小腸のビタミン
A吸
収・代謝の調節機構に関する研究
∼ニワ トリヒナ十二指腸 とヒ ト小腸様細胞株 CacO-2 BBe細 胞 を用いた場合∼
山 口 範 晃D・ 高 瀬 幸 子の。駿 河 和 仁D The study of tamn A absorption and metabolism in chick
duodenum and human inttstihal CacO-2 BBe cells
NonakiYAMAGUCHI°
,Sachiko TAKASED,Kazuhito SURUGAつ要 約
ビタミン
Aは
様 々な生理作用 を有 し、小腸 内の ビタ ミンA吸
収・代謝 に関わる各種 の酵素や タンパ ク質 についての重要性が報告 されて きたが、 これ らの発現が どの ような機序で調節 されて いるかは十分 に解明 されていない。 これまでに著者 らは、ニワ トリヒナ十二指腸及びヒ ト小腸上 皮吸収細胞様細胞株CacO-2 BBe細胞 を用い、小腸の ビタミンA及
び βカロテ ン吸収・代謝 の中 心的な役割 を演ず るβカロテン中央開裂酵素 (β…carotene 15,15生mOnOOxygenase:BCM01)の
生 理的役割 とその発現調節 メカニズムについて追究 した。その結果、ニワ トリヒナの十二指腸の孵 化後数 日において、 レチナールか らレチノイン酸 を生成す る酵素reintt dehydoЮgenase りpe l(RALDH l)と
BCMOl遺
伝子発現 が共 に増大 した。 また、ニ ワ トリ十二指腸 や CacO-2 BBe 細胞 において、各種 ホルモ ンによ りBCMOl発
現が克進 した。 これ らの研 究成果 は、小腸 内のビタミン
Aの
生体内利用効率や恒常性 の調節機構 について新 たな基礎 的知見 を提示で きる と考 えている。キ ー ワ ー ド:ビタ ミ ンA、
BCMOl、
ニ ワ トリ十 二 指 腸 、CacO-2 BBe細胞1.序
論 脂溶性 ビタ ミンの一種 であ る ビタミンAは
細胞 の分化 ・増殖 、形態形成、視覚作 用、免疫系 な どにお い て重要 な役割 を演 じ、 レチ ノール、 レチナール、 レチ ノイ ン酸 とその類縁化合物 を総称 して レチ ノイ ドと呼 ばれている。。 ビタミンAの
供給源 として、い くつかの動物性食 品 に含 まれ る レチエルエステル と、主 に緑 黄色野菜や果物 な どの植物性食品 に含 まれるカロテノイ ドの中で、 ビタ ミンAの
前駆体 プロ ビタ ミンA(β
カロテ ン、αカロテ ン、 ク リプ トキサ ンチ ン)が
あ る。 ビタ ミンAは
、 ヒ トな どの哺乳類 の生体 内で は合 成で きないので、 これ らの食 品か ら摂取 しなければな らない。現在 、温暖化 による気候変動や世界人口の急 増 によ り、将来的 に食糧不足が予測 される。現実 として、食糧不足 に悩 まされてい る開発途上国では ビタミ ンAの
摂取不足 による夜 盲症 や免疫力 の低 下、 さらに乳幼児 の発育不 良や栄養失調 は深刻 な問題 とされて いるの`9。 その一方で、 ビタ ミンA過
剰摂取 に よる全 身倦怠感、頭痛、吐 き気 な どの脳圧克進 な どの症状 も 見 られめ、特 に妊婦 の場合 には、胎児奇形が問題 になってい る°。 そのため、上記 の ような ビタ ミンA栄
養 問題 を改善す るため に も、食事 か ら如何 に して ビタ ミンAを
効 率 よ く生体内で利用 し、その生理作用 を発揮す るか を究明す る必要がある。筆者 らはこれ まで にヒ トの小腸 1)長崎県立大学 看護栄養学部 栄養健康学科 2)浜松大学 健康 プロデュース学部 健康栄養学科長崎県立大学 看護栄養学部紀要 第 9巻 (2008) 上皮吸収細胞 をモデルとしたヒ ト小腸上皮吸】踊田胞様細胞株 CacO-2 BBe細 胞やニワ トリ十二指腸 を用いて、 ビタミン
A吸
収・代謝 に関わる研究を行 ってきた。本稿 は、小腸の ビタミンA及
びβカロテン吸収・代謝 の中心を担 うβカロテン中央開裂酵素 (β―carotene 15,15■monooxygenasαBCM0 1)に
関 して、 これまでの 筆者 らの研究成果の概要 をまとめた。2.小
腸 ビタ ミンAの
吸収・ 代謝 食餌か ら摂取 されたビタミンA(レ
チエルエステル)お
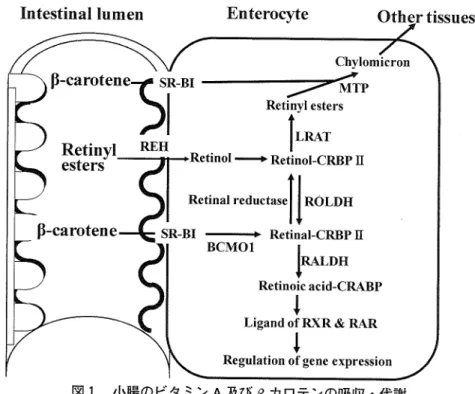
よびβカロテンは、図 1に 示す ように吸収、代 謝、輸送 される。食餌由来のカロテノイ ド(主にβカロテン)は複合 ミセル内に取 り込まれ、scavenger recep―tor dass B type I(SR―
BI)を
介 して小腸粘膜上皮細胞内に取 り込まれる0。 このβカロテンは、そのままカイロ ミクロンに組み込まれ、腸管膜 リンパ管へ と輸送される経路 と、βカロテン中央開裂酵素(β―carottne 15,15'
―
monooxygenase:BCM0 1)に
より2分
子の レチナールに転換する経路があるのヽ0。βカロテンか ら転換 され たレチナールは、細胞性 レチノール結合 タンパク質タイプⅡ(cellular reinol―binding prottin type Ⅱ:CRBPII) と複合体 を形成 し9、 レチナール還元酵素
(reintt reductase)に よリレチノエルーcRBPII複 合体へ転換 され
る10ヽけ。レチノールー
cRBⅢ
複合体は、長鎖脂肪酸 とレシチンレチノールアシル転移酵素(lecithin retinol acyト transattasα
LRAT)に
よってレチエルエステルを形成するりヽ0。 このレチエルエステルは、マイクロソームトリグリセリド転送タンパク ( crosom江 伍glyceride ttansfer pЮ
tein MTP)の
作用 によって、他の脂質(ト
リグリセ リド、 リン脂質、コレステロールなど
)と
もにカイロミクロンに取 り込まれて、 リンパ管へ輸送 さ れ、血液を循環 して、各組織へ連ばれる勁。一方、 レチナール脱水素酵素 (reinal dehydrogenase:RALDH) により、 レチナールからビタミンAの
生理活性 を有するレチノイン酸へ生成 されるn。9_θね レチノイン酸
やall―″η∫レチノイン酸は、核内受容体 レチノイン酸 レセプター (reinOic acid recepto■
RAR)及
びレチノイド
Xレ
セプター (reinoid X receptor RXR)の リガン ドとして結合することで生理作用 を発揮 し、様々な遺図
1
小陽 の ビタ ミンA及
び β 力 ロテ ンの吸収・ 代謝BCMO■ β―carotene 15'15'―monooxygenase,SR―BI:scavenger rcceptor class B type I,cRBPⅡ :cellular reinoユ‐binding proteinサ pe Π,LRAT:lecithin
reinol acyltranferasc,MTP:Hucrosomal tiglycende transfer protein,ROLDH:retinol dehydrogenase,RALDHi reinal dehydrogenase,CRABPi cellu―
1をr reinoic acid―binding proゃin,REととretlnylester hydrolase,RAR:reinoic acid receptor・ RXR:reinold X FeCCptoi
htestinal lumen
F‐CarOteme_」L sR―
BI―
Reiral_cRBP ⅡBC
101 1_DH
Retinoic acidぃ
CRABP
↓
Ligand OfRXR&敵R
I
伝子発現 の転写 因子 として機 能す る。。 なお、 これ らの レチ ノイ ン酸 の中で、9‐どね レチ ノイ ン酸 は
RAR
及 び
RXRに
よる転写活性 を有す るが、証1-′胞乃∫レチ ノイ ン酸 はRARに
対 してだけ親和性 を示すり`0。 一方、
食餌 由来の レチエルエステルは、 レチニルエステル水解酵素 (俺dttlester hydrolase:REH)に よって加水分
解 されて レチ ノール とな り、小腸粘膜 上皮細胞 内 に吸収 され る。。吸収 された レチノールはレチ ノールー CRBPⅡ 複合体 を形成 し、 レチ ノール脱水素酵素 (reinol dehydrogenasei ROЫ
)H)に
よ リレチ ノイ ン酸へ合成す る経路 と、
LRATに
よって再 び レチニルエステル を形成 し、各組織へ運搬 される。3.BCM01発
現 の生理 的意 義BcM01は
脊椎動物 の細胞 質 に存 在 す る酵素 であ り、1原子 の酸素 が βカロテ ンの15,15'位 に付 加 し、 エポキシ化 され、1分
子 の水分子 の付加 によ り開裂 し、その結果2分
子 の レチ ナールが生成 され る20。 多 く の杏椎動物 において、BCMO lは
小腸上皮吸1跡巴胞 で高い酵素活性 を示 し、その他 に肝臓 や、肺 、脳 、腎臓 な どに も酵素活性が確認 されてい る20、勃。 また、各種のBCMO l遺
伝子が同定 された こ とで、BcMOl遺
伝 子 の発現性 や発現調節 の機序 な どの遺伝子工学 を用 いた研 究が進 め られた。 ヒ トやマ ウスのBCMOl遺
伝 子 の5'上流 プロモー ター解析 を行 った ところ、脂質代謝 に関わる転写 因子ペ ルオキ シソーム増殖剤 活性化 レセプター γ(perOxisome prolif∝ ねr activtted receptor γ :PビN)と
RXRの
二量体の応答領域が確認 された20、20。 Lindqvistら は、 ヒ ト
BCMO l遺
伝子 の変異型 (ア ミノ酸配列170番 目のス レオニ ンが メチ オニ ンに 置換 した型)で
は、βカロテ ンの ビタ ミンAへ
の転換効率が著 しく減少 し、体 内 に βカロテ ンが過剰 に蓄 積 され、低 ビタミンA状
態 に陥 る可能性 について報告 した20。 さらに、BCM0 1遺
伝子 ノ ックアウ トマ ウス を用 いた研 究 で は、低 ビタ ミンAレ
ベ ルに陥 るだけではな く脂質代謝の恒常性が崩壊 し、脂肪肝 を発症す るこ とも報告 されてい る20。 この ように、BCM0 1は
体 内の βカロテ ン及 び ビタ ミンAレ
ベ ル を調節す る だけではな く、脂質代謝 の調節 に も関わつていることが示 された。BCMO lは
ビタミンA恒
常性 において重要 な生理 的 な調節機 能 を持 ち、βカロテ ンを過剰 に摂取 して も ビタ ミンA過
剰 に よる疾患 は発生 しない と言 われてい る。その根拠 とされてい る研 究 として、過剰 の レチ ノイ ン酸や レチエルエステル を投与 した場合 、 ラ ツ ト小腸 のBCMOl酵
素活性 は低 くな り、逆 にRARア
ン タゴエス トや シ トクロームP450(レ
チ ノイ ン酸 を異化す る酵素)の
活性化剤 を投与 した ラ ッ トで はBCMO
l活性 は上昇 した2の 。最近の研 究で は、 ビタミンA恒
常性 を調節す る因子の一つ として、小腸 に特異 的 に発 現す る Intesine specittc homeodomein(lsx)が 、BCM0 1及
びSR…BIの発現 を負 に調節 し、 さらに体 内の ビタミン
Aレ
ベルに応 じて これ らの遺伝子発現 も変動す ることが報告 された20。 この こ とか ら、Isxは小腸吸 収細 胞 内の βカロテ ンの取 り込 みか らレチ ノイ ドヘ の転換 までの調節 に関わる因子 であ るこ とを示 唆 して い る。 著者 らのBCM0 1発
現 に関わ るこれ までの研 究 は29-30、 ニ ヮ トリ胚及 び ヒナの小腸 を用 いて、発達過程 にお けるBCMOl遺
伝 子発現性 の変動 を解析 した。 さらに、各組織 の成熟化 に レチ ノイ ン酸 の生理作 用 が 必須 であ り、その合成酵素 であ るRALDHと BCMO l遺
伝 子発現 の関連性 を検討 した。 また、ニ ワ トリ小 腸及 びヒ トの小腸上皮吸J難田胞 をモデル としたCaco-2 BBe細胞 にグル ココルチ コイ ドや 甲状腺 ホルモ ンを 投与 し、BCMOl遺
伝子 の発現性 を検討 した。長崎県立大学 看護栄養学部紀要 第 9巻 (2008)
4.ニ
ワ トリヒナ及び ヒ ト小腸上皮吸収細胞様細胞株CacO-2 BBe細
胞 のBCMol発
現 に関 する研 究
(1)ニ
ワ トリヒナ十二指腸の発達過程におけるBCMolと
RALDH l遺
伝子発現の関連性 ニワ トリ胚およびヒナの発育や各組織発達において、ビタミンA(レ
チノイン酸)は
必須の栄養素である。 孵化後 2日 までのニワ トリヒナは餌 を摂取せず、腸管腔に入 り込んだ卵黄からβカロテンまたはレチノー ルが供給 されてレチノイン酸を合成 していると考えられている3か 。 また、ニヮ トリヒナの十二指腸 を用いたr 研究では、BcMol酵
素活性 は孵化 した数 日後 に急激に上昇 し、十二指腸の絨毛細胞の成熟過程期にBcMo
l酵 素活性が誘導 されることを報告 した39、30。 そこで著者 らは、孵化前から孵化数 日後のエワ トリ胚及びヒ ナの十二指腸 を用いて、BcMo l遺
伝子発現の変動について解析 し、さらにレチナールからレチノイン酸を 生成する酵素RALDH lと
BcMo l遺
伝子発現の変動 との関連性 を検討 した8ω 。その結果、ニワ トリヒナの 孵化後 において、BcMo l遺
伝子発現量が増大 した (図 2)。 同様 に、RALDH l遺
伝子発現量 も急激 に孵化 後 に増大 した。また、血清 βカロテン濃度 もニヮ トリ孵化前後において急激に上昇することが報告 されて いる切。これらのデータか ら、孵化後数 日間はBcMo lに
よって βカロテ ンか ら転換 された レチナールか らレチノイン酸生成が活発であることが示唆される。このように、孵化前後のニワ トリヒナのBcMol発
現の生理的役割 として、未成熟の小腸 を発達 させるために、ビタミンA(レ
チノイン酸)供
給 に寄与 してい ることが考えられる。BCM01
(Days) BC 101 1 o.l Kb)' RALDH l (Day9 E18 RALDHl t2.3Kb) ● 18S (1.8Kb)8
一一
E
弱
s
韓
3 静!
機
・
・8 S い 3 , O F ︲ 却 ﹁ 2 . 0 卜 ︲ 齢 ︲ ・ , 0 卜 ︲ o ・ 5卜 ︲ OL Kb) E18 0 3 5 8 15 gDays) 2.S 将2.0 3憂
1.5 日 ど1,o H 館 0.5 0 卒Hatth Penbd OfdevdOpment HatcL Days ofdttdOpment! 3 5 8
図
2
三 ワ トリヒナ孵化前後 の十二指陽BCM01と
RALDH l遺伝 子発現 の変動BCMOlと RALDHl mRNA発現量は、ノーザンブロット法により解析 し、18S rRNA発 現量で補正した相対値をグラフ化した
(平均値
±SEM、 n=11-12)。 E18は18日胚(孵化2日前)を示す。異なるアルファベットは有意差を示す ●<o.05、 Duncan's test)。
(2)ニ
ワ トリ十二指腸のBCMol遺
伝子発現に及ぼすハイ ドロコルチゾンの影響 副腎皮質か ら分泌 されるグルココルチコイ ドゃ甲状腺ホルモンは、小腸上皮吸収細胞の分化・成熟おいて 重要な生理的役割を果た している30。 先述 したように、エワ トリヒナ十二指腸のBCMO l発
現は孵化後急激 に上昇する。従って、ニヮ トリヒナ十二指腸 において、これらのホルモンの作用によってBcMol発
現が どの ように影響するかを検討 した30。 まず、孵化前後のニワ トリヒナを用いて、ニヮ トリの生理的グルココ ルチコイ ドである血清ハイ ドロコルチゾン (hydЮ cosOne:HC)濃
度変動 を測定 したところ、BcMo l発
現変動 と類似 していることが分かった (図3A)。 次 に、ニ ヮ トリ胚 (受精卵
)に Hc投
与 した ところ、孵化 り 寄 共 喜 く 密 鋼 日 0 > 導 ミ 窮 館0 ・ 6 0 。 5 0 ・4 0 。3 0 ・2 0 ・ 1 0 命 皇 ヽ こ の ﹃ o ‘ ヽ と 目 8 ﹃ 0 0 0 目 日 喜 3 ∽ Day18 embryo control HC
8招
;湛
Kb) 0-day―old control HCH
お 別 1 ・5 ・,0 0・5 0 り ︻ り 、5 ︻ 鸞 ヅ ″ 唱 H 。 > 導 こ 巧 立 ︻ 8 0Ⅱ
れ
ch Period of developmentE18 P 3
Ⅱatch Period of development 図3
ハ イ ドロコル チ ゾンは孵化前後 のニ ワ トリヒナ十二指陽 のBCM01遺
伝 子発現 を誘 導 す る(Alニヮトリヒナ孵化前後の血清ハイ ドロコルチゾン(HC)の濃度変動をELISA(Enzymc■ ed iHImunosorbent assay)に よつて測定した
(平均値上SEM、 n〓 9-15)。 異なるアルファベットは有意差を示す ●<0.05、 Duncan's ttst)。 (B)ハイドロコルチゾンによるニワトリヒ
ナ孵化前後の十二指腸BCMOl遺伝子発現への影響。BCMOlと
RALDH l―
A発現量は、ノーザンブロット法により解析 し、18STRNA発現量 で補 正 した相対 値 を グラ フ化 した (平均 値 ±SEM、 ■=6-9)。 E18は18日胚 (孵化2日前)を示 す 。*:フ <0.05(Smdent's
t test)。 2日 前 (ニワ トリ胚18日
)及
び孵化 当 日でBCM0 1遺
伝子発現量が増大 した (図 3B)。 これ らの結 果 か ら、Hcは
BCMOl発
現 を遺伝 子 レベ ルで誘導す るこ とが示唆 された。 また、HC投
与群 で は、甲状腺 ホルモ ン の活性型である トリヨー ドサ イロニ ン (T3)の血清濃度 も上昇 していた29。 これ らの結果 か ら、孵化前後 に お けるニ ワ トリ十二指腸 のBCM0 1発
現誘導 にはHC及
びT3などのホルモ ン因子 に よつて調節 され てい る 可能性 を考 えた。(3)CaCO-2 BBe細
胞 の ビタ ミンA吸
収 。代謝 に及 ぼすT3の影響CacO-2 BBe糸田胞 は、Caco-2細 胞 か らサ ブクローニ ング された細胞 である。小腸吸収細胞 は、未分化 の ク リプ ト細胞 か ら増殖 し、絨毛先端へ と分化 ・成熟す る過程 を有す る特徴 があ る。Caco-2 BBe細胞 は ヒ トの 小腸吸収細胞 の形態学的 に良 く似 た特徴 を示すモデル として用 い られてい る30。 また、
BCM0 1酵
素活性 はCaco跡
田胞 で は検 出で きないが、Caco-2 BBe細胞 で はBCMO l酵
素活性 及 び遺伝 子発 現量 ともに Caco-2 細胞 に比べ てか な り高い°ヽ朝`3つ 。従 って、著者 らはヒ ト小腸吸1琳田胞 の ビタ ミンA及
び βカロテ ン吸収 ・代 謝 に関わる冴 究ではCaco 2 BBe細胞 を用 いてい る。 上記 の結果の ように、ニワ トリヒナ十二指腸 でのBCM0 1発
現 は、HCや
T3の影響 によって九進 され る可 能性 を示 した。そ こで、著者 らはCacO-2 BBe細胞 を用 いて、T3及び合成 グル ココルチ コイ ドであ るデキサ メタゾンに よるBCMO l発
現へ の影響 について検討 した鋤。その結果、デキサメタゾンで処理 した Caco-2BBe細
胞 で はBCM0 1発
現 は変動 しなか ったが、T3で処理 した場合 、濃 度依存 的 にBCM0 1遺
伝 子発現量 が増大 した (図4A)。 さらにCaco-2 BBe細胞 が未分化 の状態 よ りも、 コンフルエ ン ト7日 後 の成熟 した状態 で顕著 に
BCM0 1遺
伝 子発現量 が増大す る ことが分 か つた (図4B)。 また、転写 阻害剤 であるア クチ ノマイシ ン
Dと
T3で同時 に処理 した細胞 中で は、T3によるBCM0 1発
現量 の増大 を抑制 した (図4C)。 これらの結果 か ら、
BCMOl発
現 は転写 レベ ルで 吼に よつて調節 され る可 能性 を示 した。同時 にT3で処理 した CacO-2 BBe細胞 は、LRAT遺
伝子発現量 も増大 した こ とか ら (図4D)、 T3はβカロテ ンか らビタ ミンAの
転換 、更 にその腸管吸収 に関わる遺伝 子発現 を調節 してい る可 能性 を示 した。 しか し、BCM01発
現 がT3長崎県立大学 看護栄養学部紀要 第 9巻 (2008)
7 days pos卜co■muent
図
4 Caco-2 BBe細
胞 のT9によ るBCM01及
びLRAT遺
伝 子発現 への影響いCaco-2 BBe細 胞をジメチルスルホキンド(DMSO)及び1100由r T3で24時間処理 したときBcMol遺伝子発現畳。(B)コンフルエント
(細胞が培養器面を覆い尽 くした状態)当 日及びコンフルエント7日後のCaco-2 BBe細 胞をDMSO及び1004M Taで 24時間処理 したとき
のBCMOl遺伝子発現畳。(CICacO-2 BBe細 胞 をDMSO、 1004M T 3及び5 vg/期アクチ ノマイシンDで24時 間処理 した ときの
BCMol
遺伝子発現量。(Dlcaco-2 BBe細 胞 をDMSO及び100nM島で24時 間処理 した ときのLRAT遺伝子発現量。い ―(D)のBCMol及びLRAT遺
伝子発現量 は、 リアルタイムRT‐PCRによって測定 し、内部標準 bosomal PЮtein,large,PO(RPLP O)で 補正 した相対値 をグラフ化 した
(平均値 士sEM、 n=4)。 *:P<o.o5,**:p<0.01(smdent'st― test)。
確 になっていない。今後、
BcMo lプ
ロモーター領域内のTR結
合領域の同定やTR発
現ベ クターを用いた トランスフェクションなどの遺伝子工学的(な手法 を用いた更なる研究が必要 とされる。 このように、ホルモ ンによるBcMol遺
伝子発現の調節機序はヒ トやニヮ トリなどの種 によって異 なる ことが考えられるが、小腸でのビタミンA吸
収・代謝 を調節する上でこれ らのホルモンは必要不可欠であ ると考えられる。 以上のように、我々はニワ トリヒナおよび cacO-2 BBe細 胞 を用いてBcM0 1発
現の調節機序 について研 究 して きた。ニヮ トリヒナの発達のために、十二指腸でのBCMO l発
現 はビタミンA(レ
チノイ ン酸)供
給源のための重要な生理的役割 を有 していることが考えられる。また、ニヮ トリとcacO-2 BBe細 胞 を用い、 各種 ホルモ ンを投与 した結果か ら、ニヮ トリとヒ トの間にはBCMol発
現誘導のメカニズムに種差がある か もしれない。それについては更に追究 してい く必要がある。現在、我々は、cacO-2 BBe細 胞 を用いた研究 を中心 に進展 してお り、Hepttocyte nuclear factOr-4α (IINF-4α
)及
びIINF-lα に よるBcMol遺
伝 子 発現 が遺伝 子転 写 レベ ルで調節 され てい る こ とを見 出 した。 これ らの転写 因子 や ホルモ ンは、互 い に競合 ま たは相互的にBCMol発
現 を制御することで、小腸内での ビタミンAの
利用 と吸収・代謝、及び各組織ヘ の輸送 を調節 し、各生体内のビタミンAの
恒常性の維持 に寄与 している可能性が考えられる。このように、 4.0 5.0 0 0 0 0 3 2 1 り ︻Φ > 0 ︻ 奪 じ 角 国 電 口 0 > 一 やヽ ︻0 巳 ︹ ・ 0 ・ 0 ・ 0 ・ 0 0 4 3 2 1 の 巧 > 留 く Z 区 日 ● > ︺ “ 巧 出 ・ 5 ・ 0 ・ 5 0 1 1 0 ∽珂 景 喜 く Z 留 日 Φ > 攀 “ 巧 雷 ・0 お ・0 ・5 0 2 1 1 0 黎 o 景 喜 く Z 留 日 0 〓 ↓“ 巧 目 C. 2.5 2.0DLIS() 1■ 【 lo nW1 100 n I conauclnt day
我 々のニワ トリヒナお よびCaco-2 BBe細胞 を用いた研 究成果 は、 ビタミン
A生
体 内利用効率やその代 謝 の 恒常性 をについて新 たな基礎 的知見 を掲示で きる と考 えている。5.ま
と め ビタ ミンAは
小腸 の成熟化 や小腸吸収細胞 の増殖 ・分化 において必要不可欠 な生理作用 を有 す る。ニ ワ トリヒナの孵化期 における十二指腸 で は、BCMO l遺
伝 子発現が増大 し、同様 にRALDH l遺
伝子発現 の変 動 も認 め られた。 これは、十二指腸 の機能発達、成熟 のため に βカロテ ンか ら由来す る レチ ノイン酸生成 に必要である と考 え られる。 また、BCMO l発
現 はニワ トリ十二指腸や ヒ トの小腸吸収細胞 をモデル とした caco 2 BBe細胞 において、それぞれハ イ ドロコルチ ゾ ン及 び 島に よ りBCMO l発
現 を克進す るこ とが示 された。
引 用 文 献
1.武
藤泰敏:レチノイド・カロテノイド ー体内代謝と発癌予防―、 1-4、 南山堂、東京、1997。2. Miller M,Hum帥礎yJ,JOlnson E,Marnda E,Brooklneyer R,Katz J:れhy do childttn become vitalnln A deicient?J Nutr 132:2867
S-2880S,2002.
3. Sonlmer A,Dが dson FR:An■ ∝y Accords:Assessment and control of vitamin A deiciency:he Annecy Accords.J Nutr 132:2845S―
2850S,2002.
4。 SilverFnan AK,Ellis CN,Voorhees JJ:Hypervitanlnosis A syndrome:a paradigm of reinoid side erects.J Am Acad Dermatol16:
1027-1039,1987.
5.Rohman Ю,Moore LL,Shger MR,Nguyen US,Manmno S,M unsky A:Teratogenicity of high tamin A intake.N EnglJ Med,
333,1369‐1373,1995.
6. Duing A,Dawson AD,Hamson EHi Carotenoid transport is decreased and expression of he■ pid transPorter SR―BI,NPCILl,and
ABCAl is downregulated in Caco-2 cens treated witt Ezeimibe J Nutr 135:2305-2312,2005
7.Glover tt The conversion ofβ ‐carOtene nto wiamin A Vitam Horm 18:371-386,1960.
8.Goodman DS,Blomstrand R,Wemer B,Huang HS,Shato ■The intesthal absorpion and metabolism of tamin A and β―CarOtene
in rnan.J Chn lnvest 45:1615‐ 1623,1966.
9, Ong DE:A novel retAno卜bind g protein from rat.Pu icaこ on and prartial characttnzation.J Biol Chem 259:1476‐ 1482,1984 10.Crosas B,HyndmaII DJ,Gallego O,Martras S,Pares X,Hylm TG:Human aldose reductase and human smttl intesine aldose reduc―
tase are emcient reinal reductasesi consequences for rednoid lnetabo近 sm.Biochena J 373:973-979,2003.
11.Kakkad BR Ong DE:Reducion of retinaldehyde bound to cellular retinolもinttng pЮtein(type II)by HiCЮ somes hm rat smallin― testine J Biol Chen1 2631 12916‐ 12919,1988.
12. MttDonald PN,Ong DE:Evidence for a lecittin― retinol acyltransferase activity in he rat smttl intestine.J Biol Chem
263:12478-12482,1988.
13.Ong DE,Kakkad B,MacDonald PN:Acyl COA― dependent esttflcation of reinol bound to cenular reinol― binding prottin(type Ⅱ)
by H cЮsomes from rat sman intestine.J Biol Chena 262:2729-2736,1987
14. Harrison EH:Mcchanism of digestion and absorpこ on of detary vitaxm A.Annu Rev Nutr 25:87-103,2005,
15,Zhao D,McCarery R s K」,Ntte RL,Hogan R Chin岬,Drtter UC:Molecular identincatiOn of a maJor retinoic―acid― synhesizing enzyme,a retinaldehyde‐ speciic dehydrogenase.Eur J Biochen■ 15:15-22,1996
16. Chambon P:A decade of lnolecular biology of retinoic acid receptors FASEB J 101 940-954,1996.
17.Levin AA,Sturzenbecker LJ,Kazmer S,Bosakows題 呪HuseltOn C,Allenby G,Speck J,Kratzeisen C,Rosenberger M,Lovey A,
Grippo JF:9-cis reinoic acid stereoisomer binds and act ates the nuclear ttceptOr RXRα .Nature 355:359-361,1992.
18. AⅡenby G,Bocquel岨,Saunders M,Kazmer S,Speck J,Rosenberger M,Lovey A,Kastner R Crippo n Chambon R Levin AA:
Rednoic acid receptors and reinoid X receptors:interactions widl endogenous retinoic acids.Proc Nad Acad Sci USA 90:30‐ 34,1993.
19.Rigmp KM,Ong DE:A retknyl ester hydtolase act ity intrinsic to tte brush border membrane of rat small intesine.Bioche stry 31, 2920∼2926,1992.
20.Leuenberger MS,Engdoch―Jarret C,and Woggo■ WD:The reacion mechaIIIsm of the enzyme―catalyzed centtal cleavage of β―
carotene to retinal.Angew Chenalnt Ed 40,2614-2617,2001.
長崎 県立大 学 看 護 栄養 学 部紀 要 第9巻 (2008)
matography.Anal Biochcn1 241,199-205,1996
22.Duszka C,Groher R Attm EM,Alexandre― Gouわ au MC,BOrel鳥 Aztts一Braesco V:Rat intesinal卜 carOtene dioxygenase“tiviv is located p marily in he cytosol ofmaturejeiun4al enterocytcs.J Nutr 126:2550-2556,1996.
23.BOulanger A,McLemore R Copcland NG,Gilben DJ,Je撤ねs NA,Yu SS,Gendeman S,Redmond TM:Idendacation of β―CarOttnc 15,15'一monooxygenase as a peroxisome pЮliferator―actiTated receptor tareCet gene.FASEI〕 J17:1304-1306,2003.
24.COng X,Tstt SヽⅥ Yan B,Rubin LP:Coopettdon betteen MEF2 and PPARArin human intestmtt β,β Carotene 15,15'― monooxygenase gene expression BMC MOI Bio1 7:7,2006.
25. Lindqvist A,Sharlrin J,sharvill l)E,Andersson S:Loss― of―hnCioa mutaion in carotenoid 15,15'― monooxygenase identifled in a pa― ient、vidl hypercarotenerxlla and hypovitattunosis A J lWutr 137:2346-2350,2007
26.Hessel S,EichngerA,Isken A,AmenguЛ J,Hunzelmann S,Hoeller U,EIste Ч Hunzlkerコ;COrnczyk R,OberhauserⅥ
von Lintlg J, Wyss A:CM0 1 deflciency abolshes vitamnA pЮ duciOn frOm β―carotene and alters llpid metabolism in mice.J Biol Chem 28雰33553 -33561,2007.
27. Bachmann H,Desbarats A,Pa son tt Sedgelvick M,RIss G,Wyss A,Cardlnauit N,Duszka C,Gorttczyk R,GЮ lier P:Feedback
reeOulation ofβ,β―CarOttne 15,15■monooxygenase by reinOic acid in rtts and chickens.J Nur 13多 3616-3622,2002
28.seino、 Mikitt Kiyon H,Abe■ FuiimotOコ;Kimura K,Tattuchi A,Takahash、 OiSO Ч IWanaoCa■ SeinO s:Isx pゼ cipates in
he mttntenance of vit延 n A metabolism by reguladon Of β一Carotene 15,15'― monooxygenasc(Bcmo l)expression.J Biol Chem 283:
4905-4911,2008.
29.山 口範晃:県立長崎シーボル ト大学大学院 修士学位論文 “The study of development gene expression of β―Carottne cleavage
enzyme and retinoic acid signaling in chick duodenu■ l and livei"、 2005。
30.Yamaguchi N,Yamalnoto■ Sumga K,Takase S:D"elopmental changes in gene exprcssiOns of β_carotene cleavtte enzyme and red noic acid synthesizing enzymes in the chck duodenum.Comp Biochem Physiol A Mollntegr Physiol 148:690-697,2007.
31. 激衝maguch N,Suruga K:Tttiodottyronine stimulates CMO l gene expressiOn in human htestinal Caco-2 BBe cens Life Sci82:789-796,2008.
32.Tょase S,Suruga K,Suzuki R,Goda T:Relatlonship bet、 veen peinatal appearance of cellular rcdnOl― binding prote ,type Ⅱ and retl一
nal reductase activity in chick Lve■ Life Sci 58:135-144,1996
33,Taiima s,Goda■ Takase Si CoOrdinated distribuion padems of ttee enη me actlwides i耐 olved in he absOrption and metabolism of β―CarOtene and v並剛In A along he villus― cりpt ttis Of chick duOdenum.Life Sci 65:841848,1999.
34.Taiima s,coda■ Takase Si Co―ordinated inducion of β―Catotene clettage enzyme and reこ nd reductase the duodenum ofthe de―
veloping chicks.Comp Biochenl Physiol B Biochem Mol Biol 128:425-434,2001.
35. Henning SJ:Postnatal development cOOrdinaton of fceding,dlgestiOn,and lnetabottsm Am J Physio1 241:G199-G214,1981. 36. Peterson MD,Moosdor MS:An in vitro model for he anttysis of intesO■ al bmsh border assembly I.Ultrastrucmral analysis of cell
contact―induced brush border assembly in Caco-2 BBe cens,J Cell sci 105:445-460,1993.
37.During A,Abaugh G,SI h JC Jr:Charactenzation of β‐carotene 15,15■ dioxygenasc a飢 ity in TC7 clone of humm intesinal ce■