東医大誌 78(3)
: 217
-223, 2020
ミニレビュー
生化学分野ハイライト
No. 1
がん細胞におけるリソソームの機能と治療標的と しての可能性
Lysosomal functions in cancer cell biology and their possibility as the therapeutic target for cancer treat- ment
東京医科大学生化学分野 : 宮﨑誠也、宮澤啓介 Department of Biochemistry,
Tokyo Medical University : Masaya MIYAZAKI, Keisuke MIYAZAWA
は じ め に
リソソームは、オートファジーによる選択的分解
(lysophagy)やオートリソソームからのリソソーム 再 生(autophagic lysosomal reformation : 以 下 ALR と略す)など、ダイナミックな細胞内動態を示す小 器官である。リソソームは長い間、エンドサイトー シス、ファゴサイトーシス、オートファジーによっ て輸送された細胞内外由来の高分子化合物を分解す
る “消化器官” として認知されてきた 1) 。しかし近年、
シグナル伝達やがんの転移、免疫応答、細胞膜修復、
小胞の開口分泌(exocytosis)など様々な機能を調 節する小器官として注目されている。さらに、リソ ソーム機能の欠陥はリソソーム蓄積症やアルツハイ マー病、パーキンソン病、筋萎縮性側索硬化症等の 代謝変性疾患病態への関与も明らかとなってい る 2) 。
今回、この多様な機能を有するリソソームのがん における細胞内挙動を中心に、細胞生物学的意義を 概説する。また、この細胞内小器官を「治療標的」
とするがんの新規治療法の可能性について、当生化 学分野の最近の知見を踏まえて紹介したい。
リソソームについて
リソソームは約 60 種類の加水分解酵素を含み、
これらの酵素は pH 4.5〜5 が至適条件とされている。
このリソソーム内の酸性環境は、リソソーム膜上の プロトンポンプである V-ATPase や塩素イオンチャ ネル、イオントランスポーターによって維持されて いる 3) 。リソソーム膜は約 200 種類の膜タンパク質 から構成されている。主要な膜タンパク質である lysosomal
-associated membrane protein ( 以 下 LAMP
と略す) 1 及び LAMP2 は、糖鎖含量が高く加水分
解酵素群からリソソーム膜自身を保護していると考 えられている。また、リソソーム膜上にはオートファ ゴソームや細胞膜とリソソームの融合に重要な役割 を 持 つ、SNARE (soluble N
-ethylmaleimide sensitive factor attachment protein receptor)タンパク質が存在 する 4) 5) 。
リソソームは細胞質全体に広く分布している。そ の 細 胞 内 局 在 に よ り、 核 周 辺 に お け る 中 心 体
(MTOC : microtubule organizing center)近傍に存在 する不動性の集団と、細胞膜に近い末梢側に存在し 移動能に富む集団の大きく 2 つに区分されている。
核周辺のリソソームは、オートファゴソームと融合 しオートリソソームを形成することで、その内容物 の分解に関わっている。一方、末梢に存在するリソ ソームは、細胞膜修復の制御やラパマイシン標的タ ン パ ク 質 複 合 体(mammalian target of rapamycin complex 1 : 以下 mTORC1 と略す)の活性化に関与 している 6
-8) 。セリン・スレオニンキナーゼである mTORC1 は、リソソーム膜上に Ragulator タンパク 質複合体を介して局在し、インスリン刺激やアミノ 酸により活性化されるが、その活性化は Rheb と Rag の 2 種類の低分子量 G タンパク質の制御下で 行われる。まず、インスリン刺激を受けて PI3K
( phosphoinositide 3-kinase )が活性化され、 AKT 活 性化を経て、GAP(GTPase activating protein)活性 が抑制されることで GTP 結合型 Rheb が増大する。
この GTP-Rheb が mTORC1 のキナーゼドメインに 直接結合することでその活性が上昇する。一方、ア
ミノ酸は Ragulator タンパク質複合体と結合してい
る Rag GTPase を介して mTORC1 を活性化させるが、
この活性化にはリソソーム膜上の V-ATPase が必要 とされる 9) 。活性化された mTORC1 は、翻訳調節を 司る S6 キナーゼや 4E
-BP1 などをリン酸化するこ とでタンパク質合成を促進させる。
このようにリソソームは、オートファジーによる 異化経路と mTORC1 活性化による同化経路の両調 節に関わっており、がん細胞の旺盛な細胞内代謝に おいて重要な役割を担っている 10) 。また、オートファ ジーは腫瘍の成長促進や化学療法の薬剤抵抗性にも 関与するとされている 11) 。現在、オートファジー阻 害に焦点を当てたがん治療の研究が数多く行われて
いるが、臨床試験の対象となる薬剤の多くは、オー トファジーフラックスの後半部を阻害するオートリ ソソーム機能の調節に関与するものがほとんどであ る 10) 12) 。
リソソームの細胞内局在を決定する因子
一般にリソソームの細胞内の動きは “stop-and
-
go” の断続的な動きを繰り返して、その局在が決定 される 6) 13) 。リソソームの細胞内局在化を決定する 因子として、微小管上を移動する ① キネシンと
② ダイニン/ダイナクチン、そしてアクチンフィラ
Fig. 1 リソソームの細胞内局在と核近傍におけるクラスター形成
A. リソソームの細胞内局在は、核周辺の中心体近傍に局在して比較的不動のリソソーム集団と、細胞の末梢に
散在し移動性に富む集団とに大別される。微小管の⊕端への移動(黒色の矢印)にはキネシン、⊖端の移動(赤 色の矢印)にはダイニン/ダイナクチンを媒介して輸送される。ミオシンはアクチンフィラメント上に沿って輸送 する。リソソームエキソサイトーシス (点線で囲まれた箇所)では、リソソーム膜上の
SNARE
タンパク質を介 して細胞膜とのリソソーム膜融合が引き起こされ、その内容物が細胞外に放出される。B. 非小細胞性肺癌細胞株 H226
細胞に、プロテアソーム阻害剤であるボルテゾミブ(5 nM)とオートファジー阻害活性を有するクラリスロマイシン(50 µM)を同時添加後、
24
時間培養した。プロテアソーム系とオートファ ジー・リソソーム系の二大タンパク質分解機構を同時に阻害することでアグリソームが形成されるが、その際に 核近傍にリソソームのクラスターが形成される。電子密度の高い小器官(矢印)は全てリソソームである。(文献33
より引用)メント上を移動する ③ ミオシンの 3 つのモーター タンパク質が知られている。微小管はチューブリン 二量体の重合・脱重合によって形成され、その伸長 には方向性がある。チューブリン二量体が付加する 側を⊕(プラス)端、解離する側を⊖(マイナス)
端と呼ぶ。プラス方向への移動(順行輸送)はキネ シンが、マイナス方向への移動(逆行輸送)はダイ ニン/ダイナクチンが媒介することによって細胞全 体へのリソソーム輸送が行われる。一方、アクチン フィラメントは細胞膜直下に多く存在し、細胞の形 状を決定する。ミオシンは、キネシンによって細胞 膜周辺に輸送されてきたリソソームを受け取り、エ キソサイトーシスのための細胞膜との融合を制御す る(Figure 1A) 6) 14) 。リソソームの動きはこれらモー タータンパク質の他に、小胞体(ER)やゴルジ体、
ペルオキシソーム等の細胞小器官との相互接触に よっても調節を受けている 6) 15) 。
リソソームの細胞内局在や運動性は複数の条件に よって制御される。例えば、オートファジーが亢進 している条件下では、大部分のリソソームはオート ファゴソームと融合し、オートリソソーム形成に動 員されている。これによるリソソームの不足を補う ために、リソソーム再生(ALR)が活発化するが、
この機構にもキネシンとダイニンが関わってい る 6) 。また、細胞質のアルカリ化はリソソームを核 周 辺 に 分 布 さ せ、 酸 性 化 で は 末 梢 に 分 散 さ せ る 16) 17) 。さらに、アグリソーム形成や飢餓条件下で は、核周辺部でのクラスター化が誘導され、不良タ ンパク質処理や異化のために動員されていると考え られる(Figure 1B) 2) 。
リソソーム膜の透過性の亢進
(
LMP : lysosome membrane permeabilization
)と リソソームストレス応答(
ELDR : endo
-lysosomal damage response
)リソソーム膜透過性亢進(以下 LMP と略す)の 特徴は、様々な原因によりリソソーム膜が損傷を受 け、リソソームの膨化が惹起され、その内容物が細 胞質に放出されることである。LMP を引き起こす 内因性物質として、活性酸素種(ROS)やアポトー シス促進因子 Bax が知られている 18) 。過度なストレ スを受けることで、時にリソソーム膜の破裂が引き 起こされる。これにより、リソソーム内容物の加水 分解酵素カテプシン B や D 等の漏出により、細胞
質の酸性化やリソソーム機能の低下、さらに重度な 場合はネクローシスや Bid や Bax の活性化により アポトーシスが誘導されることもある。このように LMP を起点とする細胞質へのカテプシン漏出を伴 う細胞死を、リソソーム 依存性細胞死(LDCD : lysosome-dependent cell death)と呼ぶ 19) 。
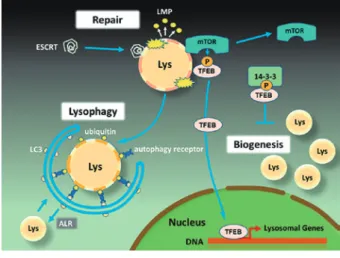
リソソームが受ける様々なストレスに対し、三種 類のリソソームストレス応答が惹起される(Figure 2) 18) 19) 。 一 つ 目 は、 リ ソ ソ ー ム 膜 損 傷 に よ り、
ESCRT タンパク質群がリソソーム膜の小さな穴に
動員され、膜修復が惹起される。二つ目は、リソソー ム膜修復による再生が不可能な状況において、リソ ファジー(lysophagy)により損傷したリソソーム の 分 解・ 排 除 が 行 わ れ る。 三 つ 目 の 応 答 は、
mTORC1
-TFEB 経路の活性化によりリソソーム生
合成が行われ、新規のリソソームが補充される。
転写因子 TFEB はリソソーム生合成やオートファ ゴソーム形成における “master regulator (主要制御因 子) ” であり、CLEAR と呼ばれる 10 塩基からなる 配列( 5 ′
-GTCACGTGAC
-3 ′ )に結合する。リソソー ム関連遺伝子の 96 個のうち 68 個がこの CLEAR を プロモーター領域に有し、TFEB による転写制御を
Fig. 2
リ ソ ソ ー ム ス ト レ ス 応 答(ELDR : endo-lysosomal
damage response)
種々のストレスによりリソソーム膜が障害される と ① ESCRTタンパク質が動員されて、膜修復が行 われる。また、② mTORC1のリソソーム膜からの 解離により
TFEB
の脱リン酸化が起こり、TFEBは 核内移行してリソソーム新生のための関連遺伝子の 転写活性が起こり、リソソーム数が増加する。また、③ ダメージを受けたリソソームはオートファジー による選択的分解(lysophagy)を受け、一部は新た なリソソームへと再生される(ALR : lysosomal ref-
ormation)。Lys :
リソソーム、LMP : lysosomal mem-brane permeabilization、TFEB : transcription factor EB、
mTOR : mammarian target of rapamycine
受けている。実際、TFEB を過剰発現させた細胞で は、リソソーム酵素やリソソーム膜タンパク質の遺 伝子発現が上昇し、リソソーム数も増大することが 明らかにされている 12) 。通常、TFEB はリソソーム 膜上に局在する mTORC1 によるリン酸化を受け、
14
-3
-3 タンパク質と結合することで細胞質に留まっ ている。しかし、リソソーム膜損傷に呼応したリソ ソーム膜からの mTORC1 の解離は、TFEB の脱リ ン酸化を促す。これにより TFEB が核内移行して、
新規のリソソーム生合成やオートファゴソーム形成 を誘導する(Figure 2)。
このような複数のストレス応答が作動することで リソソームの機能と数の安定性が維持されてい る 18) 19) 。
がんとリソソーム
クローン性増殖をきたすがん細胞では、正常細胞 に比べその旺盛な増殖により、高いエネルギー要求 を満たすためにオートファジー・リソソームへの依 存度が高い 20) 。特にリソソームは、細胞内エネルギー 代謝における異化の中核を担い、アミノ酸プールを 生み出すことでがんの生存・増殖に大きく寄与して いる 21
-23) 。このようながん細胞では、TFEB ファミ リータンパク質の MiT-TFE 遺伝子の過剰発現が見 られる 24) 25) 。この転写因子の高発現は、オートファ ジー誘導の亢進によるエネルギー源の確保と、Rag
GTPase を介した mTORC1 活性化によるタンパク質
合成の両経路を活性化する 1) 。これら異化と同化の 両経路の異常な活性化は、がん細胞の成長とエネル ギー代謝を大きくサポートすると考えられてい る 1) 26) 。
また、リソソームエキソサイトーシス(lysosomal
exocytosis )とは、リソソームが細胞膜と融合し、
リソソームの内容物を細胞外に分泌するシステムで ある。これによって加水分解酵素(カテプシン D, B, S, K, L)が放出され、基底膜成分を破壊することで、
がん細胞の転移や血管新生を促進する 12) 27) 。さらに、
がん細胞ではリソソームの一部機能不全も示されて いる 28) 。Eda M. らによる横紋筋肉腫細胞株を用い た実験では、糖鎖からシアル酸を遊離させるシアリ ダーゼの低下によって、シアル酸がリソソーム膜の
LAMP1 に蓄積することが観察された。これにより、
リソソームエキソサイトーシスが増強され、がん細 胞の転移能や浸潤能を高めることが示唆されてい
る 29) 。
Gotink K.J. らによる腎癌、結腸癌細胞株を用いた
分子標的薬スニチニブ(sunitinib)の耐性化に関す る報告では、スニチニブがリソソーム内に蓄積して 隔離されることで、薬剤抵抗性を獲得していること が示されている 12) 30) 。それに関連して、これまで我々 を含めた多くの研究者は、オートファジーの抑制に より各種抗がん剤の薬剤感受性が高まることを報告 している 11,31
-33)
発生母地やがん種により差があるものの、がん細 胞と正常細胞を比較すると、一般的にはがん細胞で は TFEB 関連遺伝子の転写活性が高く、それによる オートファジーの亢進、リソソーム数の増加傾向が 見られる 24) 25) 。また、リソソームの酵素活性が高く 維持されている傾向にあるが、一部のリソソーム に機能障害が見られる 28) 。このように、がん細胞に おいてリソソームの変化は、転移や浸潤そして薬剤 抵抗性など様々な役割を果たす。よって、リソソー ムを標的とするがん治療研究の重要性が今後さらに 高まっていくことが予想される。
リソソームを標的とするがん治療の可能性
mTOR は細胞内エネルギー代謝の中心的な調節因 子として機能し、TFEB の転写制御を司る。既に mTOR 阻害剤であるエベロリムス( everolimus )は、
腎癌、乳癌の症例で使用されている。また、クロロ キン(CQ)やヒドロキシクロロキン(HCQ)も “オー トファジー阻害剤” として、各種抗がん剤との併用 による臨床試験が進行中である 10) 。化学構造上で
CQ、HCQ の共通骨格であるキノリン環の二つをト
リアミンリンカーで繋げたビスアミノキノリン化合
物 Lys05 は極めて強力なオートファジー阻害剤とし
て開発された 34) 。さらに同様のビスアミノキノリン
化合物 DQ661 を用いた悪性黒色腫細胞株に対する
実験においては、異化経路であるオートファジーと 同化経路である mTORC1 を同時に遮断することで 抗腫瘍効果を発揮する事が報告されている 35) 。この 作用機序に関わる DQ661 の分子標的は、タンパク 質の脱パルミトイル化に必要な palmitoyl-protein thioesterase (PPT1)で、DQ661 はこの酵素活性を阻 害する。これにより、V
-ATPase サブユニットのリ ソソーム膜からの解離を促し、リソソーム機能阻害 によりオートファジーが抑制される。これと同時に、
Ragulator 複合体を破壊することで、リソソソーム
膜上への mTORC1 の局在化と mTORC1-Rheb 間の 分子会合も阻害するため mTORC1 活性が阻害され る。このように同薬剤は、がん細胞において、異化 経路と同化経路を同時に遮断することで抗腫瘍効果 を発揮する点で特徴がある 35) 。
当生化学分野では、アジスロマイシン(AZM)
を含むマクロライド抗生剤にはオートファジー阻害 効果があることを発見し、がん治療応用の可能性を 提言してきた 32) 33) 36) 37) 。これらマクロライドはオー トファジーの後半のプロセスを阻害することでオー トファジーフラックスを止める。Takeda A. らは、
AZM とプロトンポンプ阻害剤のランソプラゾール とを各種がん細胞株に同時に作用させると、細胞質 内にリソソーム自身を貪食したオートリソソームが 著しく増加し(lysophagy)、かつ、強力な殺細胞効 果が発揮されることを報告した(Figure 3A)。興味 深いことに、この殺細胞効果はアポトーシス誘導に 因るものではなく、 LMP を介したネクローシス様 の細胞死であった 37) 。この分子背景には、ランソプ ラゾールによるリソソーム膜障害で TFEB が活性化 し、リソソーム新生によりその数の増加をきたすこ とと、加えて、AZM のよるオートファジーフラッ クス阻害作用による lysophagy 自体の破綻が同時に 起こることに起因しているようである。この結果、
細胞質内に大量のダメージを受けたリソソームが蓄 積して、カテプシン等の加水分解酵素の大量放出に より、いわゆる LDCD が誘導されたと考えられる。
また、当分野の Hino H. らは、現在、進行・再発 乳癌治療に臨床使用されている CDK4/6 阻害剤アベ マシクリブ(abemaciclib)が、肺癌細胞株や乳癌細 胞株において、リソソームの著しい膨化を伴う強力 な非アポトーシス性の細胞死を誘導することを報告 した(Figure 3B) 38) 。このアベマシクリブの in vitro での殺細胞効果は、他の CDK4/6 阻害剤であるリボ シ ク リ ブ や パ ル ボ シ ク リ ブ よ り 強 力 で、 か つ、
CDK4/6 阻害剤とは独立した off
-target 効果による
“リソソーム膨化” に依存していた。これよりリソ
ソームの機能不全が起点となる新規の細胞死様式が 示唆されている 38) 。
ま と め
クローン性増殖を続けるがん組織では高分子化合 物の合成が必須である。リソソームは細胞内におい て、異化経路と同化経路のハブ的存在として機能し ており、がん細胞にとって極めて重要な小器官であ る。これを巧みに制御することで、有効かつ効率的 ながん細胞死を誘導することが可能である。しかし、
リソソームの役割は非常に複雑であり、がん細胞に おける細胞生物学的理解をさらに深めることは、難 治性腫瘍に対する新規治療法を確立する上で重要で ある。
謝 辞
電子顕微鏡撮影に協力いただだいた國場寛子先生
(東京医科大学 電子顕微鏡室)、貴重なアドバイス
Fig. 3
リソファジ―(lysophagy)の亢進および膨化したリソソームの増加により誘導されるがん細胞死の透過 型電子顕微鏡像
A. ランソプラゾール(100 µM)と AZM(50 µM)
で
24
時間処理後のA549
細胞(非小細胞性肺癌細胞 株)。オートリソソーム(破線領域)が細胞質に多数 観察され、内部には未消化の細胞内容物やリソソー ム自身(矢印)が自食されている(lysophagyの亢進)。(國場寛子氏撮影)。
B. A549
細胞をアべマシクリブ(10 µM)で12
時 間処理後。未消化内容部を含む膨化した空胞を多数 認め、これらはLAMP2
陽性であることから、リソ ソームであることが判明した(文献38
より引用)。右パネルは左ボックス内の拡大像。Lys : lysosome、
N : nucleus.
をいただいた東京医科大学生化学分野の高野直治講 師、論文作成に協力いただいた教室秘書の廣田綾子 さんに深謝いたします。
筆者の
COI
開示本論文発表内容に関しては特に申告なし。
文 献