マダイの品種改良に関する研究
89
0
0
全文
(2) マダイの品種改良に関する研究. 近畿大学水産研究所 家戸敬太郎 (主査:熊井英水教授). S t u d i e sont h eB r e e d i n go ft h eRedS e aBream, Pagrusmajor. K e i t a r oKato. 恥1 a r c h , 2002. F i s h e r i e sL a b o r a t o r yo fK i n k iU n i v e r s i t y ( A d v i s o r :P r o f .HidemiKumai). i n k iU n i v e r s i t y,t of u l f i l lt h e S u b m i t t e dt ot h eG r a d u a t eS c h o o l,K r e q u i r e m e n tf o rt h eD o c t o r a t eD e g r e e ..
(3) Bul . 1F i s h .L a b .K i n k iU n i v .,No. 8 ,1 7 3-2 5 9. ( 2 0 0 2 ). マダイの品種改良に関する研究 * 1 家戸敬太郎. S t u d i e so nt h eB r e e d i n go ft h eRedS e aBream ,Pagrusmajor K e i t a r o KATO*2. 目次. 緒 論 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 7 5. 第 I章. 選択系マダイと非選択系マダイの量的・質的形質およびその飼育. ………………. 1 7 8. ・1.材料および方法 I. … … . . . ・ ・...………・……-……-……・……・……………………………. 1 7 8. 1 2 . 結果および考察. ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ … … … … . . . ・ ・ . . . . ・ ・ . . 180. 第I I章. H. H. 染色体操作による品種改良. H. … … ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・. 1 9 0. I I -1.第二極体放出阻止型および第一卵割阻止型雌性発生二倍体マダイの生残,. . . . . ・ ・ . . . 190 成長および外部形態 ・・……………・・…………・……・……・・………………...・ ・ I I l・1.材料および方法 •.•.•.•.•..•.•..••.•..••.••.•.•....••.•.•.••.•.•.•.••.•.....•.•..•••.••.•..•••..• 190 H. H. H・I ・2 . 結果. ・ ・ ・ … . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 9 3. . 考察 I I l・3. … … . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 199. I I ・ 2 . クローンマダイの作出およびその飼育 ………………………………………………… 200 I I ・ 2・1.材料および方法 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 一 . . . . . . . . . . . . . . . . . . . . . . . . … ・ 一 . . . . . . . . . . . . . . . . . . 200 I I 2・2 . 結果. ••..••••.•.....•.••.•••.••.•.•.•.•....•••.••.•.•.•..•.•..•....•.•.••...•.••.••...•..•..•.•••...•. 202. I I 2 3 . 考察. . . ・ ・ . . … … … … … ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・. 204. H. I I・ 3 . DNAフィンガープリント法によるクローン性の証明. ・ 3・1.材料および方法 I I. ………………………………. 205. ・・……-……・・……-……-…………-……-………………一………. 206. ・3 2 . 結果 I I. 207. I I 3 3 . 考察. 2 1 1. I I・ 4 . クローンマダイの諸形質. ……………・・…・………・……・・・・・・・・…………………………. 2 1 1. . 外部形態 I I 4 -1. ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ … . . . . . . . . . . . . . . . . . . . . . 212 I I ・ 4・ 1 ・1.材料および方法 •..•..•.•••.......•..•....•.••.•.•..••...•.•.••.•.•..•..•..•.••.•.•••..•... 212. I I ・ 4・ 1 ・ 2 . 結果. … ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ー ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ … … . . . . . . . . . . . . . . . . . . 213. 本論文は近畿大学審査学位論文である。 * 2白浜実験場 ( F i s h e r i e sL a b o r a t o r y,K i n k iU n i v e r s i t y ,3 1 5 3,Shirahama,Nishimuro,Wakayama6 4 9 ・ 2 2 1 1,J a p 佃). 事1. -173一.
(4) 近大水研報. (2002). 7 18 1S 18 12 23 23 24 25 2 222222222. n3. G. 成⋮⋮数 組法⋮⋮指法 分ホ⋮⋮隷か ⋮戒び⋮⋮殖び 今よ .. 空よ. ・. 1Vli----. 化おびお 察の料果察ょ料果察. 4z 山 -. 酒田丹斗ぺ. 考附材結考潟材結考 2 3 主1 1 2 ・3 3 包子﹁ 1 ・ I ニ-・2・ 2 2A1333 4 之 4441444 H4HHE4HHE 第I I I章. 8号. マダイの性統御. ……-…………・……・…...・. H. ・ … . . . . . ・. H. ・ . . . . . . . ・. H. ・-………・………・・. …・……………………………………....・ ・-……… 1 1 1 1 .選抜育種マダイの性分化過程 1 1 1 ・ I ・ 1 .材料および方法 . . . . ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ .結 果 …………・・・・・・・・.........................................................・・………...・ ・ . 1 1 1 1・2 .考 察 …・・…・・・・..................................................................................・・ I I I l・3 H. e. H. H. 2 2 7 2 2 7 2 2 7 2 2 8 2 3 1. 3 3 1 1 1 2 .1 7 α ・メチノレテストステロン経口投与による全雄群誘導 ・ ・ ・-…………....・ ・... 2 1 I I 2 -1 . 材料および方法 ……・...................................・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 2 3 3 H. ・2 .結 果 1 I I 2 ・ 2 ・3 .考 察 1 1 1. 第. H. H. …・…一………・・・…・…・・…・・・……一・・……・・一…-…..…・・…・・…………・・…... . . . ・. H. ・ . . . ・ ・…一......…・・・…・・…・…ー・………・...・ ・...一...…・・…ー…-……….. H. H. IV章 クローンマダイの量産と養殖用品種としての展望 …………………………….. 2 3 9 I V -1 .親魚の選定 … ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 2 39 IV-2. 第一卵害a~且止型雑性発生二語体の作出と親魚養成 … ・ ・ ・・ . . . . . ・ ・ … . . . ・ ・-…… 2 4 0 IV ・3 .ホモ接合型クローンの作出とその性統御 …・…...・ ・ … … . . . . . . . . . ・ ・ . . . . . ・ ・ . . . ・ ・ . 2 4 1 H. H. H. H. 要. 約. Summary 辞. 文 献. H. H. H. H. ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・. 2 4 2. ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・. 244. … . . . . . . . . . . . . 一 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . … . . . . . . . . . . . . . . . ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・. 2 4 6. ・ … ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・. 2 4 8. ・ … . . . . ・. 249. 4 .ヘテロ接合型クローンの大量生産 I V. 謝. 2 3 5 2 3 8. a. e. e. H. ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・.
(5) 家戸:マダイの品種改員に関する研究. 緒論. マダイ Pagrusmajor タイ科マダイ韮科マダイ属)は,最大全長が 1mに達する魚で,体は 楕円形で俣1肩,根幅域は隆起し,背鰭蘇はいずれも強国で鋭く,体は紅色で体上半部にコバルト 色の小斑紋が散在,星鰭後縁が黒いという特徴を有し,本邦では北海道東・北部および沖縄を除 く日本各地に分布する九マダイはこのように色が赤くて美しく,体形が整い,日本近海に広く 生息することから,我が国では古くから慶事にはかかせない魚として最も珍重されてきた。 マダイ養殖の歴史は比較的新しく,海面鱗生糞による本格的な養殖が行われるようになったの は1 9 6 5年頃からといわれているヘ 1 9 7 0年には初めてマダ、イの養殖生産量が農林水産統計年表. 6 0 tとわずかで、あった。その後養殖生産量は著しく増大し, 1 9 8 0年 に登場し,同年の生産量は 4 には約 3 0告の 1 4, 7 5 7t ,1 9 9 0年には 5 1, 6 3 6t ,さらに 1 9 9 9年には 8 7, 2 3 2tに も 達 し て い る ヘ こ のようにマダイ養殖が嬢発的に進展した背景には,種苗生産技術の向上による人工撞苗の安定供 給と質の向上があげられる。種菖生産尾数は, 1 9 8 5年には約 1 . 10 8万尾であったものが 1 9 9 5年. 0信を越える 1{ 意1 , 8 4 8万尾が生産されるようになったヘしかし,近年では不況,魚 にはその 1 病の蔓廷,さらには生産過剰による市場価格の低迷などによって,マダイの養殖経営は議しい状 況に陥り,これを打開するためコスト削減や商品の差別化による市場缶格の向上に繋がるより優 れた形質を備えた養殖品種の確立が望まれている c マダイは,我が留における代表的な養殖魚種 で、あって,人工種菖生産技術もほぼ確立されており,この点は品種改良を図る上から有利な魚種 と考えられる。 マダイの品種改良に関する研究として,近畿大学水産研究所において 3 0年間以上に亘る選抜 育種が行われ,成長の速い系統が作出されている ( M u r a t ae ta l .4 ) )0 近年,本系統は西宮本各地 で養殖品種として用いられているが,こうした中で,選抜育種されたマダイの高成長をもたらし ている具体的要国や,また,非選抜の通常マダイとの各種形質の異罰を明らかにすることは更な る品種改良を考えた場合に重要になると考えられるが,これまでのところ選抜脊種マダイと非選 抜マダ、イとの比較に関するデータは少ない。 新たな品種改良の手段として注目され, 1 9 8 0年代以降活発に研究されているものに染色体操 作があるヘ染色体操作とは,第一卵割あるいは第二減数分裂の祖止および配偶子の遺伝的不活 性化により,染色体の数と組み合わせを統御する技街体系をいうもので,魚類においても信数体, 雄性発生二倍体,雄性発生二倍体などが作出されている ι4ヘマダイにおいても,不妊化に伴う 成長の向上,二次性徴(体色黒化)の発現による商品錨値低下の防止,細胞体積の増大に伴う体 重の増加などが期待され三倍体が作出されたが,成長の向上はみられず,また,不妊化はしたも ののそのほとんどが雄になり二次性徴が現れるなど養殖品種としての有用性は認められていな 3 1,山間の目 い 21,へ一方,雄性発生二倍体は,全雌生産町内側および短期間での近交系作出 30, の 的で作出されてきた。雑性発生二信体は,遺伝的に不活性化した精子で卵を媒精後,第一卵害j 担止あるいは第二極体放出の阻止により作出される牧刻。前者では雑親由来の 1セットの染色体 を体細胞分裂の睦止により倍化するのですべての遺伝子産でホモ接合型となる値捧が作出され, わずか 2世代でクローンを得ることができる。一方,後者では減数第一分裂時の遺伝子座-動原 体関の組換え率によって部分的にヘテロ接合型を示す個体が生じ,これらの個体はクローン作出. -175一.
(6) 近大水研報. 8号. (2002). のための親魚として用いることはできない。しかしながら. この第二極体放出阻止型の雌性発生. を 2世代あるいは 3世代繰り返すことにより遺伝子型は完全に同一ではないが固定化が進み,ク 5 5 ) ローン化した実用的な近交系が得られるぺ雌性発生二倍体は多くの魚種において作出され 49,. それらに関する総説 49, 5 3 ・ 5 5 ) も多くみられるが,その作出方法に関してはまだ検討すべき多くの問 題を残している。また,長期間の飼育に基づいて雌性発生二倍体の成長や生残率を調べた報告は ほとんどみられず,海産魚において,雌性発生二倍体の飼育に関する報告がなされているのはヒ i v a c e u s仰 7, 48) のみである。従って,今後は養殖対象魚種の雌性発生二倍体 ラメ ,P a r a l i c h t h y s o/. についての実用的な情報を蓄積する必要があると考えられる。雌性発生二倍体マダ、イの作出方法 についてもこれまでに報告されている州 1) が,雌'性発生二倍体マダイの成長,生残,外部形態な ど養殖用品種としての評価に関する報告はまだ皆無といえる。 クローンは,免疫学,内分泌学,発生生物学,分子生物学および遺伝学において重要な実験動. ヘ. 5 物である。また近年,魚類において養殖用品種としてのクローンの利用が注目されている 45,. ホモ接合型クローン(以下,ホモクローンと略記)は,ホモ接合型の雌性発生二倍体雌より得た 卵から第二極体放出阻止型の雌性発生二倍体を作出することにより得られ. ヘテロ接合型クロー. ン(以下,ヘテロクローンと略記)はホモ接合型雌性発生二倍体の雌雄を交配することにより得 られる。ヘテロクローン集団はヘテロ接合性を保ちながら遺伝的には均一で、あるので,表現型分 散の減少や発育安定性の向上が期待される仰い九染色体操作によって,淡水魚では,ゼブラフイツ シュ Danior e r i oペ メ ダ カ 0 η' z i a s/ a t 伊ω 2ペ ア ユ P / e c o g / o s s u sa l t i v e l i s14),コイ C y p r 的u sc a r p i o5ヘ ,. アマゴ O n c o r h y n c h u sr h o d u r u s50),ニジマス O n c o r h y n c h u sm y k i s s58) およびティラピア O r e o c h r o m i s n i l o t i c u s仰 9) のクローンが作出されている。しかしながら,海産魚でクローンが作出されている のはこれまでのところヒラメ ,P a r a l i c h t h y sp l i v a c e u s仰 8) のみである。 染色体操作,特に雌性発生技術により,優良形質を固定した改良品種を大量生産するには,ニ. i 1 v e rb a r b,P u n t i u sg o n i o n o t u s削などで試みられたような性統御が必要と ジマスベヒラメ制, s なる。また,マダイの雄は産卵期およびその前後に二次性徴により体色が黒化し商品価値が低下 する 2) ので,マダ、イ養殖には優れた経済形質を備えた全雌集団の作出が有効と考えられる。しか しながら,マダイの性統御に関する研究はまだみあたらない。性統御を行う場合には,対象魚の 性決定機構を知る必要があるが,マダイは異型染色体をもたず 6へどのような性決定様式をもつ のか明らかではない。性統御の 1つである雄化技術の手法を確立すると通常卵と雄化雌精子との 交配が可能となる。得られた魚の性比の解析によって本種の遺伝的性決定機構を決めることが可 能かどうかが分かり,性決定機構が遺伝による場合にはホモ接合型がどちらであるかを識別する のに有効である仏へ性決定機構が雌ホモ型である場合には,性転換雄が一度成熟すれば,その 精子と通常雌の卵との受精により全雌群が得られる ω,件ぺ一般に雌雄異体性の魚類の性転換は, 性分化の期間中およびその前後を通じて性ステロイドを投与することにより誘導されるぺ従っ て,性統御技術を確立する場合にはその基礎的知見として性分化過程を知る必要がある。 Matsuyamae ta / .ω) は,マダ、イ天然集団の生殖腺発達,性成熟および性比について調べ,マダ、イ. は雌雄同体となる稚魚期をもっ雌雄異体性魚であることを報告しているが,解化からの成長に伴 う性分化の詳細については調べられていなし、。また,飼育条件下におけるマダイの成長は選抜育 種系統の方が非選抜系の天然集団に比べ著しく速い 4) が,この選抜育種系統のマダ、イの性分化に ついて調べた例はまだない。. -176一.
(7) 家戸:マダ、イの品種改良に関する研究. 以上の観点から,本研究では,より優れた形質を備えたマダ、イの養殖用品種確立のための基礎 的および応用的知見の集積を試みた。 第 I章では, 30年間に亘り選抜育種されてきたマダイおよび選抜育種されていないマダイを 用いて,両者の遺伝的変異および外部形態を比較するとともに,異なる給餌条件で両系統を飼育 し,成長,飼料効率などを比較し,選抜育種マダ、イの特性を明らかにした。 第I I章では,新たな品種改良の方法として染色体操作を用いて,雌性発生二倍体の作出およ. びそれを用いたクローンの作出について検討するとともに,それらの養殖魚としての特性を評価 した。 第I I I章では,性統御に関する知見を得ることを目的として,選抜育種マダ、イの性分化過程に. ついて調べるとともに. 性ホルモン処理による全雄群誘導の方法について検討した。. 第 IV章では, 1-I I I章での結果を踏まえ,クロ)ンマダイの量産とその養殖用品種への応用. に関する問題点と展望について述べた。. -177一.
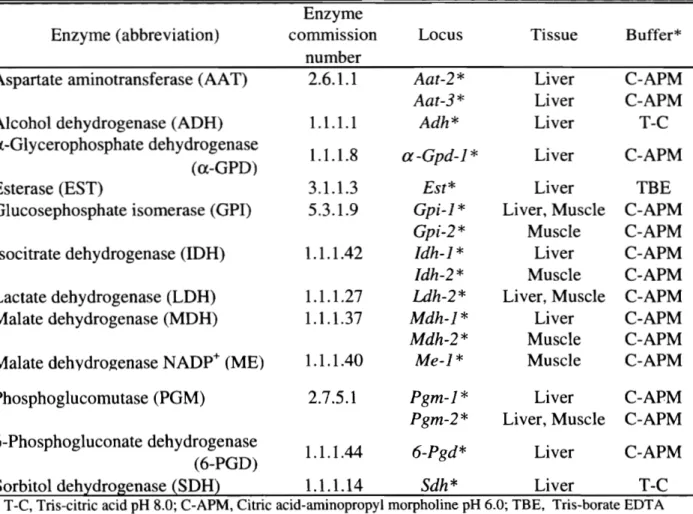
(8) 近大水研報. 第 I章. 8号. (2002). 選択系マダイと非選択系マダイの量的・質的形質およびその飼育制. Muratae ta l .4)は,近畿大学水産研究所において 30年間以上に亘るマダイの選抜育種を行い,. 高成長系統の作出に成功している。この選択系マダイにおいては,高成長に貢献する何らかの遺 伝的要素が固定されていると考えられるが,選択系と非選択系との問の成長差について詳細に調 べた報告はまだない。そこで本研究では,選択系と非選択系マダイとの遺伝的変異,外部形態, 成長,飼料効率などについて比較した。. I ・1.材料および方法. (1)供試魚. 6および 7世代目の選抜育種マダ‘イ親魚 4) ( 19 9 1年産雌雄各 1 4尾)および瀬戸内海(明石周辺) で捕獲された天然親魚(推定 5 7歳魚 1 0 4尾,平均魚体重 1 . 2 6k g ) から,それぞれ自然産卵に より得られた卵を用いた。 1 9 9 5年 6月 8日に解化した両系統を近畿大学水産研究所大島分室で 約 6ヶ月間飼育した後,同白浜実験場に輸送して実験に用いた。 ( 2 ) 遺伝的変異. 0尾ずつの肝臓および筋肉の解凍ドリップを用いた。電気泳動, アイソザイム分析には両系統 6 a n i g u c h iandSugamaη) 染色および電気泳動ノミターンの解読については谷口および岡田川,並びに T. の方法に従った。検出した 1 2酵素,遺伝子座,検出に用いた組織および電気泳動に用いた緩衝 a b l e1 ・ 1に示した。電気泳動の結果に基づいて, 1 7遺伝子座を Shakleee ta l .73) に従い命名 液を T. した。両系統が HardyWeinbergの法則刈 l こ基づいて推定した遺伝モデ、ルに適合するかどうかを 調べるために,各遺伝子座の遺伝子型の観察値と上記の法則に従って算出した期待値との聞のど 検定を行った。また,変異性の指標となる 1遺伝子座当たりの平均対立遺伝子数,多型的遺伝子 座率および平均ヘテロ接合体率を比較した。さらに,対立遺伝子頻度の異質性に関するど検定 を行い両系統間で比較した。 ( 3 ) 外部形態. 外部形態の測定には両系統 32尾ずつを用いた。計数的形質として,背鰭,腹鰭および尻鰭の 赫および軟条数,胸鰭および尻鰭の軟条数,並びに側線上方横列鱗数および側線下方横列鱗数を それぞれ計数した。また,計測的形質として,全長,尾文長,標準体長,頭長,吻長,眼径,虹 門前長,両眼間隔,上顎長,背鰭前長,体高,体幅,尾柄長,尾柄高,背鰭および尻鰭基底長, 胸鰭長および腹鰭長をそれぞれ測定した。選択系と非選択系とのプロポーションを比較するため に,両系統から平均標準体長がほぼ等しくなるように 1 0尾ずつを選び 1 7形質の標準体長に対す る比を算出した。各形質の計数および計測は中坊の方法制 l こ従って行った。. -178一.
(9) 家戸:マダイの品種改良に関する研究. Table1 1 .Th el i s to fenzymesexamined, l o c id e t e c t e d, t i s s u ea s s a y e dandb u f f e rs y s t e m su s e d f o re l e c t r o p h o r e s i so fr e ds e abreams a m p l e s 互nzyme( a ちb r e v i a t i o n ). A s p a r t a t ea m i n o t r a n s f e r a s e(AA T). Enzyme commlss1on number .1 2 . 6 .1. Locus. T i s s u e. B u f f e r *. 1 . 1 . 1 .1. A a t 2 * A a t 3 * Adh*. L i v e r L i v e r L i v e r. C-APM 五4 C-AP τ-C. A l c o h o ld e h y d r o g e n a s e(ADH) α G l y c e r o p h o s p h a t ed e h y d r o g e n a s e (α-GPD) E s t e r a s e(EST) 予h o s p h a t ei s o m e r a s e( G P I ) G l u c o s e. 1 . 1 . 1 .8. a-Gpd-l*. L i v e r. C APM. 3 .1 . 1 .3 5. 3 . 1 .9. I s o c i t r a t ed e h y d r o g e n a s e(IDH). 1 . 1 . 1 .42. L a c t a t ed e h y d r o g e n a s e(LDH) M a l a t ed e h y d r o g e n a s e(MDH). 1 . 1 . 1 .27 1 . 1 . 1 .3 7. M a l a t edehydro~enase NADP+(ME). 1 . 1 . 1 .40. E s t * G p i 1 * Gpi ・2 * I d h 1 * I d h 2 * Ld h 2 * Mdh-1* Mdh-2* Me・1*. L i v e r L i v e r,Muscle Muscle L i v e r Muscle L i v e r,Muscle L i v e r Muscle 民1 u s c 1 e. TBE C-APM C-APM C-APM C-APM C-APM C-APM C-AP ちf C-APM. P h o s p h o g l u c o m u t a s e(PGM). 2 . 7 . 5 . 1. Pgm-l* Pgm-2*. C-APM L i v e r L i v e r ,M uscle C-APM. 由. 6 P h o s p h o g l u c o n a t ed e h y d r o g e n a s e 1 . 1 . 1 . 44 6-Pgd* L i v e r C-APM (6-PGD) S o r b i t o ld e h y d r o g e n a s e(SDH) 1 . 1 . 1 .1 4 Sdh* L i v e r T-C *TC , T r i s c i t r i ca c i dpH8 . 0 ;C-APM, C i t r i ca c i d a m i n o p r o p y lm o r p h o l i n epH6 . 0 ;TBE ,T r i s b o r a t eEDT A p H 8 . 9 . ( 4 ) 或長比較 6ヶ月齢の両系統 326尾ずつをそれぞれ近畿大学水産研究所白浜実験場地先の海面講生賓 ( 3. X3X3m) ~こ収容し,市販のマダイ用配合館料を給餌して 8 ヶ完関飼育した。飼育開始時の魚 体重 ( g,平均±標準偏差〉は,選択系では 5 9 . 2: : t9 . 2,非選択系では 3 6 . 1: : t6.4であった。魚体 重の測定は 1ヶ丹あるいは 2ヶ月目毎行った。. ( 5 )飼育試験 飼育実験では次のような 4試験区を設定した。試験区上選択系飽食給餌医;試験区 2,選択 系制設給餌区;試験区 3,非選択系飽食給餌区;試験区 4,非選択系制捜給餌区。選択系および 2尾ずつを FRP 非選択系の試験開始時の平均体重はそれぞれ 432gおよび 260gで、あった。各区 1 3 製 2m 容水槽にそれぞれ収容し 8週間流水飼育した c 飼育水温は 1 6 2 3Cの範囲で、あった G 飼 0. 料には市販のマダイ用配合飼料(桓タンパク質含量, 54%;粗脂質含量, 17%) を用い. 1日 1. 回試験区 1および 3~こは飽食量を給餌し,試験区 2 および 4 は両区の給餌率が等しくなるように 魚体重の 0. 3-0.9%の給餌率で給餌した。体重測定を 2週間吾毎に行い,各区の増重率,飼料効 率およびタンパク質効率を算出した。試験開始時および終了時に各区 3~ ずつの全魚体の一般成. 分組成を測定し,見かけのタンパク質および脂質の蓄積率を算出した。飼育試験終了後,各区の.
(10) 近大水研報. 8号. (2002). 残りの魚に 0 .5%の Cr203を含む飼料を試験期間中と同様の条件で給餌した。摂餌 6時間後にお ける胃組織および内容物のペプシン様酵素活性. 並びに腸内容物のトリプシン様酵素およびアミ. ラーゼ活性をそれぞれ測定した。酵素活性は,組織あるいは内容物 Ig当たり,魚体重 1 0 0g当 たりおよびタンパク質 1mg当たりの u n I tで表示した。また,摂餌 6時間後の腸内容物および糞 の見かけのタンパク質および糖質の消化率も測定した。一般成分,消化酵素活性および Cr203は , それぞれ吉中および佐藤 76),Kawai and I k e d a77,沌)および福原および塚原閣の方法に従って測定 した。. I ・ 2 . 結果および考察. (1)遺伝的変異 検出した 1 2酵素 1 7遺伝子座のうち変異のみられた遺伝子座は,選択系で 6遺伝子座,非選択. 1遺伝子座で、あった。それぞれの遺伝子座における各対立遺伝子頻度および遺伝子型頻度 系で 1 a b l e1 ・ 2に示した。遺伝子型頻度の観察値と期待値との差の の観察値と期待値との差のど値を T Weinbergの法則に ど検定では,両集団ともいずれの遺伝子座にも有意差は認められず, Hardy-. 基づく遺伝モデ、ルとしての適合性が示された。 次に,両系統の変異性の指標として, 1 7遺伝子座における 1遺伝子座当たりの平均対立遺伝 子数,変異遺伝子座率,多型的遺伝子座率および平均ヘテロ接合体率を算出して T a b l e1 ・3に示 した。 1遺伝子座当たりの平均対立遺伝子数は,非選択系の値が顕著に高かったロまた,変異遺 伝子座率 ( p )においても,非選択系は選択系において単型で、あった αG p d J * ,G p i J *,I d h J * ,. M d h J *および S d h *の 5遺伝子座で変異が認められ,非選択系が高かった。しかし,最大対立 遺伝子頻度が 0 . 9 5以下の多型的遺伝子座率 ( p * )では逆に非選択系の方が低い値を示した。一方, H o )では両系統聞に顕著な差異はみられず 平均ヘテロ接合体率の観察値 (. 平均ヘテロ接合体率. ! H e )では,非選択系の方がわずかに低い値を示した。 の観察値と期待値との比 (Ho さらに ,A a t 2 *,A d h *,M e J , * E s t * および S d h *の 5遺伝子座において選択系と非選択系との T a b l e1 -4)。その結果,両系統聞には 聞の各対立遺伝子頻度の異質性に関するが検定を行った ( 1 2対立遺伝子に有意差が認められ,特に A a t 2 * 1 0 0,A d h * 1 0 0,A d h * J 3 0,M e l * J 2 0,E s t * J 3 0. および E s t * Xの頻度に著しい差異がみられた。 以上の結果から,選択系と非選択系との聞には 1遺伝子座当たりの対立遺伝子数および変異遺 伝子座率において顕著な差がみられ,選択系における低頻度遺伝子の消失が示唆され,それぞれ の親魚集団のもつ遺伝子プールには大きな量的差異があるものと考えられる。一方,そのような 保有対立遺伝子数の違いにも関わらず,平均ヘテロ接合体率の観察値 (Ho)に両系統間で差異が みられなかった事実は,選択系における非常に高い異型接合性の存在を示している。谷口ら制 は,選択系マダイにおける 1遺伝子座当たりの平均対立遺伝子数および多型的遺伝子座率の低下 を報告しており,これらの遺伝的変異の低下は主に低頻度対立遺伝子の消失によるものであり, 再生産に係わった親魚数が少ない時に起こるピン首効果によるとみている。本研究における選択 系の遺伝的変異の特徴は谷口ら制の報告と類似していた。また, 5遺伝子座においてみられた対 立遺伝子頻度の異質性より,両系統聞の遺伝的な類似性は極めて低いことが示された。. -180一.
(11) 家戸:マダイの品種改良に関する研究. 言 ztzt I I. 0 0 0 0 5 5 寸2 χ一 0 002χ -AUAW/ 3210 ,98x ,. *. e ' a g s E. Table1 2 .A l l e l ef 詑弓u e n c i e sa tp o l y m o r p h i cl o c iandc h i s 司u a r ev a l u e sf o r d i f f e r e n c eち 己t weeno b s e r v e dande x p e c t e df r e q u e n c i e so fg e n o t y p e s o ft h es e l e c t e dandn o n s e l e c t e ds t r a i n so fr e ds e abreams 持o n NonS e l e c t e d S e l e c t e d s e l e c t e d s e l e c t e d l l e l e l l e l e Locus A Locus A S紅 白n s t r a l n s t r a l n s t r a l n 9 9 2 l 120 0 . 0 0 8 Aat ・2 * 1 0 0 0 . 8 4 2 一勺. a h T * O 0 . 1 5 8 100 2 0 0 0 . 9 9 2 0 . 0 0 8 χ 2 χ 2 0 . 2 3 0 . 0 0 0 . 0 0 0 . 1 1 7 0 . 2 5 8 Mdh-1* 100 0 . 9 8 3 6 0 Adh* 1 0 0 0 . 2 8 3 0 . 5 1 7 80 O 0 . 0 1 7 2 0 . 0 2 1 3 0 0 . 2 2 5 0 . 6 0 0 χ χ Z 0 . 2 8 Me-1* 120 0 . 1 7 8 0 . 3 9 8 0 . 1 0 100 0 . 4 7 4 0 . 3 4 7 α-Gpd-1* 120 0 0 . 0 2 5 0 . 2 1 2 90 0 . 2 2 9 0 . 9 7 5 100 Z 2 0 0 . 1 1 9 0 . 0 4 2 χ 0 . 0 4 χ 2 2 . 6 0 0 . 1 2 5 0 3. 4 8 0 . 0 4 2 Pgm-1* 110 0 0 . 0 0 8 0 100 0 . 8 4 2 0 . 8 5 8 0 . 0 8 3 0 . 0 4 2 90 0 . 1 5 8 0 . 1 1 7 0. 0 . 3 8 1 4 08 0 . 0 1 7 80 0 0 . 1 6 9 0 . 0 4 2 χ 2 2 . 1 2 3 . 6 4 0 . 2 4 6 0. 3 42 120 0 0 . 1 8 4 0 . 1 1 9 6-Pgd* . 1 5 8 0 0 . 8 0 8 100 0 2 . 3 8 3 . 7 5 . 8 4 2 在宿志 0 . 0 0 8 O 0 80 G p i 1 * χ 2 2 . 1 0 0 . 0 3 0 . 9 8 3 0 . 0 0 8ー でS d J 百 * O 0 . 0 4 2 O 250 1 0 0 0 . 9 5 8 0 . 0 2 χ z 0 . 1 1 *R a r ea l l e l e s( 12 5,1 1 5,90,80a n d75)i nEst*l o c u sw e r ep o o l e dt oX. ヲ t. I. Table1 3 .G e n e t i cv a r i a b i l i t yo ft h es e l e c t e dandn o n s e l e c t e ds t r a i n so fr e ds e a breamsb a s e don1 7l o c io fisozyme Average Num ちe r numbero f o ff i s h a l l e l e sp e r l o c u s. P. p*. Ho. S e l e c t e d 0 . 3 5 3 0 . 3 5 3 0 . 1 6 4 1 .706 60 s t r a l n Nons e l e c t e d 0 . 6 4 7 0 . 2 9 4 0 . 1 6 2 2 . 5 2 9 60 s t r a l n P :P r o p o r t i o no fp o l y m o r p h i cl o ci . P * :P r o p o r t i o no fp o l y m o r p h i cl o c i( m a x i m u ma l l e l ef r e q u e n c y<0 . 9 5 ) . H o :A v e r a g eh e 詑r o z y g o s i t yo b s e r v e d . H e :Av e r a g eh e t e r o z y g o s i t ye x 予e c 記d . 圃. He. lH e Ho. 0 . 1 6 0. 1 .030. 0. 167. 0 . 9 7 1.
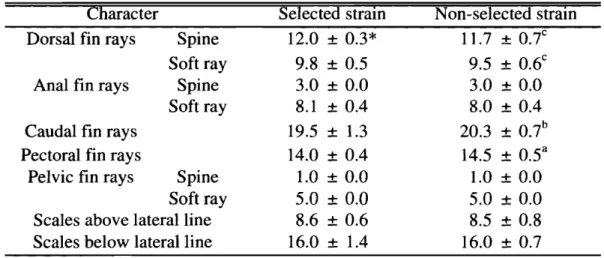
(12) 近大水研報. 8号. (2002). Table1 ・ 4 .C h i s q u a r ev a l u e sofh e t e r o g e n e i t yt e s tf o ra l l e l e おq u e n c i e sa t5polymorphicl o c ibetweens e l e c t e d f andn o n s e l e c t e ds t r a i n sofr e ds e abreams Locus A a t 2 *. A l l e l e 1 0 0. 主. χvalues. 1 7 . 6 7 3a b 7 . 9 0 4 a 1 3 . 6 1 1 a 3 4 . 8 1 6 1 3 . 9 6 5a C 3 . 9 3 8 0 . 0 9 9 4 . 3 6 3C a 1 5 . 7 4 2 c 5 .1 9 4 1 .69 0 . 1 8 1 1 0 . 3 4 1b 2 . 6 3 6 1 5 . 1 2 7a C 5 . 1 0 6. 6 0 1 0 0 1 3 0 120 Me-l* 100 90 2 0 130 E s t * 120 110 100 95 85 X* s ' d h * 1 0 0 a . b . cS i g n i f i c a n t l yd i f f e r e n tb e t w e e ns e l e c t e da n dn o n s e l e c t e ds t r a i n s Adh*. ( a , pく 0 . 0 0 1 ;b, pく 0 . 0 1 ;c , pく 0 . 0 5 ) . ネ. R a r ea l I e l e s( 12 5,1 1 5, 90, 80a n d75)i nEst*l o c u sw e r e予o o l e dt oX.. ( 2 ) 外部形態 選択系と非選択系との計数的形質を比較して T a b l e1 ・5に示した。選択系の背繕蘇および軟条 数は非選択系よりも有意に高く,逆に尾鰭軟条数および臆鰭軟条数は非選択系の方が高かった c. Table1 ・ S.Com 予a r i s o nofm e r i s t i cc o u n t sbetweens e l e c t e dandn o n s e l e c t e d s t r a i n sofr e ds e abreams C h a r a c t e r S e l e c t e ds t r a I n N o n s e l e c t e ds t r a I n c 1 2 . 0: f :0 . 3 * S p i n e 1 1 .7: f :0 . 7 Dors a 1f i nr a y s c S o f tr a y 9 . 8: f :0 . 5 f :0 . 6 9 . 5: Analf i nr a y s S p i n e 3 . 0: f :0 . 0 3 . 0: f :0 . 0 8 . 1: f :0 . 4 S o f tr a y f :0 . 4 8 . 0: Caudalf i nr a y s 1 9 . 5: ! :1 .3 2 0 . 3: f :0 . 7b a P e c t o r a lf i nr a y s 1 4 . 0: t0. 4 1 4 . 5: t0 . 5 S p i n e P e l v i cf i nr a y s 1 .0: f :0 . 0 1 .0: ! :0 . 0 S o f tr a y 5 . 0: t0 . 0 5 . 0: t0 . 0 S c a l e sabovel a t e r a l l i n e 8 . 6: t0 . 6 t0 . 8 8 . 5: 1 6 . 0: f :1 .4 S c a l e sbelowl a t e r a l l i n e t0 . 7 1 6 . 0:. *Mean: t :SD( n = 3 2 ) . S i g n i f i c a n t 1yd i f f e r e n tf r o mt h ev a l u e so fs e l e c t e ds t r a i n( a , pく0 . 0 0 1 ;b , p<O.OI;c ,. . b . c. 邑. p < 0 . 0 5 ) .. -182-.
(13) nF3. Q0. 且 &Et. A a r. ρ i v e E E ' F i v. ・ AU. Qリ. 、.・・. e a ρ L V. 冒. n o. G. n. JU. n a. r ・. )3. 3C. A 剛﹄ト. r 、L す・且. 且 4E ・ 且 4. & 0 一 85291741328372973 ・ ・ h7 83211711547013910. e2 dden WInse--n. 記 n km 一 デ52697090235572814 ιιL 仏 仏 4 仏 1 2 1 1 l 仏 11.2.Z 討はま FIJu-o e 一十一+一+一+一+一+一+一+一+一+一+一+一+一+一+一+一+一+一. 目. = 一. 陀 ほ 芝一 1 0 3 1 1 6 1 1 4 4 1 2 1 5 1 2 2. 且. A. 守 且. ,、 、 . , ノ = e ヨ = r nu--叩m =一 /Et=e9sU- 一 mm= 一. bbmk 凶印刷化問附刷付加問取. P. m苛 印 時 m が ee LL Le L. 白h. LZFZ払 む2 宮山m. 向山一. 一削一切加的. 割. mb. -泊. 加. ω一 ⋮⋮忠則仰いおにおお M山 M口. 引約一. KLg akD T. x. 家戸:マダ、イの品種改良に関する研究. N o n s e l e c t e ds t r a i n 1 3 0 . 9: t5 . 2 .9 : t5. 3 1 21 1 0 3 . 5: t4 . 3 t1 . 2 3 2 . 5: 11 .7 : t0 . 7 t0 . 5 1 0 . 9: 6 8 . 1: t4 . 5 11 .1: t0 . 8 11 .0 : t1 .0 44. 4: t1 . 2 4 3 . 3: t2 . 6 1 7 . 2: t0 . 9 2 0 . 2: t2 . 3 11 .7 : ! :0 . 6 ! :3 . 9 5 4 . 5: ! :1 .7 2 0 . 0: 2 5 . 8: ! :2 . 3 21 .2 : t0 . 6. *Mean:!:SD( n = 1 0 ) . Table1 7 .Comparisooo fp r o p o r t i o o a lmeasurementsbetweens e l e c t e dand000s e l e c t e ds t r a i n so fr e ds e abreamse x p r e s s e da sar a t i ot ot h es t a n d a r d l e n g t h Nons e l e c t e ds t r a i n S e l e c t e ds t r a i n 1 .265 : ! :0 . 0 3 0 a 1 . 2 2 8: t0 . 0 2 5 * T o t a l l e n g t h a ! :0 . 0 2 2 1 . 1 7 9: 1 .1 4 8: ! :0 . 0 2 2 Forkl e n g t h 0. 31 4: t0 . 0 1 1 0 . 3 1 2: t0 . 0 1 1 Headl e n g t h 0 . 1 1 2: ! :0 . 0 0 6 t0 . 0 0 6 S n o u tl e n g t h 0 . 1 1 5: 0. 105 : t0 . 0 0 6 0. 108 : t0 . 0 0 6 Eyed i a m e t e r 0 . 6 5 2: t0 . 0 2 6 t0 . 0 2 2 0 . 6 5 4: P r e a n a l l e n g t h 0 . 1 1 0: ! :0 . 0 0 5 0 . 1 0 7: t0 . 0 0 6 I n t e r o r b i t a lw i d t h : t 0 . 0 0 8 0 . 1 0 8 0 . 1 0 7: t0 . 0 0 9 Lengtho fu p p e rjaw 0. 427 : t0 . 0 1 2 P r e d o r s a ll e n g t h 0. 438 : t0 . 0 1 2 0. 41 6: t0 . 0 2 4 0. 427 : t0 . 0 1 6 Bodyd e p t h 0 . 1 7 1: ! :0 . 0 0 8 0 . 1 6 6: ! :0 . 0 0 5 Bodyw i d t h 0 . 1 9 3: ! :0 . 0 2 2 0 . 1 9 7: ! :0 . 0 1 7 L e n g t ho fc a u d a lp e d u n c l e 0. 11 3土 0 . 0 0 3 0. l1 3: t0 . 0 0 5 Deptho fc a u d a lp e d u n c l e 0 . 5 3 0: ! :0 . 0 2 2 t0 . 0 0 9 0 . 5 1 4: Lengtho fd o r s a lb a s e Lengtho fa n a lb a s e 0 . 1 9 2: t0 . 0 1 3 0 . 1 9 4: t0 . 0 1 5 a L e n g t ho fp e c t o r a lf i n 0 . 2 1 0: t0 . 0 2 4 t0 . 0 2 2 0 . 2 4 9: 0 . 1 9 7: t0 . 0 2 8 0 . 2 0 4: t0 . 0 0 4 Lengtho fp e l v i cf i n *Mean:!::SD( n = 1 0 ) . aS i g n i f i c a n t 1yd i f f e r e n tf r o mt h ev a l u e so fs e l e c t e ds t r a i n(pく0 . 0 1 ) . 曲. -183一.
(14) 8号. 近大本研報. (2002). 平均標準体長がほぼ等しくなるように選んだ両系統 1 0尾ずつの計測的形質の測定檀および標 準体長に対する 1 7形質の比を T a b l e1 ・6および T a b l e1 7にそれぞれ示した。非選択系における 標準体長に対する全長,尾叉長および牌鰭長の比は選択系よりも有意に高かった。これらの結果 は,選択系の尾態および絢鰭が非選択系のそれらよりも小さいことを示している。魚類において 尾鰭および腕鰭はそれぞれ遊泳の推進および制御のために用いられるへ従って,選択系の遊泳 能力は非選択系よりも劣るのかもしれない. Q. ( 3 )成 長 海面縞生賛における 8ヶ月間の飼育期間中の生残率は選択系 67.7%,非選択系 63.0%で、あった。 餌育期間中の両系統の平均体重の推移は, F i g .1 1,こされるように飼育期間を通して選択系が非 選択系よりも有意に高かった。. 4 0 0 S e l e c をe ds t r a i n. 一 ー -Non-selectedsrain ま. 一 -. 3 0 0 CD. s CD ω 津. ぎ200 a. ω. CD c ' : s s 圃. Q , i. >. 4宅. 1 0 0. ︽U. F.O.@@. ∞.@@ F.. F.h ・ . @ 。. F.@.@@. F.6.60. F. 守.@@. F. 約.@@. 。. F.mw.@. でで@@. F.mwF.mw@. Feedingp e r i o d F i g .1 ・1 .C o m p a r i s o nofgrow 由 b e t w e e ns e l e c t e da n d n o n s e l e c t e ds 泊 i n so fr e ds e ab r e a m s .. ( 4 )飼育誤験. s適関の飼育試験の結果をまとめて Table1・8に示した。飽食給餌区において,選択系の日間 摂餌率は 0.94%と非選択系の 0.80%よりも高かった。制限給餌区においては設定どおり選択系お よび非選択の給餌率がいずれも 0.57%と等しかった。選択系の増重率は飽食および鰐眼給餌区の. -184ー.
(15) 家戸:マダ、イの品種改良に関する研究. Table1 ・ 8 .Resultsoffeedingexperimentfor8weeks S e l e c t e ds t r a i n S a t i a t i o n R e s t r i c t e d f e e d i n g f e e d i n g. Non-selecteds t r a i n S a t i a t i o n R e s t r i c t e d f e e d i n g f e e d i n g. Av e r a g ef i o d yweight( g ). 4 2 9 . 5: t 6 0 . 2 * 1 5 5 8 . 8: t 7 0 . 1 Dailyf e e di n t a k e( % ) * 2 0 . 9 4 Weightg a i n( % ) 3 0 . 0 Feede f f i c i e n c y(% ) * 2 5 7 . 8 r o t e i ne f f i c i e n c yr a t i o 1 . 0 7 P' I n i t i a l F i n a l. 4 3 3 . 7: t 4 8 . 0 t 6 2 . 3 5 5 4 . 7: 0 . 5 7 2 7 . 9 8 9 . 9 1 .6 6. 2 6 4 . 8: t5 4 . 3 : t 5 0 . 5 3 2 2 . 4 0 . 8 0 21 .7 5 0 . 8 0 . 9 4. 2 5 4 . 3: t 4 6 . 6 3 0 7 . 7: t5 2 . 5 0 . 5 7 21 .0 6 9 . 6 1 .2 9. *1M ean : tSD( n = 1 2 ) .. * 2Dryd i e tb a s i s . 80. 70. 尻 u. nwnunununu. a w. ( ま)35EgESω ﹄冨ω﹄豆島司. a. ﹃. qu. 内,h. 10. o. SS. SS. NS. NS. S a t i a t i o n R I 回t r i c t i o n S a t i a t i o n R e s 廿i c t i o n. P r o t e i nr e t e n t i o n. L ip i dr e t e n t i o n. F i g .1 ・ 2 .C o m p a r i s o no fa p p a r e n tp r o t e i na n d1 i p i d r e t e n t I o nr a t eb e t w e e ns e l e c t e ds t r a I n s( S S ) a n dn o n s e l e c t e ds 位a i n s( N S )o fr e ds e a b r e a m s . いずれにおいても非選択系よりも高く,また,両系統において制限給餌区よりも飽食給餌区の方 が増重率はわずかに高かった。飼料効率およびタンパク質効率はいずれも選択系制限給餌区>非 選択系制限給餌区〉選択系飽食給餌区>非選択系飽食給餌区の順となった。 各区の見かけのタンパク質および脂質の蓄積率を F i g .1 ・ 2に示した。見かけのタンパク質蓄積. -185一.
(16) 8号. 近大水研報. 6000. (2002). 2000. 60. a. aaTqd. nuwaua ︾. 勾&. ︾. 弓 4. 500. 10. R. o. od. 4E'. n g u e R. 程。. 竜 一 日. 口. SSS a t i a t i o n. •. gsg色 槌 畠 ¥ 芝 居 冨. a--45.. aaTqu. nununv n︾ nvn nununu. M、 @Emmm事 町 怠何回冨. 1000. 。. 50. nunu nvauv Rvhu. 草MW判。偉keeaMWCS¥お宮E. 5000. 盟関 Satiation. o. F i g .1 ・ 3 .C o m p a r i s o nofp e p s i n l i k ee n z 戸n ea c t i v i t i e so fg 出 回ct i s s u e6h o u r sa f t e rf e e d i n gb e t w e e n 鎚 l e c t e ds t r a i n s( S S )a n dn o n s e l e c t e ds t r a i n s (NS) o fr e ds e ab r 弱m s .E r r o rるa r si n d i c a 句 s 組n d a r dd e v i a t i o n . 似 l y福 島: r e n tヤ く0.05). •V a l u e sm a r k e dw i t ht h el e t t e ra r es i g n i f i c. 50. 50. 1 . 8. 1 . 6 40. . : d 40 T , I ). 1 . 4. g t 2. ・固唾. ω. 偲. ‘ -. 偉. t : ' l. 主30. ~30. ・田園. 弓 = T J J 、. a. ~. 邑 島. T J J. h. ・田圃. 0. 孟1.0. 0. 20. 冨. 冨0 . 8 、 、. g20. t : ' l. 苦0.6. ・ 司 、 、 F. 冨. z. 前. 10. 喝... g10. 0 . 4. &亀. a o c. &亀. Qd. r egu R. o o • SSS a t i a t i o n 口SSR e s t r i c t i o n 瞳 NSS a t i a t i o n. N. o. ロ. 0 . 2. F i g .1 4 .C o m p a r i s o no fp e p s i n l i k eenzymea c t i v i t i e so fg a s 佐i cd i g e s t a6h o u r sa f t e rf e e d i n gb e t w e e n s e l e c 包 ds 出 血s ( S S )a n dn o n s e l e c t e ds 泊 i n s( N S )o fr e ds e ab r e a m s .E r r o rb郡 i n d i c 絢 S飽n ゐr d. 率は選択系制限給餌区が 3 7.1%と最も高く,次いで、非選択系昔話限給餌区の 26.4%であった。飽食 給餌区では両系統ともに 23.5%であった。見かけの脂質蓄積率は非選択系制鼓給餌区>選択系制 限給餌区>選択系飽食給餌区>非選択系飽食給餌区の頬となり,非選択系飽食給餌区は著しく低. -186一.
(17) 家戸:マダイの品種改良に関する研究. 3 . 0. 5 0. 70. eaωCS¥SEZ 芸 Mgbpkm唱. 45. 冨. 32.9 0 』. ~. 2 1 5. 、. 4. ・・. U ︽ nunu AU R u a a y q J曹 内. 母制油田O槌叫唱槌¥組制何回同ヨ. 10. 組. 宮1.0 E. 0 . 5. 5. o. o. o SSS a t i a t i o n. •. 2 . 5. nuauavzuauRuav ,h 吟4 4 a-Tququn 4. 60. ロSSRestriction 瞳 NSSat. 訟をi o n. 口NSRe抑 制ion. F i g .1 5 .C o m p a r i s o no f住y 予s i n l i k ee n 勾 mea c t i v i t i e so fi n t e s t i n a ld i g e s t a6ho ぽ sa 会e rf e e d i n g 切 i n s( N S )o fr e ds e ab r e a m s .E r r o rb a r s b e t w e e ns e l e c t e ds t r a i n s( S S )a n dn o n s e l e c t e ds i n d i c a t es t a n d a r dd e v i a t i o n . 。 V a l u e smarkedw i t ht h esamel e 伐e ra r es i g n i f i c a n t l yd i f f e r e n t 伊< 0 . 0 5 ).. 2 . 4. 2 . 5. 1 . 0. 旨 :1 . 6. 38.8. S 。 £ ・田園. =. Qi. 喝-. a. 。 伊 ¥. ~1.5. 唱. Z12. ・田圃. 電 器 島. 、 -1.0 ~. Q. Q. ご0.8. ・声圃. 冨. 冨. = 0 h. e ~. 、 、. O•6. 的. . 4 苦0 Z. 磁. 唱団. = 0 . 4. 0 . 2. o. 9. ・圃圃. 0 . 5. 冨. o •. 1 . 2. 2 . 0. 恒. 2 . 0. a. SSS a t i a t i o n. 口SSRestricをion 盟 NSSatiation 口NSRest耐. 言i g .1 ・ 6 .C o m p a r i s o no fa m y l a s ea c t i v i t i e so fi n t e s t i n a ld i g e s t a6加盟'S a f t e rf e e d i n gb e t w e e ns e l e c t e d. s 回 i n s( S S )a n dn o n s e l e c t e ds 凶 I n s( N S )o fr e ds e ab r e a m s .E r r o rb a r si n d i c a t es 包n 白r d d e v i a t i o n . 。 V a l u e smarkedwi 也 t h esamel e t t e ra r es i g n i f i c a n t l yd i 首e r e n t 争< 0 . 0 5 ).. い値を示した。 各区の摂餌 6時間後における胃組織および内容物のペプシン様酵素活性を Fig. 1 3および F i g .. I ・4にそれぞれ示した。両系統において飽食給額亙の胃組織のペプシン様酵素活性は制限給餌区.
(18) 近大水研報. 8号. P r o t e i n. (2002). S u g e r. 70 nunununununu a u E u a a T q d曹 司 4 4 E. (ヌ )b ヨa宮 ωω宅 冨ω﹄豆島司. o. S S S a t i a t i o n. SS R e s t r i c t i o n. NS. NS. S a t i a t i o n. R e s t r i c t i o n. F i g . I ・ 7 .C o m p a r i s o no fa p p a r e n td i g e s t i b i l i t yo fp r o t e i na n ds u g a ri nt h ei n t e s t i n a ld i g e s t a6h o u r sa f t e rf e e d i n g b e t w e e ns e l e c t e ds 回 i n s( S S )a n dn o n s e l e c t e ds 回 i n s( N S )o fr e ds e ab r e a m s .. P r o t e i n. 9 5 . 4. nuEunU. A36000. w a a︿ (ヌ )bzZ音色宅相国ωm. 9 5. S u g e r. ﹄. 75. SS S a t i a t i o n. SS R e s t r i c t i o n. NS. NS. S a t i a t i o n. R e s t r i c t i o n. F i g .1 8 .Comparisonofa p p a r e n td i g e s t i b i l i t yo fp r o t e i na n ds u g a ri nt h ef e c e sb e t w e e ns e l e c t e d紺 ヨi n s( S S ) 位a i n s( N S )o fr e ds e ab r e a m s . a n dn o n s e l e c t e ds. -188一.
(19) 家戸:マダイの品種改良に関する研究. のそれよりも低かった凸胃内容物においても有意差はみなれなかったものの胃組織のペプシン様 酵素活性と同じような傾向がみられた。各区の摂餌 6時間後における腸内容物のトヲプシン様酵 素活性を F i g .1 5に示した. 9. 非選択系飽食給額区のトリプシン様酵素活性は著しく低かった。ま. た,同区のアミラーゼ、活性も低かった ( F i g .1 ・6 )。 各区の摂餌 6時間後の揚内容物および糞におけるタンパク質および糖震の見かけの消化率を. F i g .1 ・7および F i g .1 ・8にそれぞれ示した c 非選択系飽食給餌区において,腸内容物および糞と もに見かけのタンパク質および糖糞の消化率が他区よりも低かった c 本研究の結果から,飽食給餌した場合に,選択系の摂餌率,増重率,飼料効率,タンパク質効 率および消化率はし、ずれも非選択系よりも高いこと,および制限給餌した場合には選択系は飼料 脂質をエネルギー源として利用しタンパク質を蓄積する能力が高いことが示された。. B r e t ta n dG r o v e s82) は,幼魚期に飽食給餌した場合には体に蓄積されるエネルギー量が代謝で、 消費されるエネルギー量よりも多いこと,両エネルギー量は初期において比較的高く,サイズお よび年齢の増加に伴い減少するが,その減少の程度は両者間で、異なっており,やがて蓄積される エネルギーよりも代諒エネルギーの方が大きくなることをみておち,その根本的な要因がサイズ の増加ー飼料効率の減少という基本的な原因に基づくと報告している c 本研究における飼育試験 では,選択系と非選択系との聞にサイズの差異がみられた。従って,本研究の結果のみに基づい て選択系の特徴を議論することは難しい. E. しかしながら,飼料効率などはサイズの大きかった選. 択系が非選択系に比べて高く飼育成績が優れていたこと。この点から同サイズで比較した場合に おいても選択系の方が優れた飼育成績を示す可能性が高いと考えられる。. -189一.
(20) 近大水研報. 第I I章. 8号. (2002). 染色体操作による品種改良. 前章では,選択系マダイが非選択系マダイよりも摂餌量や飼料効率が高い結果,成長の褒れる ことを明らかにした. D. しかしながら,近年の不況,魚病の蔓延,さらには生産過剰による市場髄. 格の低迷などによち,マダイの養殖経営は厳しい状況にあち,より護れた特性を錆えた品種が養 魚家より要求されてきている c 本章では染色体操作を用いた新たな養殖用品種作出の可能性につ いて検討した。. E・1.第二極体放出題止型および第一卵嵩題止型雌性発生二倍体マダイの生残,或長および外部. 形態回 雌性発生二倍体は,遺伝的に不活性化した精子で通常卵を媒精し,その後第一卵誤を姐止(以 下 , m i t o t i c G 2 Nと略記)あるいは第二極体放出を担止〈以下, m e i o t i c G 2 Nと略記)して作出さ 5 ヘ雌性発生二倍体マダイの作出方法についてはこれまでに報告されている 37, 4I)が,雄性 れる 49,. p l e g n a t h u s 発生二倍体マダイの飼育に関する報告はまだない。そこで本研究では,イシダイ O f a s c i a t u s 精子の遺伝的不活性化に有効な紫外線照射条件について検討するとともに, m e i o t i c G2Nおよび m i t o t i c G 2 Nマダイを作出して,それらの生残,成長および外部形態を通常二信体 o n n a l ・ 2Nと略記〉と比較した 5 (以下, n. 1 1 1 -1.材料および方法 (1)親魚および採卵・採精. 6および 7世代目の選抜育種マダイ. 4 ). 5歳魚の雌一尾(尾叉長, 5 8. 4cm;体重, 4, 450g ) および. 070g ) よりそれぞれ搾出法により採卵および採精した。雑性 雄一尾(尾叉長, 56.0cm;体重, 4, 7 . 6 発生二倍体の作出には,雄性発生誘導の確認を容易にするためにイシダイ 7議魚(尾叉長, 3 cm,体重, 2, 260g ) の精子を用いた。なお,予備実験において,イシダイ精子とマダイ卵は受 精するが, F1仔魚は奇形となることを確認した c. ( 2 ) イシダイ清子の遺伝的不活性化のための最適繋外線照射量 1出 fのイシダイ精子を海産魚用 BSS( b a l a n c e ds a l ts o l u t i o n )で 5 0信に希釈後,小野里および山 2 5e r g/mm Isの紫外線 (UV) を黒射した。 羽却の方法に従い 6. uv照射量は 0,250,500, 1, 000,. 2 1 , 500,2, 000,3, 000,4, 000,5, 000,6, 000,8, 000および 1 6, 000e r gImm ( 0・2 6 3s ) とした。処. , 300粒のマダイ卵に媒精した。各照射量について約 1 5 0粒の卵を 141容 理した精液 2mlを約 1. 8Cでインキュベートし,解化率を求めた c ビーカー 2個ずつに収容して 1 0. ( 3 ) 臨eiotic-G2Nおよび mitotic-G2Nの作出 1尾の雌から得た卵を 3群に分け,そのうちの 2群は雌性発生二倍体作出のためにじV 処 理 2 ( 3, 000e r g/mm 8Cでインキュベートした o m e i o t i c G 2 N ) したイシダイ精子で媒精し,その後 1 0. -190一.
(21) 家戸:マダイの品種改良に関する研究. は媒精後 3分から 30分間 ltで抵温処理して作出した。 m i t o t i c G 2 Nは媒精後 46分から 5分 30 2 秒間フレンチプレス(大岳製作所,東京〉を用いて 700均 /cm の高水圧処理 41)を行って作出し. た。残りの 1群の卵には通常マダイ精子を媒精し,対照魚として通常二倍体 ( n o n n a l・2N) を作 出した。 meiotic-G2N,m i t o t i c G 2 Nおよび nonnal-2Nの作出に用いた卵数は,それぞれ約 26, 000, 39, 000および 1 3, 000粒で、あった。各群について約 1 5 0粒の卵を 11容ビーカー 2留ずつに i 収容し 0. て1 8Cでインキュベートした c 卵発生および群化を投影機で観察・計数し,怪体形成率 ( E S ), 解化率 (H 畏〉および正常鮮化率 (HRN) を以下の式により算出した。 王S (%)= 100xNe/Nt. HR(%)= 100x(N a'る+Nn)/Nt 0 0xNn/(Nab+Nn) HRN(%)= 1 Ne=怪体形成卵数・ Nt=総卵数:Nab=異常解化仔魚・ Nn. 正常解化仔魚. uv処理精子で媒精後処理をしなかったものも同様に観察し,無処理対照として用いた. c. ( 4 ) 仔魚および稚魚の飼育 nonnal ・2N 1 0, 000尾 , ,meiotic-G2N 6, 700尾および m i t o t i c G 2 N4, 500の仔魚を, 5001容水槽 ( n o n n a l 2 N ) と 2001容 水 槽 ( m e i o t i c G 2 Nと m i t o t i c G 2 N ) 2基ずつに収容して飼育した。各区 i g .I I l~こ示した。飼育水には, の飼育水温,餌飼料系列および換水率を F. uv殺菌処理した議過. 海水を用い, 50日齢までは水温が 1 8Cを下巨らないように石英ヒーターを水槽に投入した。飼 0. 育水の塩分は 2 2 . 8 3 3 . 6 (平均±標準偏差, 3 2 . 9土1.7 ) であったむ飼育水の換水率は魚の成長に 合わせて 0 2 0へと増加させた ( F i g .I I -I)。鮮化後 30日目までは飼青水 i こN a n n o c h l o r o p s i ss p . を添加し, 500, 000 c e l l s/mlとなるように維持した。水槽の上面には蛍光灯を設置し,午前 8a 寺 から午後 6時まで点灯した。醇化後 2-40日自までワムシ B r a c h i o n u s平.を, 18-40S目までブ ラインシュヲンプ A r t e m i as a l i n aを , 20日目以降は市販の配合飼料をそれぞれ給餌した。 n o n n a l 2N区においては,飼育密度調整のため解化後 29日吉および 62日自に半数の魚を水槽かち取り. 上げた。解化後 90SeIまでの各区の生残率を記録した。 ( 5 ) 第一卵割阻止裂雌性発生二倍体誘導の確認. m i t o t i c G 2 N誘導の確認はアイソザイムマーカ一分析によって行った。検出した 5酵素,遺イ云. 子座および電気泳動に用いた緩衝液を T a b l eI I lに示した。第 I章に示した方法で,各区 20尾 ずつの Aat ・2 , * Adh , *E s t矢 Pgm-l* および 6-Pgd*遺伝子座を検出した c. ( 6 ) 外部形態 外部形態の調定には各区とも 9 1 日齢の魚を用いた。魚体重,標準体長および体高を測定し, 背鰭,胸鰭,腹鰭,尻鰭および尾鰭の軟条数を計数した。測定結果から,平均,標準需差,分散 および変動係数を算出し,試験区間で比較した。形態異常を示した魚の出現率も同時に記録した。 ( 7 ) 鎮育詰験. 実 験 1:各区 30尾ずつを 2001容 水 槽 に 収 容 し 30 日間涜水式で飼育した。 m i t o t i c G 2 N区 ,. 一 一. 1 9 1一 一.
(22) 近大水研報. 8号. (2002). •. Formulatedd i e t. ・ 司 .. 2rinesMEP. p. • ‘ R o t i f e r. 25. 記 20 ~. 20~ミ. h. g. a. 場-. a. 唱. @. ‘ h、. h. 1 5 3. Q.. E 4 占. z t 5 h d 主. 一置-. N o r m a lー2 N. : r 。 r . I ). 一・一括e i o t i c G 2 純 一会-. ~. 10. M i t o t i c G 2 副. e. 唱 占. g. =. Z. 5 誌 唱 占. h. ~. a. o~. 10. o. 10. 20. 30. 50 60 ま e rh a t e h i n g D a y sa f i 40. 70. 90. 80. F i g .s ・ 1 .W a t e rt e m p e r a 知詑 o fe a c hg r o u p ,f e e d i n gr e g i m ea n dw a t e re x c h a n g er a 健sf o r詑 a r i n gr e ds e a b r e a m企'Om 0t o9 0d a y sa f t e rh a t c h 訟g .. Table1 1 ・1 .Theenzymesexamined,l o c id e t e c t e dandるu f f e rs y s t e m su s e df o r e l e c t r o p h o r e s i si nr e ds e abreams a m p l e s Enzyme Enzyme( a b b r e v i a t i o n ) commission Locus B u f f e r * number 1 . 1 . 1 .1 Adh* T-C A l c o h o ld e h y d r o g e n a s e(ADH) 予紅t a t ea m i n o t r a n s f e r a s e(AAT) 2 . 6 . 1 . 1 Aat ・2 * C-APM As . 1 .3 E s t * TBE E s t e r a s e(EST) 3 .1 Pgm-l* C-APM P h o s p h o g l u c o m u t a s e(PGM) 2 . 7 . 5 . 1 4 4 6-Pgd* CAPM 6 P h o s p h o g l u c o n a t ed e h y d r o g e n a s e(6-PGD) 1 . 1 . 1. 由. *T-C,Tris-citricacidpH8.0;C-APM,Citricacid-aminopropylmorpholinepH6.0;TBE,Trisb o r a t eEDTA予H 8 . 9 .. m e i o t i c G 2 N区 お よ び n o r m a l 2 N区の実験開始時の魚体重(平均±標準備差〉は,それぞれ 9 . 8 +2 . 9g ,1 0 . 9+ 0 . 2gおよび 7 . 6+ 0 . 2 gで、あった。急体重を 1 0日目毎に測定した。魚体重の演j 定結果に基づいて,魚体重の 3% の市販マダイ用配合飼料(丸紅飼料株式会社製,粗タンパク質,. -192-.
(23) 家戸:マダイの品種改良に関する研究. 90. 8 0 70 S80 司J. ~ h. 50. OD. 去 40 ω 遣30 20 1 0. 。 。. ぜト」. 1 0 0 02 0 0 0 3000 4000 5 0 0 06 0 0 07 0 0 08 0 0 0 2. UVdose (erg/mm). o. さトーJ 2 0. 40. 6 0. 8 0. 1 0 0. DurationofUV-irradiation( s e c ) F i g .1 1 ・ 2 .E f f e c t so fUVd o s eu s e dt oi r r a d i a t eJ 勾a n e s ep a r r o tf i s hs p e n nont h eh a t c h i n gr a t e ( m e a n% , n = 2 )o fr e ds e abreame g g s .. 5 6 . 1% ;4 0 0 . 6k c a l l1 0 0g ) を給餌したc 各区の日間摂餌率,増重率, 日開成長率および飼料効率 をそれぞれ算出した。 実験 2:解化後 2 10日目に, m i t o t i c G 2 N区倒産, m e i o t i c G 2 N区 1 0 4尾および n o n n a l 2 Ng. 9 9尾を近畿大学水産研究所白浜実験場地先の海面縞生賓 (3X3X3m) にそれぞれ収容し,約 2 年間餌青した q 収容時の m i t o t i c G 2 N区 , m e i o t i c G 2 N区および n o n n a l 2 N区の魚体重(平均±標 準偏差)は,それぞれ 1 1 6. 4+ 2 3 . 8g,1 2 8 . 2+ 2 3 . 9gおよび 1 2 7 . 9+ 1 9. 4gであった。 1ヶ月あ るいは 2ヶ月呂毎に魚体重を測定するとともに生残率も記録した。. ( 8 ) 統計解析 試験区間の分散を F i s h e r ' sFで,平均値の差を S c h e f f sFでそれぞれ有意差検定した(p< 0 . 0 針 。 有意差検定はコンピュータプログラム S t a t V i e w 5 . 0 (SASI n s t i t u t eI n c .USA ,C a r y ) によった。 1 1 1・2 .結 果. (1)イシダイ精子の遺伝的不活性化のための最適紫外線照射量 マダイ卵の解化率に及ぼすイシダイ精子へのじV 照射量の影響を F i g .I I ・ 2に示した c 未照射 の精子で媒精した卵の鮮化率は 8 4.5%であった D 照射量が 250erg/mm の時に癖化率は著しく 2.
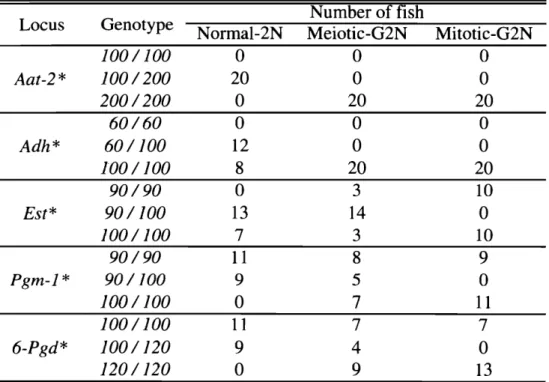
(24) 近大水研報. 8号. (2002). Table1 1 ・ 2 .S u r v i v a lr a t e sa tembryonics t a g e(ES), t o t a lhatchingr a t e s(HR) andhatchingr a t e sofnormallarvae(HRN)i nnormal-2N, m e i o t i c G2Nandmitotic-G2Ngroupsofreds e aむream(mean% , n=2) Totalnumberof ES*l HR*2 Ploidy HRN*3 eggsexamined Haploid 3 0 2 7 6 . 4 9 7 2 . 1 9 0 . 0 0 Normal-2N 9 4 . 8 6 3 0 3 8 5 . 1 5 8 3 . 5 0 Meiotic-G2N 310 4 5 . 7 6 5 0 . 9 7 3 8 . 0 6 Mitotic-G2N 312 3 5 . 0 0 5 0 . 3 2 1 2 . 8 2 * 1N umbero fd e v e l o p i n ge m b r y o sIT o t a lnumbero fe g g se x a m i n e dX1 0 0 . *2( Numbero fn o r m a l l a r v a e+Numbero fa b n o r m a l l a r v a e )IT o t a lnumbero fe g g s. e x a m i n e dX1 0 0 . *3N umbero fn o r m a l l a r v a eI(Num ちe ro fn o r m a l l a r v a e+Numbero fa b n o r m a l l a r v a e ) X1 0 0 .. 100. 一 ー -Normal-2N 一 ・ -Meiotic-G2N 一 会 -Miま.otic-G2N. ¥ ¥ , 話 ) ・ ' 、 ‘ 、. 60 ト. ¥. 。 場 需 』 d. U ・ 寄 2 ' = 2. 判. 2 0 G. o. 10. 20. 30. 40. 50. 60. 70. 80. 90. Daysaを 1 Ie l ・hatching F i g .1 1 ・ 3 .Th es u r v i v a lr a t e s( m e a n% , n = 2 )o fn o n n a l 2 N,m e i o t i c -a n dm i t o t i c G 2 N担 r e ds e a. b 間 出n合o m0 t o90 d a y sa 会e rh a t c h i n g .. 2 2 低くなり(14.0%),500e r gIm m で再び上昇し始め ,2, 000e r gIm m 7 4.3%)となった C で最大 (. 2 3, 000から 8 , 000e r gIm m までは 55%前後で推移した s 従って,これ以降の実験では u v照射量 2 を 3, 000e r g/m m ~こ定めた。. ( 2 ) 第二護体放出題止歪および第一那覇題止型雌性発生二信体の生残および醇化率 norm a 1 -2N区 , m e i o t i c G 2 N区および m i t o t i c G 2 N区の経体形成率,解化率および正常癖化率を T a b l eI I ・ 2に示した。じV 処理したイシダイ精子で媒精後いずれの処理もしなかった無処理対照. -194-.
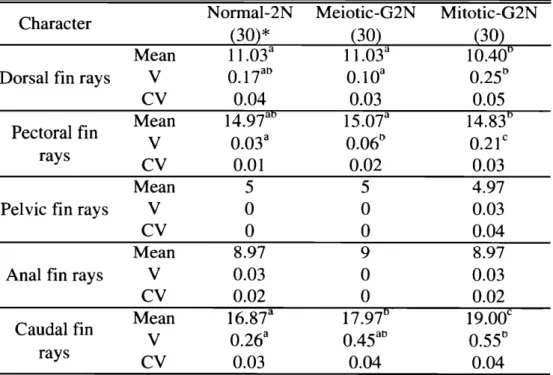
(25) 家戸:マダイの品種改良に関する研究. Table1 1・3 .GenotypesoftheAat・2てAdh*,E s t * ,Pgm-1*and6 -Pgd* l o c ii nn o r r n a l2N,m e i o t i c -andmitotic-G2Ngroupsofr e d s e abream Numberoff i s h Genotype Locus Norrnal-2N Meiotic-G2N Mitotic-G2N 四. A a t 2 *. Adh*. E s t *. Pgm-1*. 6-Pgd*. 100/100 100/200 200/200 60/60 60/100 100/100 90/90 90/100 100/100 90/90 90/100 100/100 100/100 100/120 120/120. O 20. O O 12 8. O 1 3. 7 1 1 9. O 1 1. 9 O. 。 。 。. 。 。 O. 20. 20. O. O. 20 3 14 3 8 5. 20 10. 7 7. 1 1. O 10. 。 9. 4. 7 O. 9. 1 3. 区の匪体形成率および解化率はいずれも 70私以上の檀を示したが正常仔魚は認められなかった。. n o r m a l ・2 N区がすべての指標で最も高い値を示した。 m i t o t i c G 2 N区の鮮化率および正常解化率は 2 . 8および 35.0%であった o n o r m a l 2 N区 , m e i o t i c ・G2N区および m i t o t i c 最も低く,それぞれ 1 G2N区の生残率は. それぞれ解化後 20日匡に 4 5 . 0 .6 . 3および 1 1 .3%,5 0S自は 3 2 . 8 .5 . 8およ. び 9.6%,90自自では 3 2 . 6,5.3および 8.6%で、あった ( F i g .I I ・3 )。 ( 3 ) 第一卵割阻止型雌性発生二倍体誘導の確認. n o r m a l ・2N区 ,. meiotic・G2N 区および mitotic-G2N 区の Aat-2 矢 Adh キ~. E s t * ,P g m l*および 6 P g d *. 遺伝子座の遺伝子型を T a b l e1 1 3に示した。 A a t 2 *および Adh*遺伝子座においては meiotic-G2N および m i t o t i c G 2 Nともに全ての個体がホモ接合型で、あった。 E s t * ,P g m l *および 6 P g d *遺 伝 子 座において, m e i o t i c G 2 N区 20尾中それぞれ 1 4, 5および 4患がヘテロ接合型であったが,. m i t o t i c G 2 N区においては全個体がホモ接合型であった。 n o r m a l ・2 N区 , m e i o t i c G 2 N区および m i t o t i c G 2 N区の E s t *の電気泳動像を F i g .1 1 -4に示した。. ( 4 ) 外部形態 n o r m a l ・2N区 , m e i o t i c G 2 N区および m i t o t i c ・G2N区の魚体重,標準体長および体高の平均,分 a b l eI I ・ 4に示した。 m e i o t i c G 2 N区および m i t o t i c G 2 N区の分散は,いずれ 散および変動係数を T の形質においても n o r m a l 2 N区よりも有意に高く, m i t o t i c G 2 N区は変動係数も最も高かった。 各区の計数的形質の平均,分散および変動係数を T a b l e1 1 ・5に示した。 m i t o t i c G 2 N区の胸鰭軟 条数および尾鰭軟条数の分散は n o r m a l 2 N区よりも有意に高かった白. n o r m a l ・2N民 m e i o t i c G 2 N区および m i t o t i c G 2 N区の形態異常魚の出現率を T a b l e1 1 ・6~こ示し.
(26) 近大水研報. 8号. (2002). +. o F i g .11-4. E l e c t r o p h e r o g r a m so fEst寧 ( e s t e r a s e )f o rt h en o r m a l 2N( A ),m e i o t i c G 2 N( ヨ )a n dm i t o t i c -G2N( C )g r o u p s o fr e ds e ab r e a m .. bFEhu. ・. FBPE. Gι 一aoz 一aO. i---1j. l u 一&日. JS. ︹ リ 一N v リゆ一ぴ d15-NU66 rτ3Jq ニ29j 511 一. ob. 6 z -. -S孟. -a. O. AU. e W C v d 、也. B. Table1 1 4 .M o r p h o m e t r i c so fo f f s p r i n g( 9 1d a y so l d ) ;Mean,v a r i a n c e , ( V )andc o e f f i c i e n to fv a r i a t i o n(CV)o fbodyw e i g h t s t a n d a r dl e n g t handるodyd e p t hi nnormal-2N, meiotic-G2N andm i t o t i c G 2 Ng r o u p so fr e ds e abream Normal-2N- Meiotic-G2N-M i t o t i c G 2 N C h a r a c t e r ( 1 5 1 ) * ( 1 7 5 ) * a 0 7 . 5 2 1 0 . 2 0 Mean a 0 v 2 . 7 8 1 0 . 0 2 2 2 0 . 3 1 CV 0. a 孔1 e a n 6. 35 6 . 9 10 S t a n d a r dl e n g t h a V 0 . 2 4 0 . 8 70 ( c m ) CV 0. 0 8 0 . 1 4 a 0 Mean 2. 33 2 . 6 2 ヨodyde予t h b a V 0 . 0 5 0 . 1 0 ( c m ) cv 0 . 0 9 0 . 1 2. *Numberoffish.. a b cM eansw i t h i nal i n en o tmarkedbyt h esames u p e r s c r i p tl e t t e ra r es i g n i f i c a n t l y. d i f f e r e n t( p< 0 . 0 5 ) .. た 。 meiotic-G2N区および mitotic-G2N区の短躯症,脊柱側室事症および頭部形態異常の出現率は normal-2N区よりも高かった。また,すべての種類の形態異常魚の出現率は mitotic-G2N区が最. も高かった。. -196-.
(27) 家戸:マダイの品種改良に関する研究. Table1 1 ・ 5 .Morphometricso fo f f s 予r i n g( 9 1d a y so l d ) ;Mean,v a r i a n c e ( V )andc o e f f i c i e n tofv a r i a t i o n(CV)o fm e r i s t i cc o u n t si n r o u p so fr e ds e a normal-2N,meiotic-G2Nandmitotic-G2Ng bream Normal2N Meiotic-G2N Mitotic-G2N C h a r a c t e r ( 3 0 ) ( 3 0 ) 。 Q r : a a 1 0 . 4 0 0 1 1 . 0 3 Mean 1 1 . 0 3 ao a 0 V 0 . 1 7 0 . 1 0 0 . 2 5 D o r s a lf i nr a y s CV 0 . 0 4 0 . 0 3 0 . 0 5 ao a 1 4 . 8 3 0 民1 e a n 1 4 . 9 7 1 5 . 0 7 P e c t o r a lf i n a 0 C V 0 . 0 3 0 . 0 6 0 . 2 1 r a y s CV 0 . 0 1 0 . 0 2 0 . 0 3 Mean 5 5 4 . 9 7 V O 0 . 0 3 P e l v i cf i nr a y s CV 0 . 0 4 Mean 8 . 9 7 9 8 . 9 7 V 0 . 0 3 0 . 0 3 Analf i nr a y s CV 0 . 0 2 0 . 0 2 a C 1 7 . 9 7 6 Mean 1 6 . 8 7 1 9 . 0 0 Caudalf i n a ao 0 V 0 . 2 6 0 . 4 5 0 . 5 5 r a y s CV 0 . 0 3 0 . 0 4 0 . 0 4 *Numbero ff i s h . a b cM e a n sw i t h i nal i n en o tm a r k e db yt h es a m es u 予e r s c r i p tl e t t e ra r es i g n i f i c a n t l y d i f f e r e n t(pく0 . 0 5 ) . 置. 。 。. 。 。 。. .Thei n c i d e n c e so fd e f o r m i t i e si nnormal-2N,meiotic-G2N Table1 1・6 andmitotic-G2Ng r o u p sofr e ds e abream(%) Number S h o r t 0 _ _ 1 ! _ _ ! _ Deformity La c 1 ιof S c o l i o s i s c c i p u t operculum off i s h s t a t u r e ~-~--~--- ofo Normal-2N 513 0 . 1 9 0 0 . 9 7 0 Meiotic-G2N 328 3 . 6 6 5 . 1 8 4 . 5 7 0 Mitotic-G2N 202 7 . 9 2 7. 43 7. 4 3 1 .98. ( 5 )飼 育 誌 験 実験 1:・各匿の餌育試験の結果を Tぬ l e1 1 7にまとめて示した。増重率,. 日開成長率および飼. 料効率はいずれも m i t o t i c G 2 N区で最も低く, n o n n a l 2 N区で、最も高かった。 実験 2:n o n n a l 2 N区 , m e i o t i c G 2 N区および m i t o t i c G 2 N区の魚体重の変動を T a b l e1 1 8に示 した。鮮化後約 1年経過した 3 6 8日齢における m i t o t i c G 2 N区の魚体重〈平均±標準偏差, 1 9 4 . 9. : t4 0. 3g )は , n o n n a l 2 Nr K( 2 3 3. 4+4 3. 3g ) および m e i o t i c G 2 N区 ( 2 4 5 . 8: t3 7 . 0g ) よりも有 意に低かった。解化後 9 2 2日自には, n o n n a l ・2 N区の魚体重が 1 7 2 2 . 6+ 2 7 8 . 2gで最も高く, m e i o t i c G 2 NrKが 1 5 3 7 . 9土 2 1 0 . 2gでこれに続き, m i t o t i c G 2 N区は 9 5 3 . 2+ 2 3 9 . 8gと最も低かっ o r τ n a l 2 N亙 , m e i o t i c G 2 N区および m i t o t i c G 2 N区の額育期間中の生残率は,それぞれ 4 0, 4 . た 。 n 6 0 . 6および 1 7 . 9 %で、あった。. 一1 9 7一.
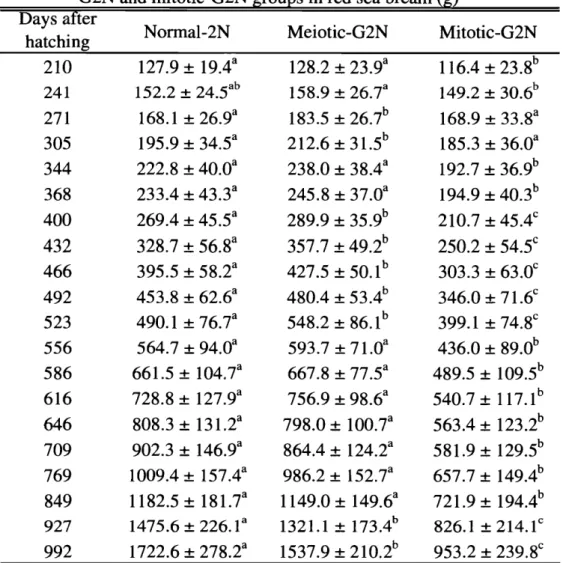
(28) 近大水研報. 8号. (2002). TableI I ・ 7 .R e s u l t so ff e e d i n gt r i a lo fnormal-2N,meiotic-G2Nandm i t o t i c G2Ng r o u p si nr e ds e abream Normal-2N Meiotic-G2N Mitotic-G2N Av e r a g ebodyw e i g h t( g ) f :0 . 2 * 1 0 . 9: f :0 . 2 9 . 8: f :0 . 1 7 . 6: I n i t i a l f :0 . 7 2 9 . 2: f :1 .1 2 4 . 8: f :0 . 4 F i n a l 2 2 . 2: b a a b D a i l yf e e d i n gr a t e(%) 2 . 5 0: f :0 . 0 5 2 . 5 6: f :0 . 0 3 2 . 5 9: f :0 . 0 1 c a b 1 6 7 . 5: ! :5 . 9 1 5 3 . 9: 1 9 2 . 5: f :3 . 4 f :4 . 9 Weghtg a i n(%) b a 3.15±0.06b D a i l ygrowthr a t e(%) 3 . 3 6: f :0 . 0 7 2 . 9 8: f :0 . 0 8 b b a 1 1 5 . 0: 4: f :4 . 7 134. f :3 . 5 f :3 . 8 Feede f f i c i e n c y(%) 1 2 3 . 3: n = 3 ) . tSD( *Mean: a b cM r es i g n i f i c a n t l yd yt u p e r s c r i e t t e ra i f f e r e n t i t h i nal i n en o tm a r k e db h es a m es e a n sw 戸l O . 0 5 ) . ( p<. fa v e r a g ebodyw Table1 1 8 .Comparisono e i g h to fnormal2N m e i o t i c , G2NandmitotIC-G2Ng r o u p si nr e ds e aるream包} Daysafter Normal-2N Meiotic-G2N Mitotic-G2N h a t c h i n g a a b f :1 9 . 4 1 2 8 . 2: 210 1 2 7 . 9: f :2 3 . 9 1 1 6 . 4: f :2 3 . 8 a a b b t2 4 . 5 241 1 5 2 . 2: t2 6 . 7 1 4 9 . 2: t3 1 5 8 . 9: 0 . 6 a b a f :2 6 . 9 1 8 3 . 5: f :2 271 1 6 8 . 1: 6 . 7 1 6 8 . 9: t3 3 . 8 a b a t3 4 . 5 2 1 2 . 6: f :31 1 9 5 . 9: .5 t3 1 8 5 . 3: 305 6 . 0 a a b 344 2 2 2 . 8: t4 0 . 0 2 3 8 . 0: f :3 1 9 2 . 7: 8. 4 f :3 6 . 9 a a b 2 3 3 . 4: 368 f :4 3. f :3 2 4 5 . 8: 3 7 . 0 1 9 4 . 9: f :4 0 . 3 a b c 400 2 6 9 . 4: f :4 5 . 5 2 8 9 . 9: f :3 5 . 9 2 1 0 . 7: f :4 5 . 4 a b c 432 3 2 8 . 7: f :5 6 . 8 3 5 7 . 7: f :4 9 . 2 2 5 0 . 2: f :5 4 . 5 a b c 466 3 9 5 . 5: f :5 8 . 2 4 2 7 . 5: f :5 0 . 1 3 0 3 . 3: f :6 3 . 0 a b c 492 4 5 3 . 8: f :6 2 . 6 4 8 0 . 4: f :5 3 . 4 f :71 .6 3 4 6 . 0: a b c f :7 4 9 0 . 1: 523 6 .7 5 4 8 . 2: f :8 6 . 1 3 9 9 . 1: f :7 4 . 8 a a b 556 5 6 4 . 7: f :9 4 . 0 5 9 3 . 7: f :71 .0 f :8 4 3 6 . 0: 9 . 0 a a b .5: f :1 586 661 0 4 .7 6 6 7 . 8: f :7 7 . 5 4 8 9 . 5: f :1 0 9 . 5 a a b f :1 2 7 . 9 616 7 2 8 . 8: 7 5 6 . 9: f :9 8 . 6 5 4 0 . 7: f :1 1 7 . 1 a a b f :1 31 8 0 8 . 3: 7 9 8 . 0: 646 .2 f :1 0 0 . 7 5 6 3 . 4: f :1 2 3 . 2 a a b 709 9 0 2 . 3: f :1 4 6 . 9 8 6 4 . 4: f :1 2 4 . 2 581 .9: f :1 2 9 . 5 a a b f :1 5 7 . 4 9 8 6 . 2: f :1 1 0 0 9 . 4: 5 2 . 7 6 5 7 . 7 : ! :1 4 9 . 4 769 a a b 1 1 8 2 . 5: 849 f :1 81 .7 1 1 4 9 . 0: f :1 4 9 . 6 721 .9: f :1 9 4 . 4 a b 927 1 4 7 5 . 6: t2 2 6 . 1 .1: t1 7 3 . 4 8 2 6 . 1: t2 1 4 . 1c 1321 a b c 992 1 7 2 2 . 6: f :2 7 8 . 2 1 5 3 7 . 9: f :2 1 0 . 2 f :2 9 5 3 . 2: 3 9 . 8 圃. *Mean~SD.. a b cM yt i t h i nal o tm u p e r s c r i p tl e a n sw i n en a r k e db h es a m es e t t e ra i g n i f i c a nt r es 1y d i f f e r e n t(p<0.05).. 一 198一.
図


+7



Outline
関連したドキュメント
これらの協働型のモビリティサービスの事例に関して は大井 1)
そのため本研究では,数理的解析手法の一つである サポートベクタマシン 2) (Support Vector
よって、製品の器種における画一的な生産が行われ る過程は次のようにまとめられる。7
介護問題研究は、介護者の負担軽減を目的とし、負担 に影響する要因やストレスを追究するが、普遍的結論を
機械物理研究室では,光などの自然現象を 活用した高速・知的情報処理の創成を目指 した研究に取り組んでいます。応用物理学 会の「光
今日のお話の本題, 「マウスの遺伝子を操作する」です。まず,外から遺伝子を入れると
この論文の構成は次のようになっている。第2章では銅酸化物超伝導体に対する今までの研
氏は,まずこの研究をするに至った動機を「綴
