熊本大学学位論文
水チャネル aquaporin の機能調節機構に関する研究
2007 長井 一史
Study of Functional Regulation of Water Channel,
Study of Functional Regulation of Water Channel, Aquaporin Department of Chemico-pharmacological Sciences,
Graduate School of Pharmaceutical Sciences
Kazufumi Nagai
Aquaporins (AQPs) are a family of water channel proteins that allow the rapid movement of water in response to osmotic gradients. To date, 13 homologous members of the AQP family have been identified and they are expressed in various organs.
Phenotype studies of transgenic AQP knockout mouse models have demonstrated essential roles for AQP in mammalian physiology and pathophysiology. For instance, mice lacking AQPs that are expressed in kidney are polyuric because they were unable to concentrate their urine. AQP5 is expressed in exocrine glands and alveolar type I cells.
AQP5-deficient mice exhibit reduced secretion of saliva, sweat and airway fluids. AQP4 is expressed in astrocyte endfeet in the brain. AQP4-deficient mice show reduced traumatic brain injury-induced cerebral edema. Consequently, these studies have been suggested that AQPs are potential targets for new diuretic reagents and novel drugs for disorders that involve water movement, such as brain edema and abnormal airway secretion. However, the functional regulation of AQP has not been fully understood and the clinical modulators of AQP have not been identified. Therefore, in this study, to find new modulators of AQPs, I examined the effects of kampo medicine, various metals and nitric oxide on AQP activity.
First, I focused on Goreisan that is one of Japanese Kampo medicine to treat patients with body water imbalance. I examined the effect of Goreisan on membrane water permeability in mammalian cells and aquaporin (AQP) water channel activity. Goreisan and its component Atractylodis lanceae rhizoma (ALR) decreased membrane water permeability without changing the membrane potential. ALR decreased water permeability in AQP5-containing proteoliposomes, but did not do so in control liposomes.
Therefore, it is likely that ALR reduced membrane water permeability by the direct
inhibition of AQP5. In addition, an 80% ethanol-insoluble fraction of ALR showed the
results suggest that the bivalent metal ions contained in Goreisan and ALR decrease cell membrane water permeability via inhibition of AQP5, and this effect might contribute to the pharmacological effects of Goreisan.
Next, I examined the effect of various metal ions (Li+, Ca2\ Mn2+, Co2+, Ni2+, Cu2+, Zn2+, Ag+ and Hg2+) on water permeability of AQP1-5 in Xenopus oocytes expressing AQP1-5.
In result, many metal ions showed isoform-selective inhibition of AQPs. Hg2+ and Ag+
selectively inhibited AQP1 and AQP2, Mn2+, Ni2+ and Zn2+ inhibited AQP4 and AQP5.
AQP3 showed wide sensitivity for metal ions. Mn2+, Zn2+ and Ni2+ inhibited AQP4 and four extracellular amino acids, Glu63, Asp69, His151 and Glu228 were identified as responsible sites for AQP4 inhibitors. Furthermore, Cys183 was identified as responsible site for inhibitory effect of Mn2+ and Hg2+ on AQP5. These results indicated that metal ions were very useful tools for investigations of the regulation of AQP water permeability.
Finally, I investigated the effect of Nitric oxide (NO) on functional regulation of AQP5.
NO donors decreased water permeability of AQPS in Xenopus oocyte expression system
and induced S-nitrosylation of AQP5 at Cys6 and CysMr\ In addition, NO donors
significantly reduced the expression of AQP5 at plasma membrane and membrane water
permeability in mouse lung epithelial cells (MLE-12). Cell surface biotinylation and
immunofluorescence strongly suggested that NO changes the distribution of AQPS from
plasma membrane to intracellular fraction. This reduction of AQP5 was inhibited by
simultaneous treatment of methyl-beta-cyclodextrin but not by ODQ. These results
suggested that NO decreases AQP5 activity through both S-nitrosylation and
internalization of AQP5.
目次
第 1 章 緒論... 1
第2章 AQP を介した細胞膜水透過性に対する五苓散の作用についての検討... 5
2.1.本章の目的... 5
2.2.五苓散および五苓散構成生薬の細胞膜水透過性に対する作用 ... 6
1) 五苓散の細胞膜水透過性に対する作用 ... 6
2) 五苓散構成生薬の細胞膜水透過性に対する作用 ... 8
3) 五苓散および蒼朮の細胞膜電位に対する作用... 10
4) 蒼朮の AQP5 プロテオリポソーム水透過性に対する作用... 11
5) 蒼朮分画エキスの細胞膜水透過性に対する作用 ... 12
6) 蒼朮 80%エタノール不溶性分画の細胞膜水透過性に対する作用 ... 14
2.3.考察 ... 15
第 3 章 金属イオンの AQP 阻害作用についての検討 ... 17
3.1.本章の目的... 17
3.2.種々の金属イオンの AQP 水透過性に対する作用 ... 18
3.3.ほ乳類細胞の細胞膜水透過性に対する金属イオンの作用... 20
3.4.AQP 点変異体に対する金属イオンの作用... 23
1) AQP4 点変異体に対する Mn2+,Zn2+および Ni2+の作用... 24
2) AQP5 点変異体に対する Hg2+および Mn2+の作用 ... 27
3.5.考察 ... 29
4.2.AQP5 のニトロソ化についての検討 ... 34
1) MLE-12 細胞における AQP5 のニトロソ化についての検討 ... 34
2) AQP5 システイン変異体におけるニトロソ化の検討 ... 36
4.3. AQP5 水透過性に対する NO の作用 ... 38
1) AQP5 水透過性に対する NO ドナーの作用... 38
2) NO の AQP5 水透過性抑制機構における cGMPpathway の関与 ... 40
4.4.考察 ... 41
第 5 章 細胞膜上 AQP5 の発現量に対する NO の作用... 44
5.1. 本章の目的... 44
5.2. AQP5 の細胞膜上発現量に対する NO の作用 ... 45
1) NO の細胞膜上 AQP5 発現量に対する作用... 45
2) NO の細胞膜上 AQP5 発現抑制作用のメカニズムについての検討 ... 47
3) NO による AQP5 の局在変化についての検討 ... 49
4) NO の細胞膜水透過性に対する作用 ... 51
5.3.考察 ... 53
第 6 章 総括... 56
第 1 章 緒論
すべての生命体にとって,水は必要不可欠な物質である.水は生体を構成する物質 の中で最も多く含まれており,生命現象を司る化学反応の場を提供し,また水そのも のが種々の化学反応の基質となっている.水は飲料や食物として摂取され,体内に分 布した後に最終的には尿や汗として体外へ排出される.生体内の水は細胞内,細胞間 隙,血液の 3 つの領域に存在し,それらの領域間を静水圧および浸透圧勾配に従って 常にダイナミックに移動して動的平衡を保っている.しかし,この平衡が崩れ,細胞 内,細胞間隙および皮下組織に水が過剰に貯留すると浮腫を生じる.臨床的に浮腫を 生じる原因は多数存在し,例えば,腎機能障害による尿生成異常を初めとして,肝硬 変時の腹水の貯留,頭部損傷や脳梗塞による脳浮腫の形成,急性肺傷害や心不全によ る肺水腫の形成などほとんどすべての臨床領域に及んでいる.特に,脳浮腫や肺水腫 の形成はそれぞれ頭蓋内圧の亢進による神経組織の圧迫や,呼吸不全などを引き起こ す致死的な疾患であり,患者の予後を決定する上で非常に重要である.従来,脳浮腫 に対する薬物療法は,マンニトールおよびグリセロール溶液などの高浸透圧溶液や,
利尿薬を用いて腎臓における尿生成量を増加させ,余剰な水分を排出させることを目 的として行われてきた.また,肺水腫に対する治療も強心薬や利尿薬が用いられてい る.しかしながら,高浸透圧溶液による治療効果は十分なものではなく,科学的根拠 に乏しいことが指摘されている.また,利尿薬による薬物療法は尿量の増加により全 身の水分量を減少させるため,高ナトリウム血症や低カリウム血症および心不全など の副作用が問題となっている.このような背景から,現行の薬物療法に加えて,浮腫 に対する新たな治療薬が求められている.しかしながら,浮腫に対する薬物療法はこ の数十年間大きな進歩は見られていない.その原因の1つとして浮腫治療に対する明 確な分子標的が無いことが挙げられる.
従来,水分子は浸透圧勾配に従って脂質二重層を単純拡散すると考えられていた.
しかし,1992 年に Agre らが赤血球膜から細胞膜の水輸送を促進するタンパク質と
れている2.ヒトでは各 AQP アイソフォームは全身に広く分布しており,組織特異的 な発現パターンを示している3.AQP は 6 回膜貫通型のタンパク質であり,細胞膜上 で 4 量体を形成して存在し,各単体が水を透過させる孔(ポア)をその構造に含んで いる4.そのポア構造の中心部分にはアスパラギン(N),プロリン(P),アラニン(A)の 3 つのアミノ酸からなる NPA ドメインを持ち,これは AQP ファミリーに保存された 領域である.AQP ファミリーは大きく2つのグループに分けられ,1 つは水分子を 選択的に透過させるグループで AQP1,2,4,5,8 が含まれる.もう 1 つのグループは 水分子以外にもグリセロールや尿素など他の小分子も透過させるグループで AQP3, 7,9,10 が含まれる5-8.その他のアイソフォームに関しては未だその選択性が明らか になっていない.AQP 欠損マウスの表現系解析や,AQP 遺伝子の変異による疾患が 解明されるにつれて,AQP の生理学的および病態生理学的な役割が明らかになりつ つある.例えば,腎臓に発現している AQP(AQP1,2,3 および 4)を欠損したマウス では尿濃縮障害による多尿が認められ9-12,特に AQP2 遺伝子はヒトにおいても変異 が見つかっており,腎性尿崩症の原因遺伝子であることが知られている13-15.また,
AQP4 は脳に多く発現しており16-18,AQP4 欠損マウスは中大脳動脈結紮および急性 水中毒により惹起される脳浮腫が軽減し,死亡率を改善することも報告されている19. さらに,AQP5 は唾液腺,汗腺,涙腺,気道粘膜下腺および肺胞上皮 I 型細胞など腺 組織に発現しており,AQP5 欠損マウスではそれら腺組織における水分泌が低下し,
唾液,汗,涙の分泌不全となる20-23.また,肺胞-血管間の水透過性が 10 分の 1 に低 下する24.さらに,シェーグレン症候群の患者の腺組織では AQP5 の異常な局在が認 められており,それが唾液や涙の分泌不全の原因ではないかと考えられている25-27.
これらの報告は AQP が生体内の水分代謝に重要な役割を果たしていることを示して おり,加えて AQP が種々の水分代謝異常に対する治療薬の創薬ターゲットになる可 能性を支持している.例えば,腎臓に発現している AQP を阻害する物質は電解質バ ランスに影響の少ない利尿薬となり得る.また AQP4 を阻害する物質は脳浮腫に対 しての予防薬および治療薬となることが考えられる.また,AQP5 の機能を亢進する 物質はドライアイやドライマウスに対する治療薬となる可能性がある.さらに,上記 の AQP 欠損マウスには致死的な生理学的および発生学的異常が認められないため,
AQP 阻害薬には重篤な副作用がないと考えられる.これらのことから種々の水分代 謝異常に対する治療薬として AQP 機能調節薬の開発が望まれている.
今までAQP機能調節物質に関して得られている知見として,水銀化合物を初めとし た金属イオンがAQPの水透過性を阻害することが知られている.水銀化合物はAQP1 分子内のポア構造近傍に存在するシステイン残基に共有結合することでAQP1の水透 過性を抑制する28,29.銀および金化合物もAQP1を阻害することが報告されている30,31. さらに,銅およびニッケルがAQP3を阻害することが知られており,その阻害作用に はAQP3の細胞外ループに存在するヒスチジン,セリンおよびトリプトファン残基が 関与することも報告されている32,33.これらの報告は,各AQPアイソフォームが異な った金属イオン感受性を持つことを示しており,それは各AQPアイソフォームが水透 過性を維持するための独自の構造を持つことを示唆している.AQP機能調節物質に関 する知見としては金属イオンの他にもtetraethylammonium,acetazolamideおよび dimethylsulfoxideなどがAQPを阻害するとの報告もなされたが34-38,その後,その 結果を覆す報告もなされており31,現在までに確立されたAQP機能調節物質は上記の 金属イオンのみである.これら金属イオンは生体に対して様々な反応性を持つことや,
その毒性の高さから医薬品への応用は難しいが,AQPの水透過性阻害機構を調べる上 での有用なツールになると考えられる.すなわち,各金属イオンのAQP阻害機構を詳 細に解明することで,AQPアイソフォーム選択的な阻害薬の開発につながると考えら れる.また,AQPをターゲットとした創薬においては,医薬品への応用を目指した新 規AQP機能調節物質の探索とAQPの水透過性調節機構の解明の両面からのアプローチ
このような背景のもと,本研究では,AQP機能調節薬の開発を究極的な目的として 実験を行なった.第2章では,新規AQP機能調節物質を見出すことを目的として,漢 方方剤の1つである五苓散を出発物質として探索を行なった.第3章では金属イオンに よるAQPの水透過性阻害機構についての知見を得ることを目的として,既知のAQP阻 害剤を含めた一価および二価の陽イオンを中心に,AQP1˜5の各アイソフォームに対 する阻害作用を検討した.第4章および第5章では内因性のAQP機能調節機構を解明す ることを目的として,Nitricoxide(NO)によるAQP機能調節機構に関して検討した.
以下に得られた知見の詳細を記す.
第2章 AQP を介した細胞膜水透過性に対する五苓散の作用についての検討
2.1.本章の目的
AQP 機能調節物質が種々の水分代謝異常を伴うような疾患に対しての治療・予防 薬となり得ることは前章においてすでに述べた.AQP 阻害物質として最もよく知ら れるものに水銀化合物がある.水銀化合物は 20 世紀の初めまで実際に利尿薬として 浮腫治療などに用いられてきたが,その作用には腎臓に発現する AQP の阻害が寄与 していた可能性は十分に考えられる.しかし,水銀化合物は非特異的な作用も多く持 っており,その副作用の強さから現在は利尿薬としては用いられていない.従って,
医薬品として応用可能な AQP 機能調節物質は見出されていないのが現状である.
五苓散は 5 種類の生薬から構成される漢方方剤であり,日本では水分代謝異常を伴 う疾患に対して使用されてきた.五苓散についての臨床的データおよび基礎的データ には興味深い結果が含まれている.第1に,ヒトに五苓散を投与し,尿量の変化を測 定すると,代表的な西洋利尿薬であるフロセミドと同程度の尿量増加が認められた.
さらに興味深いことに,五苓散は浮腫状態においては尿量を増加させ,逆に脱水状態 では尿量を減少させて体内に水分を保持するように作用した.第2に,ヒトに五苓散 を投与するとカリウムおよびナトリウムの排泄傾向は増加するものの,血漿カリウム 濃度の減少傾向は西洋利尿薬に比べて弱く,血漿電解質濃度はコントロール群との間 に有意な差を認めない.これらの報告は五苓散の利尿作用の機序が既存の西洋医学的 な利尿作用とは異なり,電解質の再吸収阻害に依存していないことを示唆している.
そこで本章では五苓散および五苓散を構成する生薬が AQP 機能調節物質を含んで いるのではないかという仮説のもとに,五苓散および五苓散に含まれる生薬の細胞膜 水透過性および AQP5 の水透過性に対する作用を検討した.
2.2.五苓散および五苓散構成生薬の細胞膜水透過性に対する作用
1) 五苓散の細胞膜水透過性に対する作用
AQP 機能調節物質を探索するにあたって,五苓散に注目した.五苓散が MLE-12 細胞の細胞膜水透過性に影響を与えるか否かについて stopped-flow 法を用いて検討 した.Stopped-flow 法はほ乳類細胞の細胞膜水透過性を測定する方法の1つであり,
従来から,AQP の細胞膜水透過性亢進作用や,AQP 阻害物質の阻害作用を調べるた めにも用いられてきた39-42.本実験の概要を簡単に記す.細胞懸濁液と高浸透圧溶液
(600mM マンニトールを含んだ緩衝液)を stopped-flow 装置にて急速に混合し,
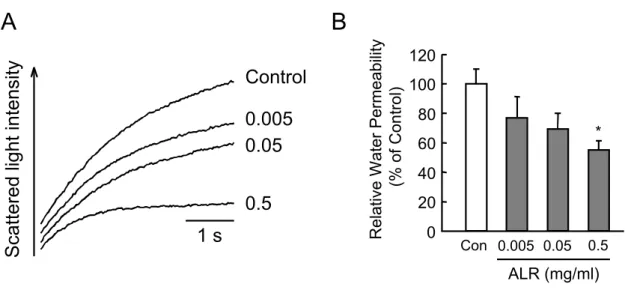
細胞に高浸透圧負荷を与えると細胞内から水が流出し,細胞体積は減少する.細胞体 積の減少に伴って細胞からの 90 度方向への散乱光は増加し,その散乱光強度の初期 変化速度から細胞膜の水透過性を算出した.MLE-12 細胞に五苓散を 0.3,1.0 およ び 3.0mg/ml の濃度で 2 分間処理し,その後,細胞膜水透過性を測定した.五苓散 は濃度依存的に MLE-12 細胞の細胞膜水透過性を抑制し(Fig.1A),1.0mg/ml の 五苓散は細胞膜水透過性をコントロールの約 50%に抑制した(Fig.1B).これは代 表的な AQP 阻害剤である HgCl2
(0.5mM)を 2 分間処理した場合とほぼ同程度の抑 制効果であった.また,五苓散の処理時間を 10,20,30 分と変えて細胞膜水透過性の 抑制作用を検討したが,2 分間処理時と比べて顕著な変化は得られなかった(データ は示さず).さらに五苓散そのものは溶液の散乱光強度,浸透圧,pH には影響を与え なかった(データは示さず).Figure 1. Effect of Goreisan on membrane water permeability in MLE-12 cells.
MLE-12 cells were subjected to a 300 mOsm inwardly directed osmotic gradient at 20°C Scattered light intensity was measured by stopped-flow apparatus. (A) MLE-12 cells were incubated with each concentrations of Goreisan (0.3-3.0 mg/ml) for 2min or HgCl
2(0.5 m M ) f o r 2 m i n b e f o r e m e a s u r e m e n t o f m e m b r a n e w a t e r p e r m e a b i l i t y b y stopped-flow light scattering method. (B) Relative water permeability for experiments as in (A). The data are means ± SD from six to eight experiments. * indicates a significant difference from the control group.
Scattered light intensity 1 s
Control 0.3 1.0 3.0
0 20 40 60 80 100 120
Con 0.3 1.0 3.0 HgCl
2Relative water permeability (% of control)
*
*
**
*
Goreisan (mg/ml)
2) 五苓散構成生薬の細胞膜水透過性に対する作用
五苓散は沢冩(AlismatisRhizoma),桂皮(CinnamomiCortex),猪苓(Polyporus),
蒼朮(AtractylodisLanceaeRhizoma(ALR))および茯苓(Hoelen)の5種類の生薬 から構成される漢方方剤である.次に,これら構成生薬のうち,どの生薬が細胞膜水 透過性抑制作用を持つのかを明らかにするために,各生薬の水抽出エキスの作用につ いて検討した.各生薬は全て最終濃度が0.5mg/mlになるようにMLE-12細胞懸濁液 に加え,2分間,20℃で処理した後,stopped-flow法により細胞膜水透過性抑制作用 を調べた(Table2).その結果,蒼朮(ALR)が最も強い抑制作用(コントロールの 53%に抑制)を示し,次いで猪苓(polyporus)および茯苓(Hoelen)にも抑制作用が 認められた.しかしながら,茯苓処理により細胞の凝集体が形成されることが顕微鏡 下で確認された(データは示さず).また,各生薬エキスの細胞毒性を細胞外に放出さ れたlactatedehydrogenase(LDH)活性を指標に測定すると,猪苓(>0.5mg/ml)に は細胞毒性が認められたが,蒼朮については認められなかった(データは示さず).ま た,蒼朮は濃度依存的な細胞膜水透過性抑制抑制作用を示した(Fig.2)ことから,五 苓散構成生薬のうちAQP阻害物質の探索には蒼朮が最も適していると考えられた.
Herbal extract Inhibitory effect (%)
Cinnamomi cortex 17.1±4.8
Hoelen 30.3±6.7
Alismatis rhizoma 21.8±4.2
Polyporus 50.7±15.2 *
1 s
Control 0.005 0.05
Scattered light intensity
0.5
0 20 40 60 80 100 120
Con 0.005 0.05 0.5 ALR (mg/ml) Relative Water Permeability (% of Control)
*
A B
Figure 2. Effect of ALR extract on membrane water permeability in MLE-12 cells.
(A) MlE-12 cells were incubated with ALR extract at indicated concentration for 2 min before
measurement of membrane water permeability by stopped-flow light scattering method. (B)
Relative water permeability for experiments as in (A). The data are means ± SD from six to
eight experiments. * indicates a significant difference from the control group.
3) 五苓散および蒼朮の細胞膜電位に対する作用
前項の結果より,五苓散および蒼朮(ALR)に細胞膜水透過性抑制作用があること が見出された.細胞膜を介した水の移動は様々なイオンチャネル・トランスポーター のイオン輸送に伴った浸透圧勾配により生じる.しかしながら,本実験においては五 苓散あるいは蒼朮がこれらのイオン輸送機序を活性化し,二次的に細胞膜水透過性に 影響している可能性は排除できていない.そこで五苓散および蒼朮が細胞膜を介した イオン輸送に影響するか否かについてパッチクランプ法により細胞膜電位を測定する ことによって評価した.MLE-12 細胞の表面にパッチ電極を軽く接触させてナイスタ チン穿孔法により膜電位を記録しながら細胞外液を 1mg/ml 五苓散あるいは 0.5 mg/ml 蒼朮を含有するものに置換した.Fig.3 に示すように五苓散および蒼朮は細 胞膜電位に著明な変化を生じなかった.従って,五苓散および蒼朮による細胞膜水透 過性抑制作用がイオン輸送の活性化によるものではないことが示唆された.
-40mv
20 mv
Goreisan
ALR
-40mv
4) 蒼朮のAQP5プロテオリポソーム水透過性に対する作用
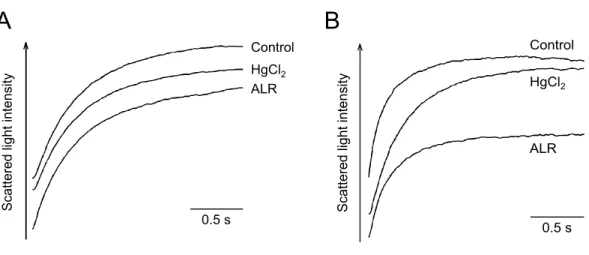
蒼朮が AQP5 の水透過性を直接抑制するか否かを検討するため,AQP5 含有プロ テオリポソームの水透過性に対する作用を stopped-flow 法により測定した.プロテ オリポソームと同様の方法を用いて作成した AQP5 を含まないコントロールリポソ ームについても比較検討を行なった.まず AQP5 含有プロテオリポソームについて 水銀感受性があるか否かについてコントロールリポソームと比較検討を行なった結果,
HgCl2は AQP5 含有プロテオリポソームの水透過性を抑制したが,コントロールリ ポソームの水透過性は抑制しなかった(Fig.4).蒼朮についても同様に検討したとこ ろ,蒼朮も AQP5 プロテオリポソームの水透過性を抑制したが,コントロールリポ ソームの水透過性は抑制しなかった(Fig.4).このことから,蒼朮は AQP5 に直接阻 害作用を示して細胞膜水透過性を抑制する可能性が示唆された.
Scattered light intensity
Control
HgCl
2ALR
0.5 s Scattered light intensity
0.5 s Control
HgCl
2ALR
A B
Figure 4. Effect of ALR and HgCl
2on membrane water permeability in control liposomes and AQP5-containing proteoliposomes.
Control liposomes (A) and AQP5 containing-proteoliposomes (B) were incubated with ALR (0.5
mg/ml) or HgCl
2(1 mM) for 2 min before measurement of membrane water permeability by
stopped-flow light scattering method.
5) 蒼朮分画エキスの細胞膜水透過性に対する作用
次に,蒼朮のどの分画に細胞膜水透過性抑制作用があるのか調べた.蒼朮水抽出エ キスをacetoneextract(A),MeOHextract(M),50%MeOHextract(50M),water extract(W),80%EtOHprecipitatefraction(P)の5つに分画し,各分画の細胞膜 水透過性に対する作用を stopped-flow 法にて検討した.各分画の処理濃度は回収率 から,蒼朮水抽出エキス 1.0mg/ml に相当する濃度になるように PBS を溶媒に用い て調整し(acetoneextract は DMSO にて調整した),2 分間,20℃で処理した.Fig.
5A より,Pfraction に最も強い細胞膜水透過性抑制作用が認められ,他の分画には 抑制作用はほとんど認められなかった.Pfractionの細胞膜水透過性抑制作用につ いてさらに詳しく検討した.まず,0.003,0.03,0.3mg/mlPfraction を用いて濃 度依存性について調べたが,Pfraction は濃度依存的に細胞膜水透過性を抑制した (Fig.5B).0.3mg/mlPfraction により細胞膜水透過性はコントロールの約 50%に まで低下した.Pfraction に含まれる疎水性成分ついて HPLC により同定を試みた が,Pfraction にはほとんど何も検出されなかった(データは示さず).
Figure 5. Effects of ALR extract fractions on membrane water permeability in MLE-12 cells.
Relative water permeability was calculated from the time course of changing scattered light intensity measured by stopped-flow light scattering method. (A) Cells were treated with ALR acetone-eluted (0.01 mg/ml dissolved in DMSO; A), methanol-eluted (0.03 mg/ml;
M), 50% methanol-eluted (0.04 mg/ml; 50M), water-eluted (0.5 mg/ml; W) and 80%
ethanol-insoluble (0.3 mg/ml; P) fractions for 2 min, after which membrane water permeability was measured. (B) Cells were treated with 0.003, 0.03 or 0.3 mg/ml ALR 80%
ethanol-insoluble fraction for 2 min. The data are means ± SE from six to eight experiments.
* indicates a significant difference from the control group.
0 20 40 60 80 100 120
Con M 50M W P Relative water permeability (% of Control)
A
*
0 20 40 60 80 100 120
Con 0.003 0.03 0.3 Relative water permeability (% of Control)
P fraction (mg/ml)
*
*
A B
6) 蒼朮80%エタノール不溶性分画の細胞膜水透過性に対する作用
Pfraction は極めて親水性の高い分画であり,植物由来の成分であることから金属 イオンが豊富に含まれる可能性が考えられた.また,過去の報告においても,水銀,
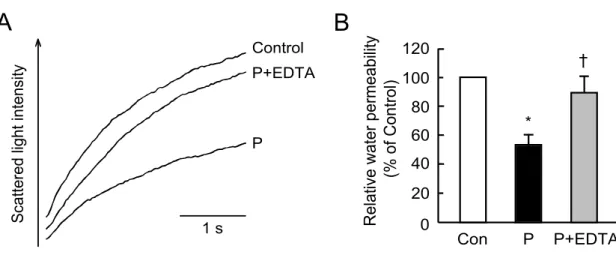
金,銀およびニッケルなどの金属イオンが AQP を阻害することが示されている.そ こで,Pfraction の細胞膜水透過性抑制作用に金属イオンが関与するか否か検討する ために,金属キレート剤である EDTA 存在下における Pfraction の細胞膜水透過性 抑制作用について検討した.その結果,EDTA 存在下において Pfraction の細胞膜 水透過性抑制作用は消失した(Fig.6).このことから,Pfraction に含まれる細胞膜 水透過性抑制作用を持つ物質が二価の金属イオンであることが示唆された.
Scattered light intensity
1 s
Control P+EDTA
P
0 20 40 60 80 100 120
Con P P+EDTA Relative water permeability (% of Control)
†
*
A B
Figure 6. Effect of EDTA on decrease in membrane water permeability induced
2.3.考察
1.五苓散の細胞膜水透過性抑制作用について
本章においてまず,五苓散が濃度依存的に MLE-12 細胞の細胞膜水透過性を抑制 することが明らかになった(Fig.1).また,五苓散は 2 分間という短い処理時間で抑 制作用を現し,その処理時間を 10~30 分に変えても抑制作用は同程度であった(デ ータは示さず).これは水銀化合物による AQP 阻害作用とよく似たものであった.さ らに五苓散を構成する 5 つの生薬エキスについても同様に細胞膜水透過性に対する作 用を比較検討したところ,蒼朮に最も強い抑制作用が認められ,それは濃度依存的で あった(Table2)(Fig.2).猪苓,茯苓にも抑制作用が認められたが,猪苓には高濃 度で細胞毒性が認められたこと(データは示さず),また,茯苓は顕微鏡下で細胞凝 集作用が確認されたこと(データは示さず)から,AQP 機能調節物質の探索には蒼朮 が最も適していると判断した.五苓散および蒼朮は細胞膜電位に対して影響を与えな かったこと(Fig.3),また,コントロールリポソームの水透過性には影響せず,AQP5 を含むリポソームの水透過性を抑制したこと(Fig.4)から,蒼朮は AQP5 の水透過 性を直接的に阻害している可能性が高いと考えられた.さらに蒼朮に含まれる活性成 分を同定するために蒼朮水抽出エキスを 5 つに分画し,細胞膜水透過性に対する作用 を比較検討したが,抑制作用は主として蒼朮 80%エタノール不溶性分画(Pfraction) に認められた(Fig.5).Pfraction は 5 つの分画の中で最も親水性の高い分画であり,
また HPLC によっても Pfraction にはほとんど有機化合物が検出されなかったこと (データは示さず),また,Pfraction の細胞膜水透過性抑制作用は EDTA 存在下で消 失したこと(Fig.6)から,本分画の細胞膜水透過性抑制作用は金属イオンによるも のであることが考えられた.
2.金属イオンによる AQP 阻害作用について
金属イオンの AQP 阻害作用については今まで水銀化合物を初めとして銀,金,銅 およびニッケルを含む化合物が知られている28,30,32,33.銀および金については植物由 来の小胞膜(植物 AQP を含む)およびヒト由来赤血球の細胞膜水透過性を抑制する ことが示されており,これはおそらく植物 AQP およびヒト AQP1 を阻害することに よるものと考えられている.また,銅およびニッケルは AQP3 を特異的に阻害する ことが示されている.本実験において用いた Pfraction に含まれる金属イオンを ICP-MS によって分析したところ,マンガン,コバルトおよびクロムが含まれている ことが明らかになった.しかしながら,これらの金属イオンについては未だ AQP 阻 害に関する知見は得られておらず,また,水銀化合物以外に AQP5 を特異的に阻害 する金属イオンも見出されてはいない.過去の報告と今回の知見を合わせて考えると,
多くの金属イオンが AQP アイソフォーム選択的な阻害作用を持っている可能性が考 えられる.今後,各 AQP アイソフォームに対する金属イオンの阻害作用のプロファ イルを検討することが必要であると考えられる.本実験の結果より金属イオンが AQP を阻害することで,五苓散の薬理作用の一部を担っている可能性が考えられた.今ま で,生薬および天然物の薬理作用を考える場合,その活性成分は主として有機化合物 に目が向けられてきた.しかし,本章の結果は漢方方剤または生薬エキスに含まれる 無機成分が薬理作用の一部を担う可能性があることを示唆している.また,利水作用 を持つといわれる漢方方剤には蒼朮を含むものが多く,AQP 阻害作用を持つ金属イ オンが利水作用に寄与している可能性が考えられる.今後,各漢方方剤に含まれる金 属成分とその作用を比較検討することが必要かもしれない.
第 3 章 金属イオンの AQP 阻害作用についての検討
3.1.本章の目的
AQP はその分子内に水分子を透過させるポア構造を有しており,その中心部に NPA ドメインと呼ばれるアスパラギン,プロリンおよびアラニンからなる各 AQP アイソ フォーム間において保存された領域を持つ4.また,AQP1,2 および 5 は NPA ドメ イン近傍にシステイン残基を持っており,このシステイン残基に水銀が共有結合する ことで水透過性が抑制されると従来から考えられてきた28.さらに,近年になって銀 および金化合物が AQP1 阻害作用を持つことや30,銅およびニッケル化合物が AQP3 の水透過性を特異的に抑制することも報告されている.本研究でも前章に記したよう に蒼朮に含まれる金属成分が AQP5 を阻害することを明らかにしたが,これらの知 見は AQP が種々の金属イオンにより阻害されることを示唆している.また,これま でに報告された水銀とニッケルの阻害作用を比較すると,その AQP アイソフォーム 選択性や阻害の機序も異なることが推定されるが AQP の金属イオン感受性について は従来,十分に調べられているとは言い難い.
そこで本章ではアフリカツメガエル卵母細胞発現系を用いて,AQP1-5 の各アイソ フォームに対する種々の金属イオンの作用を一価および二価の陽イオンを中心に調べ た.
3.2.種々の金属イオンの AQP 水透過性に対する作用
本章では,アフリカツメガエル卵母細胞発現系を用いて種々の金属イオンの各 AQP アイソフォーム水透過性に対する作用を評価した43.本実験法では,まず AQP を発 現させたアフリカツメガエル卵母細胞を実体顕微鏡下に置き,その細胞外液(200 mOsm)に 4 倍量の蒸留水を加えて 5 倍希釈することで低浸透圧処理(40mOsm)し た.その後,浸透圧勾配に従って卵母細胞内へ流入した水分量を,これに伴って生じ る細胞体積の増加として記録した.
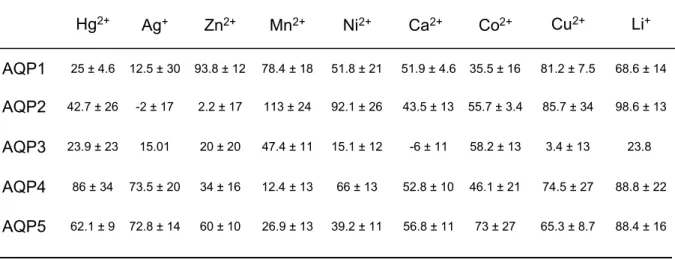
各金属イオン(Hg2+;0.3mM,Ag+;5
µ
M,その他;1mM)は卵母細胞に低浸透圧 刺激を加える 5 分前に細胞外液に加え,低浸透圧刺激のための蒸留水にも金属イオン を加えた.各金属イオンの AQP 阻害作用は金属イオン非存在下に測定した細胞膜水 透過性に対する金属イオン存在下の細胞膜水透過性の割合(%)で示した(Table3).その結果,多くの金属イオンが AQP アイソフォーム選択的な阻害作用を持つことが 明らかとなった.従来の報告と一致して,Hg2+は AQP4 を除く AQP アイソフォー ムを阻害し,Ag+も AQP1 に対して強い阻害作用を示した.しかし,これらの金属イ オンによる阻害作用は AQP1 および AQP2 に比べ,AQP4 および AQP5 では弱かっ た.一方で,Mn2+および Zn2+は AQP4 および AQP5 に対して強い阻害作用を示した.
また,AQP3 は広い金属感受性を持つと考えられた.
AQP4 は今まで阻害物質が見出されていなかったことから,特に強い阻害作用を示 した Mn2+に注目し,AQP4 阻害作用について濃度依存性を調べた結果,Mn2+は濃度 依存的に AQP4 を阻害した(Fig.7).
Table 3. Effects of various metal ions on AQPs water permeability.
Relative AQPs water permeability (% of control) of metal ion treated AQP-expressing oocyte membrane water permeability was calculated based on the water permeability of AQP-expressing oocyte under control condition. Oocytes were injected with water or 5ng cRNA encoding each AQPs (AQP1-5). Osmotic water permeability was measured after a 48-h incubation at 18ºC. In each experiment an oocyte was subjected to 80% hypotonic challenge (from 200 mOsm to 40 mOsm). Indicated metal ions (Hg
2+; 0.3 mM, Ag
+; 5 µM, others; 1 mM) were added to the outside medium 5 min prior to hypotonic challenge. Each data represents mean ± S.E.
AQP1 AQP2 AQP3 AQP4 AQP5
Zn
2+Ca
2+Co
2+Ag
+Cu
2+Li
+Hg
2+Mn
2+Ni
2+25 ± 4.6 12.5 ± 30 93.8 ± 12 78.4 ± 18 51.8 ± 21 51.9 ± 4.6 35.5 ± 16 81.2 ± 7.5 68.6 ± 14 42.7 ± 26 -2 ± 17 2.2 ± 17 113 ± 24 92.1 ± 26 43.5 ± 13 55.7 ± 3.4 85.7 ± 34 98.6 ± 13
-6 ± 11
23.9 ± 23 15.01 20 ± 20 47.4 ± 11 15.1 ± 12 58.2 ± 13 3.4 ± 13 23.8
12.4 ± 13 74.5 ± 27
34 ± 16 66 ± 13
86 ± 34 73.5 ± 20 52.8 ± 10 46.1 ± 21 88.8 ± 22
26.9 ± 13 65.3 ± 8.7
60 ± 10 39.2 ± 11 56.8 ± 11 73 ± 27
62.1 ± 9 72.8 ± 14 88.4 ± 16
RelativeAQPswaterpermeability(%)
Figure 7. Effect of MnCl
2on membrane water permeability of Xenopus oocyte expressing AQP4 Oocytes were injected with water or cRNA encoding AQP4. Oocytes were incubated at 18ºC for 48 h. In each experiment an oocyte was subjected to 80% hypotonic challenge (from 200 mOsm to 40 mOsm). Indicated concentrations of MnCl
2were added to the outside medium 5 min prior to hypotonic challenge. Each data represents mean ± S.E.
0 20 40 60 80 100 120
Con 0.1 0.5 1.0
MnCl
2(mM)
Relative water permeability (% of control)
3.3.ほ乳類細胞の細胞膜水透過性に対する金属イオンの作用
前節の結果より,金属イオンは AQP アイソフォーム特異的な阻害作用を持つこと が明らかになった.AQP は各アイソフォームが組織および細胞種選択的に発現して おり,金属イオンによる AQP アイソフォーム選択的な阻害作用は細胞種選択的な水 透過性阻害作用を示す可能性が考えられる3.そこで,金属イオンによる細胞膜水透 過性抑制作用について赤血球および肺胞上皮細胞株(MLE-12)を用いて調べた.赤 血球は AQP1 を高発現しており,HgCl2感受性の高い細胞膜水透過性を持つことが 報告されている44.また,Ag2+などの AQP 阻害物質の評価においても用いられてい ることから30,赤血球を用いて金属イオンの AQP1 に対する阻害作用を評価するには 妥当であると考えられた.ラット末梢血より赤血球を単離し,1%ヘマトクリットと なるように HEPESbufferに懸濁した.金属イオンは 5 分間処理し,その後600mM マンニトールを含む HEPESbufferと stopped-flow 法により混合し,細胞膜水透 過性を測定した.また,MLE-12 細胞も同様に検討した.Fig.8 に示すように赤血球 は高浸透圧刺激に対して急激な細胞体積変化を示し,高い細胞膜水透過性を持つこと が示された.HgCl2処理によって赤血球の細胞膜水透過性はコントロールの約 40%ま で抑制されたが,ZnCl2およびMnCl2処理では抑制されなかった(Fig.8).一方,
MLE-12 細胞においては HgCl2,MnCl2およびZnCl2はすべてコントロールに比べ 30˜40%程度の細胞膜水透過性抑制作用を示した(Fig.9).また,HgCl2に対する感 受性は MLE-12 細胞よりも赤血球の方が高かった(Fig.8D,9D).これらの結果はア フリカツメガエル卵母細胞発現系を用いた実験結果とよく相関しており,金属イオン の AQP アイソフォーム選択的阻害作用がほ乳類細胞を用いた実験においても示され
Scattered light intensity 1 s
ZnCl
2Control
Scattered light intensity 1 s
Control
MnCl
2Scattered light intensity
1 s
HgCl
2Control
0 20 40 60 80 100 120 140
Con MnCl
2ZnCl
2HgCl
2Relative water permeability ( % of Control)
A B
C D
Figure 8. Effects of metal ions on membrane water permeaebility in red blood cells.
Red blood cells were incubated with 1 mM ZnCl
2(A) or 1 mM MnCl
2(B) or 1 mM HgCl
2(C) for 5 min before measurement of membrane water permeability by stopped-flow light
scattering method. (D) Relative water permeability calculated from the time course of
changing scattered light intensity. The data are means ± SD from six to eight experiments.
Scattered light intensity 1 s
HgCl
2Control
Scattered light intensity 1 s
MnCl
2Control
Scattered light intensity 1 s
ZnCl
2Control
Relative water permeability ( % of Control) 0 20 40 60 80 100 120
Con MnCl
2ZnCl
2HgCl
2A B
C D
Figure 9. Effects of metal ions on membrane water permeaebility in MLE- 12 cells.
MLE-12 cells were incubated with 1 mM ZnCl (A) or 1 mM MnCl (B) or 1 mM HgCl
3.4.AQP 点変異体に対する金属イオンの作用
次に,金属イオンの AQP 点変異体に対する阻害作用を検討した.AQP1 は,その ポア構造付近にシステイン残基を持ち,このシステイン残基に Hg2+が結合すること で水透過性が阻害されることが報告されている28.また,AQP5 も AQP1 と同様にポ ア構造付近にシステイン残基を持っているが,このシステイン残基が金属イオンによ る阻害作用に関与しているかどうかは明らかではない.そこで,金属イオンの AQP5 阻害作用におけるシステイン残基の関与を調べるために,AQP5 の 183 番目のシス テイン残基をアラニンに置換した変異体(AQP5C183A)を作成し,前節の結果から 強い AQP5 阻害作用を示した Mn2+および Hg2+の阻害作用を検討した.一方,AQP4 は水銀感受性のシステイン残基を持っておらず,また,今まで阻害物質に関する報告 はなされていない.Ni2+による AQP3 の阻害作用には AQP3 分子の細胞外領域に存 在するヒスチジン残基が関与していることが示されている32.また,酸性アミノ酸も,
従来からタンパク質の金属配位に関与することが広く知られていることから,
AQP4 については,細胞外領域に存在するヒスチジンおよび酸性アミノ酸に注目し,63 番 目のグルタミン酸,69 番目のアスパラギン酸,151 番目のヒスチジン,228 番目の グルタミン酸および 230 番目のヒスチジンをそれぞれアラニンに置換した変異体 (E63A,D69A,H151A,E228A および H230A)を作成した.これら 5 つの AQP4 変 異体を用いて,前節の結果から AQP4 阻害作用を示した Mn2+,Zn2+および Ni2+の阻 害作用について検討した.1) AQP4点変異体に対するMn2+,Zn2+およびNi2+の作用
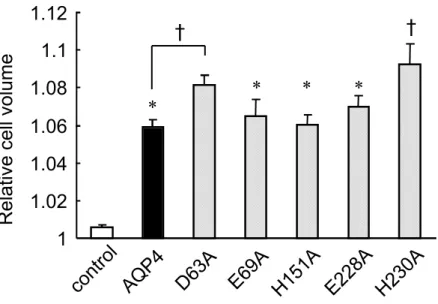
今回作成した種々の AQP4 点変異体の膜上での発現量には著明な差は認められな かった(データは示さず).野生型 AQP4 および AQP4 点変異体を用いて水透過性を 比較したところ,水透過性亢進作用は E69A,H151A および E228A では,野生型と 著明な差は認められなかったが,興味深いことに,D63A および H230A は野生型よ りも著明な水透過性亢進作用が認められ,特に H230A の亢進作用が顕著であった (Fig.11).これらの結果は,63 番目のグルタミン酸および 230 番目のヒスチジンが AQP4 の透過性亢進作用に一部抑制的に関与している可能性を示唆している.
一方,Mn2+,Zn2+および Ni2+による阻害作用について調べると,E63A,D69A, H151A の3つの点変異体では,Mn2+による阻害作用は消失した(Fig.12).すなわ ち,Mn2+は 63 番目のグルタミン酸,69 番目のアスパラギン酸および 151 番目のヒ スチジンの少なくとも3つのアミノ酸を介して阻害作用を現していることが示唆され た.また,E63A,D69A,H151A および E228A の4つの点変異体では,Zn2+の阻害 作用が消失した(Fig.12).このことから,AQP4 の Zn2+による阻害には,63 番目の グルタミン酸,69 番目のアスパラギン酸,151 番目のヒスチジン,228 番目グルタ ミン酸の少なくとも4つのアミノ酸が関与していることが示唆された.さらに,E63A および D69A の 2 つの点変異体では,Ni2+による阻害作用が消失した(Fig.12).従 って,Ni2+の阻害に,63 番目のグルタミン酸および 69 番目のアスパラギン酸の少 なくとも 2 つのアミノ酸が関与していることが示唆された.
これらの結果を併せて考えると,AQP4 分子中の 63 番目のグルタミン酸および 69 番目のアスパラギン酸は,金属イオンによる阻害に必須の機序に関与すると考えられ,
Figure 10. Hourglass model for AQP4 membrane topology
1 1.02 1.04 1.06 1.08 1.1 1.12
control AQP4 D63A E69A H151A E228A H230A
Relative cell volume
* * * *
† †
Figure 11. Effects of mutated AQP4 expression on Osmotic water permeability in Xenopus oocytes.
Summary of osmotic water permeabilities. Increase rate of cell volume was calculated from increase volume of cell expressing mutated AQP4s. Ordinate indicates rate of cell volume increase (cm
3/min) Each data represents mean ± S.E., with a numbers of oocyte 4-10. * and † indicate significant difference from the control and from AQP4, respectively.
H
2N HOOC
A N N P A
P
H
2O
AQP4
D H
H E E
230 63 228
69
151
0 20 40 60 80 100 120 140 160 180
AQP4 E63A D69A H151A E228A H230A
Relative water permeability (% of control)
Mn
2+(1.0 mM)
0 20 40 60 80 100 120 140 160 180
AQP4 E63A D69A H151A E228A H230A
Relative water permeability (% of control)
Zn
2+(1.0 mM)
0 20 40 60 80 100 120
AQP4 E63A D69A H151A E228A H230A
Relative water permeability (% of control)
Ni
2+(1.0 mM)
Figure 12. Inhibitory effect of metal ions on Osmotic water permeability of Xenopus oocytes expressing AQP4 and AQP4 mutants.
Oocytes were injected with water or cRNA
encoding AQP4 and AQP4 mutants. Oocytes
were incubated at 18ºC for 48 h. Effect of 1.0
mM MnCl
2, ZnCl
2and NiSO
4on increase
volume of cell expressing AQP4 and AQP4
mutants was examined. Inhibition rate was
calculated based on the water permeability of
oocyte non-treated with each ions. Ordinate
2) AQP5点変異体に対するHg2+およびMn2+の作用
AQP5 においても AQP1 同様,Hg2+による阻害にはシステインが関与すると考 えられているが,実験的にそれを示した報告はない.そこで,AQP5 の 183 番目の システイン残基をアラニンに置換した変異体 AQP5C183A に対する Hg2+および本 研究において強い AQP5 阻害作用を示した Mn2+の作用を検討した.
Westernblotting 法により調べた細胞膜上の AQP5 量は野生型と点変異体との間 で著明な差は認められなかった(データは示さず).また,AQP5 による水透過性の 亢進も,野生型 AQP5 および AQP5C183A では同程度の作用が認められ(Fig.14),
183 番目のシステイン残基が AQP5 の水透過性亢進に必須ではないと考えられた.
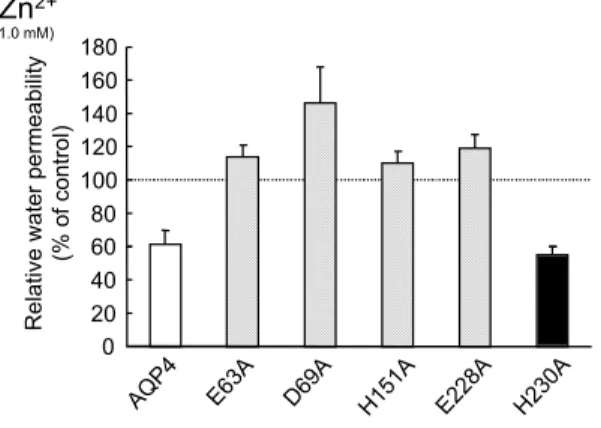
しかし,Hg2+および Mn2+による阻害作用は,AQP5C183A では消失した(Fig.14).
従って,AQP5 の Hg2+および Mn2+両金属イオンによる AQP5 阻害には,少なくと も 183 番目のシステインが関与していることが示唆された.
Figure 13. Hourglass model for AQP5 membrane topology.
0 0.01 0.02 0.03 0.04 0.05 0.06 0.07 0.08 0.09
control AQP5 C183A
control 0.3 mM Hg
2+Cell volume increased (cm
3/min) *
*
†
0 0.01 0.02 0.03 0.04 0.05 0.06 0.07 0.08 0.09
control AQP5 C183A
control 1.0mM Mn2+
Cell volume increased (cm
3/min) *
†
*
A B
H
2N HOOC
A N N P A
P
H
2O
C
Hg
2+AQP5
183
3.5.考察
1 金属イオンによる AQP アイソフォーム選択的な阻害作用について
本章では既知の AQP 阻害作用を持つ金属イオンに加えて一価および二価の陽イオ ンを中心に AQP1˜5 の各アイソフォームについての金属イオン感受性を検討した.
その結果,既知の AQP 阻害作用を持つ金属イオン以外にも多くの金属イオンが AQP 阻害作用を持つことが明らかになった.また,AQP は各アイソフォーム間で異なる 金属イオン感受性を持つことも明らかになった(Table3.).まず,AQP3 は以前より 知られていた銅,ニッケル以外にも本実験で検討した金属イオンのほとんどすべてに 感受性を持っており,幅広い金属イオン感受性を示した.AQP3 はアクアグリセロポ リンとも呼ばれ,水分子以外にもグリセロールも透過させることが知られており,本 実験で比較した他の AQP アイソフォームとは構造上も異なる領域を持つことが広い 金属感受性を持つ理由であると考えられた43.一方,AQP4 は水銀感受性システイン 残基をその構造内に持っておらず,AQP4 阻害物質は今まで見出されていなかったが,
本実験においてマンガンおよび亜鉛が AQP4 を強く阻害することが初めて見出され た.また,AQP5 は水銀感受性のシステイン残基を持つことから,水銀が阻害作用を 持つことは従来から考えられていたが,本実験において初めてその阻害作用が明確に 示された.さらに,マンガンおよびニッケルが AQP5 を阻害することを初めて見出 し,その阻害作用は水銀よりも著明であった.さらに,ほ乳類細胞においても細胞膜 水透過性の金属イオン感受性が細胞により異なることが示された.本章では,HgCl2, ZnCl2および MnCl2に対する感受性を赤血球および MLE-12 細胞を用いて検討した が,両細胞は異なった金属イオン感受性を示した(Fig.8,9).赤血球は AQP1 を,
MLE-12 細胞は AQP5 を発現しており,アフリカツメガエル卵母細胞発現系におけ る実験結果と併せて考えると,両細胞の金属イオン感受性は発現している AQP アイ ソフォームに依存していることが示唆された.しかしながら,AQP1 と AQP5 の2 つのアイソフォームおよび 3 種類の金属イオンについての結果であるため,さらなる 検討が必要である.しかし,単一の AQP アイソフォームのみを発現している細胞株 は他にはほとんど知られていないため,AQP 過剰発現細胞を作成して実験すること
イオン感受性は大きく 3 つにわけることができる.AQP3 は幅広い金属イオン感受性 を持ち,AQP1 および 2 は水銀および銀によって強い阻害作用が認められ,一方で AQP4 および AQP5 は亜鉛およびマンガンによって強い阻害作用が認められた.ま た,金属イオンの中でも特に亜鉛は幅広い AQP アイソフォームに対して阻害作用を 持つことが明らかになった.このことから AQP アイソフォーム間には共通した亜鉛 感受性の構造を持つことが示唆された.
2 AQP 点変異体に対する金属イオンの作用について
金属イオンの AQP 阻害における作用点を明らかにするため,種々の AQP 点変異 体を用いて実験を行なった.以前の報告から,水銀は AQP1 のシステイン残基変異 体に対しては阻害作用を持たないことが示されており,このことから水銀の作用点が ポア構造付近に存在するシステイン残基であると考えられてきた28.また,本研究に お い て も AQP1 の 水 銀 感 受 性 シ ス テ イ ン 残 基 を ア ラ ニ ン に 置 換 し た 変 異 体 (AQP1C189A)は水銀に対して感受性を持たないことが確認できた(データは示さ ず).また,AQP5 においてもシステイン残基変異体(AQP5C183A)を作成し,水銀 の阻害作用を検討したところ,AQP5C183A は水銀により阻害されなかったことから AQP5 においても AQP1 と同様に水銀の作用点がポア構造付近のシステイン残基で あることが確認された(Fig.14).さらに AQP5C183A はマンガンに対する感受性が 消失した(Fig.14).このことから,マンガンも水銀と同様にシステイン残基を作用 点としていることが明らかになった.しかしながら,マンガンは AQP1 に対しては 阻害作用が認められなかったことはマンガンの作用点はシステイン残基だけではなく 他のアミノ酸の関与も必要であることが示唆された.
ンの阻害作用には 63 番目のグルタミン酸,69 番目のアスパラギン酸および 151 番 目のヒスチジンが関与していた.この結果は,AQP4 と AQP5 に対するマンガンの 作用点が異なることを示している.また,亜鉛の阻害作用には 63 番目のグルタミン 酸,69 番目のアスパラギン酸,151 番目のヒスチジンおよび 228 番目のグルタミン 酸が関与しており,ニッケルの阻害作用には 63 番目のグルタミン酸および 69 番目 のアスパラギン酸が関与していた.これらの結果より,金属イオンはそれぞれ独自の 作用点を持っていること,また,AQP にはシステイン残基だけでなく多くの金属イ オン感受性アミノ酸残基が存在することが明らかになった.AQP4 におけるマンガン,
亜鉛およびニッケルの作用点のうち,63 番目のグルタミン酸と 69 番目のアスパラギ ン酸は 3 つの金属イオンに共通の作用点であること,151 番目のヒスチジンはマンガ ンおよび亜鉛の特異的作用点であること,また,228 番目のグルタミン酸は亜鉛に特 異的な作用点であることが示された.また,どの金属イオンも AQP4 に対して複数 の作用点をもっており,1つでも欠損するとその阻害作用は消失したことから,AQP4 阻害において1つの金属イオンに対し,複数のアミノ酸が関与しており,さらに全て のアミノ酸が必要であると考えられた.
本章の結果から,多くの金属イオンが AQP アイソフォーム選択的な阻害作用を持 つことが明らかとなり,特に今まで阻害物質が見出されていなかった AQP4 に対す る阻害物質も見出すことができた.金属イオンそのものが AQP 阻害薬として医薬品 として応用は難しいが,AQP4 阻害作用における金属イオンの作用点が明らかにな ったことで AQP4 阻害剤のモデルを提唱することは可能であると考えられる.第 1 章でも述べたように,AQP4 阻害物質は脳浮腫治療および予防薬となり得ることが示 唆されている.本章で得られた知見を元に考えると,マンガンが最も強い AQP4 阻 害作用を示したことから AQP4 の 63 番目のグルタミン酸,69 番目のアスパラギン 酸および 151 番目のヒスチジンの立体構造を元にした特異的阻害剤を考案すること ができると考えられる.また,AQP 変異体を用いた実験からも示されたように,多 くの金属イオンは AQP の細胞外に存在するアミノ酸残基に結合することで阻害作用 を示すと考えられ,また同じ金属イオンでも AQP アイソフォームの違いによってそ
るための構造を独自に持っていることを示唆しており,AQP アイソフォーム選択的 な阻害薬の開発の可能性が考えられる.AQP がアイソフォーム選択的に組織に分布 していることを考えると,AQP アイソフォーム選択的な阻害薬は組織選択性が高く,
副作用の少ない薬となりうると考えられる.