PLD4 欠損マウス脳の組織学的解析
Histological analysis of the brain in
PLD4-deficient mice
研究分野
薬理学
指導教授
馬場 広子
略 語 表
APS: ammonium persulfate
BSA: bovine serum albumin
CD: cluster of differentiation
DAB: 3, 3’-diaminobenzidine
DAPI: 4’, 6-diamidino-2-phenylindole
EAE: experimental autoimmune encephalomyelitis
GFAP: glial fibrillary acidic protein
HE: hematoxylin-eosin
Iba1: ionized calcium-binding adapter molecule 1
LPS: lipopolysaccaride
MAG: myelin associated glycoprotein
MBP: myelin basic protein
MOG: myelin-oligodendrocyte glycoprotein
MS: multiple sclerosis
PBS: phosphate-buffered saline
PCR: polymerase chain reaction
PFA: paraformaldehyde
PH: pleckstrin homology
PLD: phospholipase D
PLP: myelin proteolipid protein
PVDF: polyvinylidine difluoride
PX: phox homology
RT-PCR: reverse transfection-PCR
SDS: sodium dodecyl sulfate
SDS-PAGE: sodium dodecyl sulfate-polyacrylamide electrophoresis
SPF: specific pathogen free
siRNA: small interfering RNA
tetO: tetracycline operator
TBS: Tris-bufferd saline
T-TBS: 0.1 % Tween 20/TBS
3 ら れ て い る 酵 素 タ ン パ ク 質 で ,ミ ト コ ン ド リ ア 膜 に 存 在 す る 脂 質 カ ル ジ オ リ ピ ン を 加 水 分 解 し , シ グ ナ ル 分 子 で あ る ホ ス フ ァ チ ジ ン 酸 を 産 生 す る 2 0 ). さ ら に ミ ト コ ン ド リ ア の 融 合 過 程 お よ び 精 母 細 胞 発 達 過 程 に 生 じ る ミ ト コ ン ド リ ア の 変 化 に 働 い て い る こ と が 知 ら れ て い る 2 1 ). PLD4 は ,当 研 究 室 と 理 化 学 研 究 所 脳 セ ン タ ー の 古 市 貞 一 博 士 (現 在 は 東 京 理 科 大 学 教 授 ) の グ ル ー プ と の 共 同 研 究 に よ り , ミ ク ロ グ リ ア 以 外 の 他 の グ リ ア 系 細 胞 お よ び ニ ュ ー ロ ン に は 発 現 が 認 め ら れ ず ,活 性 化 し た ミ ク ロ グ リ ア に 発 現 増 加 す る タ ン パ ク 質 と し て 同 定 さ れ た 分 子 で あ る .PLD4 は 他 の PLD と は 異 な り ,小 脳 で は 生 後 3 日 目 か ら 発 現 レ ベ ル が 上 昇 し 始 め ,生 後 7 日 目 で そ の 発 現 量 が 最 大 と な り ,そ の 後 は 漸 減 し ,生 後21 日 目 に は 消 失 す る (Fig.3; Yoshikawa et al., 2010). 特 に , 発 達 過 程 の マ ウ ス 小 脳 白 質 に 存 在 す る 活 性 化 ミ ク ロ グ リ ア に 発 現 し て お り ,ま た 中 枢 神 経 の 脱 髄 時 に 増 加 す る 活 性 化 ミ ク ロ グ リ ア に も そ の 存 在 が 認 め ら れ て い る 2 2 ). 脳 組 織 以 外 で は , 脾 臓 , 胸 腺 お よ び 肝 臓 な ど の 細 網 内 皮 系
組 織 に も 発 現 し て い る (Fig.3).PLD4 は 503 の ア ミ ノ 酸 残 基 か ら 構 成 さ れ ,N 末 端 側 に 1 つ の 膜 貫 通 domain を 有 し , C 末 端 側 に は 2 つ の HKD モ チ ー フ が あ る (Fig.2; Yoshikawa et al., 2010 ). 古 典 的 な PLD フ ァ ミ リ ー が リ ン 脂 質 を 加 水 分 解 す る 酵 素 活 性 を 示 す の に 不 可 欠 な 領 域 で あ る HKD モ チ ー フ を 持 っ て い る に も 関 わ ら ず PLD4 は PLD と し て の 酵 素 活 性 を 示 さ な い た め ,そ の 機 能 は 未 だ 不 明 で あ る . こ れ ま で に lipopolysaccharide (LPS) に よ り 活 性 化 さ れ た 培 養 系 の ミ ク ロ グ リ ア で は ,発 現 量 の 増 加 し た PLD4 は ま ず 核 質 部 に 存 在 し ,bioparticle を 貪 食 さ せ る と 食 胞 に 集 積 す る こ と が 明 ら か に な っ て い る 2 2 ). 最 近 , 比 較 的 日 本 人 に 多 い 自 己 免 疫 性 疾 患 で あ る 全 身 性 強 皮 症 の 罹 患 率 の 指 標 と し て PLD4 遺 伝 子 が 報 告 さ れ た 2 3 ). ウ シ で は PLD4 遺 伝 子 の ナ ン セ ン ス 変 異 に よ り , 亜 鉛 欠 乏 症 に 似 た 症 状 を 示 す 2 4 ) な ど 免 疫 系 が 関 与 す る 病 態 と の 関 連 も 報 告 さ れ て い る .し か し な が ら , 発 達 期 の 脳 の 白 質 に 特 徴 的 な 発 現 様 式 を 示 す PLD4 の 機 能 お よ び 脳 の 発 達 過 程 に お け る 役 割 に つ い て は 明 ら か に さ れ て い な い . そ こ で ,生 体 内 に お け る PLD4 の 機 能 を 明 ら か に す る た め に ,当 研 究 室 で PLD4 欠 損 マ ウ ス が 作 製 さ れ た .PLD4 は ,マ ウ ス 胎 仔 の in situ hybridization 法 に よ る mRNA 発 現 デ ー タ ベ ー ス で 胎 生 14.5 日 目 か ら 胸 腺 に 強 く 発 現 す る こ と が 示 さ れ て い る [genepaint (http://www.genepaint.org/), EH4997, Embryo C4027 3 4A] . そ の た め ,PLD4 欠 損 マ ウ ス で は 胎 生 致 死 に 至 る 可 能 性 も 考 え ら れ た こ と か ら コ ン デ ィ シ ョ ナ ル ノ ッ ク ア ウ ト マ ウ ス と し て 作 製 さ れ た .コ ン デ ィ シ ョ ナ ル ノ ッ ク ア ウ ト マ ウ ス の 作 製 に は Flexible Accelerated STOP Tetracycline Operator (tetO)-knockin (FAST) system 2 5 ) (Fig.4) の タ ー ゲ ッ テ ィ ン グ ベ ク タ ー を 用 い た 2 6 ).
6
Fig. 1 The three states of microglia. (modified from Neuroglia pp.86, 1995)
7
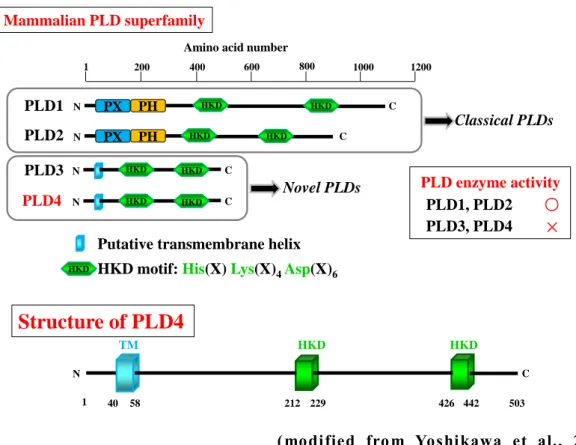
(modified from Yoshikawa et al., 2010) Fig. 2 Structural comparisons of PLD superfamily members.
Protein strucures and domains of mouse PLD family members (PLDs); PLD1, PLD2, PLD3, and PLD4. Top line r epresents a mino acid position. Mammalian PLDs ar e divided into two types; the classical type including PLD1 and PLD2 and the novel type including PLD3 and PLD4. PLD4 is a protein composed of 503 amino acids and has a transmembrane domain and two HKD motifs but dose not have either a PX or PH domain. The HKD motif is necessary for the enzymatic activity of PLD1 and PLD2 but PLD4 does not exhibit phosphatidyl choline-specific phospholipase activity.
PLD4 PLD1 PLD2 PLD3 Novel PLDs Classical PLDs 1 200 400 600 800 1200
Amino acid number
1000 N C N PX PH C N PX PH C N C HKD HKD HKD HKD HKD HKD HKD HKD
Putative transmembrane helix HKD
N C
1 40 58 212 229 426 442 503
Structure of PLD4
TM HKD HKD
HKD motif: His(X) Lys(X)4Asp(X)6
PLD enzyme activity PLD3, PLD4 PLD1, PLD2 ○
×
8
(Yoshikawa et al., 2010) Fig. 3 Developmental change in cerebellum and tissue distribution of PLD family mRNA.
Semi-quantitative real time PCR (qRT -PCR) analysis of PLD fa mily mRNA in
cerebellum at embryonic day18 (left), postnatal day (P) 0, P3, P7, P12, P15, P21, and P56. The expression of PLD4 was det ected gradually from P0, peaked at P7 and rapidly decreased to adult level at P21 in mouse cerebellum. Analysis of PLD fa mily mRNA in various tissues at P7 (right). PLD4 was detect ed in the thymus, liver, and spleen.
PLD4
PLD3
PLD1
PLD2
E18
P0
P3
P7
P12
P15
P21
P56
Brain
Th
ymus
Lu
ng
Hear
t
Liver
Sp
leen
Ki
dney
T
este
s
Cerebellum
Tissue Distribution (P7)
Novel PLDs
9
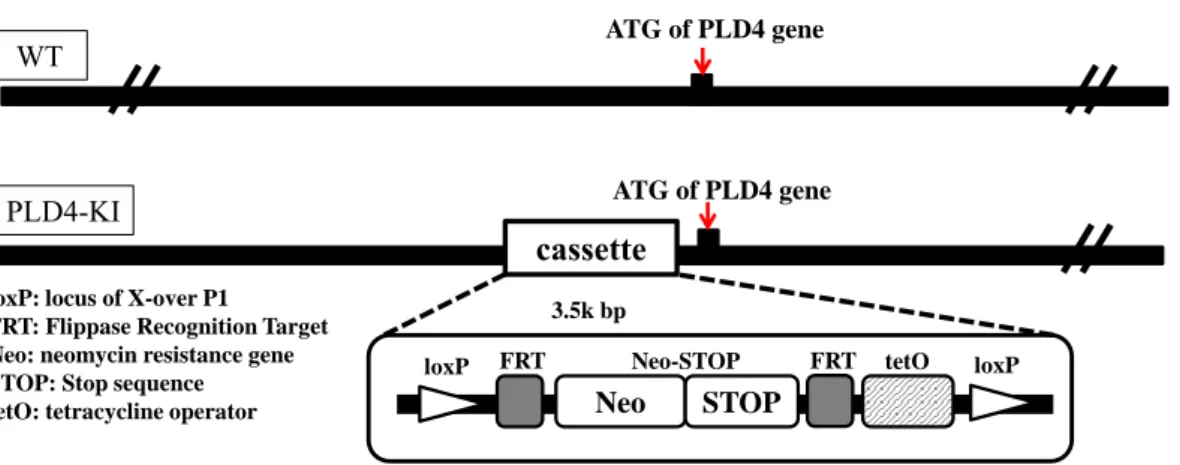
Fig. 4 Genome structure of the PLD4 gene in PLD4 knock-in (PLD4-deficient) mice.
The PLD4 knock-in (PLD4-deficient) mice (PLD4 -KI) were generated by the homologous recombination using t he PLD4-targeting vector containing a STOP-tetO cassette inserted i n front of the start codon, The STOP-tetO cassette contained sequences for ; loxP, FRT, a neomycin resistant gene, STOP, FRT, tetO, and loxP in order. WT, wild type.
WT ATG of PLD4 gene
PLD4-KI
cassette loxP: locus of X-over P1
FRT: Flippase Recognition Target Neo: neomycin resistance gene STOP: Stop sequence
tetO: tetracycline operator Neo STOP
loxP FRT Neo-STOP FRT tetO loxP 3.5k bp
10
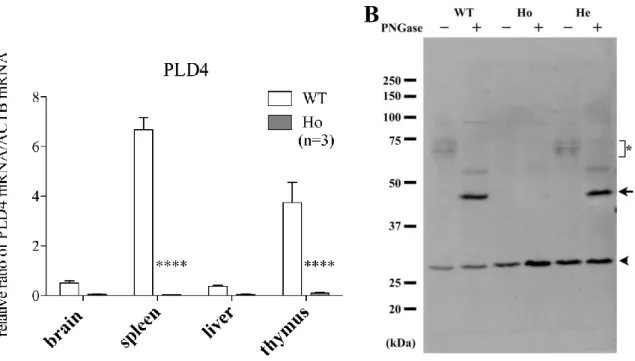
Fig. 5 Elimination of PLD4 mRNA and protein in PLD4-deficent mice.
A, Expression of PLD4 mRNA by qRT-PCR in brain, spleen, liver and thymus of wild type (WT) and PLD4-deficient (Ho) mice. Graphs show the relative ratio of the level of PLD4 and β-actin mRNA from samples run in triplicate from 3 independent experiments. In wild type, PLD4 mRNA was expressed in brain and various reticuloendothelial tissues, inculuding the spleen, liver and thymus . In PLD4-deficient mice, PLD4 mRNA was complet ely eliminated in these tissues. B, PLD4 protein levels in adult wild type (WT), homozygote (Ho), and heterozygote (He) mice. Spleen homogenates (10 µg) with or without deglycosylation were separated by 10.5% sodiu m dodecylsulfate polyacryla mide gel electrophoresis (SDS -PAGE) and Western blot analysis was perfor med using an anti -PLD4 antibody. Since PLD4 has multiple glycosylation sites, various PLD4 -related bands (70~75 kDa; indicated by *) wer e shown in non-treated sa mples. The arrow indicates deglycosylated PLD4 (~45 kDa) by PNGase F (peptide-N-glycosidase F) treatment. The arrowhead indicates an unrelated protein product detect ed by the anti -PLD4 antibody. Note that this pritein was also detected in PLD4-deficient spleen sa mples while PLD4 bands with or without sugars were completely eli minated.
11
Fig. 6 The anatomy of the cerebellum
Schematic diagrams of overview of brain (A), median sagittal section of cerebellum (B), and three layers of cerebellar cortex (C) were depicted.
medulla oblongata pons
Bergmann glia molecular layer
Purkinje cell layer granule cell layer
White matter climbing fiber mossy fiber
cerebellum
cerebellar nucleus
deep white matter folial white matter
12
第
1 節 実 験 材 料 及 び 実 験 方 法
1-1 試 薬 類
特 に 明 記 し て い な い 一 般 試 薬 は 和 光 純 薬 , 関 東 化 学 お よ び シ グ マ -ア ル ド リ ッ チ ジ ャ パ ン の 特 級 を 使 用 し た .各 種 溶 液 の 調 製 に は 超 純 水 (MilliQ 水 ; Millipore) を 用 い た .1-2 実 験 動 物
系 統 維 持 用 の C57BL/6N マ ウ ス は , 日 本 エ ス エ ル シ ー か ら 購 入 し た . 当 研 究 室 で 作 製 し た PLD4 KI マ ウ ス 2 6 ) に 関 し て は 交 配 に よ り 生 ま れ た 仔 の 遺 伝 子 型を polymerase chain reaction (PCR) 法 で 診 断 し , 野 生 型 (WT) お よ び ホ モ 接 合 体 (Ho, PLD4 欠 損 マ ウ ス ) を 実 験 に 使 用 し , ヘ テ ロ 接 合 体 (He) を マ ウ ス コ ロ ニ ー の 維 持 に 用 い た .
DNA の 抽 出 は , ア ル カ リ 溶 解 法 に よ り 行 っ た . 生 体 試 料 (生 後 4 週 齢 マ ウ ス の 尾 ,2 mm 程 度 ) に 50 mM NaOH を 180 µl 加 え 混 合 し , 95℃ , 10 min イ ン キ ュ ベ ー ト し た .そ の 後 ,1 M Tris-HCl (pH 8.0) を 20 µl 加 え 混 合 し ,12000 rpm, 5 min で 遠 心 分 離 し た (MX-200,TOMY).上 清 を DNA 抽 出 液 と し て 用 い ,下 記 の 条 件 で PCR を 行 っ た .
<診 断 用 プ ラ イ マ ー >
13 <PCR 条 件 > PCR 後 の 溶 液 は ,1/5 量 の 6×loading buffer を 添 加 し 1.2% ア ガ ロ ー ス ゲ ル に て 135V, 30 分 間 電 気 泳 動 を 行 い (i-MyRun.NC, コ ス モ ・ バ イ オ ), エ チ ジ ウ ム ブ ロ マ イ ド 染 色 を 行 っ た .野 生 型 で は PLD4-upstrea m と PLD4-downstrea m の プ ラ イ マ ー か ら 1.5-kbp に ,PLD4 欠 損 マ ウ ス で は tetO-upstream と PLD4-downstrea m の プ ラ イ マ ー か ら 850-bp に PCR 産 物 と し て バ ン ド が 検 出 さ れ る . 本 実 験 で 使 用 さ れ た 実 験 動 物 は , 東 京 薬 科 大 学 実 験 動 物 施 設 内 の specific pathogen free (SPF) 区 域 に お い て 恒 温 (23±1℃ ), 恒 湿 (55±5%), 12 時 間 周 期 の 定 時 照 明 下 で 飼 料 と 水 を 自 由 に 与 え て 飼 育 さ れ た .動 物 の 取 り 扱 い に 関 し て は , 東 京 薬 科 大 学 動 物 実 験 規 程 に 基 づ い て 行 わ れ た .本 実 験 計 画 は 東 京 薬 科 大 学 実 験 動 物 委 員 会 で 審 査 さ れ , 学 長 か ら の 承 認 を 受 け た 後 に 実 施 さ れ た (P12-23, P13-31, P14-09, P15-26, P15-27).
1-3 遺 伝 子 組 換 え 実 験
本 研 究 計 画 は 東 京 薬 科 大 学 組 換 え DNA 実 験 安 全 委 員 会 で 審 査 さ れ ,学 長 か ら の 承 認 を 受 け た (Y2012-12, Y2013-9, Y2014-23, Y2015-23). 本 実 験 は 「 東 京 薬 科 大 学 組 換 え DNA 実 験 安 全 管 理 規 則 」 に 則 っ て 行 わ れ た .1-4 灌 流 固 定
生 後 0, 3, 5, 7 お よ び 10 日 目 (P5, 7, お よ び P10)の マ ウ ス を ジ エ チ ル エ ー テ ル に よ っ て 吸 入 麻 酔 し ,仰 臥 位 で 保 定 し た 後 に 開 胸 し た .右 心 耳 を 切 開 し ,先 端 を 鈍 化 し た 26G の 針 を 心 尖 部 よ り 左 心 室 内 に 刺 入 後 ,最 終 濃 度 が 0.1 %に な る よ う に ヘ パ リ ン (持 田 製 薬 ) を 加 え た 室 温 の 生 理 食 塩 水 (大 塚 製 薬 ) を 体 重 と 同 量 灌 流 し , 血 液 を 洗 い 流 し た . 引 き 続 き , 氷 上 に て 冷 却 し た 体 重 2 倍 量 の 4% KOD 2×PCR buffer 7.5 µ l dNTP 3 µ l pr imer up (upstr ea m) 0.3 µ l pr imer low (downstr ea m) 0.3 µ lMilliQ 2.6 µ l
KOD Fxneo (T OYOBO) 0.3 µ l
14
paraformaldehyde (PFA)/0.1 M phosphate buffer (PB) (pH 7.4) で 灌 流 固 定 し , 脳 を 摘 出 し た . 摘 出 さ れ た 脳 組 織 を 4% PFA/PB で 24 時 間 浸 漬 固 定 し た .
1-5 パ ラ フ ィ ン 切 片 の 作 製
1-4 で 浸 漬 固 定 し た 組 織 サ ン プ ル を Tissue-Tek VIP (サ ク ラ フ ァ イ ン テ ッ ク ジ ャ パ ン) を 用 い パ ラ フ ィ ン 包 埋 し た . 室 温 下 , ミ ク ロ ト ー ム (大 和 光 機 工 業 ) で 厚 さ 6 µ m に 組 織 を 薄 切 し た . そ の 切 片 を MAS コ ー ト ス ラ イ ド ガ ラ ス あ る い は FRONTIER コ ー ト ス ラ イ ド ガ ラ ス (松 浪 硝 子 工 業 ) に の せ ,37℃ に 設 定 し た 伸 展 機 (大 和 光 機 工 業 ) で 1 晩 乾 燥 さ せ た 後 , 組 織 解 析 に 使 用 し た .1-6 免 疫 組 織 化 学 的 染 色
1-5 で 作 製 し た パ ラ フ ィ ン 切 片 を 室 温 下 , 脱 パ ラ フ ィ ン 処 理 し (キ シ レ ン 15 分 間 3 回 ),続 い て 親 水 性 処 理 (100% エ タ ノ ー ル 3 分 3 回 ,90%,80%,70% エ タ ノ ー ル 各 3 分 1 回 ) を 行 っ た .続 い て 抗 原 賦 活 化 の た め に 0.1 M ク エ ン 酸 溶 液 に 浸 漬 し ,電 子 レ ン ジ で 沸 騰 さ せ た 後 ,容 器 ご と 40 分 間 放 置 し ,室 温 に 戻 し た . そ の 後 ,phosphate buffered saline (PBS; 137 mM NaCl, 81 mM Na2HPO4, 2.68mM KCl, 1.47 mM KH2PO4, pH7.4) に て 5 分 間 洗 浄 後 , 3% H2O2 中 で 10 分 間 反
応 さ せ る こ と に よ り 内 因 性 の ペ ル オ キ シ ダ ー ゼ を 失 活 さ せ ,PBS で 5 分 間 洗 浄 し た .そ の 後 ,Blocking 溶 液 [10% normal donkey serum (Dako),0.3% TritonX-100, PBS] で 1 時 間 処 理 す る こ と に よ り 抗 体 の 非 選 択 的 吸 着 を 防 止 す る た め の blocking を 行 っ た . Blocking 溶 液 で 希 釈 し た 一 次 抗 体 を 4℃ で 一 晩 反 応 さ せ た . そ の 後 ,室 温 に て PBS で 5 分 間 ず つ 3 回 洗 浄 し ,ビ オ チ ン 化 二 次 抗 体 を 30 分 間 反 応 さ せ ,ABC-kit (Vector Laboratory) の ABC 混 合 液 で 30 分 間 処 理 し た .PBS で 3 回 洗 浄 し た 後 , 3,3-dia minobentidine (Dako)・ H2O2 反 応 液 を 用 い て 顕 微 鏡 下
15
1-7 Hematoxilin-Eosin (HE)染 色
1-5 で 作 製 し た パ ラ フ ィ ン 切 片 を 脱 パ ラ フ ィ ン 処 理 (キ シ レ ン 15 分 間 3 回 )後 に ,親 水 性 処 理 (100% エ タ ノ ー ル 3 分 3 回 ,90%,80%,70% エ タ ノ ー ル に 各 3 分 1 回 ) を 行 い , hematoxylin (Mayer ’s hematoxylin solution; Sigma) に 10 分 間 浸 し た 後 ,流 水 で 10 分 間 洗 浄 し た .さ ら に 純 水 を 満 た し た 染 色 バ ッ ト で 軽 く 洗 浄 し た 後 ,Eosin (1% eosin Y solution; Sigma) に 1 分 間 浸 し ,そ の 後 純 水 に て 1 分 間 洗 浄 を 行 い , 脱 水 お よ び 透 徹 し ,MGK-S で 封 入 し た . 画 像 は デ ジ タ ル カ メ ラ 付 き 光 学 顕 微 鏡 で 撮 影 し た (Axio Imager M1 with AxioCa m HRc あ る い は BZ-X700).
1-8 凍 結 切 片 の 作 製
1-4 で 作 製 し た 浸 漬 固 定 後 の 組 織 を PBS に て 5 分 間 3 回 洗 浄 し , 4℃ の 10% sucrose/PBS,15% sucrose/PBS ,20% sucrose/PBS に 一 晩 ず つ 浸 し ,Tissue-Tek O.C.T. compound (サ ク ラ フ ァ イ ン テ ッ ク ジ ャ パ ン ) に 包 埋 し ,ド ラ イ ア イ ス で 凍 結 さ せ た . ク ラ イ オ ス タ ッ ト (LEIC A CM 1850; Leica) で 10 µm に 薄 切 し た 組 織 切 片 を MAS コ ー ト あ る い は FRONTIER コ ー ト し た ス ラ イ ド ガ ラ ス 上 で 風 乾 さ せ た 後 , 使 用 時 ま で -40℃ で 保 存 し た .
1-9 蛍 光 免 疫 組 織 染 色
16
1-10 リ ゾ レ シ チ ン 投 与 し た 坐 骨 神 経
抗 arginase 1 抗 体 に よ り 小 脳 を 免 疫 染 色 す る 際 の ポ ジ テ ィ ブ コ ン ト ロ ー ル と し て lysolecithin 投 与 後 ICR マ ウ ス 坐 骨 神 経 の 切 片 を 使 用 し た .用 い た 切 片 の 作 製 法 は 以 下 の 通 り で あ る . 8 週 齢 雄 の ICR マ ウ ス に ソ ム ノ ペ ン チ ル 注 射 液 (共 立 製 薬 ) を 生 理 食 塩 水 で 10 倍 に 希 釈 し た 溶 液 を 体 重 あ た り 10 ml/kg に な る よ う に 腹 腔 内 投 与 し て 麻 酔 し た .1% lysolecithin [L-α-Lysophosphatidylcholine (Sigma)]/Locke’s solution [154 mM NaCl, 2 mM CaCl2, 5.6 mM KCl/10 mM 4 -(2-hydoroxyethyl)-1-piperazethane17
1-11 抗 体
免 疫 組 織 染 色 で 使 用 し た 抗 体 は 以 下 の 通 り で あ る .カ ッ コ 内 に は 希 釈 倍 率 を 示 す .
一 次 抗 体
rabbit polycl ona l a nt i-ioniz ed ca lciu m-bindi ng a dapt er mol ecul e 1 (Iba 1) a nt ibody ( 和 光 純 薬 )
(1:800) rat monoc lona l a nt i- myeli n basic pr ot ei n (MBP) a nt ibody (C hemic on) (1:100) rabbit polycl ona l a nt i-gl ia l fibr illar y a cidic pr ot ei n (GF AP ) a nt ibody (DAKO) (1:10) rabbit polycl ona l a nt i-ca lbindi n D-28K a nt ibody (C hemicon) (1:200) rat monoc lona l a nt i-clust er of diff er ent ia t ion 68 ( C D68 ) a nt ibody ( Abca m) (1:100) goa t pol ycl ona l a nt i-ar gi nase 1 a nt ibody (Sa nt a Cruz Biot echnology) (1:50) rabbit polycl ona l a nt i-NG2 pr ot eoglyca n a nt ibody (C he mic on) (1:500) rabbit polycl ona l a nt i-Olig2 a nt ibody (IBL) (1:200) mouse monocl ona l a nt i-a denoma t ous pol yposi s coli ( APC /CC 1) a nt ibody
(Mer ck Mil lipor e)
(1:100)
二 次 抗 体
19
1-14 GFAP 陽 性 像 輝 度 測 定 方 法
Image J を 用 い , GFAP 陽 性 像 の 輝 度 を 測 定 す る た め に 視 野 内 の 陽 性 細 胞 が 集 ま る 白 質 領 域 (緑 ) を DAPI 染 色 像 (青 ) を も と に 決 定 し (下 図 ),そ の 面 積 と 領 域 中 の 蛍 光 輝 度 を 測 定 し た . 輝 度 を 面 積 で 割 り , 単 位 面 積 (1 µm2) あ た り の 輝 度 を GFAP 蛍 光 強 度 と し た .1-15 Olig2 陽 性 細 胞 数 測 定 方 法
観 察 視 野 内 の 小 脳 深 部 白 質 の Olig2 陽 性 細 胞 数 を 計 測 し ,DAPI 陽 性 の 全 細 胞 数 に 対 す る 割 合 を 算 出 し た .1-16 組 織 ホ モ ジ ネ ー ト の 調 製
生 後 7 日 目 の 野 生 型 と PLD4 欠 損 マ ウ ス を ジ エ チ ル エ ー テ ル に よ っ て 吸 入 麻 酔 し 断 頭 後 , 脳 を 摘 出 し , そ こ か ら 小 脳 の み を 採 取 し た . 小 脳 重 量 当 た り 9 倍 量 の 氷 冷 し た Homogenization buffer [0.32 M sucrose, 0.75 µM aprotinin, 1 µ M leupeptin, 1 µM pepstatin A, 0.4 µM phenylmethylsulfonyl fluo ride, 1 mM dithiothreitol (DTT)] を 加 え , ポ ッ タ ー 型 テ フ ロ ン ― ガ ラ ス ホ モ ジ ナ イ ザ ー (Digital Homogenizer, Iuchi) を 用 い て 氷 冷 下 で ホ モ ジ ナ イ ズ し た . そ の 後 , 冷 却小 脳 白 質 内 GFAP 陽 性 (緑 )領 域 を 決 定 す る
20
遠 心 分 離 機 (MX-200, TOMY) に て 遠 心 分 離 (1000 ×g, 10 min, 4℃ ) し , 核 を 除 い た 上 清 を ホ モ ジ ネ ー ト と し た . 得 ら れ た 組 織 ホ モ ジ ネ ー ト は bovine serum albumin (BSA) を 基 準 と し た BC A 法 (PIERCE あ る い は タ カ ラ バ イ オ ) で タ ン パ ク 質 濃 度 を 定 量 し た 後 に 分 注 し , -80℃ に て 保 存 し た .
1-17 Western blot 法
1-17-1 SDS-PAGE
21
Table.1 Recipes for polyacrylamide gels for SDS -PAGE S eparat ing gel 12. 0%
4×Tr is -HC l, pH8.8 1.5 ml 30% a cr yla mi de/
2.4 ml 0. 8% bisa cr yla mide mixt ur e
10% S DS 0.06 ml Mill iQ 水 2.04 ml TEMED 0.004 ml 10% APS 0.02 ml
Sta cki ng gel 4.0% 4×Tr is -HC l, pH6.8 0.9 ml 30% a cr yla mi de/
0.48 ml 0. 8% bisa cr yla mide mixt ur e
10% S DS 0.036 ml Mill iQ 水 2.16 ml TEMED 0.0036 ml 10% APS 0.018 ml APS: ammonium persulfate
TEMED: N,-N,-N’,-N’-tetramethylethylendiamine
1-17-2 転 写
22
1-17-3 免 疫 化 学 発 光 法 に よ る タ ン パ ク 質 の 検 出
1-17-2 で タ ン パ ク 質 が 転 写 さ れ た PVDF 膜 を 0.1% Tween 20/Tris -buffered saline (T-TBS) (0.1% Tween 20, 150 mM NaCl, 20 mM Tris -HCl, pH 7.5) で 洗 浄 し た 後 ,Blocking 溶 液 (0.3% skim milk (Becton Dickinson), T -TBS) 中 に て 室 温 で 1 時 間 処 理 し た .そ の 後 ,Blocking 溶 液 で 希 釈 し た 一 次 抗 体 と PVDF 膜 を 室 温 に て 60 分 間 反 応 さ せ た . 一 次 抗 体 反 応 後 , PVDF 膜 を T-TBS で 5 分 間 3 回 洗 浄 し た . 更 に ペ ル オ キ シ ダ ー ゼ 標 識 二 次 抗 体 を T-TBS で 希 釈 し , 洗 浄 後 の PVDF 膜 に の せ ,室 温 に て 60 分 間 反 応 さ せ た 後 ,T-TBS で 5 分 間 3 回 洗 浄 し た .そ の 後 ,PVDF 膜 に ECL 溶 液 (Amersham, GE Healthcare) を 1 cm2 あ た り 0.05 ml の
せ て 1 分 間 反 応 さ せ ,ル ミ ノ イ メ ー ジ ア ナ ラ イ ザ LAS-3000 (富 士 フ ィ ル ム ) を 用 い て 発 光 シ グ ナ ル を 検 出 し た .画 像 の 解 析 は ImageGauge Ver. 4.23 に て 行 っ た . MBP 遺 伝 子 は 1 つ の 転 写 産 物 か ら 選 択 的 ス プ ラ イ シ ン グ に よ り 異 な る 分 子 量 の タ ン パ ク 質 と な る こ と が 知 ら れ て い る . 主 要 な 4 本 の バ ン ド (14, 17, 18.5, 21 kDa) の 輝 度 を 測 定 し ,そ の 合 計 を 算 出 し た .内 因 性 control と し て β-actin を 用 い て 定 量 化 し た .
Western blot 法 で 使 用 し た 抗 体 は 以 下 の 通 り で あ る .カ ッ コ 内 に は 希 釈 倍 率 を 示 す .
一 次 抗 体
rat monoclonal anti-MBP antibody (CHEMICON) (1:4000) rabbit polyclonal a nti-β-actin antibody (和 光 純 薬 ) (1:2000) 二 次 抗 体
HRP-conjugated goat anti-rabbit IgG antibody (Jackson Immuno Research) (1:10000) HRP-conjugated goat anti-rat IgG antibody (Jackson Immuno Research) (1:10000)
1-18 統 計 学 的 解 析
23
1-19 ク プ リ ゾ ン モ デ ル の 作 製
1-2 で 診 断 し た マ ウ ス の う ち ,野 生 型 と PLD4 欠 損 マ ウ ス を 8~ 10 週 齢 ま で 通 常 の 飼 料 で 飼 育 し た . そ の 後 , ク プ リ ゾ ン 混 餌 を 5 週 間 , 自 由 摂 食 さ せ た .
ク プ リ ゾ ン (bis-cyclohexanone-oxaldihydrazone; Sigma -Aldrich) は 動 物 飼 育 用 の MF 飼 料 (オ リ エ ン タ ル 酵 母 工 業 ) に 0.2% (w/w) の 含 量 で 混 合 製 造 し た も の を 使 用 し た . 比 較 対 照 の マ ウ ス は . 通 常 の CE-2 (日 本 ク レ ア ) を 摂 取 さ せ た .
1-20 FluoroMyelin 染 色
25
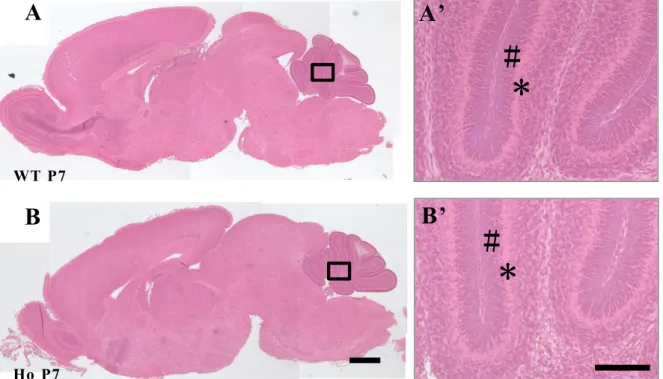
Fig. 7 Gross brain structures in PLD4 -deficient mice at P7.
Comparison of gross brain structures observed in wild type (WT; A) and PLD4-deficient mice (Ho; B) at P7. Paraffin sections were stained with hematoxylin and eosin. No significant differences in gross brain anatomy were observed. High magnification images of cerebellar cortex (blac k squares) are shown in the right panels (A’, B’). No significant differences in external granular layer (#) and Purkinj e cell layer (*) were observed. Scale bars in B and in the higher magnification image at
right represent 500 µm and 100 µ m, respectively.
27
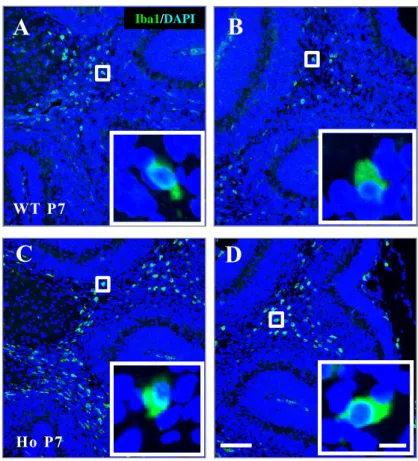
Fig. 8 Iba1-positive microglial activation in the cerebella of PLD4 -deficient mice at P5.
Frozen sections of deep whit e matter (A, C) or folia l white matter (B, D) of the cerebella obtained from wild type (A, B) and PLD4 -deficient (C, D) mice wer e immunostained using an anti-Iba1 antibody. A representative Iba1 -positive cel l indicated by the white square in each panel is shown at higher magnific ation in the lower right corners. Note that the microglia at P5 are the rounded activated type. Scale bars represent 100 µm (D) or 10 µm (in the white square of D).
28
Fig. 9 Iba1-positive microglial activation in the cerebella of PLD4 -deficient mice at P7.
Frozen sections of deep whit e matter (A, C) or folia l white matter (B, D) of the cerebella obtained from wild type (A, B) and PLD4 -deficient (C, D) mice wer e immunostained using an anti-Iba1 antibody. A representative Iba1 -positive cel l indicated by the white square in each panel is shown at higher magnification in the lower right corners. Note that the microglia at P7 are still the rounded activated type. Scale bars represent 100 µm (D ) or 10 µm (in the white square of D).
29
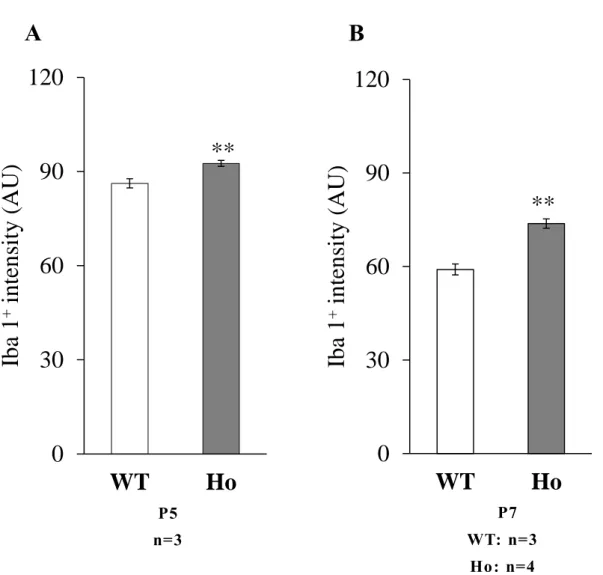
Fig. 10 Q uantitative intensity analysis of Iba1 -positive microglia in the deep white matter.
Intensities of Iba1 immunoreactivity in individual cells (see Materials and Methods 1-1-12) were measured in the deep cerebellar white matter at P5 (A) and P7 (B). Graphs indicate the mean ±SEM. Data were obtained from: 92 WT cells and 98 Ho cells at P5, a nd 125 WT cells and 130 Ho cells at P7 (n: number of mice). Statistica l analysis was perfor med using a Student t test. **P<0.01.
30
2-1-3 発 達 期 の PLD4 欠 損 マ ウ ス 小 脳 白 質 に 局 在 す る 活 性 化 ミ
ク ロ グ リ ア の 表 現 型
Iba1 陽 性 ミ ク ロ グ リ ア が 刺 激 を 受 け て 活 性 化 さ れ る と , M1 ミ ク ロ グ リ ア あ る い は M2 ミ ク ロ グ リ ア と 呼 ば れ る 2 つ の タ イ プ に 分 け ら れ る こ と が 知 ら れ て い る 3 7 ).M1 タ イ プ の ミ ク ロ グ リ ア は 炎 症 誘 発 を 介 し て 組 織 障 害 性 の 働 き を す るタ イ プ で あ り ,inducible nitric oxide synthase (iNOS) , tumor necrosis factor -α (TNFα),あ る い は int erleukin-1β (IL-1β) な ど の 炎 症 性 の サ イ ト カ イ ン の 産 生・分 泌 や CD68, CD32 あ る い は CD86 な ど の 炎 症 を 促 進 さ せ る 細 胞 表 面 マ ー カ ー を 発 現 さ せ る .一 方 ,M2 タ イ プ の ミ ク ロ グ リ ア は 抗 炎 症 作 用 に 関 与 す る タ イ プ で あ り ,IL-4, IL-10 あ る い は arginase 1 な ど の 炎 症 抑 制 性 因 子 を 産 生 す る 3 7 , 3 8 , 3 9 ).
31
32
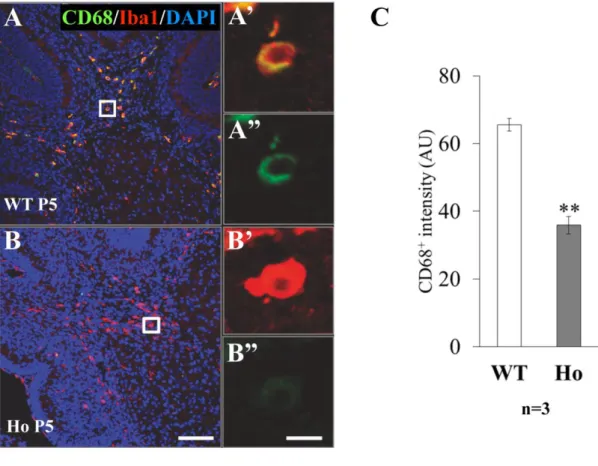
Fig. 11 CD68 immunoreactivity of Iba1 -positive microglia in cerebellar deep white matter at P5.
The deep whit e matter of P5 cerebellum obtained from WT (A) or PLD4 -deficient (B) mice was double-immunostained by anti-Iba1 (red) and anti-CD68 (green) antibodies. Higher magnification images of representative Iba1+/CD68+ cells were from the whit e
squrares in A, B. Scale bars indicate 100 µm (B) or 10 µm (B ’’). All Iba1-positive cells were also positive for CD68, however, the staining intensity of CD68 (see Materials and Methods 1 -1-13) in PLD4-deficient cells was significantly lower tha n that in wild-type cells (C). Data were obtained from: 78 WT cells and 65 Ho cells in the deep whit e matter (n: number of mice). Graphs indicate mean ± SEM. Statistical analysis was perfor med using a Student t test. **P<0.01.
B
B’
B’’
WT P5
33
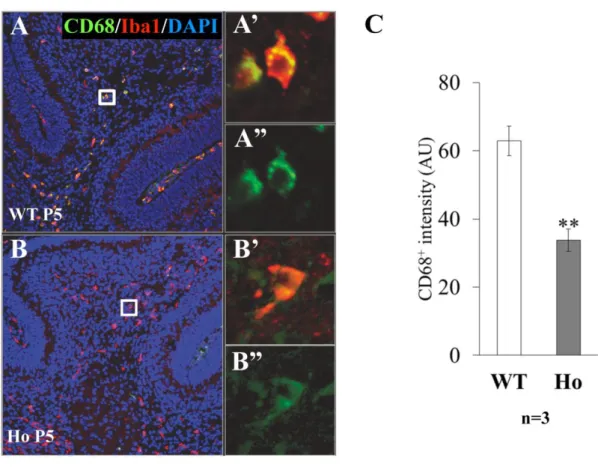
Fig. 12 CD68 immunoreactivity of Iba1 -positive microglia in folial white matter at P5.
The folial white matter of P5 cerebellum obtained from WT (A) or PLD4 -deficient (B) mice was double-immunostained by anti-Iba1 (red) and anti-CD68 (green) antibodies. Higher magnification images of representative Iba 1+ CD68+ cells were from the whit e
squrares in A, B. Scale bars indicate 100 µm (B) or 10 µm (B ’’). All Iba1-positive cells were also positive for CD68, however, the staining intensity of CD68 (see Materials and Methods) in PLD4 -deficient cells was significantly lower than in wild-type cells (C). Data were obtained from: 25 WT cells and 32 Ho cells in the folia l white matter (n: number of mice). Graphs indicate mean ± SEM. Statistical analysis was perfor med using a Student t test. **P<0.01.
B
B’
WT P5
34
Fig. 13 Arginase 1 immunoreactivity of Iba1 -positive microglia in cerebellar white matter at P5.
The deep white matter (A, C) and folial white matter (B, D) of P5 cerebellum obtained from WT (A, B) or PLD4 -deficient (C, D) mice were double-immunostained by anti-Iba1 (green) and anti-arginase 1 (red) antibodies. No Iba1 -positive microglia showed arginase 1 immu noreactivity in either genotype at P5. The positive control image of the section from the demyelinated lesion of lysolecithin -injected sciatic nerve was demonstrated (E). Representative arginase 1 -positive cells indicated by the white square is shown at higher magnification in the lower right corner in (E). Scale bars indicate 50 µm (D, E) or 10 µm (in the white squares of E).
B
C
D
WT Folia
Ho
Deep white matter
Ho Folia
Lysolecithin-injected sciatic nerve
Iba1/arginase1/DAPI
A
WT
Deep white matter
36 calbindin
Fig. 14 Purkinje cells in the cerebella of PLD4 -deficient mice.
Representative calbindin-positive Purkinje cells in wild type (WT; A, B) and PLD4-deficient (Ho; C, D) cerebella at P5 (A, C) and P7 (B, D) are exhibited. A ’ to D’ are higher magnification images of the white squares in A to D. No apparent differences were observed between wild type and PLD4 -deficient mice at both ages. Scale bars indicate 100 µm (D) or 20 µm (D ’).
37
Fig. 15 Astrocytes in the cerebella of PLD4 -deficient mice.
Representative GFAP -immunoreactive images of astrocytes in the deep whit e matter of wild type (A, B) and PLD4 -deficient (C, D) cerebella at P5 (A, C) and P7 (B, D) ar e exhibited. The sharp (#) in D represents astrocyte signal in the whit e matter. The asterisk in D represents Bergmann glia signals. Higher magnification images of the white squares are shown in the lower right corners. Scale bars indica te 100 µm (D), or 20 µ m (in the white squares of D).
38
Fig. 16 Quantitative intensity analysis of GFAP -positive area in the deep white matter.
Intensities of GFAP immunoreactivity in the deep cerebella white matter of P5 (A) and P7 (B) cerebella obtained from wild type (WT) and PLD4 -deficent (Ho) mice were quantified (see Materials and Methods 1 -14). GFAP+ intensities are represent ed as
intensity per unit area (1 µm2). Graphs in A and B indicate the mean ± SEM.
Statistical analyses with Student t tests showed no significant difference between the two genotypes at P5 (A) and P7 (B).
40
Fig. 17 Characteristics of microglia, Purkinje cells, and astrocytes in the cerebell a of PLD4-deficient mice at P10.
Representative images of Iba1 -positive microglia in wild type (WT; A, B) and PLD4-deficient (Ho; C, D) cerebella at P10 were exhibi ted. Higher magnification of a representative Iba1-positive microglia is shown in the lower right corner. At P10 process-bearing ra mified microglia were found in both types of mice. Purkinje cells in WT (E) and Ho (F) cerebella at P10 were immunostained by anti-calbindin antibody (E, F). E’ and F’ are higher magnification images of the white squares in E and F. GFAP immunoreactive astrocytes in the deep white matter (G, I) and folial whit e matter (H, J) of WT and Ho mice were immunostained by anti -GFAP antibody. Higher-magnification images of whit e squares (G, H, I, J) were indicated in the lower right corners. No apparent differences of calbindin a nd GFAP positive staining wer e observed between two genotypes. Scale bars indicate 100 µm (D, F, J) or 10 µm (in the white squares of D) or 20 µm (F ’, in the whit e squares of J).
43
Fig. 18 Developmental change of microglial activation in normal C57BL/6 mouse cerebellum.
Frozen sagittal sections of P0, P3, P5, P7, P10 cerebella obtained from normal
44
Fig. 19 Progress of myelination in normal C57BL/6 mouse cerebellum.
45
Fig. 20 Relationship between myelination and microglial activation during mouse cerebellar development.
Double immunostaining of the whit e matter in cerebellar folia at P7 with anti -Iba 1 (red) and anti-MBP (green) antibodies. The DAPI staining pattern is overlaid to indicate the cerebellar region. B and C are higher magnification images of the whit e squares in A. In mice, clusters of rounded Iba1-positive activated microglia ar e present in the area where myelination is in progress (C). Scale bar in A represents 100 µm or B represents 20 µ m.
A
47
Fig. 21 Protein expression of MBP in the cerebella of PLD4 -deficient mice at P7.
Proteins in P7 cerebellar homogenates (10 µg) from wild type (WT) or PLD4-deficient (Ho) mice (three mice from each group) were separated by 12% SDS -PAGE and Western blot was performed using an anti -MBP antibody (A). After stripping off the antibody, the same blot was restained with an anti -β-actin antibody for quantification. The total intensity of each MBP band (14, 17, 18.5, and 21 kDa) was quantified for each animal and the ratio of MBP to β-actin for each genotype was calculated (B) (see Materials and Methods 1 -17). Graph indicates mean ± SEM. Statistical analysis with a Student t test shows no significant difference between the two groups.
48
*
*
MBP/DAPI
Fig. 22 Decrease of MBP-positive myelinated fibers in cerebellum of PLD4-deficient mice.
MBP immunoreactivities in wild type (A, C, E) and PLD4 -deficient (B, D, F) cerebella at P5 (A, B), P7 (C, D) and P10 (E, F) were compared. Cell nuclei were stained wit h DAPI (blue). Representative images of 5 mice from each genotype were select ed. As shown in Fig.19, myelination in the deep white matter is in progress at P5 and
49
50
2-1-8 PLD4 欠 損 マ ウ ス 小 脳 に お け る オ リ ゴ デ ン ド ロ サ イ ト の
分 化
オ リ ゴ デ ン ド ロ サ イ ト の 系 譜 は ,脳 形 成 の 過 程 に お い て グ リ ア 系 の 幹 細 胞 か ら 生 じ る オ リ ゴ デ ン ド ロ サ イ ト 前 駆 細 胞 (oligodendrocyte progenitor cell, migrating progenitor) か ら 始 ま る . オ リ ゴ デ ン ド ロ サ イ ト の 分 化 過 程 で は , そ の 分 化 に 伴 い 突 起 数 が 増 加 し[ 初 期 前 駆 細 胞 (early progenitor) → 後 期 前 駆 細 胞 (late progenitor)], ミ エ リ ン タ ン パ ク 質 の 発 現 (ミ エ リ ン 形 成 前 オ リ ゴ デ ン ド ロ サ イ ト ,premyelinating oligodendrocyte) 後 ,軸 索 と の 相 互 作 用 を 介 し て 最 終 的 に ミ エ リ ン 形 成 オ リ ゴ デ ン ド ロ サ イ ト (myelinating oligodendrocyt e) へ と 成 熟 す る (Fig. 23). 前 駆 細 胞 の 段 階 で は 分 裂 能 を 有 し , 早 期 に は 移 動 能 も 高 い が , ミ エ リ ン タ ン パ ク 質 が 発 現 す る 段 階 に 至 る と こ れ ら の 能 力 が 失 わ れ て い る .オ リ ゴ デ ン ド ロ サ イ ト 系 譜 の 細 胞 で は , 転 写 因 子 の Olig2 を 発 現 し て お り , 各 分 化 段 階 は 様 々 な マ ー カ ー に よ り 特 徴 づ け ら れ る 4 1 ). PLD4 欠 損 マ ウ ス の ミ エ リ ン 形 成 に お け る 変 化 が オ リ ゴ デ ン ド ロ サ イ ト の 分 化 段 階 に 及 ぼ す 影 響 を 調 べ る た め に 代 表 的 な マ ー カ ー を 用 い て 解 析 し た . ま ず , オ リ ゴ デ ン ド ロ サ イ ト の 数 自 体 に 変 化 が な い か ど う か を 調 べ る た め に ,オ リ ゴ デ ン ド ロ サ イ ト 系 譜 細 胞 マ ー カ ー の 抗 Olig2 抗 体 を 用 い て 深 部 白 質 に お け る Olig2 陽 性 オ リ ゴ デ ン ド ロ サ イ ト の 細 胞 数 を 計 測 し た . そ の 結 果 ,P5 で は 野 生 型 と PLD4 欠 損 マ ウ ス で オ リ ゴ デ ン ド ロ サ イ ト 系 譜 細 胞 数 に 違 い は 見 ら れ な か っ た (Fig. 24A-C). P7 で も 同 様 の 結 果 と な っ た (Fig. 24D-F).
51
Fig. 23 Differentiation of oligodendrocyte lineage cells in vivo and their markers used in this experiment.
Premyelinating Oligodendrocyte Migrating Progenitor Late Progenitor Myelinating Oligodendrocyte Early Progenitor Olig2 NG2 Olig2 NG2 Olig2 NG2 Olig2 MBP CC1 Olig2 MBP CC1
52
Fig. 24 Olig2 expression in oligodendrocytes in cerebellum.
53
Fig. 25 Presence of NG2-positive oligodendrocyte progenitors in folial white matter.
54
Fig. 26 CC1 expression in oligodendrocyte in cerebellum
Representative CC1 staining patterns in P7 cerebella of wild type (A) and PLD4-deficient (B) mice were exhibit ed. Fewer CC1 -positive cells were present in the PLD4-deficient folia, although many CC1 -positive cells were found in the deep cerebellar white matter of both types of mice. Scale bars in A represents 100 µm.
CC1
WT P7
A
B
56
57
Fig. 27 Corpus callosum structures in adult PLD4-deficient mice.
Comparison of corpus callosum structures in wild type (WT; A) and PLD4 -deficient mice (Ho; B). Frozen sections were stained wit h hematoxylin and eosin. No significant anatomical differences in corpus callosum were observed. High magnification images of corpus callosum (black squares) are shown in the lower panels (A’, B’). Scale bars indicate 200 µ m (B) or 100 µ m (B’).
A
B
A’
B’
58
Fig. 28 Demyelination in the corpus callosum of cuprizone-treated adult PLD4-deficient mice.
Frozen coronal sections of corpus callosum obtained from adult wild type (A, C; WT) and PLD4-deficient (B, D; Ho) mice after 5 weeks of nor mal (A, B) or cuprizone (C, D, CPZ) feeding were stained by FluoroMyeli n. A’ to D’ are higher magnification images of the white squares in A to D. Representative i mage from each group (n=3) was shown. No apparent differences in myelin staining were observed bet ween A and B. In C and D, myelin staining was significantly reduce d. Note that almost no stained area was observed in C (* ). Scale bars indicate 200 µm (D) or 100 µm (D’).
59
Fig.29 Relationship between demyelination and microglial activation in the corpus callosum of cuprizone-treated adult PLD4-deficent mice.
Frozen coronal sections of corpus callosum obtained from adult wild type (A, C; WT) and PLD4-deficient (B, D; Ho) mice after 5 weeks of normal (A, B) or cuprizone (C, D: CPZ) feeding were doublei mmunostained by anti-MBP (A-D, red) and anti-Iba1 (A’-D’, green). Nuclei were counterstained with DAPI (blue). A representative Iba1-positive cell indicated by the whit e square in each panel is shown at higher
60
63
66
参 考 文 献
1) Azevedo FA., Carvalho LR., Grinberg LT., Farfel JM., Ferretti RE., Leit e RE., Jacob Filho W., Lent R., Herculano-Houzel S., J Comp Neurol, 513, 532-41 (2009).
2) Mark F. Bear., Barry W. Connors., Michael A. Paradiso., Translation supervised by Kato K., Goto K., Fuji S., Yamazaki Y., “Sinkei Kagaku, ” Nishimura-shot en Co., Ltd., Tokyo, 2007, pp. 19 -42.
3) Haydon PG., Nat Rev Neurosci, 2, 185-93 (2001).
4) Kudo Y., Koizumi S., Wada K., Hashimoto K., Folia Pharmacol Jpm, 130, 185-192 (2007).
5) Tanaka H., Ma J., Tanaka KF., Takao K., Komada M., Tanda K., Suzuki A., Ishibashi T., Baba H., Isa T., Shigemoto R., Ono K., Miyakawa T., Ikenaka K., J
Neurosci, 26, 8363-71 (2009)
6) Ito M., “Nou Shinkei Kagaku, ” Miwa-shoten Co., Ltd., Tokyo., 2003, pp.108-127. 7) Shibata K., Sugawara T., Fujishita K., Shinozaki Y., Matsukawa T., Suzuki T.,
Koizumi S., PLoS One, 6, e23510 (2011).
8) Mikoshiba K., Shimizu T., “Nounohassei・bunka・kasosei,” Kyoritsu-shuppan Co., Ltd., Tokyo, 2002, pp. 67 -111
9) Tuohy VK., Lu Z., Sobel RA., Laursen RA., Lees MB., J Immunol, 142, 1523-7 (1989).
10) Mendel I1., Kerlero de Rosbo N., Ben-Nun A., Eur J Immunol, 25, 1951-9 (1995) 11) Nimmerjahn A., Kirchhoff F., Helmchen F., Science, 308, 1314-8 (2005).
12) Pont-Lezica L., Béchade C., Belarif-Cantaut Y., Pascual O., Bessis A., J
67
13) Marco Prinz & Josef Priller., Nat Rev Neurosci, 15, 300-12 (2014).
14) Suzuki K., Sugihara G., Ouchi Y., Nakamura K., Futatsubashi M., Takebayashi K., Yoshihara Y., Omata K., Matsumoto K., Tsuchiya KJ., Iwata Y., Tsujii M., Sugiya ma T., Mori N., JAMA Psychiatry, 70, 49-58 (2013).
15) Dudvarski Stankovic N., Teodorczyk M., Ploen R., Zipp F., Schmidt MH., Acta
Neuropathol, 131, 347-63 (2016).
16) Yoshikawa F., Banno Y., Otani Y., Yamaguchi Y., Nagakura -Takagi Y., Morita N., Sato Y., Saruta C., Nishibe H., Sadakata T., Shinoda Y., Hayashi K., Mishima Y. , Baba H., Furuichi T., PLoS One, 5, e13932 (2010).
17) Gomez-Cambronero J., Di Fulvio M., Knapek K., J Leukoc Biol, 82, 272-81 (2007).
18) Pedersen KM., Finsen B., Celis JE., Jensen NA., J Biol Chem, 273, 31494-504 (1998).
19) van der Lee SJ., Holstege H., Wong TH., Jakobsdottir J., Bis JC., Chouraki V., van Rooij JG., Grove ML., Smith AV., Amin N., Choi SH., Beiser AS., Garcia ME., van IJcken WF., Pijnenburg YA., Louwersheimer E., Brouwer RW., van den Hout MC., Oole E., Eirkisdottir G., Levy D., Rotter JI., Emilsson V., O'Donnell CJ., Aspelund T., Uitterlinden AG., Launer LJ., Hofman A., Boerwinkle E., Psaty BM., DeStefano AL., Schelt ens P., Seshadri S., van Swieten JC., Gudnason V., van der Flier WM., Ikram MA., van Duijn CM., Nature, 520, E2-3 (2015).
20) Choi SY., Huang P., Jenkins GM., Chan DC., Schiller J., Frohman MA., Nat Cell
Biol, 8, 1255-62 (2006).
21) Gao Q., Frohman MA., BMB Rep, 45, 7-13 (2012)
22) Otani Y., Ya maguchi Y., Sato Y., Furuichi T., Ikenaka K., Kitani H., Baba H.,
PLoS One, 6, e27544 (2011).
68
Furukawa H., Kochi Y., Ota Y., Ikari K., Sato S., Tohma S., Ya mada R., Yama moto K., Kubo M., Ya manaka H., Kuwana M., Tsuchiya N., Matsuda F., Mimori T., Arthritis Rheum, 65, 472-80 (2013).
24) Jung S., Pausch H., Langenmayer MC., Schwarzenbacher H., Majzoub-Altwec k M., Gollnick NS., Fries R., BMC Genomics, 22, 623 (2014).
25) Tanaka KF., Ahmari SE., Leonardo ED., Richardson-Jones JW., Budreck EC., Scheiffele P., Sugio S., Inamura N., Ikenaka K., Hen R., Biol Psychiatry, 67, 770-3, (2010).
26) Otani Y., Dr. Thesis, (2012).
27) Eric R. Kandel., James H. Schwartz., Thomas M. Jessell., Stev en A. Siegelbaum., A.J. Hudspeth., “Principles of neural science, ” Fifth ed., McGraw-Hill, 2013, chapter forty two.
28) Sidman RL., Rakic P., Brain Res, 62, 1-35, (1973).
29) Ya mada K., Watanabe M., Anat Sci Int, 2, 94-108, (2002).
30) Peferoen LA., Gerritsen WH., Breur M., Ummenthum KM., Peferoen-Baert RM., van der Valk P., van Noort JM., Amor S., Acta Neuropathol Commun, 3, 87, (2015).
31) Sinha S., Boyden AW., Itani FR., Crawford MP., Karandikar NJ., 10, 619, (2015).
32) Kariya Y., Kariya Y., Saito T., Nishiyama S., Honda T., Tanaka K., Yoshida M., Fujihara K., Hashimoto Y., BBA Clin, 3, 126-34 (2015).
33) Tanaka T., Yoshida S., Biomol Concepts, 5, 289-98, (2014).
34) Praet J., Guglielmetti C., Berneman Z., Van der Linden A., Ponsaerts P., Neurosci Biobehav Rev, 47, 485-505, (2014).
69
Padovani-Claudio D., He T., Choi K., Lane TE., Miller RH., Ransohoff RM., Nat Neurosci, 13, 319-26, (2010).
36) Matsushima GK., Morell P., Brain Pathol, 11, 107-16, (2001).
37) Crain JM., Nikodemova M., Watters JJ., J Neurosci Res, 91, 1143-51 (2013). 38) Liao B., Zhao W., Beers DR., Henkel JS., Appel SH., Exp Neurol, 237, 147-52
(2012).
39) Miron VE., Franklin RJ., J Neurochem, 130, 165-71, (2014).
40) Pont-Lezica L., Béchade C., Belarif-Cantaut Y., Pascual O., Bessis A., J
Neurochem, 119, 901-8, (2011).
41) Brnce D. Trapp., Steven E. Pfeiffer., Mihaela Anit ei., Graha me J. Kidd., “MYELIN BIOLOGY and DISORDERS ”, San Diego, California, 2004, chapter two.
42) Ahrendsen JT., Macklin W., Neurosci Bull, 29, 199-215, (2013). 43) Nave KA., Werner HB., Annu Rev Cell Dev Biol, 30, 503-33, (2014).
44) Shigemoto-Mogami Y., Hoshikawa K., Goldman JE., Sekino Y., Sato K., J Neurosci, 34, 2231-43, (2014).
45) Taveggia C., Feltri ML., Wrabetz L., Nat Rev Neurol, 6, 276-87, (2010). 46) Holness CL., Simmons DL., Blood, 6, 1607-13, (1993)
47) Matsumoto H., Kumon Y., Watanabe H., Ohnishi T., Shudou M., Ii C., Takahashi H., Imai Y., Tanaka J., J Neurosci Res, 5, 994-1009, (2007).
48) Graeber MB., Streit WJ., Kiefer R., Schoen SW., Kreutzberg GW., J Neuroimmunol, 27, 121-32, (1990).
70
Neuroinflammation, 12, 114, (2015).
50) Appel SH., Zhao W., Beers DR., Henkel JS., Acta Myol, 1, 4-8, (2011).
51) Mikita J., Dubourdieu-Cassagno N., Deloire MS., Vekris A., Biran M., Raffar d G., Brochet B., Canron MH., Franconi JM., Boiziau C., Petry KG., Mult Scler, 1, 2-15, (2011).
52) Butovsky O., Landa G., Kunis G., Ziv Y., Avidan H., Greenberg N., Schwartz A., Smirnov I., Pollack A., Jung S., Schwartz M., J Clin Invest, 4, 905-15, (2006). 53) Miron VE., Boyd A., Zhao JW., Yuen TJ., Ruckh JM., Shadrach JL., van
Wijngaarden P., Wagers AJ., Willia ms A., Franklin RJ., ffrench-Constant C., Nat Neurosci, 9, 1211-8, (2013).