P < 0.05 vs. calcitriol. +, P < 0.05. (c) CYP24A1 mRNA expression in TSA201 cells and ALP, OPN, COL1, and RANKL in MC3T3-E1 cells stimulated by calcitriol and falecalcitriol. RLU, Relative luciferase unit. **, P < 0.01; ***, P < 0.001 vs. calcitriol. 第二章 考察 本章では、VDR の外因性リガンドとして臨床で使用されている全 7 種の VD3製剤を対象とし、WT-VDR を介した転 写活性を評価指標として比較した。次いで、この効果を実際の臨床に直接還元することを目的として VDDRII に注目し、 この疾患をもつ患者に関する症例報告に基づいて、転写活性を指標として治療効果を推測した。 本研究の結果、カルシポトリオール、タカルシトール、ファレカルシトリオール、マキサカルシトールについては、 WT-VDR に対して生理的な aVD3であるカルシトリオールと同等か、もしくはそれ以上の転写活性を認めた。一方でア a b c
14
ルファカルシドールとエルデカルシトールの 2 製剤は、WT-VDR に対する転写活性が非常に弱かった。理由として、ア ルファカルシドールはプロドラッグであり、肝臓での水酸化を経て活性型となることから [38]、本研究で使用したヒト胎 児腎細胞由来 TSA201 内では活性化されないためと考えられた。またエルデカルシトールは、VDR に対する結合親和 性がカルシトリオールと比較して約 1/2 であるため [39]、転写活性が弱かったと推察された。
VDDRII の既報では、LBD 領域に変異を有する VDDRII の症例の内 I314S は薬理作用量の VD2で、Q259E、 H305Q、G319V、R391C は高濃度のカルシトリオールで治療可能であったとされる [40–42]。一方で I268T と E420K は、 高濃度のカルシトリオール投与でも症状が改善されなかったことが報告されている [43,44]。本研究で再現した mVDR 発現プラスミドによるタンパク質発現量をウエスタンブロッティングで解析したところ、全ての mVDR が核内に移行するこ とを確認した。 前述の通り、Q259E、H305Q、R391C の 3 症例では、高濃度のカルシトリオール投与によって症状が改善されたと報 告されている。本研究においても、カルシトリオールでは、高濃度の刺激においてのみ、未刺激と比較して有意に転写 量が増加した。このことから、カルシトリオールの添加量増加により転写活性が促進されることが、in vitro でも再現され た。I268T は高濃度のカルシトリオール投与でも治療効果が無いと報告されている [43]。本研究における結果でも、 I268T における高濃度カルシトリオールによる転写活性は、未刺激と比較して 4.2 倍と、前述の Q259E、H305Q、 R391C の 3 変異と比較して、転写量があまり増加しなかった。一方で、ファレカルシトリオール 10.0 nM 刺激による I268T の転写活性は、カルシトリオール 10.0 nM 刺激に対して 37.4 倍と有意に高かった。これらの結果から、ファレカルシトリ オールはカルシトリオールとは異なる受容体結合性を有しているために、高い転写活性を有する可能性が推察された。 ファレカルシトリオールの詳細な作用メカニズムを解明するために、mammalian two-hybrid assay による WT-VDR とコフ ァクターの相互作用を解析した。その結果、いずれの薬剤についても濃度依存的にコアクチベーターのリクルートが促 進されたが、GRIP1 と SRC1 においては、ファレカルシトリオールのリクルート促進作用はカルシトリオールと比較して有 意に高かった。また、カルシトリオール及びファレカルシトリオール濃度依存的にコリプレッサーの解離が促進され、そ の作用はファレカルシトリオールでより強かった。以上のことからファレカルシトリオールは、コアクチベーターのリクルー ト、及びコリプレッサーの解離を促進することで、転写活性促進に寄与していることが示唆された。I268T は、コアクチベ ーターである SRC1 と DRIP205 のリクルートが阻害されていることが報告されているが [43]、今回の結果でも、ファレカ ルシトリオールは I268T とコファクターのリクルートに影響を及ぼさなかった。このことからファレカルシトリオールは、 WT-VDR ではコファクターとの相互作用に関与して VDR の転写活性に影響する一方で、I268T に関しては、コファク ターとの相互作用には関与せず、他のメカニズムによって転写活性の増加をもたらしている可能性が推察された。 VDR を介したファレカルシトリオールの標的遺伝子転写制御について、VDR の標的遺伝子である CYP27B1、 CYP24A1、ALP、OPN、COL1 及び RANKL 遺伝子を用いて、カルシトリオールによる発現制御との比較検討を行った。 その結果、ファレカルシトリオールはカルシトリオールよりもそれぞれの遺伝子発現量を有意に増加させた。ファレカル シトリオールの VDR との結合親和性はカルシトリオールの 1/3 だが [45]、骨形成作用がカルシトリオールよりも強いと いう報告があることから [46]、ファレカルシトリオールは VDR を介しながらも、転写に関与する他の要因も関わっている 可能性が示唆されている [47]。 以上、本研究により、ファレカルシトリオールは WT-VDR に対してカルシトリオールよりも高い転写活性を有する可能 性が示唆された。ファレカルシトリオールは VDR に対して、カルシトリオールとは異なる結合様式で結合し、何らかの因 子を介して機能していることが推察された。本研究では、既存医薬品と実際の症例の治療効果の予測を、in vitro の実 験系における転写活性の比較検討によって試みた。本研究成果により、実際の臨床症例において適切な薬物治療を 実施することで、救済可能な VDDRII 症例が存在する可能性が示唆された。
15
第三章
転写活性に基づく生薬由来成分 cinnamaldehyde の内分泌・代謝領域における薬効の予測
序論 第二章の活性型ビタミン D3のように、外因性リガンドも内因性リガンドと同様に、内分泌・代謝領域において様々な 生理作用を発揮、あるいは修飾していることは明らかである。本章では、アジアで古来より、ハーブや生薬の構成成分 として用いられてきた桂皮 (Cinnamomum cassia) による抗糖尿病効果について検討を行った。 桂皮は、ヒトにおいて空腹時血糖と血中脂質を減少させることが報告されている [48]。桂皮に含まれる芳香成分の 一つである cinnamaldehyde (CA) を、肥満 2 型糖尿病モデル db/db マウスに投与した結果、空腹時血糖や血中インス リン値の改善、体重の減少と、HDL-コレステロール値の上昇が見られた [49]。これらの証左から、桂皮の抗糖尿病効 果や、CA がもつ脂質代謝に及ぼす効果は、核内受容体の一種である peroxisome proliferator-activated receptors (PPARs) を介した脂質・糖代謝作用である可能性があると示唆された。本章では、PPARs の外因性リガンドとしての CA の機能について、転写活性の視点から評価・解析を試みた。 第一節 CA が PPAR の転写活性に及ぼす影響とその分子メカニズムの検討 【目的】 CA が PPARs の転写活性に及ぼす影響についてスクリーニングし、さらに PPAR のリガンドとして作用する際の結合 ドメインへの CA の結合性について、LBD 領域に変異を導入した PPARγ (mPPARγ) を用いて検討した。 【材料及び方法】 CA (#031-03453) は和光純薬工業より購入した。PPARs の陽性コントロールである GW590735 (#622402-22-6) は Cayman chemical、L-165041 (#076K4605) は Sigma、pioglitazone (#ALX-270-367) は Enzo Life Sciences よりそれぞ れ購入した。これらの薬剤は全て DMSO に 50 mM の濃度で溶解し実験に供した。CA は、細胞の生存及び増殖に影 響のないことを確認した 50 μM を上限として、刺激濃度を設定した。PPARα、δ、γ 発現プラスミドである pCMX-PPARs、 変異 PPARγ (mutation PPARγ; mPPARγ) 発現プラスミドである pCMX-mPPARγ、PPARs の LBD と Gal4 を接続した Gal4-PPARs は、Dr. Evans RM (Salk Institute) より譲渡されたものを、PPAR 応答配列 (PPAR response element; PPRE) をもつレポータープラスミド PPRE-tk-Luc は、Dr. J. Szwaya (Dana Farber Cancer Institute) より譲渡されたものを 使用した [50]。Gal4 応答配列レポータープラスミド UAS-E1b-TATA-Luc は、第二章第二節で使用したものを用いた。 レポーターアッセイ及び統計解析は、前節までと同様の方法で行った。【結果】
CA は、濃度依存的に、PPARδ、PPARγ の転写活性を増加させたが (Figure 13a,b)、PPARα の転写活性には影響を 及ぼさなかった (data not shown)。さらに、CA によって Gal4-PPARδ、γ の転写活性も濃度依存的に増加したことから (Figure 14a,b)、CA は PPAR の LBD に直接結合することによりその転写を活性化していることが確認された。また、 S342A 変異に対しては、ピオグリタゾン刺激は転写活性が増加しなかったのに対し、CA 刺激により未刺激と比較して 有意に転写活性が増加した (Figure 15、S342A; 2.8 倍 vs. vehicle) 。
16
Figure 13. The stimulatory effects of CA on the transcriptional activity of PPARs. Either a) pCMX-PPARδ or b) pCMX-PPARγ were cotransfected into TSA201 cells with the reporter gene, PPRE-tk-Luc, in the presence of CA with increasing doses. **, P < 0.01.
Figure 14. The stimulatory effects of CA on the gene transcription mediated by the PPARs-LBD. Either a) Gal4-PPARδ or b) Gal4-PPARγ was cotransfected into TSA201 cells with the reporter gene, UAS-E1b-TATA-Luc, in the presence of CA with increasing doses. **, P < 0.01.
Figure 15. Effects of CA on the function of PPARγ mutants. PPARγ expression plasmid (pCMX-mPPARγ) for the indicated mutants and reporter were transiently cotransfected into TSA201 cells together in the absence or presence of various ligands. *,P < 0.05; **, P < 0.01.
a
b
17 第二節 CA と外因性リガンドが PPARs の転写活性に及ぼす相加相乗効果の検討 【目的】 前節において、CA は陽性コントロールと異なる接合性を持つことが示唆された。そこで本節では、PPARs における CA と陽性コントロールと相加相乗効果の有無について検討を行った。 【材料及び方法】 細胞培養及びレポーターアッセイは、前節までと同様の方法で行った。 【結果】
PPARγ において、CA と陽性コントロールとの相加相乗効果は確認されなかった (data not shown)。一方で、PPARδ に対して、陽性コントロールである L-165041 単独刺激と比較して、CA を同時添加した時の転写活性は、1.3 倍 (CA 10 μM)、2.1 倍 (CA 30 μM)、4.7 倍 (CA 50 μM) に増加した (Figure 16)。
Figure 16. Synergistic effect of CA and L-165041 on PPARδ activation. TSA201 cells, which were cotransfected with pCMX-PPARδ (50 ng), PPRE-tk-luc (100 ng) and treated with CA (10–50 μM) and low concentration of L-165041 (10 μM) for 24 h. 第三節 CA による RXRα 活性化作用の検討 【目的】 本節では、PPAR のヘテロダイマーパートナーである RXRα に対する CA の活性化作用についても検討を行った。さ らに、コアクチベーターである SRC1 などの転写共役因子との相互作用に及ぼす影響についても検討を行った。 【材料及び方法】 RXRα 発現プラスミドである pCMX-RXRα は Dr. Evans RM より譲渡されたものを、RXRα 応答配列レポータープラス ミドである direct repeat 1; DR1-tk-Luc は Dr. J. Szwaya より譲渡されたものを使用した。mammalian two-hybrid assay を 行うために、DNA 結合ドメインである Gal4 DBD との融合タンパク質発現プラスミドとして構築した、Gal4-RXRα、 Gal4-SRC1 を使用した。また、リガンド依存性を確認するために VP16 AD との融合タンパク質発現プラスミドとして、 VP16-RXRα 及び VP16-SRC1 を使用した。RXRα の陽性コントロールとして、9-cis-レチノイン酸 (9-cis-RA、LKT Laboratories、#R1777) を用いた。 【結果】 RXRα は、PPAR を含む数種類の核内受容体とヘテロダイマーを形成し、DNA 上の応答配列に結合することで転写 を活性化することが知られているが、一方で、9-cis-RA をリガンドとしてホモダイマーを形成し、同様に応答配列に結合 することがわかっている [51,52]。本実験の結果から、CA 50µM によっても RXRα はホモダイマーを形成し、転写を活 性化すると示唆された (Figure 17a)。さらに CA は、RXRα のコアクチベーターである SRC1 との相互作用についても、 リガンド依存的に促進すると示唆された (Figure 17b, c)。
18
Figure. 17 a) CA activates RXR (DR1-tk). Both RXRα (50 ng) and DR1-tk-Luc reporter (100 ng) were cotransfected into TSA201 cells, followed by 9-cis-RA or CA. b, c) In a reciprocal manner, Gal4-RXR expression plasmids (50 ng) were cotransfected into TSA-201 cells together with the Gal4-responsive reporter gene, UAS-E1b-TATA-luc (100 ng), and 50 ng of VP16-SRC1 in the escalation dose of CA (c). These results represent at least three transfections performed in triplicate. **, p < 0.01.
第四節 CA による PPARγ 下流遺伝子の転写調節
【目的】
CA による PPARγ の標的遺伝子発現に対する作用について検討を行った。標的遺伝子には、脂肪細胞特異的脂肪 酸結合タンパク質で、分化した脂肪細胞に発現する adipocyte protein 2 (aP2) と、脂肪酸輸送タンパク質の一種で、遊 離脂肪酸を骨格筋や幹細胞に輸送する働きをもつ cluster of differentiation 36 (CD36) を用いた [53]。
【材料及び方法】
マウス脂肪前駆細胞 3T3-L1 は TSA201 細胞と同様の培地、条件下で継代培養した。6 well plate に播種してコンフ ルエントに達した後、さらに 2 日間培養し、脂肪細胞分化誘導 high-glucose DMEM [10% FBS、1 µM dexamethasone、 500 µM isobutylmethlxanthine、5 µg/ml insulin 含有] で 3 日間培養し、脂肪細胞への分化誘導を行った。その後 CA 50 μM を添加して 24 時間培養し、total RNA を抽出後、リアルタイム qPCR に供した。使用した遺伝子特異的プライマ ーを Table 3 に示す。
【結果】
CA 刺激により vehicle と比較して aP2 は 1.5 倍、CD36 は 1.3 倍と、それぞれ有意に mRNA 発現が増加した (Figure 18)。
a
b
19 Figure 18. CA mildly induced the expression of direct target genes of PPARδ in differentiated 3T3-L1 adipocytes. After 8 days of differentiation, 3T3-L1 adipocytes were treated with vehicle (DMSO) and CA (50 μM) for 48 h. **: P < 0.01.
第三章 考察
本章では桂皮の主成分の一つである、CA の抗糖尿病作用に着目した。先行研究から、db/db マウスに対する CA の 4 週間継続投与による血糖値の低下効果が確認されている [49]。また、db/db マウスに対する桂皮抽出物の投与により、 PPAR を介して糖、脂質代謝が調節されているとの報告がある [54]。これらのことから本研究では、CA の標的分子とし て PPAR に着目し、CA の転写活性について評価、検討を行った。PPAR は脂質、糖質等の細胞内代謝と細胞の分化 に関与する転写因子群で、α、δ、γ の 3 つのサブタイプが知られているが、それぞれ発現する部位や機能、特異的リガ ンドが異なる。いずれのサブタイプも、リガンド存在下で RXRα とヘテロダイマーを形成し、標的遺伝子プロモーター上 の PPRE に結合することでその発現を調整している。PPARδ は全身に幅広く発現しており、脂質やリポタンパク質代謝 に寄与し、肥満モデルマウスにおいては耐糖能やインスリン感受性を改善することがわかっている [55]。PPARγ は、脂 肪細胞に多く発現し、脂肪細胞分化に対して中心的役割を果たしている [56]。また、2 型糖尿病の標的分子として、治 療薬であるチアゾリジン系薬剤が結合する核内受容体であり、且つインスリン感受性を改善する [57]。先行研究にお ける db/db マウスへの CA 投与の結果、インスリン抵抗性指標である homeostasis model assessment as an index of insulin resistance; HOMA-IR の改善やそれに伴う血中インスリン量低下、HDL-コレステロールの増加等、肥満や 2 型 糖尿病の改善効果が認められたが、その作用メカニズムは未解明のままであった。本研究により、CA は PPARδ/γ、さら に RXRα の転写活性を促進することで効果を発揮していることが示唆された。 CA は官能基を有する小分子である。一方 PPARs は、LBD 分子構造として大きなリガンド結合ポケットを有するため、 様々なリガンドが結合できる分子であることが明らかになっている [58]。本研究において、LBD に変異を導入した PPARγ (S342A)では、陽性コントロールであるピオグリタゾンでは転写が活性化されなかった一方で、CA により活性化 された。さらに PPARδ の活性化に対し、CA と陽性コントロールである L-165041 は相乗効果を有していた。これらの結 果から、各リガンドが異なる様式で PPARs に結合し、その転写に関与している可能性が示唆された。
また、CA のエネルギー代謝に及ぼすアウトプットとして、PPARγ によって発現調節を受けることがわかっている aP2 や CD36 に注目して解析した [53]。脂肪細胞において、これらの mRNA 発現が CA によって促進されたことから、CA は PPARγ を介して脂質代謝改善作用やインスリン抵抗性を改善する作用があることが示唆された。
以上の結果から CA は、PPARs の天然由来外因性リガンドとして PPAR の各アイソフォームの転写活性を促進するこ とで、抗肥満、抗糖尿病作用を発揮している可能性が示唆された。
20
終章
本研究において申請者は、内分泌・代謝領域の用途をもつ医療用医薬品や、医薬品としても使用される内因性リガ ンド、天然由来の外因性リガンドを対象として検討を行った。それぞれの適応は異なるものの、シグナル伝達を介して その下流に位置する遺伝子の転写に影響を及ぼすという点で共通している。このことから、各リガンドについて転写活 性を指標とした薬理効果の予測が可能であるか、検討を試みた。その結果、以下の結論を得た。 1) 内分泌領域の臨床で汎用される GH は、直接的作用として転写因子 STAT5B を介して、UCP2、UCP3 の転写を 促進することが明らかになった。さらに、UCP2 プロモーター上で新規の STAT5s のコンセンサス配列が機能して いることを示唆する所見を得た。GH による直接的作用の存在はこれまでにも示唆されていたが、転写因子 STAT5B を介して標的遺伝子 UCP2、UCP3 に影響し、エネルギー代謝促進に寄与していることを示した。 2) 臨床上の適応を有している VD3製剤 7 種を用いて、VDDRII の改善効果について転写活性を評価指標とした 比較検討を試みた。その結果、各 VD3製剤により VDR を介した転写量が異なっていたことから、VDDRII に対 する改善効果も各 VD3製剤によって異なる可能性を示した。中でもファレカルシトリオールは、カルシトリオール よりも VDR を介した転写活性が高く、症例によっては VDDRII の臨床症状の改善効果が高い可能性を示した。 3) 抗糖尿病作用を有する生薬は多数知られている。今回、その内の一つ、桂皮に含まれる主要な物質である CA を用いて、脂質、糖質代謝改善作用について評価を試みた。その結果、CA は PPARδ/γ や RXRα を介して、そ の下流にある標的遺伝子の発現を促進することを示した。このことから構成生薬由来の外因性リガンドも、転写 活性によってその作用の予測が可能であることを示した。 以上のように、内分泌・代謝領域に関連するホルモン・リガンド類は、受容体を介して転写因子、転写共役因子に影 響を及ぼし、標的遺伝子の転写を制御する。本研究において申請者は、様々なホルモンやリガンドの転写活性を解析 した。これにより、内分泌・代謝領域の生理活性物質やホルモンの医薬品としての作用を、シグナル伝達に関わる転写 因子や標的遺伝子の活性化を用いて評価出来ることを示した。 引用文献[1] W.H. Daughaday, Endocrinology--the way we were: a personal history of somatomedin., Growth Horm. IGF Res. 16 Suppl A (2006) S3–5. doi:10.1016/j.ghir.2006.03.005.
[2] N. Møller, J. Gjedsted, L. Gormsen, J. Fuglsang, C. Djurhuus, Effects of growth hormone on lipid metabolism in humans., Growth Horm. IGF Res. 13 Suppl A (2003) S18–21.
[3] N. Møller, L.C. Gormsen, O. Schmitz, S. Lund, J.O.L. Jørgensen, N. Jessen, Free fatty acids inhibit growth hormone/signal transducer and activator of transcription-5 signaling in human muscle: a potential feedback mechanism., J. Clin. Endocrinol. Metab. 94 (2009) 2204–7. doi:10.1210/jc.2008-2624.
[4] C. Carter-Su, J. Schwartz, L.S. Smit, Molecular mechanism of growth hormone action., Annu. Rev. Physiol. 58 (1996) 187–207. doi:10.1146/annurev.ph.58.030196.001155.
21
[5] M. Azam, H. Erdjument-Bromage, B.L. Kreider, M. Xia, F. Quelle, R. Basu, et al., Interleukin-3 signals through multiple isoforms of Stat5., EMBO J. 14 (1995) 1402–11.
[6] A.L. Mui, H. Wakao, A.M. O’Farrell, N. Harada, A. Miyajima, Interleukin-3, granulocyte-macrophage colony stimulating factor and interleukin-5 transduce signals through two STAT5 homologs., EMBO J. 14 (1995) 1166–75.
[7] X. Liu, G.W. Robinson, K.U. Wagner, L. Garrett, A. Wynshaw-Boris, L. Hennighausen, Stat5a is mandatory for adult mammary gland development and lactogenesis., Genes Dev. 11 (1997) 179–86.
[8] S. Teglund, C. McKay, E. Schuetz, J.M. van Deursen, D. Stravopodis, D. Wang, et al., Stat5a and Stat5b proteins have essential and nonessential, or redundant, roles in cytokine responses., Cell. 93 (1998) 841–50. [9] G.B. Udy, R.P. Towers, R.G. Snell, R.J. Wilkins, S.H. Park, P.A. Ram, et al., Requirement of STAT5b for
sexual dimorphism of body growth rates and liver gene expression., Proc. Natl. Acad. Sci. U. S. A. 94 (1997) 7239–44.
[10] M. Kawai, N. Namba, S. Mushiake, Y. Etani, R. Nishimura, M. Makishima, et al., Growth hormone
stimulates adipogenesis of 3T3-L1 cells through activation of the Stat5A/5B-PPARgamma pathway., J. Mol. Endocrinol. 38 (2007) 19–34. doi:10.1677/jme.1.02154.
[11] J.T. Zhao, M.J. Cowley, P. Lee, V. Birzniece, W. Kaplan, K.K.Y. Ho, Identification of novel GH-regulated pathway of lipid metabolism in adipose tissue: a gene expression study in hypopituitary men., J. Clin. Endocrinol. Metab. 96 (2011) E1188–96. doi:10.1210/jc.2010-2679.
[12] A.F. Attanasio, S.W. Lamberts, A.M. Matranga, M.A. Birkett, P.C. Bates, N.K. Valk, et al., Adult growth hormone (GH)-deficient patients demonstrate heterogeneity between childhood onset and adult onset before and during human GH treatment. Adult Growth Hormone Deficiency Study Group., J. Clin. Endocrinol. Metab. 82 (1997) 82–8. doi:10.1210/jcem.82.1.3643.
[13] P. V Carroll, E.R. Christ, B.A. Bengtsson, L. Carlsson, J.S. Christiansen, D. Clemmons, et al., Growth hormone deficiency in adulthood and the effects of growth hormone replacement: a review. Growth Hormone Research Society Scientific Committee., J. Clin. Endocrinol. Metab. 83 (1998) 382–95.
doi:10.1210/jcem.83.2.4594.
[14] G. McGauley, R. Cuneo, F. Salomon, P.H. Sönksen, Growth hormone deficiency and quality of life., Horm. Res. 45 (1996) 34–7.
[15] C. Hioki, T. Yoshida, A. Kogure, Y. Takakura, T. Umekawa, K. Yoshioka, et al., Effects of growth hormone (GH) on mRNA levels of uncoupling proteins 1, 2, and 3 in brown and white adipose tissues and skeletal muscle in obese mice., Horm. Metab. Res. 36 (2004) 607–13. doi:10.1055/s-2004-825905.
[16] M. Onishi, T. Nosaka, K. Misawa, A.L. Mui, D. Gorman, M. McMahon, et al., Identification and
characterization of a constitutively active STAT5 mutant that promotes cell proliferation., Mol. Cell. Biol. 18 (1998) 3871–9.
[17] T. Tagami, Y. Park, J.L. Jameson, Mechanisms that mediate negative regulation of the thyroid-stimulating hormone alpha gene by the thyroid hormone receptor., J. Biol. Chem. 274 (1999) 22345–53.
22
[18] J.E. Darnell, Reflections on STAT3, STAT5, and STAT6 as fat STATs., Proc. Natl. Acad. Sci. U. S. A. 93 (1996) 6221–4.
[19] J.N. Ihle, The Stat family in cytokine signaling., Curr. Opin. Cell Biol. 13 (2001) 211–7.
[20] W.J. Leonard, Role of Jak kinases and STATs in cytokine signal transduction., Int. J. Hematol. 73 (2001) 271– 7.
[21] I. Fukuda, N. Hizuka, T. Muraoka, A. Ichihara, Adult growth hormone deficiency: current concepts., Neurol. Med. Chir. (Tokyo). 54 (2014) 599–605.
[22] C. Lee, X. Ding, D.S. Riddick, The role of cytochrome P450-dependent metabolism in the regulation of mouse hepatic growth hormone signaling components and target genes by 3-methylcholanthrene., Drug Metab. Dispos. 41 (2013) 457–65. doi:10.1124/dmd.112.048835.
[23] J.B. Cheng, M.A. Levine, N.H. Bell, D.J. Mangelsdorf, D.W. Russell, Genetic evidence that the human CYP2R1 enzyme is a key vitamin D 25-hydroxylase., Proc. Natl. Acad. Sci. U. S. A. 101 (2004) 7711–5. doi:10.1073/pnas.0402490101.
[24] K. Takeyama, S. Kitanaka, T. Sato, M. Kobori, J. Yanagisawa, S. Kato, 25-Hydroxyvitamin D3 1alpha-hydroxylase and vitamin D synthesis., Science. 277 (1997) 1827–30.
doi:10.1126/science.277.5333.1827.
[25] A.J. Brown, D.W. Coyne, Vitamin D analogs: new therapeutic agents for secondary hyperparathyroidism., Treat. Endocrinol. 1 (2002) 313–27. doi:10.2165/00024677-200201050-00004.
[26] Y. Tsukamoto, [Vitamin D therapy in renal osteodystrophy]., Clin. Calcium. 14 (2004) 21–6. doi:CliCa040913211326.
[27] J.L. O’Neill, S.R. Feldman, Vitamine D analogue-based therapies for psoriasis., Drugs Today (Barc). 46 (2010) 351–60. doi:10.1358/dot.2010.46.5.1473264.
[28] H. Morii, T. Inoue, T. Nishijima, T. Tomokuni, T. Ishikawa, K. Moriya, et al., Management of calcium and bone abnormalities in hemodialysis patients., Semin. Nephrol. 24 (2004) 446–8.
doi:10.1016/j.semnephrol.2004.06.016.
[29] K. Matsumoto, K. Hashimoto, M. Kiyoki, M. Yamamoto, K. Yoshikawa, Effect of 1,24R-dihydroxyvitamin D3 on the growth of human keratinocytes., J. Dermatol. 17 (1990) 97–103.
doi:10.1111/j.1346-8138.1990.tb03714.x.
[30] M.R. Haussler, G.K. Whitfield, C.A. Haussler, J.C. Hsieh, P.D. Thompson, S.H. Selznick, et al., The nuclear vitamin D receptor: biological and molecular regulatory properties revealed., J. Bone Miner. Res. 13 (1998) 325–49. doi:10.1359/jbmr.1998.13.3.325.
[31] C.R. Scriver, T.M. Reade, H.F. DeLuca, A.J. Hamstra, Serum 1,25-dihydroxyvitamin D levels in normal subjects and in patients with hereditary rickets or bone disease., N. Engl. J. Med. 299 (1978) 976–9. doi:10.1056/NEJM197811022991803.
[32] P.J. Malloy, J.W. Pike, D. Feldman, The Vitamin D Receptor and the Syndrome of Hereditary
23
[33] K. Umesono, K.K. Murakami, C.C. Thompson, R.M. Evans, Direct repeats as selective response elements for the thyroid hormone, retinoic acid, and vitamin D3 receptors., Cell. 65 (1991) 1255–66.
doi:10.1016/0092-8674(91)90020-Y.
[34] J.D. Chen, K. Umesono, R.M. Evans, SMRT isoforms mediate repression and anti-repression of nuclear receptor heterodimers., Proc. Natl. Acad. Sci. U. S. A. 93 (1996) 7567–71. doi:10.1073/pnas.93.15.7567. [35] T. Tagami, P. Kopp, W. Johnson, O.K. Arseven, J.L. Jameson, The thyroid hormone receptor variant alpha2 is
a weak antagonist because it is deficient in interactions with nuclear receptor corepressors., Endocrinology. 139 (1998) 2535–44. doi:10.1210/endo.139.5.6011.
[36] T. Tagami, W.H. Lutz, R. Kumar, J.L. Jameson, The interaction of the vitamin D receptor with nuclear receptor corepressors and coactivators., Biochem. Biophys. Res. Commun. 253 (1998) 358–63.
doi:10.1006/bbrc.1998.9799.
[37] T. Suda, N. Takahashi, N. Udagawa, E. Jimi, M.T. Gillespie, T.J. Martin, Modulation of osteoclast
differentiation and function by the new members of the tumor necrosis factor receptor and ligand families., Endocr. Rev. 20 (1999) 345–57. doi:10.1210/edrv.20.3.0367.
[38] H. Orimo, E. Schacht, The D-hormone analog alfacalcidol: the pioneer beyond the horizon of osteoporosis treatment., J. Rheumatol. Suppl. 76 (2005) 4–10.
[39] S. Hatakeyama, M. Yoshino, K. Eto, K. Takahashi, J. Ishihara, Y. Ono, et al., Synthesis and preliminary biological evaluation of 20-epi-eldecalcitol [20-epi-1alpha,25-dihydroxy-2beta-(3-hydroxypropoxy)vitamin D3: 20-epi-ED-71]., J. Steroid Biochem. Mol. Biol. 121 (2010) 25–8. doi:10.1016/j.jsbmb.2010.03.041.
[40] L.C. de Macedo, F.C. Soardi, N. Ananias, V.M.S. Belangero, S.Z.P. Rigatto, M.P. De-Mello, et al., Mutations in the vitamin D receptor gene in four patients with hereditary 1,25-dihydroxyvitamin D-resistant rickets., Arq. Bras. Endocrinol. Metabol. 52 (2008) 1244–51. doi:10.1590/S0004-27302008000800007.
[41] P.J. Malloy, T.R. Eccleshall, C. Gross, L. Van Maldergem, R. Bouillon, D. Feldman, Hereditary vitamin D resistant rickets caused by a novel mutation in the vitamin D receptor that results in decreased affinity for hormone and cellular hyporesponsiveness., J. Clin. Invest. 99 (1997) 297–304. doi:10.1172/JCI119158. [42] G.K. Whitfield, S.H. Selznick, C.A. Haussler, J.C. Hsieh, M.A. Galligan, P.W. Jurutka, et al., Vitamin D
receptors from patients with resistance to 1,25-dihydroxyvitamin D3: point mutations confer reduced transactivation in response to ligand and impaired interaction with the retinoid X receptor heterodimeric partner., Mol. Endocrinol. 10 (1996) 1617–31. doi:10.1210/mend.10.12.8961271.
[43] P.J. Malloy, R. Xu, L. Peng, S. Peleg, A. Al-Ashwal, D. Feldman, Hereditary 1,25-Dihydroxyvitamin D Resistant Rickets due to a Mutation Causing Multiple Defects in Vitamin D Receptor Function, Endocrinology. 145 (2004) 5106–5114. doi:10.1210/en.2004-0080.
[44] P.J. Malloy, R. Xu, L. Peng, P.A. Clark, D. Feldman, A novel mutation in helix 12 of the vitamin D receptor impairs coactivator interaction and causes hereditary 1,25-dihydroxyvitamin D-resistant rickets without alopecia., Mol. Endocrinol. 16 (2002) 2538–46. doi:10.1210/me.2002-0152.
[45] T. Katsumata, T. Tanaka, E. Ishibashi, M. Nakatsuka, The biological activity of ST-232, the main metabolite of falecalcitriol., KISO TO RINSHO. 30 (1996) 2991–2996.
24
[46] Y. Miyamoto, T. Shinki, Y. Ohyama, T. Kasama, H. Iwasaki, Regulation of vitamin D-responsive gene expression by fluorinated analogs of calcitriol in rat osteoblastic ROB-C26 cells., J. Biochem. 118 (1995) 1068– 76. doi:10.1093/jb/118.5.1068.
[47] C. Carlberg, Genome-wide (over)view on the actions of vitamin D., Front. Physiol. 5 (2014) 167. doi:10.3389/fphys.2014.00167.
[48] A. Khan, N.A. Bryden, M.M. Polansky, R.A. Anderson, Insulin potentiating factor and chromium content of selected foods and spices., Biol. Trace Elem. Res. 24 (1990) 183–8.
[49] J. Li, T. Liu, L. Wang, X. Guo, T. Xu, L. Wu, et al., Antihyperglycemic and antihyperlipidemic action of cinnamaldehyde in C57BLKS/J db/db mice., J. Tradit. Chin. Med. 32 (2012) 446–52.
[50] S. Drori, G.D. Girnun, L. Tou, J.D. Szwaya, E. Mueller, K. Xia, et al., Hic-5 regulates an epithelial program mediated by PPARgamma., Genes Dev. 19 (2005) 362–75. doi:10.1101/gad.1240705.
[51] J.M. Lehmann, X.K. Zhang, G. Graupner, M.O. Lee, T. Hermann, B. Hoffmann, et al., Formation of retinoid X receptor homodimers leads to repression of T3 response: hormonal cross talk by ligand-induced squelching., Mol. Cell. Biol. 13 (1993) 7698–707.
[52] X.K. Zhang, J. Lehmann, B. Hoffmann, M.I. Dawson, J. Cameron, G. Graupner, et al., Homodimer formation of retinoid X receptor induced by 9-cis retinoic acid., Nature. 358 (1992) 587–91. doi:10.1038/358587a0. [53] L. Nagy, P. Tontonoz, J.G. Alvarez, H. Chen, R.M. Evans, Oxidized LDL regulates macrophage gene
expression through ligand activation of PPARgamma., Cell. 93 (1998) 229–40.
[54] S.H. Kim, S.Y. Choung, Antihyperglycemic and antihyperlipidemic action of Cinnamomi Cassiae (Cinnamon bark) extract in C57BL/Ks db/db mice., Arch. Pharm. Res. 33 (2010) 325–33. doi:10.1007/s12272-010-0219-0. [55] T. Tanaka, J. Yamamoto, S. Iwasaki, H. Asaba, H. Hamura, Y. Ikeda, et al., Activation of peroxisome
proliferator-activated receptor delta induces fatty acid beta-oxidation in skeletal muscle and attenuates metabolic syndrome., Proc. Natl. Acad. Sci. U. S. A. 100 (2003) 15924–9. doi:10.1073/pnas.0306981100. [56] P. Tontonoz, E. Hu, R.A. Graves, A.I. Budavari, B.M. Spiegelman, mPPAR gamma 2: tissue-specific regulator
of an adipocyte enhancer., Genes Dev. 8 (1994) 1224–34.
[57] B.M. Forman, P. Tontonoz, J. Chen, R.P. Brun, B.M. Spiegelman, R.M. Evans, 15-Deoxy-delta 12,
14-prostaglandin J2 is a ligand for the adipocyte determination factor PPAR gamma., Cell. 83 (1995) 803–12. [58] R.T. Nolte, G.B. Wisely, S. Westin, J.E. Cobb, M.H. Lambert, R. Kurokawa, et al., Ligand binding and
co-activator assembly of the peroxisome proliferator-activated receptor-gamma., Nature. 395 (1998) 137–43. doi:10.1038/25931.