博士学位論文
Tabebuia avellanedae
由来天然物およびその類縁
化合物からの JAK/STAT 経路阻害剤の探索と評価
近畿大学大学院
農学研究科 応用生命化学専攻
田 原 照 之
1
目次
目次 ... 1 第1章 序論 ... 3 第1節 Tabebuia avellanedae について ... 3 第2節 Tabebuia avellanedae 由来の天然物の臨床試験および薬理作用 ... 8 第3節 Tabebuia avellanedae に含まれる抗腫瘍活性天然物 NQ801 について ... 10 第4節 JAK/STAT 経路について ... 12 第5節 臨床応用されている JAK/STAT 経路阻害剤 ... 14 第6節 STAT3 の阻害剤結合部位 ... 17 第1項 SH2 ドメインに結合する化合物 ... 18 第2項 DNA バインディングドメインに結合する化合物 ... 21 第3項 クルクミンバインディングサイトに結合する化合物 ... 23 第4項 hinge 領域に結合する化合物 ... 24 第5項 STAT3 阻害剤研究の今後の可能性 ... 29 第7節 BBI608 の生物活性 ... 30 第2章 本論 ... 32 第1節 研究の目的 ... 32 第2節 ウェスタンブロットによる評価 ... 33 第3節 WST8 assay によるがん細胞に対する増殖抑制評価 ... 39 第4節 キナーゼアッセイ ... 44 第5節 STAT3 および STAT1 の SH2 ドメインに対するバインディングアッセイ 46 第6節 Autodock による STAT3 中の結合部位の推定 ... 51 第1項 NQ801 の結果 ... 52 第2項 BBI608 の結果 ... 54 第3項 化合物 25 の結果 ... 552 第4項 化合物 26 の結果 ... 56 第5項 【参考】4 化合物の重ね合わせ ... 57 第7節 レポータージーンアッセイによる NQ801 および BBI608 の評価 ... 58 第1項 レポータージーンアッセイの概略 ... 58 第2項 IL-6 濃度の最適化 ... 60 第3項 化合物の濃度設定 ... 60 第4項 ポジティブコントロールの選定 ... 62 第5項 レポータージーンアッセイの結果 ... 62
第8節 Reportergene assay による JAK/STAT 阻害剤の探索 ... 64
第1項 ヒット化合物の同定 ... 64 第2項 レポータージーンアッセイによるスクリーニング結果 ... 68 第9節 総括 ... 69 第10節 実験項 ... 72 第1項 細胞培養 ... 72 第2項 Western Blot ... 72 第3項 WST8 assay の手順 ... 74 第4項 Graphpadprism の設定について ... 74 第5項 キナーゼアッセイの手順 ... 83 第6項 Binding assay ... 85 第7項 レポータージーンアッセイ ... 86 第11節 Abbreviations ... 89 第3章 参考文献 ... 90 謝辞 ... 2
3
第1章 序論
第1節 Tabebuia avellanedae について
Tabebuia avellanedae はシソ目ノウゼンカズラ科の巨木で、冬季に白紫の花を
咲かせる。アマゾン川流域を中心とし、アルゼンチン、ブラジル、ペルー、パラ グアイ、エクアドル、ボリビアなどに自生している(Figure 1)。Pau d’arco, Red Lapacho, Ipe などの別名を持ち、その樹皮は、伝承薬として、バクテリアおよび
真菌による感染症、梅毒、マラリア、発熱、胃腸障害などに用いられてきた。1)
Figure 1. Tabebuia avellanedae
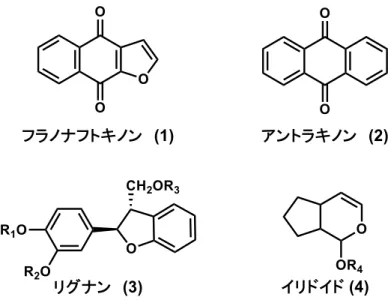
Tabebuia avellanedae はフラノナフトキノン、アントラキノン、リグナン、イリ ドイドなどの多様な天然物を含有し、今日までに様々な天然物が単離・構造決定
されている。1) Tabebuia avellanedae の抽出液は古来から伝承薬として利用され
ていた経緯があることから、その生理活性については多くの先行研究が報告さ れている。
4
Figure 2. Tabebuia avellanedae に含まれる天然物の基本骨格
抽出エキスの臨床応用の古い文献報告として、1873 年に内科医である Pinto は
Tabebuia avellanedae 抽出エキスが発熱、消化器潰瘍、性感染症、関節リウマチに
対して有効であると述べている。2)
Tabebuia avellanedae の in vivo での抗腫瘍活性効果については、ブラジルの
Queiroz らが報告している。3) エーリッヒ腹水がん移植モデルマウスモデルにお いて、Tabebuia avellanedae 抽出エキスの強制経口投与はがんの成長を抑制した。 また筆者らは同じマウスモデルで、Tabebuia avellanedae に含まれる天然物、β-lapachone の 1~5 mg/kg の静脈注射により同様ながんの成長抑制を報告している。 興味深いことに Tabebuia avellanedae 抽出エキスと、がんの成長抑制作用の用量 依存性は見られず、120 mg/kg 投与で最もがんを抑制し、500 mg/kg では顕著な 抑制作用を示さなかった。
5 Figure 3. β-lapachone の構造 Tabebuia avellanedae 抽出エキスのヒトを対象とした臨床研究もわずかながら 報告されている。4) 50 代男性のステージ 1b 期の食道がん患者に対し、Tabebuia avellanedae 抽出エキスを 12 g/day で経口摂取したところ、食道がんは、摂取 19 日目に病巣の一部が退縮していることが確認された。さらに摂取 84 日目には食 道がんの 4 か所の病変部位のすべてで退縮が確認された。当該患者に対して手 術は行われず、その後の 5 年間の経過観察中でのがんの再発は認められなかっ た。
Kangwon National University の Byeon らは Tabebuia avellanedae 内皮の水抽出 エキスが in vitro および in vivo で抗炎症作用を示していることを 2008 年に報告
している。5 ) Tabebuia avellanedae 内皮の水抽出エキスは LPS 刺激による
RAW264.7 細胞(マウスマクロファージ由来)の PGE2産生を用量依存的に抑制
した。また mRNA レベルでも COX2 の低下が確認されたが、TNF-α の mRNA レ ベルには影響を与えなかった。また、Tabebuia avellanedae 内皮の水抽出エキス は LPS 刺激による RAW264.7 からの NO(一酸化窒素)の産生を用量依存的に 抑制した。LPS 刺激による ERK のリン酸化レベルを Tabebuia avellanedae 内皮の 水抽出エキスは用量依存的に抑制したが、p38、IκBα、AKT のリン酸化には影響 を与えなかった。このことから Byeon らは ERK が標的タンパク質となりうる可
6
能性を指摘している。筆者らはさらに in vivo での耳介浮腫による評価を行って いる。興味深いことに Tabebuia avellanedae 内皮の水抽出エキスはアラキドン酸 誘発の耳介浮腫を有意に抑制したが、クロトン油で誘発された耳介浮腫に対し ては抑制作用を示さなかった。
Santa Catarina 大学の Freitas らは Tabebuia avellanedae の抽出エキスの経口投与
により、マウスの代表的なうつ病モデルで効果を示したと報告している。6) 強
制水泳試験(forced swimming test, FST)および尾懸垂試験法(tail suspension test, TST)において、Tabebuia avellanedae の抽出エキスの経口投与は用量依存的に無 同化時間を短縮させ、抗うつ作用の可能性が示された。
Sejong 大学の Park らは Tabebuia avellanedae の水抽出エキスが DSS (dextran
sulfate sodium)誘発大腸炎モデルに対して有効であったと報告している。7) B6 マウスに対して Tabebuia avellanedae の水抽出エキスを5日間経口投与し、DSS により大腸炎を誘発させたところ、プラセボ投与群と比較して、腸腺窩の損失、 腸粘膜での浮腫、免疫担当細胞の遊走などの大腸炎の兆候を軽減させた。また、 腸間膜のリンパ節において、Tabebuia avellanedae の水抽出エキスは、マクロフ ァージからの炎症性サイトカイン、IL-12 および IL-6 産生を抑制する一方で、抑 制性サイトカインである IL-10 産生を亢進した。 近畿大学の岩本らは、Tabebuia avellanedae の抗肥満作用について報告してい
る。8) 卵巣を摘出した in vivo マウスモデルにおいて、Tabebuia avellanedae のブ
タノール抽出エキスの経口投与はコントロール群と比較して、体重増加を有意 に抑制した。一方で、コントロール群と比較して、Tabebuia avellanedae ブタノー ル抽出エキス投与群の病理所見(体重、心臓、脾臓、肝臓、卵管)に顕著な違い は認めなかった。さらに岩本らは in vitro の実験を合わせて行っており、Tabebuia
7 株)のトリグリセリドを低下させた。さらには 3T3-L1 細胞を用いたアッセイを 抽出画分に適応し、トリグリセリドを低下させる活性本体の単離を試み、1-dehydroxy-3,4-dihydroaucubigenin(6)を見出した(Figure 4)。 Figure 4. 1-dehydroxy-3,4-dihydroaucubigenin の構造 本節では、Tabebuia avellanedae の抽出エキスの生理活性の先行研究について 述べ、in vitro もしくは in vivo での抗腫瘍活性、抗うつ作用、抗炎症作用につい て述べた。しかしながら、抽出エキスに含まれている成分のうちの、どの化合物 が主たる薬効を担っているかについては報告が乏しく、さらなる研究の進展が 求められている。
8
第2節 Tabebuia avellanedae 由来の天然物の臨床試験および薬理作用
Tabebuia avellanedae 由来の天然物のうち、臨床試験に進んだ化合物について は Lapachol および β-lapachone の例がある。Lapachol は Tabebuia avellanedae 由 来の天然物であるが、インドに自生する Strereosermum suaeloens (Bignoniaceae)か らも単離されている(Figure 5)。アメリカ国立がん研究所(National Cancer Institute, NCI)主導でその抗腫瘍効果の検証および Phase I の臨床試験が実施された。経口 で 4000 mg/kg/day もの投与が実施されたが、治療効果は確認されず 1970 年に臨 床試験は終了した。9) Figure 5. Lapachol の構造 一方、β-lapachone は ARQ 761 の名称で進行固形がん患者を対象として 2012 年から 2017 年に臨床試験が実施された。10) 臨床試験の結果が開示されていな いため詳細は不明であるが、さらなる進展は報告されていない。 β-lapachone は臨床試験に移行している一方で様々な薬効および生理活性が報 告されている。ハーバード大学の Li らは、β-lapachone のトポイソメラーゼ 1 阻 害活性について報告している。11) 既知のトポイソメラーゼ阻害剤であるカンプ トテシンはトポイソメラーゼ 1 と DNA との複合体を安定化させることにより、 DNA の切断を阻害するが、β-lapachone はその安定化作用は確認されなかった。 一方でトポイソメラーゼの酵素活性は、β-lapachone とのプレインキュベーショ
9
ンにより増強されることから、β-lapachone はトポイソメラーゼ 1 と直接的に作
用しその阻害活性を発揮していると考えられる。
コロラド大学の Pink らは β-lapachone のアポトーシス誘導能と quinone
oxidoreductase (NQO1)の関連について指摘している。12) NQO1 は多くの臓器や
細胞に普遍的に存在する酵素であるが、とりわけがん細胞で多く発現している ことが知られている。β-lapachone は NQO1 の基質となりカテコール体へと還元
されるが、容易に酸化され再びβ-lapachone へと変換されるため、NQO1 が触媒
する還元反応が進行し、結果として補酵素である NAD(P)H が枯渇し、そのアポ トーシス作用が発揮されるというメカニズムが提唱されている。
近年、韓国の Lim らにより、β-lapachone およびその類縁体の Mucosa-associated lymphoid tissue lymphomatranslocation protein 1 (Malt1)阻害活性が報告されている。
13) Malt1 は B 細胞および T 細胞の NF-κB のシグナル伝達に重要なタンパク質
であり、その機能阻害は B 細胞や T 細胞由来のがんや各種免疫疾患の治療につ
ながると期待されている。β-lapachone は Malt1 のパラカスパーゼ活性を抑制し、
その IC50値は 1.9 µM であった。同様のアッセイで α-lapachone および lapachol に
ついても評価が実施されているが阻害活性は認められなかった。ドッキングシ ュミレーションおよび wash-out experiment から β-lapachone は Malt1 のシステイ ン残基 Cys464 との共有結合形成が示唆された。
10 第3節 Tabebuia avellanedae に含まれる抗腫瘍活性天然物 NQ801 について 生命資源化学研究室では、Tabebuia avellanedae に含まれる抗腫瘍活性天然物 NQ801 (Figure 6)に着目し、その合成法の開発ならびに生理活性評価を行ってき た。14 Figure 6. NQ801 の構造 NQ801 の興味深い生理活性として、がん細胞に対する選択的な細胞毒性が挙 げられる。PC-3(前立腺がん由来)、A549(肺がん由来)、MCF7(乳がん由来) に対しては IC50値が 1 µM 以下とマイトマイシンと同程度の細胞毒性を示した (Table 1)。14a) Table 1. NQ801 の細胞毒性評価 IC50 (M)
human tumor cell lines
PC-3 A-549 MCF7
NQ801 0.14 0.78 0.51
11 一方で各種ヒト正常細胞*にたいしては、マイトマイシンは 1 µM 程度の細胞 毒性を示したのに対し、NQ801 はその IC50値が 3.11 ∼ 54.5 M と弱い毒性のみ が観察された(Table 2)。14a) Table 2. NQ801 の正常細胞に対する毒性 IC50 (M)
human normal cell lines
Fb Hc MPC-5 IE Hs888Lu SVCT-M12
NQ801 11.1 11.1 29.7 54.5 5.51 3.36
マイトマイシン 0.93 1.46 2.1 1.46 0.56 0.96
* Fb: normal skin, Hc: normal liver, MPC-5: normal lung, IE: normal colon, Hs888Lu: normal lung, SVCT-M12: normal breast.
12 第4節 JAK/STAT 経路について
JAK/STAT 経路は炎症や分化、がんの悪性化に関与するシグナル伝達経路であ り、その経路の遮断はがんや乾癬、関節性リュウマチ、炎症性腸疾患(Inflammatory
bowel disease, IBD)の治療につながると考えられている。15) 正常細胞において
は、STAT3 などの STAT ファミリーは部分的もしくは一過性に活性化されてい る。IL-6, IL-17, IL-22, IL-23 などのサイトカインや、上皮成長因子(EGF), 血管新 生因子(VEGF), ヒト EGFR 関連物質 2 (HER2)などの増殖因子が細胞膜上のレセ プターに結合することにより、レセプターはリン酸化を受けて活性化され、細胞 内に存在する Januse kinase 1 (JAK1)および Januse kinase 2 (JAK2)をリン酸化する。 リン酸化された JAK1 および JAK2 はさらに STAT3 をリン酸化し、リン酸化を 受けた STAT3 は 2 量体となり核内へ移行し、遺伝子のプロモーター領域に結合 することによりその Bcl-2, Bcl-xL, Fos, Cyclin D, c-Myc, Pim1, Survivin, VEGF と いった下流の遺伝子を発現させる(Figure 7)。
13
Figure 7. JAK/STAT 経路†
† Debnath B, Xu S, Neamati N, Small Molecule Inhibitors of Signal Transducer and Activator of Transcription 3 (Stat3) Protein. J. Med. Chem. 2012;55:6645–6668.のページ 6647 から転載。
14 第5節 臨床応用されている JAK/STAT 経路阻害剤 第 4 節で議論した通り、JAK/STAT 経路の調節は種々の治療効果につながる ことが期待できる。事実、JAK/STAT 経路の阻害剤で臨床応用されているものが いくつかあり、ヒトインターロイキン 6 モノクローナル抗体製剤と JAK 阻害剤 に分類される(Table 3)。ヒトインターロイキン 6 モノクローナル抗体製剤として は、中外製薬のトシリズマブ(アクテムラ)が挙げられ関節リウマチの治療に用 いられている(Table 3)。 一方、最初に臨床応用された JAK 阻害剤はルキソルチニブ (9)である。米国 では 2011 年に骨髄線維症を適応症として承認され、2014 年には真性多血症につ いても承認を受けている。日本においても骨髄線維症および真性多血症を適応 症として承認されている。同効薬であるトファシチニブ (10)は関節リウマチお よび潰瘍性大腸炎を適応症として、バリシチニブ (11)は関節リウマチを適応症 として、デルゴシチニブ(12)はアトピー性皮膚炎を適応症として相次いで承認 された(Table 3, Figure 8)。
15 Figure 8. 臨床応用されている JAK 阻害剤の構造 Table 3 日本で臨床応用されている JAK/STAT 経路阻害剤 成分名 商品名 (開発会社) 作用機序 適応症 トシリズマブ アクテムラ (中外製薬) 抗ヒトインターロ イキン 6 モノクロ ーナル抗体製剤 関節リウマチ シルクマブ (ヤンセンファー マ) 抗ヒトインターロ イキン 6 モノクロ ーナル抗体製剤 関節リウマチ トファシチニブ ゼルヤンツ (ファイザー) JAK 阻害 関節リウマチ 潰瘍性大腸炎 バリシチニブ オルミエント JAK 阻害 関節リウマチ
16 (イーライリリー) ルキソリチニブ ジャカビ (ノバルティス) (ノバルティス/ インサイト) JAK 阻害 骨髄線維症 真性多血症‡ デルゴシチニブ コレクチム軟膏 (JT) JAK 阻害 アトピー性皮膚 炎 ‡ 血液がんの1つ。患者の約 95%に JAK の変異(V617F)が認められる。
17 第6節 STAT3 の阻害剤結合部位 ヒトインターロイキン 6 モノクローナル抗体製剤および JAK 阻害剤の臨床で の成功から、STAT3 自体も創薬標的として魅力的と考えられ、盛んに研究が実 施されている。しかしながら、現在まで承認された医薬品は未だ存在しない。 STAT3 の立体構造(Figure 9)のうち、低分子の結合サイトは現在までに SH2 ド メイン16)、DNA 結合ドメイン17)、クルクミンバインディングドメイン18)、hinge 領域19)の4つが報告されている。 Figure 9. STAT3 の立体構造。§
§ ドメインごとに次のように色分けして表示した。N-terminal Domain (red), Coiled-coil Domain
(cyan), DNA-binding Domain (Magenta), Linker Domain (green), SH2 Domain (yellow), Transactivation Domain (blue), DNA (orange)
18
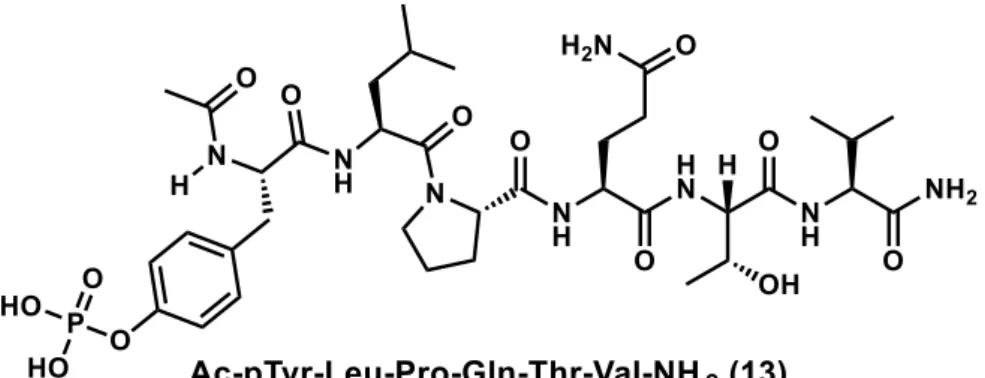
STAT3 の SH2 ドメインは、IL-6 レセプター(IL-6R)の認識部位である。IL-6 な どの刺激により活性化されると IL-6R を構成するタンパク質の1つである gp130 のチロシン残基がリン酸化されるが、SH2 ドメインは gp130 のリン酸基を認識 し、複合体を形成する。それに加え、STAT3 のダイマー化においても重要な役 割を果たしており、リン酸化された STAT3 の Tyr705 を SH2 ドメインが認識す る事によりダイマー化が促進される。このことから SH2 ドメインに結合する低 分子は gp130 との相互作用およびリン酸化された STAT3 のダイマー化の阻害に つながると考えられており、もっとも精力的に創薬研究が展開されている。 第1項 SH2 ドメインに結合する化合物 gp130 のリン酸化を受けるチロシンとその周辺の配列は SH2 ドメインとの高 い親和性を有することから、そのペプチド配列も同様な親和性を有することが 期待される。そのコンセプトをもととしたリン酸化ペプチド
Ac-pTyr-Leu-Pro-Gln-Thr-Val-NH2 (13)は、EMSA アッセイにおいて、STAT3 と DNA との結合を
IC50 = 150 nM で阻害した。20)
19
STAT3 の SH2 ドメインを標的とした他の例として、STA-21 (14)とその類縁体
が挙げられる(Figure 11)。21) STA-21 はミシガン大学の Song らにより見いださ
れた化合物である。Song らは STAT3 の SH2 ドメインに対して 429,00 化合物を バーチャルスクリーニングで評価し、さらに上位 100 化合物をレポータージー ンアッセイで評価した。天然物である Tetrangomycin (15)の類縁化合物である 14 をヒット化合物見出した。14 は 30 µM で STAT3 のリン酸化を抑制し、EMSA ア ッセイで STAT3 ダイマーと DNA の結合を阻害していることが確認された。 STA-21 の類縁体である LLL-6 (16)や 17, LLL-12 (18)についても同様な STAT3 の リン酸化抑制が報告されている。22) Figure 11. STA-21 およびその類縁体の構造 STAT3 の SH2 ドメインを標的とした創薬研究において、臨床段階に進んだ例 としては大塚製薬の OPB-31121 が挙げられる。OPB-31121 の構造は非開示であ
20
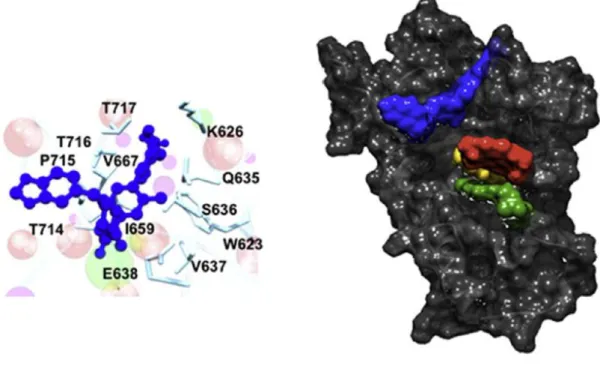
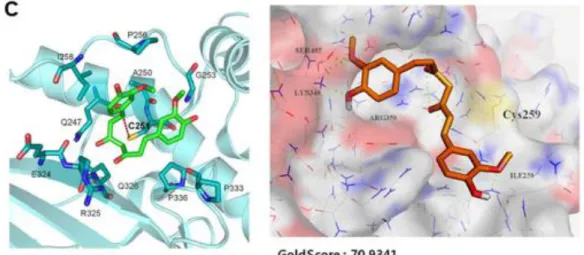
るが、Brambilla らは Autodock 4.3 により OPB-31121 の結合部位を推定し、SH2
ドメイン近傍のアロステリックサイトへの結合が示唆された(Figure 12)。23)
Figure 12. AutoDock 4.3 による OPB-31121 の STAT3 への推定結合部位**)
OPB-31121 の臨床試験については米国で進行肝臓がんを対象とした Phase I/II 試験が 2011 年から 2015 年にかけて実施されたが、Phase III への相移行は確認さ
れていない。24)
**) Brambilla L, Genini, D, Laurini E, Merulla J, Perez L, Fermeglia F, Carbone GM, Pricl S, Catapano CV. Hitting the right spot: Mechanism of action of OPB-31121, a novel and potent inhibitor of the Signal Transducer and Activator of Transcription 3 (STAT3). Mol. Oncol. 2015;9:1194–1206.のページ 1198 か ら転載。
21
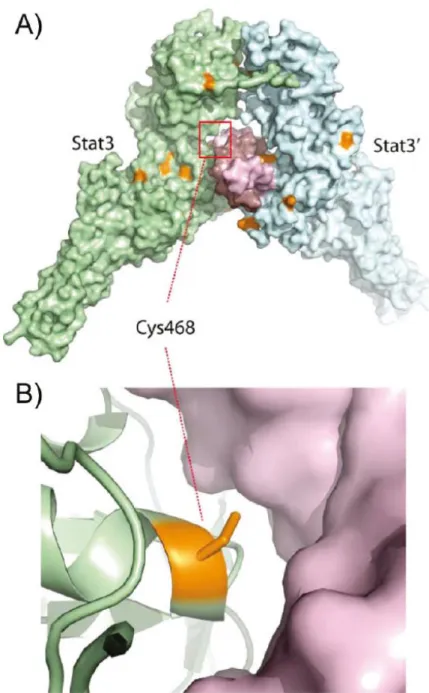
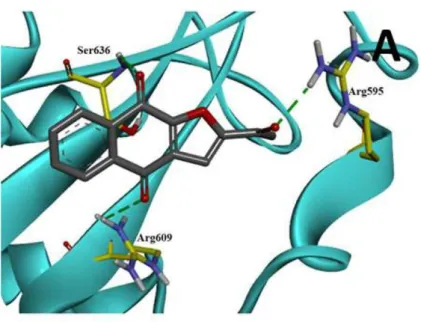
第2項 DNA バインディングドメインに結合する化合物
DNA 結合部位を阻害する低分子に関しては、Beckman Research Institute の Buettner らは、C48 (NSC-368262, )という低分子が STAT3 の DNA 結合領域のシ ステイン残基 Cys468 と結合し、STAT3 の核内移行および、JAK/STAT 経路の下
流の遺伝子発現を抑制していることを報告している。25)
22
Figure 14. C48 (NSC-368262)の STAT3 への結合様式††
†† Buettner R, Corzano R, Rashid R, Lin J, Senthil M, Hedvat M, Schroeder A, Mao A, Herrmann A, Yim
J, Li H, Yuan YC, Yakusijin K, Yakushijin F, Vaidehi N, Moore R, Gugiu G, Lee TD, Yip R, Chen Y, Jove R, Horne D, Williams JC. Alkylation of Cysteine 468 in Stat3 Defines a Novel Site for Therapeutic Development. ACS Chem. Biol. 2011;6:432-443.のページ 435 から転載.
23
第3項 クルクミンバインディングサイトに結合する化合物
近年、ソウル大学の Hahn らはクルクミンの STAT3 中の結合部位はシステイ
ン残基の Cys259 であることを報告している。18) Hahn らは Curcumin (20)中の
エノン構造が STAT3 の機能阻害および結合に重要であると述べている。 Curcumin の還元体である tetrahydrocurcumin (21)は STAT3 阻害活性を示さず、 Cys259 のアラニンへと変異させた STAT3 には阻害活性を有さなかった。
Figure 15. curcumin と tetrahydoxycurcumin の構造
Figure 16. curcumin の結合部位‡‡
‡‡ Hahn Y-I, Kim S-J, Choi B-Y, Cho K-C, Bandu R, Kim KP, Kim D-H, Kim W, Park JS, Han BW, Lee
24 第4項 hinge 領域に結合する化合物
STAT3 の Hinge 領域に結合する化合物として、Boston Biomedical Inc.で創薬さ
れた BBI608(ナパブカシン, 22)が近年報告された。19) 興味深いことに、22 と Tabebuia avellanedae から単離された 8 はいずれもフラノナフトキノン骨格を有 し、22 は微量ながら Tabebuia avellanedae から単離されたという報告がある。構 造と活性の相関を調べるうえでも、Tabebuia avellanedae の薬効面の理解におい ても大変興味深い。 Figure 17. BBI608 の構造
Boston Biomedical 社の Li らは STAT3 ダイマーとオリゴ DNA、BBI608 の 3 成 分による共結晶の作成に成功しており、BBI608 は Hinge 部分、すなわち SH2 ド メインと DNA バインディングドメインの間にあるくぼみに結合することを報 告している(Figure 18, Figure 19)。
induces apoptosis in H-Ras transformed human mammary epithelial cells. Sci. Rep. 2018;9:6409-6422.か ら転載.
25
Figure 18. BBI608 と STAT3 との共結晶§§)
§§ ) Li CJ, Yang A, Rogoff H. Method of Targeting Stat3 and Other Non-druggable Proteins. WO2017023866A1.から転載
26
Figure 19. BBI608 と STAT3 との共結晶(拡大図)§§)
2019 年に Newcastle 大学の Sabanés Zariquiey らは BBI608 の STAT3 への結合
部位を MOE を用いた分子動力学法による解析結果を報告している。26) BBI608
は DNA binding site (DBD)に高確率に存在し、H332, R335, K573 および D570 残 基との相互作用していることが推定され、このシュミレーション結果は Li らの X 線構造解析結果と良好な一致を示している。
27
Figure 20 BBI608 の STAT3 への結合***)
一方で、BBI608 は SH2 ドメインに結合するという推定結果をドッキングシュ ミレーションで予測している報告が有る。沈阳药科大学の Zhoh らは STAT3 の 単結晶構造をもとに AutoDock 4.2.6 で BBI608 との安定化構造を解析しその結合
部位は SH2 ドメインであると推定している。27)
***) Sabanés Zariquiey F, da Souza JV, Estrada-Tejedor R, Bronowska AK. If You Cannot Win Them, Join Them: Understanding New Ways to Target STAT3 by Small Molecule. ACS Omega 2019;4:13913– 13921.から転載
28
Figure 21. Zhoh らによる BBI608 のバインディングサイトの推定***)
また、四川大学の Li らは Discovery Studio (DS)3.1 ソフトウェアを用いた分子 動力学計算により BBI608 は STAT3 の SH2 ドメインに結合するものと推定し
た。28)
Figure 22. Discovery Studio (DS)3.1 による BBI608 の結合部位の推定†††)
††† Li C, Chen C, An Q, Yang Tao, Sang Z, Yang Y, Ju Y, Tong A, Luo Y. A novel series of napacucasin
derivatives as orally active inhibitors of signal transducer and activator of transcription 3 (STAT3). Euro.
29
BBI608 の STAT3 への直接的な結合を示唆する報告がある一方で、BBI608 は STAT3 の リ ン 酸 化 自 体 を 抑 制 す る と い う 報 告 が あ る 。 廈 門 大 学 (Xiamen University)の Li らは、BBI608 が Molm13 細胞(白血病由来)の STAT3 リン酸化
を抑制することを報告している。29) Boston Biomedical Inc で実施された X 線構
造解析は Tyr705 がリン酸化された STAT3 をもとに実施されており、STAT3 の リン酸化自体を抑制するという報告は BBI608 の STAT3 hinge 領域への結合とい う事象に基づいて説明することは困難であり、オフターゲットへの結合ならび に、hinge 領域とはことなる領域での STAT3 との相互作用に関して議論の余地 を残している。 第5項 STAT3 阻害剤研究の今後の可能性 第 6 節の第 1 項から第 4 項にかけて、STAT3 の低分子の標的部位に関して議 論し、現在まで 4 つの低分子が結合する部位が報告されていることを紹介した。 STAT3 阻害剤が臨床応用に至っていない理由として、以下の可能性は考慮すべ きであろう。 ①SH2 ドメインを標的とした研究に集中しているが、そもそも SH2 ドメインは リン酸化されたチロシンを認識する部位である。活性を保持しつつ物性や代謝 安定性が良好なリン酸化チロシンのバイオアイソスターの開発が求められる。 一方で、OPB-31121 のような SH2 ドメイン近傍のアロステリックサイトに結合 する低分子はリン酸基バイオアイソスターが不要と考えられ魅力的である。 ②SH2 ドメイン以外の部位へ結合する低分子の研究開発が不十分である。 cysteine468(DNA バインディングドメイン)、Cys259(クルクミンバインディン グサイト)に結合する低分子はアカデミアからのみ報告されており、周辺化合物 への創薬展開は不十分である。また、hinge 領域を標的とした創薬展開は Boston
30 Biomedical Inc,で実施され、併用療法での臨床試験が実施中である。30) 一方で、 BBI608 のバックアップとなる低分子の報告はほとんどない。分子量はわずか 240 であり、さらなる SAR 研究の余地を残している可能性があり、オフターゲット へ選択性の改善や ADME プロファイルの向上の創薬展開が期待される。 第7節 BBI608 の生物活性
第6節で議論したとおり、BBI608 は Boston Biomedical Inc.で創薬されたナフ トキノン骨格をもつ STAT3 阻害剤である。その in vitro および in vivo での薬理
作用については Li らにより報告されている。31) PaCa2(膵臓がん由来)細胞移 植マウスモデルにおいて、BBI608 を 3 日おきに 20 mg/kg を腹腔投与したとこ ろ、がん病変部位の増大を抑制した。興味深いことに、投与完了後(day 41)移行 の経過観察時において、がん病変部位の増大は確認されなかった。一方、対照と した既知抗がん剤であるゲムシタビンは、PaCa2(膵臓がん由来)細胞移植マウ スモデルにおいてがん病変部位の増大を抑制したが、投与終了後の経過観察期 間に劇的ながん病変部位の増大が確認された。BBI608 は休薬期間中も持続して がんの増殖抑制効果があるのは大変興味深い事象である。 Li らはさらに、がん幹細胞仮説に基づく検証を in vitro で行っている。がん幹 細胞の定義は不明確で仮説段階では有るが下記の2点を特徴とする。 ① がん病変を構成するがん細胞は、同一クローンを起源としながらも、一様で はなく、分化の度合いや増殖のしやすさが異なる。 ② 幹細胞能な増殖能を有するがん細胞ががんの悪性化や薬剤耐性、化学療法後 の再発に関与している。
31 がん幹細胞仮説に基づき Li らは検証を行った。まず、各種がん細胞に対して、 がん細胞の 90%程度が死滅する程度の抗がん剤で処理をし、生き残った細胞(薬 剤耐性)を培養することにより、がん幹細胞の性質を有する細胞を得た。そのが ん細胞は既存の化学療法の化合物に対しては耐性を有していたが、BBI608 に対 しては成長阻害が確認された。 一方でウェスタンブロットでは JAK/STAT 経路の下流に相当する各種タンパ ク質の発現を抑制した一方で、JAK/STAT 経路との関連が薄いと考えられてい る、Bmi-1, β-catenin, Klf などのタンパク質の発現も合わせて抑制していることか ら、オフターゲットへの作用、とりわけ Wnt シグナルへの阻害作用の可能性に ついては考慮するべきであろう。 BBI608 と NQ801 はいずれもフラノナフトキノンを持ち、構造が類似している が、興味深いことに BBI608 は Tabebuia avellanedae から単離されたという報告 があり、JAK/STAT 経路の阻害活性と Tabebuia avellanedae の関係性は興味をそ
32
第2章 本論
第1節 研究の目的 第 1 章で述べたとおり、Tabebuia avellanedae の抽出エキスには抗腫瘍効果を 始めとする様々な生理活性を有するが、エキスに含まれる成分のうち、どの化合 物が実際の生理活性を発揮しているのか不明な点が多い。本研究では、Tabebuia avellanedae に含まれる NQ801 を始めとするフラノナフトキノン類と、臨床段階の STAT3 阻害剤である BBI608 や、STAT3 の SH2 ドメインに結合するとされて いる STA-21 や LLL12 との構造類似性に着目し、「Tabebuia avellanedae に含ま れるナフトキノン類のいくつかは JAK/STAT 経路の阻害活性を有し、抽出エキ スの抗腫瘍効果の一部を担っている」と仮説を立てた。本研究では、Tabebuia
avellanedae に含まれるナフトキノン類の STAT3 リン酸化抑制作用を評価し、さ
らに標的タンパク質の推定のために、JAK のキナーゼ阻害活性、STAT3 の SH2 ドメインへの結合の評価ならびに、ドッキングシミュレーションによる標的部 位の推定を行った。SAR (structure activity relationship)解析を行い活性発現に必要 な部分構造を明らかにすることを目的とした。さらには、Tabebuia avellanedae は ナフトキノン類以外にもリグナンやイリドイド、フェニルプロパノイドなどの 天然物が含まれていることから、それら天然物からの JAK/STAT 阻害活性物質 の探索を行うことを本研究の目的とした。
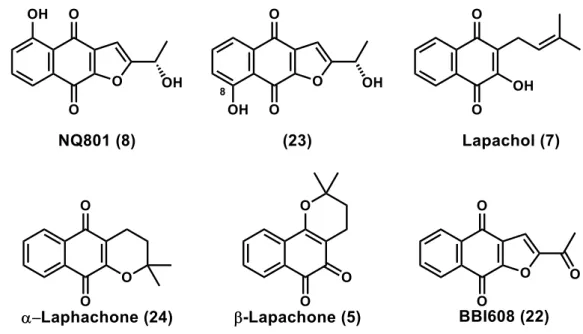
33 第2節 ウェスタンブロットによる評価
Tabebuia avellanedae に含まれる代表的なナフトキノンである NQ801 (8), (S)-8-hydroxy-2-(1-hydroxyethyl)naphtho[2,3-b]furan-4,9-dione (23), lapachol (7), α-lapachone (24), β-α-lapachone (5)および BBI608 (22)の 6 化合物(Figure 23)についてウ ェスタンブロットにより STAT3 のリン酸化抑制能の評価を行った。MCF7(乳 がん由来)を用い、無血清培地で 24 時間培養したのち、評価化合物もしくは溶 媒コントロール(0.1% DMSO)を加え、その 1 時間後に IL-6(10 ng/mL)で 1 時間刺 激し、RIPA バッファーを加え可溶化画分を回収してライセート調製をおこなっ た。
Figure 23. Tabebuia avellanedae に含まれる代表的なナフトキノン
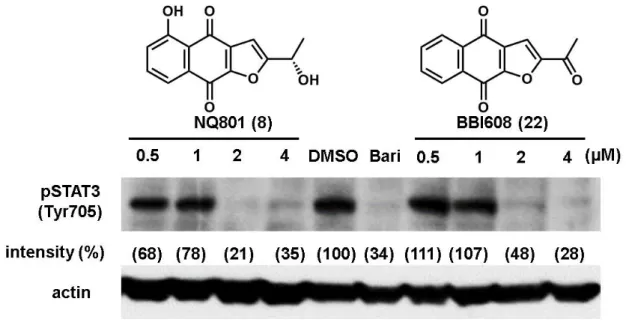
代表的なナフトキノン 6 化合物のウェスタンブロットによる評価結果を
Figure 24 に示す。IL-6 刺激により STAT3 のリン酸化の亢進が確認され、既知の
JAK 阻害剤であるバリシチニブにより pSTAT3 の抑制が確認されたことから当 該ウェスタンブロットは評価系として利用可能と判断した。評価した Tabebuia
34 avellanedae 由来ナフトキノン 6 化合物のうち、化合物 8, 23, 22 に pSTAT3 の抑 制作用が認められ、とりわけ 22 は 1 µM でほぼ完全な抑制を示した。一方化合 物 7, 24, 5 は顕著な pSTAT3 の抑制作用は確認されなかった。化合物 8, 23, 22 は いずれもフラノナフトキノン骨格を有しているのに対し、7, 24, 5 はフラン環を 有していないことから、フラノナフトキノン骨格の重要性が示唆された(Figure 25)。一方、STAT3 の総量(tSTAT3)に対する顕著な変化は確認されなかった。 Figure 24. MCF7 に対するナフトキノン類の pSTAT3 の抑制作用 Figure 25. フラノナフトキノン骨格 フラノナフトキノン骨格が pSTAT3 抑制作用に重要であると知見がえられた ことから、次にフラノナフトキノンの 2 位の側鎖の酸化段階ならびに 5 位水酸 基の有無による pSTAT3 抑制作用について評価した。
35
評価実施に際し、2-acetyl-5-hydroxynaphtho[2,3-b]furan-4,9-dione (25)および(S)-2-(1-hydroxyethyl)naphtho[2,3-b]furan-4,9-dione (26)を評価対象化合物として追加し た(Figure 26)。化合物 25 および 26 はいずれも Tabebuia avellanedae から単離さ れている天然物である。 Figure 26. 化合物 25 および 26 の構造 SAR の結果を Figure 27 に示す。化合物 8, 22, 25 および 26 はいずれも用量依 存的に STAT3 のリン酸化を抑制した。5 位に水酸基をもたない化合物 22 と 26 は、5 位に水酸基をもつ 8 および 25 と比較して、強いリン酸化抑制作用を示し、 その IC50値は 1 µM 以下であった。一方、化合物 8 と 25 の STAT3 リン酸化抑制 作用は同程度であり、かつ化合物 6 と 8 のリン酸化抑制作用は同程度であった。 2 位側鎖の酸化段階(水酸基 or ケトン)による顕著な違いは認められなかった。
36 Figure 27. 4 化合物による SAR IL-6 刺激下での MCF7 に対してフラノナフトキノン類は用量依存的に STAT3 のリン酸化抑制作用を認めたことから、次に JAK/STAT 経路が恒常的に活性化 されている乳がん由来細胞の MDA-MB-231 を用いて評価を行った。10%ウシ血 清で培養し化合物を 2 時間暴露しライセートを調製しウェスタンブロットを実 施した。化合物 8 および 22 は用量依存的に STAT3 のリン酸化を抑制した。
37
Figure 28. MDA-MB-231 に対する STAT3 リン酸化抑制作用
Tabebuia avellanedae にはナフトキノン類以外にもリグナン、イリドイド、フ ェニルプロパノイドなどの天然物が単離されている。Tabebuia avellanedae の抗 腫瘍効果はそれら天然物に基づく可能性も否定できないことから、Tabebuia avellanedae に含まれる代表的なリグナン 2 化合物 (27, 28)とイリドイド 2 化合 物 (29, 30)について STAT3 のリン酸化抑制作用について評価を実施した(Figure 29, Figure 30)。
38
Figure 29. リグナン (27, 28)およびイリドイド (29, 30)の構造
Figure 30. リグナン (9, 10)、イリドイド (11, 12)のウェスタンブロット結果
化合物 30 の 10 µM でのみ弱い STAT3 リン酸化抑制作用が確認されたが、顕 著な抑制作用は認められなかった。
39 第3節 WST8 assay によるがん細胞に対する増殖抑制評価 フラノナフトキノン類は STAT3 のリン酸化を抑制したことから、MDA-MB-231, MCF7, A549 に対する増殖抑制を検証した。アッセイ系は MTT assay に類似 した WST8 assay を選択した。その理由として、MTT assay と比較して操作が簡 便であること、データのばらつきが少ないとの文献情報があったためである。ウ ェスタンブロットで評価した Tabebuia avellanedae に含まれるナフトキノン 8 化 合物に対して評価を実施した。化合物処置後 72 時間培養したのちに吸光度を測 定した。 Table 4 WST8 assay によるがん細胞の増殖抑制評価結果
Compound Growth inhibition (IC50 µM)
MDA-MB-231 MCF7 A549
0.41 0.50 0.13
0.33 1.11 0.28
>30 >30 >30
40 1.39 0.71 1.96 0.25 0.24 0.12 0.17 0.051 0.044 1.59 0.28 0.72 WST8 assay の結果を Table 4 に示す。フラノナフトキノン骨格を持たたない 7 および 24 は最大濃度(30 µM)で顕著な増殖抑制を示さなかった。化合物 5 の IC50 値は 0.71~1.96 µM であり、中程度の増殖抑制を示した。一方、フラノナフトキ ノン骨格を有する化合物 8, 23, 22, 25, 26 はいずれも強い増殖抑制を示した。化 合物 8 と 25 の比較および化合物 22 と 26 の比較したところ、2 位側鎖がケトン 型である化合物 22 と 25 に強い増殖抑制効果が認められた。ウェスタンブロッ トによる STAT3 リン酸化抑制作用では、2 位側鎖の酸化段階は顕著な変化を示 さなかったことから、STAT3 のリン酸化抑制作用とがん細胞の増殖抑制作用が 一致しない興味深い結果となった。
41 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 8 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 4 1 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 5 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 1 . 3 9 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 3 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 3 3 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 2 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 2 5 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 7 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 > 3 0 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 5 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 1 7 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 4 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 > 3 0 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 6 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 1 . 5 9 M Figure 31. MDA-MB-231 に対するフラノナフトキノン類の細胞増殖抑制効果
42 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 8 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 5 0 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 5 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 7 1 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 3 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 1 . 1 2 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 2 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 2 4 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 7 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 > 3 0 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 5 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 0 5 1 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 4 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 > 3 0 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 6 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 2 8 M Figure 32. MCF7 に対するフラノナフトキノン類の細胞増殖抑制効果
43 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 8 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 1 3 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 5 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 1 . 9 6 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 3 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 2 8 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 2 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 1 2 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 7 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 > 3 0 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 5 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 0 4 4 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 4 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 > 3 0 M 0 5 0 1 0 0 - 9 - 8 - 7 - 6 - 5 - 4 C o m p o u n d 2 6 c o n c e n t r a t i o n ( m o l / L ) I n t e n s i t y ( % ) 0 I C5 0 = 0 . 7 2 M Figure 33. A549 に対するフラノナフトキノン類の細胞増殖抑制効果
44 第4節 キナーゼアッセイ
Tabebuia avellanedae に含まれるフラノナフトキノンに STAT3 のリン酸化抑制 作用が認められたことから、標的タンパク質の推定を試みた。第1章で議論した 通り、JAK/STAT 経路の阻害活性を有する化合物の標的部位の多くは JAK、もし くは STAT3 の SH2 ドメインである。このことから、Tabebuia avellanedae に含ま れるフラノナフトキノン類が JAK のキナーゼ活性を阻害するのかを本説で議論 し、第4節では STAT3 の SH2 ドメインへの結合能についての評価結果を述べ る。
JAK のキナーゼ活性阻害評価については、カルナバイオサイエンス株式会社 に委託し MSA (mobility shift assay)を評価法として用いた。アッセイ原理の概略 を Figure 34 に示す。
Figure 34. mobility shift assay の概略
キナーゼとは ATP をエネルギー源として基質をリン酸化する酵素である。 JAK/STAT 経路の場合、JAK がキナーゼ、STATs が基質に相当する。今回用い た MSA では、基質となる STAT3 の部分構造(フラグメント)を蛍光標識をし た組み換えタンパク質をあらかじめ作成しておき、評価したい化合物との共存 下で JAK が介するキナーゼ反応を行った。評価化合物に JAK キナーゼ阻害活性 がある場合は、リン酸化が抑制される。キャピラリー電気泳動により電荷の違い (リン酸基の有無)で分離し、それぞれのバンドを蛍光検出で定量した。STAT3
45
のリン酸化に関与しているのは JAK1 と JAK2 であるが、JAK 間での選択性確認 のため、JAK1, JAK2 に加え、JAK3 の評価も併せて行った。
NQ801 の JAK キナーゼ阻害活性の結果を Figure 35 に示す。NQ801 は、JAK1, JAK2, JAK3 に対して顕著な阻害活性を示さなかった。
Figure 35. NQ801 のキナーゼ阻害活性
同様に BBI608 は JAK1, JAK2, JAK3 に対して阻害活性を示さなかった。
Figure 36. BBI608 のキナーゼ阻害活性 0 20 40 60 80 100 120 0.1μM 1μM 10μM 100μM K in ase act ivit y (%)
NQ801
JAK1 JAK2 JAK3 0 20 40 60 80 100 120 0.1μM 1μM 10μM 100μM K in ase act ivit y (%)BBI608
JAK1 JAK2 JAK346
第5節 STAT3 および STAT1 の SH2 ドメインに対するバインディングアッセ イ
NQ801 および BBI608 は JAK1, JAK2, JAK3 に対して顕著な阻害活性を示さな かったことから、次に STAT3 の SH2 ドメインに対する結合能を TR-FRET assay
で評価した。33)
STAT3 の SH2 ドメインは IL-6R のリン酸化部位の認識ならびに、リン酸化さ れた STAT3 がホモダイマーを形成する際に相手側のリン酸基を認識する部位で ある。このことからまず、IL-6R (gp130)の部分構造(ホスホペプチド)を文献に
従い合成した。33b) このホスホペプチドを蛍光団である FAM で標識して評価
に用いた(Figure 37)。STAT1 は、IFN-γ の刺激によりリン酸化されていることが
知られている。STAT1 は IFN-γ のリン酸化チロシンを SH2 ドメインで認識する。
STAT3 と STAT1 の選択性について評価するため、IFN-γ にリン酸化チロシンを 含むペプチド配列とその FAM 標識体についても同様に合成し、バインディング アッセイの材料とした(Figure 38)。
47
Figure 37. FAM-Gly-pTyr-Leu-Pro-Gln-Thr-Val-NH2 および
48
Figure 38. FAM-Gly-pTyr-Asp-Lys-Pro-His-Val-Leu-NH2 および Ac-Gly-pTyr-Asp-Lys-Pro-His-Val-Leu-NH2 の構造(IFN-γR 由来)
49 一方、STAT3 は GST タグを融合させたリコンビナントタンパク質を準備した。 FAM-Gly-pTyr-Leu-Pro-Gln-Thr-Val-NH2 と GST 融合 STAT3 が結合した状態で 340 nm の励起波長で光照射すると FAM からテルビウムへのエネルギー遷移が 起こり、その結果として 490 nm および 520 nm での蛍光が観察される。SH2 ド メインに結合する低分子が存在する場合、490 nm および 520 nm の蛍光が減弱す るため、その蛍光強度から SH2 ドメインでの拮抗阻害を評価するアッセイ系で ある。
Figure 39. IL-6 刺激による JAK/STAT 経路の伝達
バインディングアッセイの結果を Figure 40 に示す。NQ801 は 100 µM での高 濃度で STAT3 に対して阻害率 30.8%の部分的な阻害が認められるのみであった。 一方、STAT1 の SH2 ドメイン対しては拮抗阻害を認めなかった。
50
Figure 40. STAT3 の SH2 ドメインに対するバインディングアッセイの結果
51
第6節 Autodock による STAT3 中の結合部位の推定
第2節では、NQ801 および BBI608 が STAT3 のリン酸化を抑制することを示 し、第 4 節では NQ801 と BBI は JAK キナーゼへの阻害活性を有しないこと、 第 5 節では NQ801 は STAT3 の SH2 ドメインへの結合能を有していないことを 明らかにした。そのことから、NQ801 は BBI608 と同様に STAT3 の hinge 領域 に結合している可能性がある。STAT3 の hinge 領域へ結合の推定のため、 Autodock 4.2.6 によるドッキングシュミレーションを行った。 低分子については、Chem 3D による MM2 計算で再安定のコンフォメーショ ンを算出して用いた。STAT3 タンパク質の立体構造については、マウス STAT3 ダイマーとオリゴ DNA の共結晶である 1BG1 を用い、Autodock 4.2.6 でドッキ ングシュミレーションを実施し、PyMOL でビジュアル化を行った。34)
52 第1項 NQ801 の結果
NQ801 のドッキングシュミレーションの結果を Figure 41 に示す。NQ801 はリ ンカードメイン(緑)と DNA バインディングドメイン(マゼンタ)の間かつ DNA (オレンジ)の近傍に入り込んでいる。さらに Asp334(主鎖 NH), Arg335, Asp570, Lys573, Lys574 との水素結合形成が示唆された。
Figure 41. NQ801 の推定結合部位
また、もっともクラスターランクの高いバインディングポーズの Mean Binding Energy は–6.14 kcal/mol であった。
53
Cluster rank
Lowest Binding Energy (kcal/mol)
Run Mean Binding
Energy (kcal/mol) Num in clus 1 -6.21 88 -6.14 75 2 -6.04 12 -6.04 1 3 -5.90 63 -5.75 8 4 -5.64 57 -5.34 5 5 -4.96 75 -4.95 11 6 -4.96 51 -4.96 1
N-terminal Domain (red), Coiled-coil Domain (cyan), DNA-binding Domain (Magenta), Linker Domain (green), SH2 Domain (yellow), Transactivation Domain (blue), DNA (orange)
54 第2項 BBI608 の結果
BBI608 のドッキングシュミレーション結果を Figure 42 に示す。NQ801 ど同 様に hinge 領域への結合が示唆され、Boston Biomedical Inc,の X 線構造解析結果 ならびに Sabanés ら all-atom molecular dynamic (MD) simulation による解析結果と 良好な一致を示した。もっともクラスターランクの高いバインディングポーズ の Mean Binding Energy は–6.58 kcal/mol であった。
Figure 42. BBI608 の推定結合部位
Cluster rank Lowest Binding Energy (kcal/mol)
Run Mean Binding
Energy (kcal/mol) Numin clus 1 -6.58 75 -6.58 88 2 -6.27 77 -6.14 5 3 -5.92 23 -5.90 7
55 第3項 化合物 25 の結果
同様に 25 についてもドッキングシュミレーションを実施し、hinge 領域への 結合が示唆され、もっともクラスターランクの高いバインディングポーズの Mean Binding Energy は–6.57 kcal/mol であった。
Figure 43. 化合物 25 の推定結合部位
Cluster rank Lowest Binding Energy
Run Mean Binding
Energy
Num in clus
1 -6.71 3 -6.57 87
2 -5.55 10 -5.54 9
56 第4項 化合物 26 の結果
同様に 26 ついてもドッキングシュミレーションを実施し、hinge 領域への結 合が示唆され、もっともクラスターランクの高いバインディングポーズの Mean Binding Energy は–6.06 kcal/mol であった。
Figure 44. 化合物 26 の推定結合部位
Cluster rank Lowest Binding Energy
Run Mean Binding
Energy Num in clus 1 -6.41 59 -6.06 68 2 -5.80 47 -5.65 15 3 -5.51 100 -5.50 3 4 -5.33 6 -5.32 14
57 第5項 【参考】4 化合物の重ね合わせ
第 1 項~第 4 項で述べた NQ801、BBI608、化合物 25、化合物 26 の STAT3 に対 するバインディングポーズの重ね合わせを Figure 45 に示す。
58 第7節 レポータージーンアッセイによる NQ801 および BBI608 の評価 第1項 レポータージーンアッセイの概略 第1節から第5節にかけて、ナフトキノン類の JAK/STAT 経路への阻害活性 を評価し、フラノナフトキノン骨格が STAT3 のリン酸化抑制に必要な部分構造 であることならびに、JAK キナーゼ阻害活性を有しないことならびに STAT3 の SH2 ドメインへの結合能を有していないことを明らかにし、さらには STAT3 の Hinge 領域へ結合している可能性を Autodock によるシュミレーションで明らか にした。そのことから NQ801 および BBI608 が JAK/STAT 系の下流のシグナル についても阻害しているかを確認することを目的としてレポータージーンアッ セイを実施した。 レポータージーンアッセイとは、レポーター遺伝子と呼ばれる遺伝子を細胞 に導入し、ある特定の転写産物の量を吸光度もしくは発光強度を用いて評価す るアッセイ系のことで、吸光度検出の場合は SEAP (Secreted Alkaline Phosphatase, 分泌型アルカリフォスファターゼ)が、発光検出の場合はルシフェラーゼが汎用 される。
本研究で用いた HEK-Blue IL-6 assay kit に含まれる HEK-Blue IL-6 細胞は、 HEK293 細胞に、SEAP(分泌型アルカリホスファターゼ)レポーター遺伝子があ らかじめ導入された細胞である。IL-6 刺激により JAK/STAT 経路が活性化され、 SEAP が細胞外に放出されるが、JAK/STAT 経路を阻害する低分子化合物が共存 する場合その SEAP 産生量が低下する。
59
Figure 46. HEK-Blue IL-6 細胞によるレポータージーンアッセイの概略
HEK-Blue IL-6 細胞は、SEAP をコードしたレポーター遺伝子が導入済みの細
胞であること、その遺伝子発現が安定‡‡‡であること、キットとして購入可能であ ることから今実験で使用した。 ‡‡‡ ここでいう「安定とは」HEK293 細胞に組み込まれた外来遺伝子が stable である。すなわち 一過性ではなく継代を繰り返しても外来遺伝子が失われないことを意味している。なお、HEK-Blue-IL-6 細胞は継代の際に外来遺伝子を失った HEK293 細胞に対して選択的に毒性を発揮する 抗生剤カクテル(HEK-BlueTM Selection)を共存させることによりアッセイ系の品質を担保してい る。
60 第2項 IL-6 濃度の最適化
Figure 47. IL-6 濃度の最適化
レポータージーンアッセイの実施に際し、まずは IL-6 濃度の設定を行った。 HEK-Blue IL-6 アッセイキットに記載されている IL-6 の推奨濃度は 0.03 – 10 ng/mL であることから、0.03 – 30 ng/mL の範囲で IL-6 の至適濃度を検討した。3 ng/mL のときに最大濃度(30 ng/mL)の 8 割程度の吸光度が確認されたことから 3 ng/mL に決定した。 第3項 化合物の濃度設定 第 1 項で述べたレポータージーンアッセイの原理上、評価化合物が細胞毒性 を有している場合、細胞は死滅し SEAP は産生されなくなることから「フォルス ポジティブ」として検出される。そのことから、NQ801 と BBI608 のレポーター ジーンアッセイによる評価に先立ち、HEK-Blue IL-6 細胞に対しての NQ801 と BBI608 の成長阻害を WST8 assay で評価した。 0 20 40 60 80 100 120 30 10 3 1 0.3 0.1 0.03 (%) 濃度 (ng/mL)
61
Figure 48. HEK-Blue IL-6 細胞に対する NQ801 の成長阻害
Figure 49. HEK-Blue IL-6 細胞に対する BBI608 の成長阻害
NQ801 は 1 µM から濃い濃度で、BBI608 は 3 µM から濃い濃度で顕著な成長 阻害が確認されたためレポータージーンアッセイはより低濃度の 0.1 µM で評価 し、レポータージーンアッセイで用いた 96 well プレートで WST8 assay も併せ て行うことにより 1 µM での評価も併せて実施した。 0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4 0.45 30µM 10µM 3µM 1µM DMSO
NQ801
0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4 0.45 30µM 10µM 3µM 1µM DMSOBBI608
62 第4項 ポジティブコントロールの選定 レポータージーンアッセイ実施に当たり、ポジティブコントロールとしてバ リシチニブ (11)を用いた(Figure 8)。バリシチニブは第 1 章で第 5 節で述べたと おり、臨床応用されている JAK 阻害剤である。イーライリリーで創薬された経 口の JAK1/JAK2 阻害薬であり、細胞毒性が少ないことならびに JAK/STAT 経路 への阻害活性が強力であることが選定の根拠である。 第5項 レポータージーンアッセイの結果 NQ801 および BBI608 のレポータージーンアッセイの結果を Figure 50 に示す。 NQ801 は 0.1 µM および 1 µM の濃度において JAK/STAT 経路への阻害活性を示 さなかった。同様に BBI608 は 0.1 µM および 1 µM の濃度において阻害活性を 示さなかった。なお、レポータージーンアッセイで用いた同一の 96-well プレー トで WST8 assay を実施し、0.1 µM および 1 µM の濃度域において、NQ801 およ び BBI608 はほとんど細胞毒性を示さなかった(Figure 51)。第5章第2節で議論 した通り、ウェスタンブロットで NQ801 および BBI608 は用量依存的に STAT3 のリン酸化を抑制したが、レポータージーンアッセイでは化合物の細胞毒性の ため、1 µM よりも高い濃度ではアッセイを実施することができなかった。その 理由は不明であるが、バリシチニブがほとんど毒性を示していないことから、 NQ801 および BBI608 の JAK/STAT 経路以外のオフターゲットへの影響は懸念 事項として挙げられる。
63
Figure 50. NQ801 および BBI608 のレポータージーンアッセイの結果
Figure 51. NQ801 および BBI608 の WST8 assay による生細胞数の確認
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
reportergene assay
1μM 0.1μM 0 0.1 0.2 0.3 0.4 0.5 0.6WST-8
1 μM 0.1 μMCompound DMSO DMSO Baricitinib NQ801 BBI608
IL-6 - + + + +
Compound DMSO DMSO Baricitinib NQ801 BBI608
64
第8節 Reportergene assay による JAK/STAT 阻害剤の探索 第1項 ヒット化合物の同定 第1節から第6節にかけて、ナフトキノン類の JAK/STAT 経路への阻害作用 について議論を進め、NQ801 およびその周辺化合物はウェスタンブロットで STAT3 のリン酸化を用量依存的に抑制することを明らかにし、NQ801 は JAK の キナーゼ活性は阻害しないこと、STAT3 の SH2 ドメインへの結合能を有しない ことを明らかにした。また NQ801 およびその周辺化合物は Autodock によるド ッキングシミュレーションにて、STAT3 の hinge 領域に結合するという結果を 得た。このことから、Tabebuia avellanedae の抗腫瘍活性効果は NQ801 を始めと するフラのナフトキノン類が関与している可能性が示された。 しかしながら、Tabebuia avellanedae が有する抗腫瘍活性の評価を十分に行った
とは言い難く、他の Tabebuia avellanedae 天然物やその類縁体について JAK/STAT
経路への阻害能を評価する意義はあるものと考えた。
そこで、第 7 節で議論したレポータージーンアッセイを活用してより広範囲 に JAK/STAT 経路の阻害活性を有する化合物の探索を行った。
65 Figure 52 レポータージーンアッセイの結果2 Figure 53 WST-8 の結果2 0 0.02 0.04 0.06 0.08 0.1 0.12 0.14 0.16 0.18 reportergene assay 1µM 0.1μM 0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4 0.45 0.5
WST-8
1 μM 0.1 μMCompound DMSO DMSO Baricitinib NQ011 NQ-a039 MY-k098 KU30
IL-6 - + + + + + +
Compound DMSO DMSO Baricitinib NQ011 NQ-a039 MY-k098 KU30
66 次に、KU30 の IC50値を算出するために最大濃度30 μM、公比 3 でレポータージ ーンアッセイを実施したところ、その IC50値は 0.050 µM であった。 0 5 0 1 0 0 - 8 - 6 - 4 N Q 0 0 8 m o l / L % 0 I C5 0 = 0 . 0 5 0 M Figure 54. NQ008 の IC50値 レポータージーンアッセイで NQ008 の JAK/STAT 経路への阻害活性を確認で きたため、次にウェスタンブロットで STAT3 のリン酸化への影響を評価した。 Figure 55. NQ008 の STAT3 リン酸化阻害活性
67 NQ008 は STAT3 のリン酸化を用量依存的に抑制し、8 µM, 4 µM, 2 µM での阻 害率はそれぞれ 96%, 74%, 72%であった。ウェスタンブロットにおいても阻害活 性がみられたため、次に NQ008 のがん細胞に対する成長阻害を評価した。 Table 5. NQ008 のがん細胞に対する成長阻害 Compound
Growth inhibition (IC50 µM)
MDA-MB-231 MCF7 A549
NQ008 0.50 0.77 0.49
NQ008 は MDA-MB-231, MCF7, A549 に対し同程度の成長阻害作用を示し、そ
68 第2項 レポータージーンアッセイによるスクリーニング結果 レポータージーンアッセイにより、さらなるスクリーニングを実施したが、ヒ ット化合物として得られたのは NQ008 の1化合物のみであった。レポータージ ーンアッセイについては生命資源化学研究室で保有する Tabebuia avellanedae に 含まれる天然物、合成中間体、その他の植物から単離された天然物など、約 100 化合物に対してスクリーニングを実施したが、NQ008 以外にヒット化合物とし て見出された化合物は皆無であった。