2013 年 4 月 2 日 独立行政法人 理化学研究所 国立大学法人 東京大学
21 番目のアミノ酸「セレノシステイン(Sec)」の合成メカニズムを解明
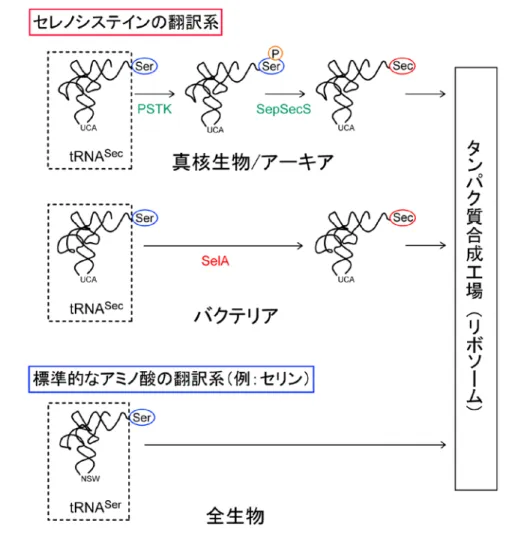
-星形の超巨大複合体が Sec 合成を一度に成し遂げる- 本研究成果のポイント ○Sec の合成に必要な酵素「SelA」の立体構造を決定 ○星形の巨大タンパク質「SelA」の4つのサブユニットが異なる作業を担い Sec を合成 ○セレン(Se)の自在な導入によるスーパー酵素の創生などへ期待 理化学研究所(野依良治理事長)と東京大学(濱田純一総長)は、バクテリアにお ける「21 番目のアミノ酸※1」と呼ばれるセレノシステイン(Sec)の合成メカニズム を解明しました。これは、理研生命分子システム基盤研究領域の横山茂之領域長(横 山茂之領域長、現:横山構造生物学研究室 上席研究員)、東京大学分子細胞生物学 研究所の伊藤弓弦助教らと米国イェール大学による共同研究成果です。 老化防止や生活習慣病予防などに有効な元素のセレン(Se)は、ヒトを含む幅広い 生物にとって微量成分として不可欠で、欠乏すると、がんや高血圧症を引き起こしま す。生体内ではSecの中に存在し、アミノ酸の配列を決める暗号(遺伝暗号)により、 一部のタンパク質に取り込まれて機能します。Secは、専用の転移RNA(tRNA)※2 であるtRNASecに結合した状態の別のアミノ酸を材料として合成されますが、ヒトを 含めた真核生物※3とアーキア※4のグループ(ヒト型)とバクテリア※5のグループ(バ クテリア型)では、その合成メカニズムが異なります。これまでヒト型については、 その詳細が明らかになってきましたが、バクテリア型については不明のままでした。 共同研究グループは、バクテリアにおいてSecを合成する酵素SelAとtRNASecの複 合体の結晶構造を解析し、SelAが10個のサブユニット※6からなる超巨大な星形をして いることが分かりました。また、サブユニットそれぞれの詳細な機能も明らかにし、 星形を構成することが、Secの合成に必要不可欠であることも突き止めました。 この成果は、Se 含有タンパク質の合成方法の開発に大きく貢献し、今まで不可能 だった天然の酵素の機能を上回るスーパー酵素の創生や、Se 欠乏を原因とする疾患 の研究などに役立つと期待できます。 本研究成果は、ターゲットタンパク研究プログラム、および、文部科学省科学研究 費補助金の支援を受けて行われ、米国の科学雑誌『Science』(4 月 5 日号)に掲載さ れます。 本件の取り扱いについては、下記の解禁時間以降でお願い申し上げます。 新聞 :日本時間 4 月 5 日(金)朝刊 テレビ・ラジオ・インターネット :日本時間 4 月 5 日(金)午前 3 時1.背 景 セレン(Se)は、周期表の酸素と硫黄の下に位置し、硫黄と似た性質を持ちますが、 より反応性に富んでいます。そのため、ヒトからバクテリアに至る幅広い生物にとっ て微量成分として不可欠で、Se の欠乏は、がんや高血圧症を引き起こします。生体 内では主にセレノシステイン(Sec)とよばれるアミノ酸に存在し、一部のタンパク 質(Se 含有タンパク質)に取り込まれます。Se 含有タンパク質は Se の高い反応性 を利用して、抗酸化作用など重要な機能を発揮します。 Sec は、タンパク質を構成する標準的な 20 種のアミノ酸に加えて、新たに発見さ れた「21 番目のアミノ酸」として知られています(図 1)。標準的なアミノ酸と同様 に、遺伝暗号に従ってタンパク質に取り込まれます。その際、アミノ酸をタンパク質 合成の場であるリボソーム※7に運搬するには、それぞれ専用のtRNA が必要で、Sec
には専用のtRNASecが存在します。通常のtRNA は、それぞれに対応するアミノ酸が
結合しますが、tRNASecは、まず一度、別のアミノ酸である「セリン(Ser)」(図 1)
が結合します(Ser-tRNASec)。その後、Ser が Sec へ変換され、Sec-tRNASecが合成
されて、リボソームに運搬されることでタンパク質に取り込まれます(図2)。 この変換メカニズムは、ヒトを含めた真核生物とアーキアのグループ(ヒト型)と バクテリアのグループ(バクテリア型)では全く異なります。ヒト型では、2 つの酵 素「PSTK」と「SepSecS」によって、2 段階で Ser から Sec へと変換されます。一 方、バクテリアでは1 つの酵素「SelA」によって、1 段階で Ser を Sec に変換します (図2)。2010 年の本研究グループの成果などにより、ヒト型のメカニズムの全容が 明らかになりました注)が、バクテリア型のメカニズムの研究は巨大タンパク質である SelA の結晶構造解析が技術的に困難であったため大きく遅れており、その解明が求 められていました。 注)2010 年 8 月 13 日プレスリリース: http://www.riken.jp/pr/press/2010/20100813/digest/ 2.研究手法と成果 共同研究グループは、バクテリアの一種 A. aeolicus(A.アエオリカス)由来の SelA 単体、および SelA と tRNASecの複合体の結晶を作製し、理研の大型放射光施設
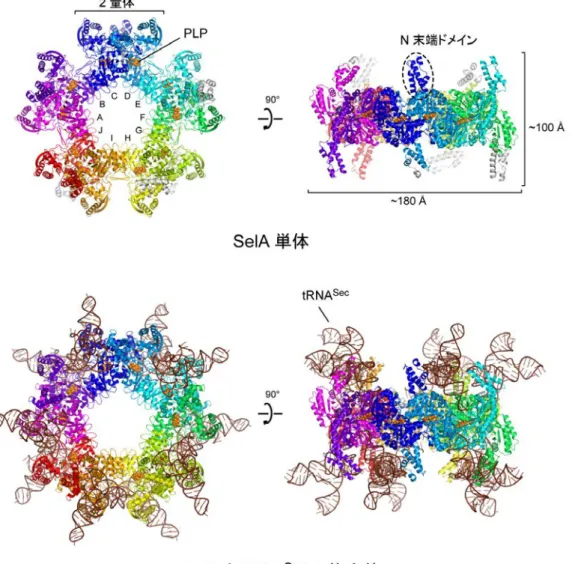
SPring-8※8のビームラインBL41XU と、高エネルギー加速器研究機構の放射光科学 研究施設フォトンファクトリー※9のビームラインBL5A、BL17A、NW12A を用いて 結晶構造を解析しました。その結果、SelA は、2 個のサブユニットからなる 2 量体 が5 個、星形に配列した 10 量体であることが分かりました(図 3 上)。全てのサブユ ニットは互いに同じ構造であるため正5 角形型の対称性を持ちます。その分子量は通 常のタンパク質が数万であるのに対し、SelA の総分子量は 50 万を超えます。そして、 SelA と tRNASecの複合体では、合計10 個の tRNASecがSelA に結合しており、総分
子量 81 万の超巨大タンパク質-RNA 複合体を形成していました(図 3 下、動画: http://youtu.be/v4a55kmbXL8)。この大きさは、全ての細胞に存在するタンパク質 合成の場であるリボソームの30S 粒子にも匹敵します。
SelA と共通の先祖を持つ他の酵素は、2 量体か 4 量体で機能します。一方、SelA は 10 量体で、このような巨大な構造は他に例がなく、なぜ超巨大な複合体が必要な
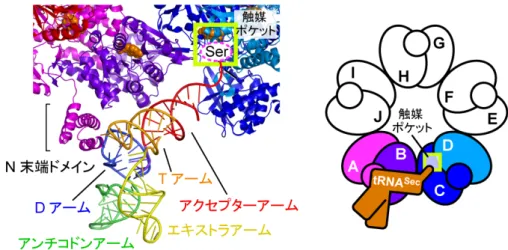
のか、その詳細な機能について調べました。その結果、SelA の中で、隣り合う 2 個 の 2 量体に含まれる 4 個のサブユニット(図 4 右 A~D)は、1 つの Ser-tRNASecに
対し、協力して4 つの異なる作業を担うことが分かりました。具体的には、サブユニ ットA が①「Ser-tRNASecを識別し」、サブユニットA と B が②「Ser-tRNASecを固
定し」、サブユニット C が③「Ser-tRNASecの先端を捕まえ」、サブユニット C と D が④「その先端にある Ser を Sec へと変換する」、という連続した作業により、1 段 階で Ser を Sec に変換すると分かりました(図 4)。また、サブユニットは全て同じ 構造であるため、隣の Ser-tRNASecに対しては、C~F の 4 個のサブユニットが 4 つ の作業を担い、サブユニットC に着目すれば、このときには①と②を担当しています。 このように各サブユニットは4 つの作業を全て担うことが出来ます。これらのサブユ ニットをそれぞれのtRNASecに対して機能させるためには、2 量体の配置が重要であ り、これを実現するためにバクテリアでは、超巨大な正5 角形型の星形構造を産み出 したことが分かりました。さらに、環状に5 つの 2 量体を配置することで、全体では、 ①~④の作業が 10 カ所で可能です。もし、直線状に配置した場合は、両端に無駄が できるため、全体で8 カ所だけとなり非効率的です。このように環状であることの重 要性も判明しました(図4)。 また、Ser から Sec への変換は、反応性に富む Se を組み込む困難な反応であると ともに、tRNASerなど他のtRNA に Se を導入しないよう、正確に識別する必要があ
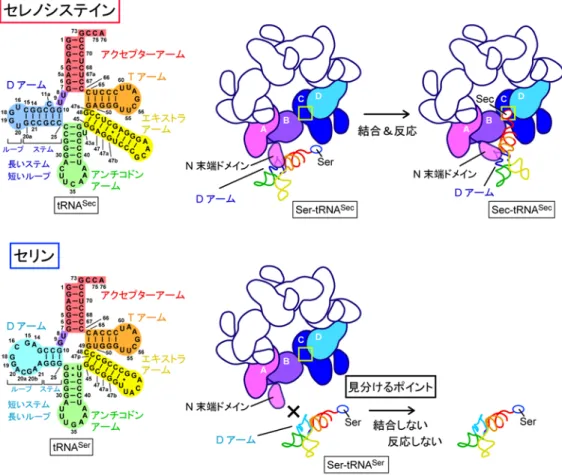
ります。そのメカニズムを詳細に調べたところ、SelA は、星型の構造から突出した 領域(N 末端ドメイン:図 3 上右、図 4 左)が、tRNASecが持つ固有の D アームと
結合することで、tRNASecを正確に識別していると分かりました(図5)。
さらに、ヒト型で働くSepSecS とバクテリア型で働く SelA では、Ser から Sec へ の変換を触媒する部位の構造が全く異なることも判明しました。Sec を合成するこれ らの酵素は、互いの構造も反応メカニズムも異なります。つまり、ヒト型とバクテリ ア型の酵素は、別々の先祖から、それぞれ独立にSec を合成できるように進化(収れ ん進化)したという非常に興味深いことも分かりました。 3.今後の期待 Se 含有タンパク質は、ヒトの生存や健康の維持に必須で、その研究は大変重要で す。しかし現状では、Sec を自在にタンパク質へ取り込むことができないため、人工 的な合成は困難です。今回、ヒト型に続きバクテリア型の生体内のSec 合成メカニズ ムも解明できました。今後、人工的な Se 含有タンパク質合成方法の開発に大きく貢 献し、Se の自在な導入によって今まで出来なかった天然の酵素の機能を上回る能力 を持つスーパー酵素の創生や、Se 欠乏を原因とする疾患の研究などに役立つことが 期待できます。 アーキアとバクテリアの多くは、基本的な20 種類のアミノ酸のうちの幾つかにつ いても、Sec 合成のように tRNA 上で他のアミノ酸を経由して合成しています。これ は原始生物の名残とされ、初期の生物は少ない種類のアミノ酸からタンパク質を合成 し、進化の過程で新しいアミノ酸を獲得していったと考えられています。タンパク質 を構成するアミノ酸の並びはDNA 上の遺伝子に規定されているため、新規のアミノ 酸の獲得には遺伝暗号とその翻訳系の進化が必要です。Sec の翻訳系は最も歴史の浅 い未熟なものであるため、Sec の合成から組み込みまでのメカニズムを詳細に調べ、
完成されたアミノ酸の翻訳系と比較することは、原始の生物が遺伝暗号を進化させな がら現在の姿に至った経緯をひも解く手がかりになると期待できます。
原論文情報:
Yuzuru Itoh, Markus J. Bröcker, Shun-ichi Sekine, Gifty Hammond, Shiro Suetsugu, Dieter Söll, and Shigeyuki Yokoyama. “The decameric SelA•tRNASec ring structure reveals the mechanism of bacterial selenocysteine formation”. Science,2013
<報道担当・問い合わせ先> (問い合わせ先) 独立行政法人理化学研究所 生命分子システム基盤研究領域 (4 月 1 日より、上席研究員研究室 横山構造生物学研究室 上席研究員) 領域長 横山 茂之(よこやま しげゆき) TEL:045-503-9196 FAX:045-503-9195 (報道担当) 独立行政法人理化学研究所 広報室 報道担当 TEL:048-467-9272 FAX:048-462-4715
<補足説明>
※1 21 番目のアミノ酸 一般に、タンパク質は 20 種類のアミノ酸を鎖状に連結させたもの。アミノ酸配列は遺伝 暗号としてDNA の塩基配列により規定され、塩基配列→アミノ酸配列への変換を「翻訳」 と呼ぶ。アミノ酸の連結は、DNA の転写物である伝令 RNA(mRNA)によりリボソーム で行われる。mRNA の塩基は、A、U、G、C の 4 種類で、その連続する 3 つの塩基をコ ドンと呼ぶ。コドンは4×4×4 の計 64 種類で、その内 61 種が 20 種類のアミノ酸のいず れかに対応し(例:UCA コドンと AGU コドンはセリンに対応)、残りの 3 種類はアミノ 酸の連結を終結させる(終止コドン)。ヒトを含む多細胞動物や、植物、原生動物、アー キア、バクテリアの一部には、セレノシステイン(Sec)というアミノ酸があり、標準的 な20 種類に加えて新たに発見されたことから「21 番目のアミノ酸」と呼ばれる。Sec のコドン(UGA コドン)は終止コドンで、mRNA 上に特別な配列がある時だけ Sec のコド
ンとして働く。
※2 転移 RNA(tRNA)
転移リボ核酸(transfer RNA)の略号。RNA の一種で、A、U、G、C が 70~100 個鎖状
に連結したものである。アミノ酸ごとに1~数種類の tRNA が存在する。tRNA はそれぞ
れ対応するアミノ酸を末端に結合し、リボソームまで運搬する。リボソームにアミノ酸を
受け渡すと、再びアミノ酸を結合して、リボソームまで運搬する働きを繰り返す。tRNA
はアクセプターアーム、D アーム、アンチコドンアーム、エキストラアーム、T アームの
ら旋構造の領域(ステム)と、先端の二重ら旋を取らない領域(ループ)を持つ。アクセ プターアームの先端にはアミノ酸が結合し、アンチコドンアームの先端の3 塩基はアンチ コドンと呼ばれ、リボソーム内でmRNA のコドンと対合する。D アームと T アームが 3 次元的に相互作用してL 字型の立体構造を取り、L 字からエキストラアームが突き出す。 ※3 真核生物 細胞内にDNA を収納した核という小器官を持つ生物の一群。ヒトを含む多細胞動物、真 菌類(カビやキノコ)、植物、原生動物などが属する。 ※4 アーキア 古細菌とも呼ばれる微生物の一群。真核生物、バクテリアと共に生物を分類する3 つの大 分類群の一員である。核を持たない点で真核生物と異なるが、アミノ酸やタンパク質の合 成など基本的なシステムは真核生物と酷似しており、バクテリアよりも真核生物に近縁で ある。メタン菌、好塩菌、超好熱菌など極限環境に生息するものが多い。 ※5 バクテリア 細菌または真正細菌とも呼ばれる核を持たない微生物の一群。大腸菌、枯草菌(納豆菌)、 乳酸菌、サルモネラ菌などが属する。 ※6 サブユニット タンパク質は1 本のアミノ酸重合鎖(ポリペプチド鎖)から成るもの(単量体タンパク質) と、複数本の鎖から成るもの(多量体タンパク質)がある。多量体タンパク質を構成する 各々のポリペプチド鎖をサブユニットと呼ぶ。 ※7 リボソーム タンパク質を合成する細胞小器官。自身も巨大なRNA-タンパク質複合体である。リボソ
ームはmRNA を取り込み、mRNA のコドンと tRNA のアンチコドンを対合させることで、
アミノ酸を結合したtRNA を呼び込み、コドンの順番通りにアミノ酸を連結する。リボソ ームは大小2 つの粒子がダルマ状に重なった構造で、バクテリアの場合、大小の粒子はそ れぞれ50S 粒子と 30S 粒子と呼ばれ、真核生物の場合は 60S と 40S と呼ばれる。 ※8 大型放射光施設 SPring-8 理研が所有する兵庫県の播磨科学公園都市にある世界最高級の放射光を生み出す施設。そ の運転管理と利用者支援は高輝度光科学研究センター(JASRI)が行っている。SPring-8
の名前はSuper Photon ring-8 GeV に由来。放射光とは、電子を光とほぼ等しい速度まで
加速し、電磁石によって進行方向を曲げた時に発生する強力な電磁波のこと。SPring-8 では、この放射光を用いて基礎科学から産業利用までの幅広い研究が行われている。 ※9 放射光科学研究施設フォトンファクトリー 茨城県つくば市にある放射光科学研究施設。日本初のX 線を利用できる放射光専用光源と して1982 年に完成した。大学や研究機関が共同で利用実験するための施設(大学共同利 用機関)として高エネルギー加速器研究機構(KEK)によって運用されている。
図 1 セリン(Ser)、システイン(Cys)、セレノシステイン(Sec)の構造式
Sec は、Ser の酸素(O)または Cys の硫黄(S)がセレン(Se)に置き換わったアミノ酸で ある。
図 2 セレノシステイン(Sec)と標準的なアミノ酸の翻訳系
上:真核生物/アーキアの Sec の翻訳系
2 つの酵素「PSTK」と「SepSecS」によって、2 段階で Ser から Sec へと変換される。
まず、PSTK が Ser-tRNASecを識別し、Ser にリン酸基(P)を転移して目印とする。次
に、SepSecS が目印のある Ser だけを Sec に変換する。
中:バクテリアのSec の翻訳系
1 つの酵素「SelA」によって、1 段階で Ser を Sec に変換する。 下:標準的なアミノ酸の翻訳系
図 3 SelA 単体および SelA と tRNASecの複合体の全体構造 上:SelA 単体 星型の正面(左)と側面(右)から見た図で、N 末端ドメインが星型の板状構造から突 き出している。10 個のサブユニットが環状に配置した星型の構造で、10 個ある触媒ポケ ットには、酵素の働きを助ける補酵素の1 種「ピリドキサールリン酸(PLP)」が結合し ている。PLP はビタミン B6が体内で形を変えたもので、SelA の他、様々な酵素の活性 を担う。
下:SelA と tRNASecの複合体
星型の正面(左)と側面(右)から見た図。
合計10 個の tRNASecがSelA に結合している。
図 4 SelA と tRNASecの相互作用の拡大図と模式図
SelA のサブユニット A と B に結合した tRNASecを拡大した図。サブユニットA の N 末端ド
メインがtRNASecの特徴的なD アームと結合して tRNASecを識別している。4 つのサブユニ
ット(A~D)は、1 つの Ser-tRNASecに対し、協力して4 つの異なる作業を担う。サブユニ
ットA が①「Ser-tRNASecを識別し」、サブユニットA と B が②「Ser-tRNASecを固定し」、
サブユニットC が③「Ser-tRNASecの先端を捕まえ」、サブユニットC と D が④「その先端
にあるSer を Sec へと変換し」、1 段階で Ser を Sec に変換する。
なお、サブユニットは全て同じ構造であるため、隣のSer-tRNASecに対しては、サブユニッ
トC が①を、サブユニット C と D が②を、サブユニット E が③を、サブユニット E と F が
④を担う。また、向かい合うSer-tRNASecに対しては、サブユニットD が①を、サブユニッ
トD と C が②を、サブユニット B が③を、サブユニット B と A が④を担う、というように、
図 5 tRNASecと tRNASerの識別メカニズム
上: セレノシステインのtRNASecの識別メカニズム
SelA の N 末端ドメインは、tRNASecが固有に持つD アームの構造と結合し、tRNASec
上のSer を Sec に変換する。D アームはステムとループから成っており、tRNASecのD
アームはステムが長く、ループが短い特徴を持つ。
下:セリンのtRNASerの識別メカニズム
tRNASerを含む標準的なアミノ酸に対応するtRNA の D アームはステムが短く、ループ
が長い。このためtRNASerのD アームは SelA と結合できず、tRNASerに付いたSer が