九州大学学術情報リポジトリ
Kyushu University Institutional Repository
カイコにおけるビタミンB_2およびB_6の恒常性と生 理
中村, 匡利
https://doi.org/10.11501/3080191
出版情報:Kyushu University, 1994, 博士(農学), 論文博士 バージョン:
権利関係:
第11章
ビタミンB2の代謝とフラボキナーゼ
第I 章にお いて明らかにしたように, カイコにはビタ ミン
B
フ の補酵素 苅1)成分に関してそのレベルを僅端に低下させないような恒常性機構が作動している. リボフラピンを欠いた飼料による飼育を5齢期から開始し でも, その後の発育経j過i品2およびぴ、, 信常'日的':1札l
められなしいミ. 3齢から開始した場介にはかなりの影響が現れるものの, カ イコは成長し, 営繭・ 産卵することができた 飼料中のリボフラビン含
ー低下により幼虫休内のビタミンBゥが減少すると 貯蔵されてい たリボ
フラビンがFMNおよびFADに1伝換され, 補酵素型Bゥレベルの著しい 低下を防ぐものと推祭された. そして, マルピーギ管に見出される極め て大量のリボフラピンがこの機構に関与して いる可能性があ るものと考 えた.
と ころで, 一般に動物では摂取されたビタミンB2は, 補酵素型であっ
ても一旦消化管内で階素により加水分解を受けてリボフラビ ンとして吸 収され, 次いで吸収されたリボフラピンが各組織に運ばる. そこにお い て補酵素却の誘導体であるFMNと
FADとに
転換されるものと考えられ ている. リボフラビンから補酵素型の誘導体が合成される経路は, リボ フラピン →FMN→FADの順である. FMNとFADは独立した補酵素と して十幾能しており, ーん-が他方の代わりになることはない. したがって双ノjそれぞれに判7iT|ゾI�が似たれているものとィラえられる. 体|付の恒常'1ゾ1:
維持機構においてう まず必袋詰のや,ljMぷ引のB勺がリボフラピン から FMNとして合成され, さらにこれからFADが作られることは保夫であ る. 第I章に掲げた各紺.織の Bっ組成解析の結呆, 幼虫のマルピーギ管と 体液およびJ31�中においてはリボフラピンが多くヲ リボフラピンは貯蔵・
輸送型Bゥであると考えられることから ここではリボ、フラピン →FMN の段階が補酵素J盟 具合成経路の仲速となっているものと考えられる. こ の反応はリボフラピンキナーゼ[EC
2.7.l.26J
(以下フラボキナーゼと称 する)によるものであり, 本酵素はリボフラピン + ATP→ FMN + ADP の反応を触媒する フラボキナーゼは一
般に紺織および細胞がリボフラ ピンを一取り込むときにも役割を栄たしているとされており(Kas氾et
al.ラ1988)
, ビタミンBゥ代謝においては主災な昨素である.以上のような背示に基づいて, 本27tではまず高含量のリボフラビンを
含むマルピーギ管を選び, 同組織よりフラボキナーゼを部分精製し, 得 られた酵素分l面を用いて性質を調べた 次いで, マルピーギ管の次ぎに フラボキナーゼ活性の|匂いr
11
)J易からも同酵素を部分精製した. さらにリ ボフラピン無添加の剣料で飼育してビタミン B2 欠乏状態とした場合の 各組織におけるフラボキナーゼ活性の変化を追究し, ビタミンBつの代謝 調節について論じた- 51 -
材料とん-U;
供試蚕品極および飼育
カイ コの品柁としてはIJrj章に述べた3砲のうち支146 号 x
H
145号ま たは日140号×文145 �j-をJ-I�いた 品荷II'Uの結果の走兵は認められなか った. 対照およびリボフラピン欠之状態卜aにおける飼育条件は前章に記 したものと同ーである.フラボキナーゼ活性の測定万法
口l酵素活性の測定は既械(McCormick, 1971) のノゴ法に準じて行った 基本反応放としては,
0.2 Mトリスーりん限緩衝液(pH 8.5)中に1
rnMZnS04ヲ0.3 mMジチオスレイトール(DTT
), 1 mM ATP, 0.1 mMリボフラ
ピンおよび 酵素液を合むもの(全 量
0.4 ml)を用いた茶褐色の試験管中
で370Cヲ30分間反応させた後, 1 N Jr\r.):t昆素酸(PCA) 0.1 m1を加えて反応
を停止させ, 生じた沈肢を述心分離(1,600g,1 0分間)により除き, 上清 を前章に述べたノj法に准じてHPLCに供して生成したF MNの量を測定 し酵素活性(nmol/hrで点ポ)に換算した HPLCの分析時間を短縮する ために短いカラム(UnisilPack F3-50A , I.D.
4.6 mm, L.50 mm ジーエルサ イエンス)を用い,流速を卜o ml/minに変更した. ATPおよびリボフラピ ンに対するKI1l値またはpH の影響を訓べる際は上記の反応液組成にお いてATP, リボフラビンの濃度または緩衝液のpHを変化させた. 金属Jイオンの影響をみる ためには上記の濃皮のZnS04の代りに程々の濃度の
ZnS04またはMgS04を
加えた K m
他の計算のためにT aylor展開を併用
した故小山乗法を)IJいた(大l川,
197 8)
タンパク質の定歪
フラボキナーゼ、のJ51554における比Y�-YI:のttJ11には280および260 nm
における|政光度(A�l)(J,A26J測定イ111からド記の計算式(管原 ・r'f�U島ヲ1977)
を用いてタンパク75波皮を算J11した
タンパク質減Ji[
(mg/m1)
=1.45
X A28U -0.74
X A260クロマトグラフィーにおいてはA"8IJにおける吸光度をモニターし, タ ンパク質濃度の目安とした リボフラピン無添加の飼料で、飼育した際の 各組織におけるフラボキナーゼ、の比活性の算出には,フェノール試薬法 (Bensadoun and Weinstein の改良法)またはピュレット法を用いた(菅
・
副島,1977). 43!??:山手jiはウシ1(11.治アルブミンを用いて作製した.フラボキナーゼの部分精 製 万 法
幼虫から保取した組織を10倍量(v/w)の5
mM
2-メルカプトエタノー ル(2-ME)および1mMフッ化フェニルメチルスルホニル(PSMF)を必
む20mMりん酸カリウム緩衝液(pH 6.8)巾で磨砕し, 25,000gで30分 間遠心分離を行ってt清を回収しガーゼで憾過して粗抽出液とした. こ れに硫酸アンモニウムを加え40%飽和とした25,000g, 30分間の速心分 離を行い, その上清にさらに硫安を加え800/0飽和とし, 同様に遠心分離 した. Iこ清を捨て沈殿を5 mM 2-MEを含む20 mMトリス-IrL酸緩衝液(pH 8.3)に溶解し, 同緩衝液に対し充分透析した 溶液中に不溶物を生
じた場合には遠心分雌
(
30,000g
, 10分間)により こ れを除き透明な上清を- 53 -
得た. 第lのカラムクロマトグラフィ一段|年?としてQ-Sepharose fast f10w カラム(直径2.6 cm X長さ 52 cm)を川いた 流速は130m1/hrとし l
画分約] 5 m1ずつを[1 r[ 1[又した. 第2のカラムクロマトグラフィ一段階と してSephacry1S -1 OOHRカラム(1内:俺2.6 cm X長さ90 cm) を適用した
流速を24m]/hr, 1山i分約5mlずつを!日l収した.
分子量の推定
Superose 12 HR 1 0/30カラムを装着したFPLC システム(ファルマシア 社) を用いてフラボキナーゼの分イ-Uの推定を行った. 0.15 M KCl. 5
rnM 2-MEおよび0.10/0 ポリオキシエチレンソルビタンモノラウレート (Tween 20)を合む20 mMりん酸カリウム緩衝液(pH 6.8)を溶出液と
し, 流速は 0. 4ml/minとした. カラムに0.1 mlの試料を添加し, 溶出液 を0.1ml ずつ分阿した. 各分画のフラボキナーゼ活性を測定し, フラボ キナーゼの浴出位置を求めた. 分寸こ量マーカーに用いたタンパク質(シ グマ社, 指弧内は分子量)は, アプロチニン(6,500)ラ チトクロームC (12,400), カーボニツクアンヒドラーゼ(29,000), アルブミン( 66 ,000),
アルコー ル脱水素酵素(150,000 ) ,
ß
-アミラーゼ(200,000)である. これらの分析は2-MEを合まない緩衝液で、行い, A280 をモニターして溶 位置を求めた.
来llf 沢
マルビーギ管よりフラボキナー ゼの部分粘製
140 ザ×支145号の
5
r給幼虫から採取したマルピーギ管15 gを用い て粗酵素液をj=HUI',しヲイ流般アンモニウム分[Ùlj (40 '" 80%飽和)を待てラ 沈殿を5mM 2- ME
を合む20mMト
リス
ー店.酸緩衝液 (
pH8. 3) 14 mlに溶解し, 同緩衝液に対し充分透析し不溶物を述心分離で除い て透明な上清を 得た. 同緩衝液で千衡化したQ-Sepharose fast flowカラムに硫安分画を添 加し, 1Iサ緩衝液により浴川して40阿分を集 めた さらに溶出緩衝液を
0. 1
M KClおよび5mM 2-MEを合む20mMトリスー指酸緩衝液(pH7.0),1 M KClおよび5mM 2-MEを合む20mMトリスー刷酸緩衝液(pH 7.0)に 順次切り干与えることにより段階的溶山を行い , それぞれ80, 40画分を集
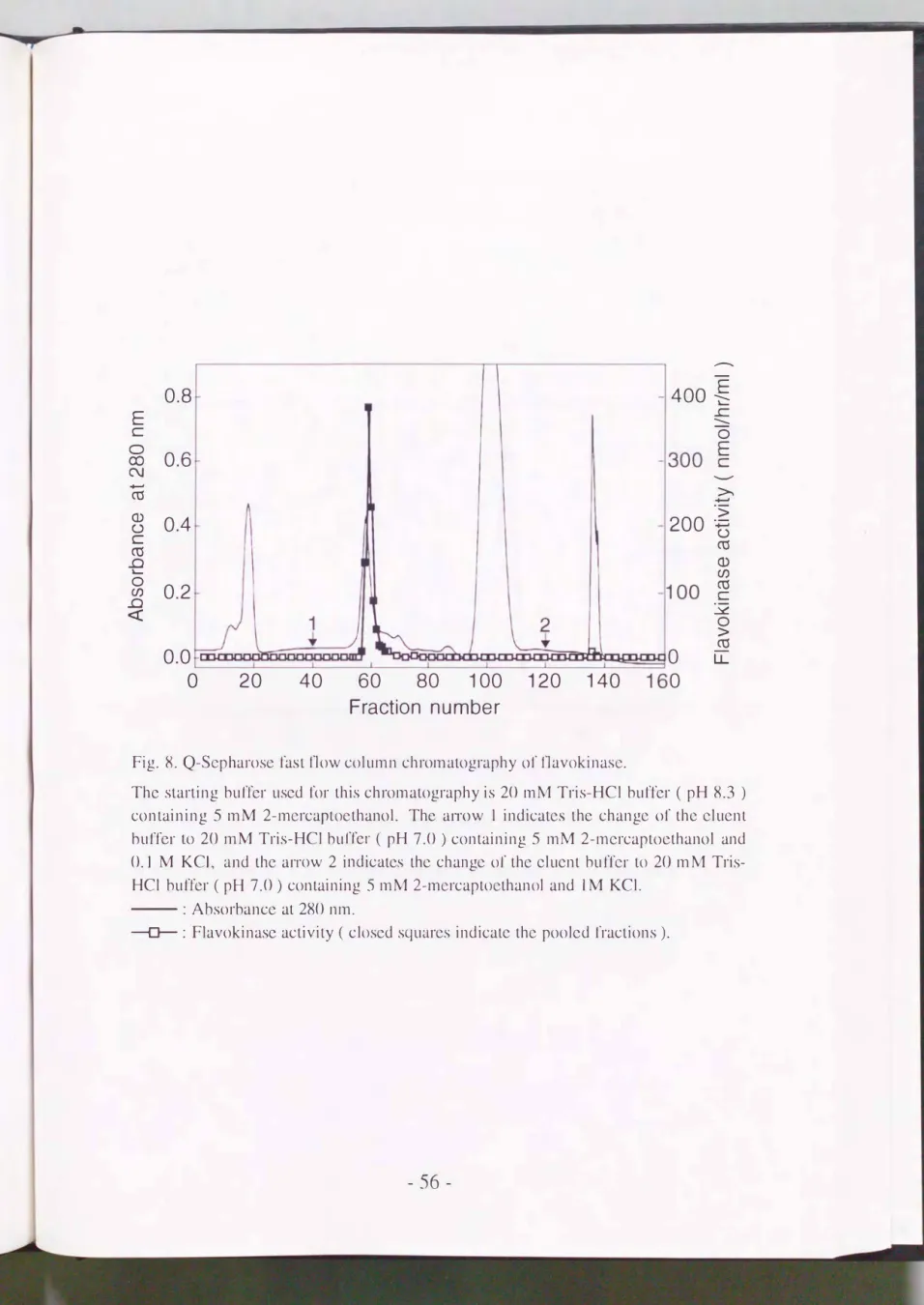
めた(Pig.8).
タンパク質は8 種類以上のピークに分離され フラボキナ ーゼ活性は0.1M KCl を合む緩衝液に切り替えた後, 最初に溶出されるタンパク官のピークにほぼ重なって浴J11した. 活性画分(第57'" 65番)を 集 め, 80%飽和になるように研し酸アンモニウムを加えた. 30,000g, 30分 間の遠心分離を行って沈殿を回収し, 4 mlの 5mM 2-MEおよび0. 10/0
Tween 20を含む20mMりん酸カリ ウム 緩衝液(pH 6.8)で溶解し, 不溶 物を遠心分雌 (12,000g, 30分間) により除い た この 上清を0. 15MKCl
5 mM 2-MEおよび0. 1% Tween 20を合む 20m M りん酸カリ ウム緩衝液 (pH 6.8)で、12衡化したSephacryl S-100HRカラムに添加し, ヒ記緩衝液で 溶I�-�した. タンパク質はFig.9に示す.ÌlÚり, 7ピークに分断tした. フラボ キナーゼ活性は最も低分子側の第7寄ピークの後半部分に溶出したのでヲ
- 55 -
。0CC心」Oω心〈
(一E\」工\一oεc)去一〉一ちのφωczxO〉の一比
nu
nu
nU
4
初 -1300
-1100 0.8
0.6
0.4
0.2
E
CO∞ω芯
ハUハU - 市
140 120
60 80 100 Fraction number 40
20 0.0
0
Fig. 8. Q-Sepharosc fast flow column chromatography 01' flavokinase.
The starting buf1'er used 1'or this chromatography is 20 mM Tris-HCl buffer ( pH 8.3 ) containing 5 mM 2-mercaptoethanoL The alTOW 1 indicates the changc 01' thc elucnt buf訂Ie口r tωo 20 luM Tris-HCl buf汀1-印Ieαr ( pH 7.0 ) cont凶aining 5 m恥M2ユ一mercaptωO巴t山hanol and 0.1 M KCI, a印nd t山.he aITOW 2 in(吋di比tじ;a叫i孔tcωS t山hc cぬha加ngc 0ωr the eluent bu1'此Iel・to20 mM Tris
HCl buf1'cr ( pH 7.0) containing 5 mM 2-mercaptoethanol and 1M KCl.
一一一一 : Absorbance at 280 nm.
-[トー: Flavokinase activity ( closed squarcs indicate the pooled fractions ).
(一E\」工\一O
EC)含一〉一ちのωωの豆半O〉の一比
600 500 400 300 200 100 0.6
0.5 0.4 0.3 0.2 0.1
εco∞ω芯ωOCの心」Oω(叫〈
。 90 80
30 40 50 60 70 Fraction number 10 20
0.0
。
Fig. 9. Sephacryl S-lOOHR column chromatography o[ f1avokinasc.
一一一一 : Absorbance at 280 nm.
--{正一: Flavokinase activity ( closcd squarcs indicate the pooled 1'ractions ).
- 57 -
これらの阿分(第61� 67 {fi=)を集め た. 本分!血iはクロマトグラフィーパ ターンか らみて均一な までに精製されていないことは明らかであるが
ビタミン B2 欠乏との刈-)J�\において昨ぷの性質を訓任するには充分であ ると思われたので, この段階で枯製操作を打ち切った. 各精製段階にお ける結呆をTable 16 にま とめる. ここまでの操作によって6 mg の標品が 得られ, 比活性は4,317nm ol/hr/ mg prote in, 精製度は450 倍, 回収率は 92%であった.
マルピーギ管フラボキナーゼ部分村製標品の性質
以上のようにして何られたマルピーギ??のフラボキナーゼ、標品につい て若干の基本的な酵素的性質を調べた まず, フラボキナーゼの基質で
あるリボフラピンの波皮を変えてf将来:1引っ:を測定したところヲ 基質j濃農度
8μM主U以J,、 卜.の倣I域2必kまで、j汀治「斤{rパげd↑v竹[1ゾ
系活性のそれぞれについて逆数フプ。ロツトをとると,
Fig. 10Bに示すよう
に直線関係が得られた よってこの酵素反応はMichaelis-Menten の式に あてはめられ, リボフラピンに対するKI1l他はl.4士0. 1μM と算出された.ラット)1 F臓のフラボキナーゼの場合リボフラピンへの Km は 1 ]μ M (Yamada et al. , 1990)であり, カイコの醇素はこれより l桁低い. フラボ キナーゼの もう一つの基質である ATPについても同様の実験を行った (Fig.]
1人B). Km値は 27
:t 3μMでありう ラット肝臓の酵素の3. 7μM (Yamada
etal. , 1990
) よりl桁両い.次 に, 分 子量の測定のため 部分粕製原品をSup erose 12 HR
10 /30カラ
ムによるゲル滅過にかけたところ, フラボキナーゼは14.3 ml のイ立置にTable 16.Summ訂y of pmi白cation of tlavokinasc from the Malpighian tubules.
Step To凶1 protein Total activity Specific activity Yield Pmitïcation (mg) (nmol/hl・) (nmollh/mg protein) (%)
u、
\。 Clude extract 2,911 27,800 9.6 100.0 1.00
(NH_J2S0.j fractionation 745 24,200 32.5 87.1 3.39
Q-Sepharose fast t10w 104 19,500 186.8 70.1 14.46
Sephacry S-lOOHR 6 25,684 4,317 92.4 449.7
3000
-、、
C 4O Lq・-4 J
2000
o_
0>
ε
エ、 、二、
E
o1000
C
、、-〆
〉
。
。 2 3 4 5 6 7 8
Riboflavin (μM) 0.0015
B
0.0010
〉\?
0.0005
0.0000
Km=1.4iO.1μM
。
1 / [riboflavin]
2
Fig. 10. Lincweaver-Burlくplots of tlavokinase [01・1ibollavin.
A: s - v P 1 0 ts.
B : Linewcaver-Burk p]ots from A.
A
-、
20000
c 4qO
L --d 〉
15000
0...
。〉
E
、工、二、
10000
o
E
C
5000
、『圃J
〉
。
。 50 100 150
ATP(μM) 0.00020
B
0.00015
ど0.00010
r-
Km=27:t3μM
0.00005
0.00000
・0.05 0.00 0.05
1 / [ ATP ]
Fig. 11. Lincwcavcr-Burk plOL� 01' ílavokinase for ATP.
A: s -v plo ts.
B : Lincweaver-Burk plots from A.
- 6] -
200
0.10
浴J11された. これを分子泣日正夫11の マーカータンパク12から求めた検量線 にあて はめると, フラボキナーゼの分r.祉は18,000であると推定された
(Pig. 12).
この分f-Jltがサブユニットのものであるか再か不明であるが,ラットのJJT臓や小腸のフラボキナーゼがそれぞれ分子量28ヲ000, 13,500 の単景休である(Merri l1& McCormic, 1980, Kasai et al., 1990) のでおそら
くカイコの本醇京もlfí_長体で存在しているものと推察される.
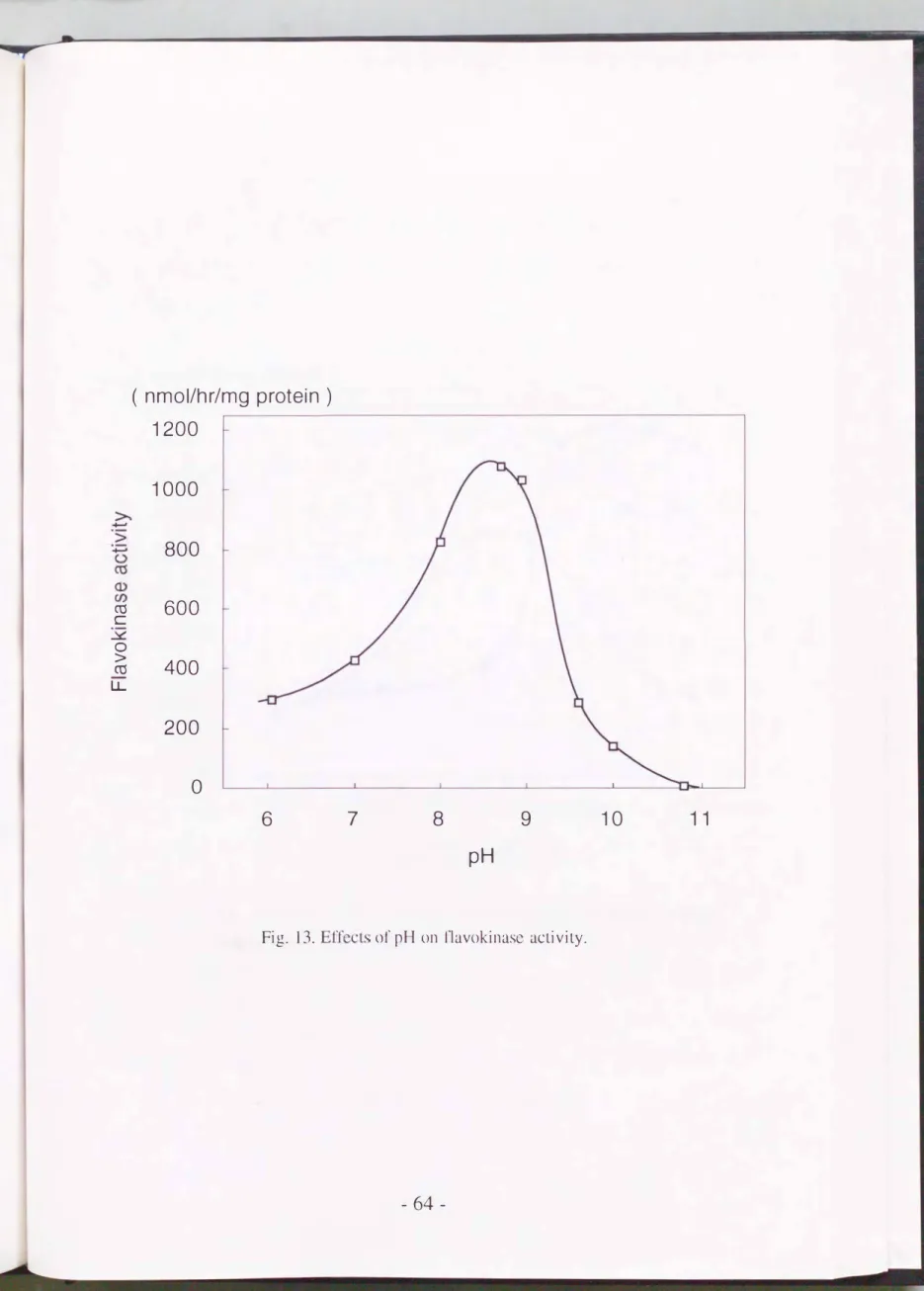
反応液のpH を6から約1 1まで変化させて酵素活性を測定すると,
Pig.
13に示す通りのゑ!?呆となった.
これよりフラボキナーゼの至適pH は8.5付近であると推定される. このイlf1はラット肝臓や小腸の同酵素の至適pHが 9.3ラ9.0である(Merril l
&McCormic, 1980, Kasai
etal., 1990)
のと比較するとやや低いが, いずれもアルカリ性11�IJにで迫pHを持つこ とは同じである.
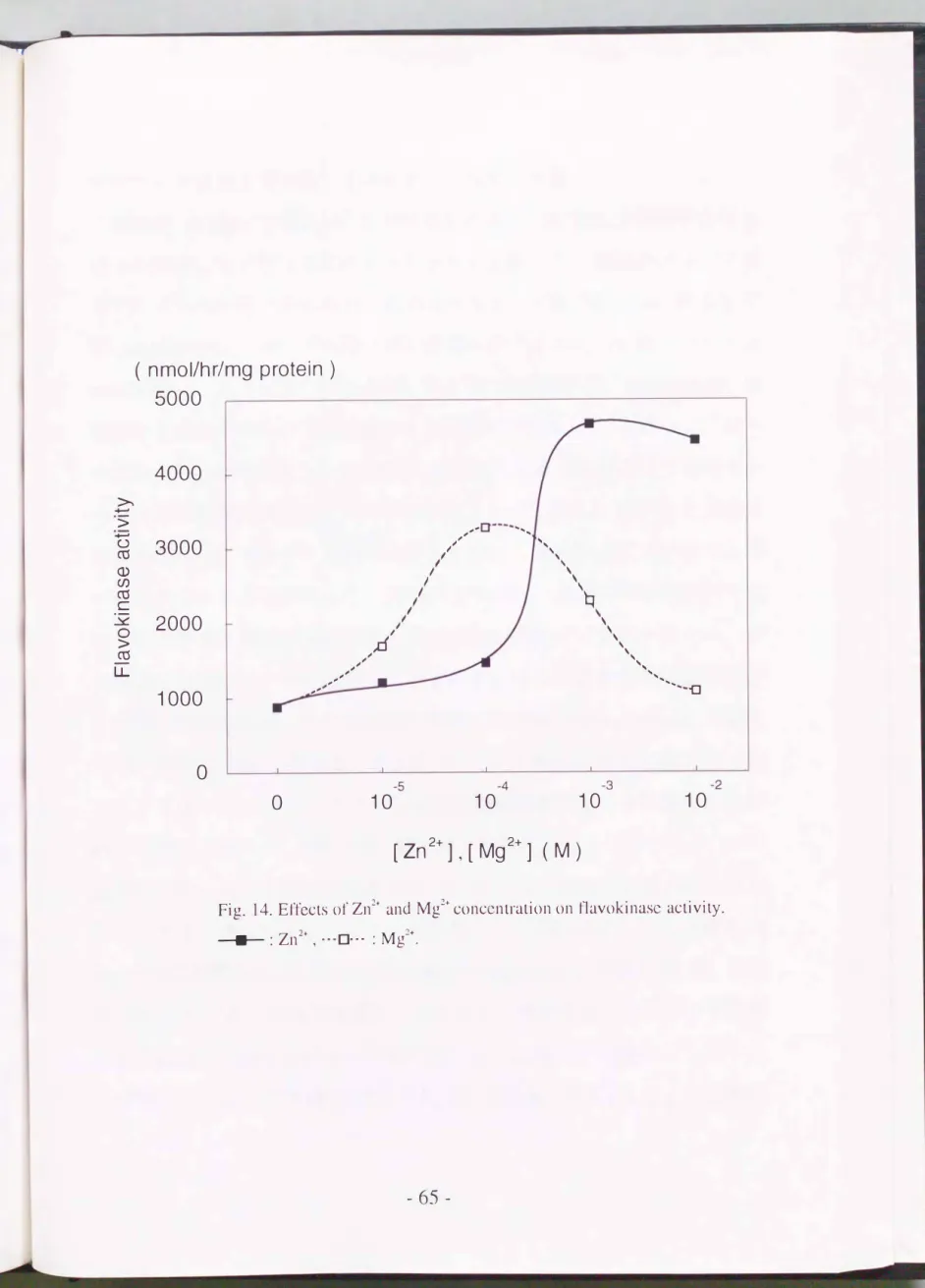
最後にd凶作イオンとマグネシウムイオンの影響を調べる目的で, 反Jじ、
液の ZnS04また はMgS04の波度を0から 10-� Mまで変化させて活性を 測定した(Fig.
14).
いずれのイオンによっても活性は上昇し , それぞれ 京高の前作1:を示す添加11せがI�jらかとなった すなわちZnS04ではlOJM,
MgS04では10-4
Mであり, こ の濃度のZn2+によって最大の活性が得られた. 本階京は活性の発現にて価重令属を安ぶすること マグネシウウム よりも亜鉛が効果的であることが切らかとなり, これら はラット肝臓や 小腸の酵素で報告されたことと一致する (Merri 11 & McCormic,
1980,
Kasai
etal.ヲ1990).
な お, 本実験における標准の活性測定条件は至適pH, L医鉛の至迫濃度,
某管(特 にリボフラピン)飽和の 条件に設定されている.
-、
ε
、、ー〆
Q) ε
コ
。>
c
。 4コ__,
μ」
17 16 15 14 13 12 1 Î
10 Î 8,000
5 10 20 50 100 200
Molecular weight ( kDa )
Fig. 12. Estimation 01' molccular wcight of flavokinase by gel tïltration.
The following mokcular weight markers wcre used. A: apl・otinin (6,500), B:
cytocromc C (12,400), C: carhonic anhydrasc (29,000), 0: albumin (66,000),
E: alchol dehydrogenasc (150,000), F: ß -amylase (200,000)
- 63 -
( nmol/hr/mg protein )
1200
1000
4〉〉- d、 4cに-u3
d
800cqn 3 c cu 600
ムζ
。>
の 400
LL
200
。
6 7 8 9 10 1 1
H nド
Fig. 13. Effeじts 01' pH on llavokinasc aじtivity.
(
nmol/hげmg protein)
5000
3000
、、、 ‘
ロ, , ,F ,, d' dF ,F 4,, d,, a'' ,,, dF ,,V 4,v d, ,, d d' ,, a' a, d, d' , , , , ,
旬、、 、 、、 、、 、、 、、 、、 、、 ‘、 ‘、 ‘、‘ ‘、 .• 』U「‘、、
‘、司、、 ‘、 、、 、、 、、 、、
4000
hv 一〉
日ocφωcc一V40〉の一比2000
1000
。
。 10 10 10 10
[ Zn 2+ ] , [ M g 2+] (M)
Fig. 14. Effects 01' Zn:!+ and孔192+ conccntration on t1avokinase activity.
一→・-: Zn2+, ---0・・・:
MI.
- 65四
マルピーギ管および'11脳フラボキナーゼ日竹の比較
標準の20μg/gリボフラピンで飼育したカイコ幼虫の5齢別における 種々の系11織についてフラボキナーゼ活JI�tを比較した 組織のタンパク質
、Iノたりでみた活性が肢も尚かったのはマルピーギ管であり(5齢4 r rで 30 nmo1/hr/mg), 次いで'11腸にltjい活性が検出された(5齢4 rrで16
nmo1Ihr/mg) .
これに比べてやや低いながら, 後部絹糸)J札 仁11部絹糸)J泉, 脂 肪体にもi1í'lil:
がみられた(詳細は次のJJll -r
で述べ る)
• したがって, 以下 の実験, すなわちピタミンB勺欠乏条件卜-における、当該組織のフラボキナー
ゼ、活竹の動態を
調べる た
めの予備段 階として
, 対照およびBっ欠
乏 幼虫の中腸からフラボキナーゼの部分村製を行い, その性質についてマルピ
ーギ管のものと比較十余討した. 村製の店法には 対照の中腸ではマルピ ーギ管のM素と同様にイ抗女分凶l, 12イオン交換クロマトグラフィー ゲ ル鴻過を迎)lJした その結,*, 約77 gの5齢幼虫中腸から70mgの部分 精製標品が伴られた そのものの比間性は270nmol/hr/mg protein, 精製皮 21倍, I叶収率420/0であった. さらに, ビタミンBゥ欠乏状態にあ る カイコ
として3齢起蚕よりリボフラピン無添加飼料で飼育し, 5齢幼虫の中腸 43 gを精製にr8いた.イ抗女分凶,陰イオン交換クロマトグラフィー, ゲル 滞、過を仁記の場合と同様の条件で行ったが, この場合さらにハイドロキ シアパタイトクロマトグラフィーをよ8}JIIした(詳細は省く) . この結栄, 3 mgの棋品が伴られヲ 比活ド1:は850nmoI/hr/mg protein, 粘製皮20倍, 凶収 蕊3%であった. これら'i1腸のフラボキナーゼの性質をマルピーギ管か ら 部分精製した酵素の場介と同様に調べた Table 17 にみられるように
リボフラピンおよびATPに刈-するKm仙, 分子量, ゴミ適pHおよび二イrlG金
、。j\
Table 17. Comparison 01' propelties of t1avokinases from thc Ma1pighian tubules and the midgut.
Tissue KlIl va1ue (μM)
Ma1pighian tubules Midgut
Midgut (( -)B2-III) 1)
Ribol1avin
1.41:tO.11 1. 53士0.15 1.12 :t 0.04 Each K'I1 value represenLc;; the mean:t S. D.
ATP
26.6 :t 2.5 25.5:t 3.9 39.4:t5.3
Molccular Optimum Zn�+
Optimum pH concentratlOn wcight
(mM)
18,000 8.5 1.0
18,000 8.5 1.0
18.000 8.3 1.0
1) Midgut of ù1e s出cworm fed on riboì1avin-deficient diet from the bcginning of thc 31・d instar.
高イオンの彪響において, マルピーギ色二と2仙のrl' )J誌のMぷ出品川にjC は認められず, 3�のフラボキナーゼはI
LÎJ
-である ものと判断した.5齢期r!'のrl� )助フラボキナーゼ活性の変動
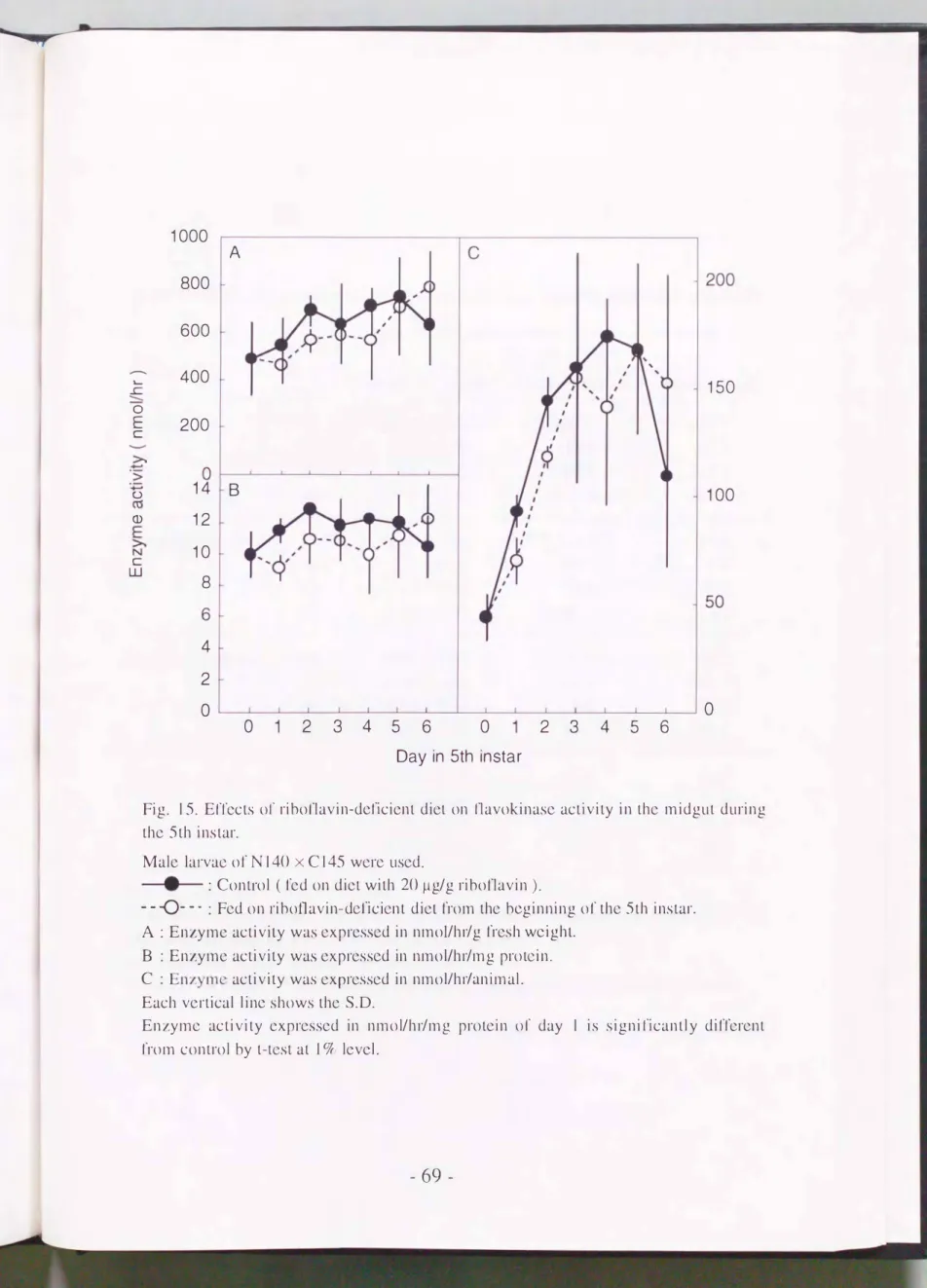
=þJJ訪のフラボキナーゼ活性は600 nmol/hr危fresh weight (Fig. 15A,実 線), 11 nmol/hr/protein (Fig. 15B,夫総)桂皮であり, 5齢朋問仁1-'大きな変 動はみられなかった. それゆえ, 組織における全活性は組織主に比例し て変動した(Fig.15C,夫総). 5齢J�蛍よりリボフラピン無添加飼料で飼 育しでも通常の飼育条件によるカイコとのI f'r]に差異は認められなか っ た
(Fig. 15A, B, C, 点線)
飼料料,のB勺令合「:UlJ止:f:を変えた条イ十刊4午1:.卜.での各系制組Il織のフラボキナ一ゼゼ、活v
マルビ一ギギ、t符i百ミ守.と|川l' )J版j助必のフラボキナ一ゼゼ、が|刊司一であるとのあ対結iケÎfl品論論自命i命)に準じ,
他の組組.織のフラボキナ一ゼも同じ分二子子であると仮定して次の実!験段を行行.つ た. まず, 3ih令起蚕よりÊftl�GI.�こ添加するリボフラピン電を変化させて飼育 を行い,
5
齢3日
のHtl.11で q,
腸, 脂肪体, 11'部および後六日絹糸)J泉のフラボキナーゼ日性を測定した(Table
18).
標準のリボフラピン添加1量である20μ
g/gの場合\ 中腸の市YJ�はほぼ]0 nmoI/hr/proteinであり, 他の来日織に比べ高い伯で、あった
中部紺糸腺の活性がI
1-'腸の次に高く, 次いで後部 絹糸JJ氏脂肪休の)11兵であった リボフラピンの添加l量を標準の1/2の10
μg/gまたは2佑-の40μg/gとしてもこれらの組織のフラボキナーゼ前性 は変化しなかった. しかし, リボフラビンの添加量を0μg/g, すなわちリ ボフラピン無添加とすると, '1')]易の フラボキナーゼ市竹1000
A
IC
800
�
ん600
'- 400
I 1/71",1/1\ Y � 150
工二、、、
。E 200
C
句-
喝〉
〉
-d
、
4c
に:_) 14
r
Bd " �
100。 12
ε 〉
N、 10
は」C
8
11'
4 2
。 。
。 2 3 4 5 6 。 2 3 4 5 6 Day in 5th instar
Fig. 15. EITccts 01' ribonavin-deficient diet on l1avokinase activity in thc midgut during thc 5th instar.
Male lar.vae of N140 x C145 wcrc uscd.
e--
: Control ( fed on dict with 20μg/g ribot1avin ).ー ー
-0
- ・・ : Fed on 1・iboí1avin-de1ïcient dict from thc beginning of the 5th instar.A : Enzymc activity was cxpres犯d in nmol/hr/g fresh wcight.
B : Enzymc activity was exprcssed in nmol/hr/mg protcin.
C : Enzyme activity was cxprcsscd in nmoVhl九nimal.
Each vcrtical line shows thc S.O.
En乙ymc activity cxprcsscd in nmol/hr/mg protcin 01' day 1 is signilïcantly diff、crcnt from control by t-tcst at 1弘lcvcl.
申69
-
Tablc 18. Ef1'ccts of graded Icvels 01' dictary rihoi1avin on t1avokinase activity in larval tIssucs.
Riboflavin Enχymc activity
Tissuc addcd to d ict
(μg/g) (nmol/Ilr/mg protcin) (nmoVhr/g fresh wt.) (nmol/hr/animal)
Midgut o 25.84 =t 2.22 1,059�.0 280.7
10 8.71 =t 2.59 340.6 92.0
20 9.67 =t 1.09 389.4 112.5
40 9.27 =t 1.44 321.9 107.2
Fat body 。 3.80 =t 1.07 145.3 16.4
10 2.07 =t 0.18 50.5 9.9
20 2.05 =t 0.34 70.4 13.0
40 2.07 =t 0.47 61.3 13.3
Middlc silk glands 。 10.00 =t 2.02 88.6 37.8
10 7.50 =t 3.0] 72.4 41.7
20 6.02 =t 0.44 68.1 42.5
40 5.85士1.18 69.1 37.1
Postcri01・silk gland只 。 5.03士0.75 101.1 38.2
10 5.72士0.38 112.7 56.6
20 4.61士0.78 106.2 54.4
40 4.69 =t 0.64 109.3 57.5
Male larva 01' C146 x N145 were uscd.
These were fed on the diet with gradcd levcls 01' libol1avin from thc beginning of thc 31・d instar and sacrificed for analysis on day 3 of thc 5th instal・-
Each specific activity reprcsents thc mean =t S.D.
* Signitïcantly diffrcnt from larvae fed dict with 20μg/g liboi1avin by t-test at 1 % lcvcJ.
昇した. 脂肪体と111却系ij糸WJ�ではわずかながら1!íltの卜.叫がみられたが,
「ド腸におけるほど著しくはなかった. 後1部、守1 祁干引 刊|日;系制紺|同j糸)服jUM山jA泉jAiのj汀fì了乃 ?引ÎIパ ,↑性|
無添加としてもほとんど変化しなかつた.
さらに, 3,4, 5齢の各Jt目玉三からリボフラピン無添力IID6J科で飼育した場 合の5齢4 lfにおける1
11 )
J持(
およびマルピーギ管
)のフラボキナーゼ活
性を比較したゑ!?米をTable19に剥げる 刈-
J!(tの創育条件の場合,組織当 たりの全店十tで、はr1-' )J易が此も大きなイ|立をイ守しており これは食餌Ijlのリ ボ、フラピンを吸収するために作動する酵素が含まれているためであろう.マ ルピ
ー
ギ管にはタンパク貨当たりおよび組織重当たりでみたフラボキ ナーゼ、間性が中腸のほぼ2倍近くも存在し 今回フラボキナーゼ活性を 測定した 5つの組織のうち肢もl匂いイ[11(で、あったが 組織が小さいため全 活性は111 )J易より低くなっていた リボフラピン無添加飼料への切換えを 5齢起蚕から行った場合には, 中腸とマルピーギ管双ノJにおいて, 対照と の聞にフラボキナーゼ活'I',tに差異は認められなかった(Pig. 15
の結呆と致する)• しかし
4
齢または3
齢 起 蚕から開 始すると斗i腸の活性は2 倍以上に上封した. け11寺 にマ ルピーギf苦の活性も 上井がみられたが, そ の上昇は30%程度にとどまっていた.考 察
E先述のようにカイコ幼虫においてはビタミン Bっ に闘して'恒常性をイ呆 つ機構が働いていると忠われる. この機備には合成, 分jýJ�, 愉送,結介|天
- 71 -
Tablc.19 E1'1'cじLs 01' ribona vin-dcfiじicnt dict on llavokinasc activity in thc midgut and the Malpighian tubules.
Tissue Dict Enzyme activity
(nmol/hr/mg protcin) (nmol/hr/g frcsh wt.) (nmol/hrhmimal)
Midgut Contl・01 16.11 :t 1.59 734:t 45 158.3士13.6 (一)B2-V 1) 15.76:t 0.96 725:t 47 170.0:t 7.2
(_ )B2-IV 2) 37.60:t 5.74 2,002:t 398 475.9:t 90.7
( - )B2-III 3) 46.80士7.41 2,412:t 480 602.8:t 71.7
孔1alpighian tubulcs Control 29.99:t 5.73 1,235:t 137 19.5:t 2.2 (-)B2-V 1) 29.26:t 4.28 1,244:t 167 24.0:t 3.2 (_ )B2-IV 2) 41.00:t 4.64 1,609:t 268 30.2:t 5.0 (一)B2-III 3) 3�L30 :t 5.71 L917:t 111 41.2:t2.4
Male larvac 01" N140 x C145 were used. Activity was assayed on day 4 01' thc 5th instar.
Each valuc represents thc mcan:t S.O.
1) Larvac wcrc fcd on 1・ihotlavin-deCicient diet from thc beginning 01' the 5th instar.
2) Larvae werc 1'cd on ribol1avin-deiïcicnt dict 1'rom thc beginning 01' the 4th instar.
3) Larvae were fcd on ribonavin-dcfiじient diet from the beginning of thc 3rd instar.
* Significantly diffrcnt 1'rom larvae fed dict with 20μg/g ribonavin by t-tcst at 1 % levcI.
子などいくつかの安|大|が凶与する 本州先においては合成過紅の訓節と いう観点からリボフラピンを基質としてFMNを形成する酵素, フラボ キナーゼに焦点をあてた. この醇ぷは刑l織および細)泡がリボフラビンを 取り込む|祭にも機能するという点でも主民である.
組織mと�たりまたは組織タンパクfT当たりでみた場合 フラボキナー ゼ活性はマルビーギ管に最も強く認められたことより マルピーギ管に 大量のリボフラピンが蓄積する際に本階京が機能しているも のと考えら れる. 次に組織当たりの全活性でみた場合には, 中腸が他の組織よりも はるかに尚いフラボキナーゼ活性をイJしていた. 食物から摂取したビタ ミンB2はリボフラピンとして中協より吸収され, さらにこれが体液を経 由して他の組織に辺ばれるはずである. すなわちけl腸はカイコの生体内 で、使われるビタミンBゥのすべてを消化管内より吸収するのであり, この ためフラボキナーゼ、活性が高いものと忠われる. このようにカイコにお けるフラボキナーゼ治性の分布はビタミン Bゥ の代謝を反映しているも のと解釈しうる.
カイコでは通常, 体液およびマルピーギ管にリボフラピンを蓄積して おり, ビタミンB2欠乏状態になるとこれが減少することから, この蓄積 刑のリボ、フラビンがフラボキナーゼのj志賀として動員できるプールを形 成しているならば, 体内における補醇コ表明ビタミン
B
勺 の恒常性維持に役 jJ_っているものと考えられる. これに関連してう マルピーギ管のリボフ ラビンが結合型でなく遊離型であるとする指J向(fl-;札1956b) は竜安で ある. 11出乳類で、はリボフラピンは特異的結合タンパク質(RFBP)と複合 体を形成しているときは, リボフラビンはフラボキナーゼの基質となり- 73 -
英fiくネ111
M ir;
Jl�!に転換できないとされる. さらに, 木市における実験の結 果, 若い齢からビタミンBゥJ!t'�添加でflld í守した場合の5齢別のように体内 のB:2欠之ががかなり進んだ条件卜においては, IIJ腸およびマルピーギ管 などにおけるフラボキナーゼ活性の以北:なl二叫が認、められたが、 このフ ラボキナーゼ活性の卜寸11.がネ111�1-ぷ7l��
Bウ のや111允に役だっているものと迎解しうる.
一方, 5 ffr令起蚕からBゥ�1Hr�添加削育を開始した場合のように休 内のB:2欠乏がまださほど進んでおらず, 補酵素型のビタミンBゥにつ い て恒常性が依然として保たれており, カイコの成長も対照と比較して遜 色ないj犬況では, フラボキナーゼ活性に変化は認められなかった. この 時, すでにビタミンBゥの↑!日行性維持機構が允動しているものとも考えら れる これにはフラボキナーゼ爪'1:'1:の仁Jill土必要とせず既存のフラボキ ナーゼで充分なのであろう. 一般にFADまたはFMNを補酵素とするフ ラピン惇素は補酵素との結合が極めて強いことが知られており, このた めリボフラビンプールからの収り込みが谷易であり フラボキナーゼLー の増加がなくても恒'日?性が保たれるという解釈もなりたつ. 極度のビタ ミン B2 欠乏に陥った場合は各組織のフラボキナーゼのよ同大によりマル ピーギ管などイ本内のリボフラビンを, 体液を通じて, できるだけ多く取 り込み補陣素型とするものと考えられる• Table 10に掲げた値から,標準 の飼料で飼育した場合の体液のリボフラピン濃度は5μMのレベルであ
ると推察され、 この値はフラボキナーゼのKm (1.4μM)よりも|毎いので,体液に浮遊する組織の衣出における醇ぷ反応はVmax にあることが期待で きる. ところが, 3齢からB2欠にすると休液のリボフラピンは0.01μM のレベルとなり Km より低くなるため, 問手素量の明大が竜安となるもの
と考えられる
rll腸におけるフラボキナーゼ、iffyl;のJill大は, 深刻なビタミンBゥ欠乏状 態のもとでIriJビタミンが版取された場合に 通常におけるよりもさらに 速やかにこのビタミンを吸収するのにィ可利であると与-えられる.
111
)J揚の フラボキナーゼのK はラット肝臓の場介よりもl桁低いの で, 11m乳類 の場合よりもさらに効半的な反応が起るものと推察される.今同FMNからFADの合成に|刻lJ-する防素であるFADピロフォスフ ォリラーゼの活性は測定し なか った. 11,-1]乳類においては, ビタミンBっ欠
乏時においてフラボキナーゼ治性には変化がないがFADピロフォスフ ォリラーゼ、の活性が1-.糾すると報(与されている(Fass
and Rivlin,
]969).
般に, 動物組織1!-,にistも多くイバ1:しているビタミンBゥ化合物はFAD であ り, 世(1<]にはFADが故もTE12に なる. したがって附乳類においては FADの量をイ確保するために FADピロフォスフォリラーゼの活性が上昇 すると与-えられている カイコにおいても, FADが最も多い組織(1Þ腸ラ 脂肪体, 絹糸腺など)を佃々に調べると, このよう な変動も観察される了 能性がある. しかし カイコで、は11,打乳動物で、は観察で、きないとされるフラ ボキナーゼ活性のと丹がみられた この速いは通常の条件下でカイコ体 内に遊離別リボフラピンが大世にみ積しており, これが体液を通じて補
酵素型に転換されるとする考えと介致する.
以上のように カイコにお けるリボフラピンの存イ1:と分布の様式およびその利)目システムの 一環を 明らかにした. これがカイコに特イ言なものかまたは他の昆虫にも みられ る一般性があるのかはイベI�
j であり今後の仰究が必
裂となる.- 75 -
11荷 主i
リボフラビンからFMNを形成するM京\ フラボキナーゼについて調 べた. 同醇素の活性はマル ピーギ符に強く認められ 大量のリボフラピ ンがここに議積する|僚に本酵素が役だ、っているものと思われた. 全活性 でみた場場.合には111 )腸j助易が|向4ι1いフラボボ、キナ一ゼ、j活百斤刑J性|
夕ミン B2の食物からのJ渋兵収に機能しているものと考考.えられる. 次に, 3 齢、からビタミンB')無添加で飼育にした場合 5齢期では中腸およびマル
ピーギ管におけるフラボキナーゼ活性が高かった. この上昇は補酵素型 B2の補充に役だっているものと抗どきされる• 5齢起蚕からBっ無添加飼育 を開始した場合にはフラボキナーゼmf|;に変化はみられず, 極度のビタ ミンB2欠乏に陥った場介にのみフラボキナーゼ活性のよl目大が生じ, リボ フラビンプールから術陣素)r�を補充するように機能するものと判断した.
第111章
ビタミンB2欠乏とキサンチン脱水素酵素活性
第I主主において, ビタミンB勺fHtjぷ}JI)
Í!lij
�Ej-で飼育した場合のカイコにお けるビタミンB守 合ftの変化のよrr 先によって, ビタミンBゥ補酵ぷ別に|却 する恒常性があることをIY-Jらかにした. カイコの成長もこの恒常性維持 の程度に依存しており, ビタミンB勺の十|liMJぷ21J!が允分呈:保証されている
間は正常に成長したが, 恒常性が}J)Jれると成長も劣るようになった. こ のように兵なるビタミンB勺レベルにあるカイコイ本内において,しいミ かなる理状態のF煮 茅主名 .異が'
1
阿、円斤円J?常jく仁, 本章ではB2 無添加例料で飼育した場合にB勺を補酵素とする酵ヨ素 素ミミ.のの
活性が変動するか子作?かについて調べた.このような酵素の代表として,ニ ワトリにおいてFADを2倒を含むことが知られており (Rajagopalan and
Handler, 1967),
カイコにおいてもFADを含むフラピン酵素である こ と が報告されている(林ヲ1962)キサンチン脱水素防:素を選んだ. これは核 酸成分の異化代謝-においてヒポキサンチンからキサンチンを経由して/λ 酸に転換することから, 昆虫, 鳥類のように窒素化合物の主要な排11lt�が尿酸である(uricotehc)生物では重安な地位を占める酵素である.
ー77 -
材料とjjU;
供試五三品穐およびflrîJif
用いたカイコの品純は1=1 140
�j' X支145号である.
対照およびリボフ ラピン欠乏状態Fにおける飼育条件は前市までと|寸ーとした.キサンチン脱水素酵ぷ活性のüW定
既報のノ7法(Rajagopalan and Handler, 1967)に準じてキサンチン脱水素 酵素前竹の測定を行った. tllUn緩衝液[0.1mMエチレンジアミン四酢酸
カリウム(EDTA-3K)を合む50 nlMりん酸カリウム緩衡法(KPB), pH
7.8J I-þ で
組織を磨砕しね�:られたホモジネ ー
トを27ラ OOOgで30 分
間遠
心 分離後, 上清を酵素液としてfけいた反応液(全量0.6ml)は50mM KPB (pH 7.8),0.1
mM EDTA-3K,0.5 mMニコチンアミドアデニンジヌクレオ
チド(NAD),
0.15
mMキサンチンナトリウムおよび 0.05 -0. 14 m1の←醇
糸液を合むように剖終した反応、被にFADを添加する場合 同化合物の濃度は 0.01mMとした 反応液を250Cに設定した分光光度計セル|付に
静置し, 反応(キサンチン+ NAD ++ HゥO→尿酸+ NADH + H+ )により生 ずる NADHを340 nmにおけるl汲光度によってモニターし酵素活性 (nmol/minで表示)を求めた. 恭子:であるキサンチンナトリウムを添加し
ないものを盲検とした 前章で述べたピュレット法を適用して(ウシ�Tl 清アルブミンを標準としつつ)タンパク質濃度を定量し, キサンチン脱 水素酵
素
の比活性を算出した川本14円。+小
カイコ幼虫の脂肪体 ql )助 後部紺糸)J以およびマルピーギ??の各組織 をそれぞれ20, 15, 15,
50
併訟の1111
出緩衝液中で1111
山し,
仁治について キサンチン脱ノ1(/(�醇ぷ出'111=を測定した(Table20). 1長:も高い活性がみらTablc 20. Activily 01' xanthinc dchydrogcnase in larva1 tissucs.
Tissuc Enχyme acti vity
(nmol/min/g frcsh wし) (nmol/min/mg protein) (nmol/min/animal)
円、け,パU
P3 門川pu
'BEE-A K M りと LU uu
O九 釦ルdiωh
r
t
・ EL'i
吋l
u1 4 . ,i nv と 'i t
'm
pb白川γ
1・
K 山 a
RMRM 858.3 1 52.8
164.り13 J�
51.61り.2 208.8士33.3
13_97 :t ()_80 3.1 510.16 1.22士0.22 5.05 1 0.21
149.1 :t 9.9 53.01 2.3 1 9.1:t2.9 3.8 :t 0.5
Ma1c 1arvac ofN140 x C145 WCl・c llscd on day 4 01' 5th instar.
Each valuc reprcscnts thc mcan士S.D.
れたのは脂肪体であり, マルビーギ管とl判j易の活性がこれに次いで尚くラ 後部絹糸mí�の活性は低かった マルビーギ管は組織の全活性(表では
nmol/animalと表示)では小さなイ[IJ[で、あった 最も活性が高い脂肪体につ いて
5
齢別問11-1
における活性の変動をよ1l跡した(Fig. 16). 組織重当たり の活性は5 齢の後半に活性が上昇する傾向jが認められたが タンパク質、lノたりの活性では5齢期間l-IIほぼ一定の他であった 全活性は組織の発 達に伴って著しく上JEll-した.
四79 -
1200
C 1000
800 200
600
,ー、、
C
ε 400
。 150
E 200
亡
、---
4〉
〉
回d、 。
+に何-コd 20
�
Bω
100
U E E 〉
AJ
』、 15
10 50
5
。 。
。 2 3 4 5 6 。 2 3 4 5 6
Day in 5th instar
Fig. 16. Changes of xanthinc dehydrogenase act ivity in thc rat body during the 5th instar.
Male larvae ofN140 xC145 werc used.
Each vcrtical line shows the S.D.
A : Enzymc activity was exprcssed in nmolJminJg fresh weight.
B : Enzymc activity was expressed in nmolJmin/mg pl・otcin.
C : Enzymc activity was expresscd in nmolJmin/animal.
次にビタミン
B2
レベルが低卜aした状態における脂肪体のキサンチン 脱水素醇ぷ1!îttについて検討する[I
(1<)で, 5出合または3齢起五三よりリボ フラピン川添加飼料でfl'iJ台し,5出合 41二!のllL5111;で、本国子:ぷ的性を測定した
(Table 21).
5齢から}![f{添加!とした場介には的性は刈']1包と差呉はなかった.酵素活性測定の|僚にキサンチンJJ}�ノkjd防素の制酵素で、ある FAD を添加 しでも測定他に変化はなかった. ーノi, 3齢からリボフラビン川添加飼料 で飼育した場介は5齢4 "における昨ぷ:111竹の低下が認められ, 来11織の ふ活性は約半分にまで低下した さらにFAD添加効果も若干認められヲ これにより酵素活性がやや111く測定された しかし, 組.織の全活性が対
照の値にまで復活することはなかった.
去J 作/J、
カイコにおけるキサンチン脱水素酔素は従来の報告(林, 1961)と同様 に脂肪体に最も高い活性が認められた 脂肪体の醇索活性は 5齢別問11' ほぼ一定であったことから, 飼料rl'の栄養素の影響を検討する|努に経過 日数の述いによる差jidは考応しなくてもよいものとした. そこで リボ フラピン無添加飼料で飼育した幼虫の脂肪休の酵素-活性について検討し たところラ51-愉起蚕からf!!t添加飼料で飼育した区では5齢4日の幼虫の脂 肪イ本の醇ヲjt-活性には対J!前との間に左兵はみられなかった. よって この 飼育条件下ではキサンチン脱水素醇ぷ活性に対する彩響はほとんどない ものと結論した. Mぷ活YI:: rlllJ定の反応液にFADを添加しでも隣家;11I'性別1
- 8] -
Table 21. Etlects of ribollavin-dcficicnt dict on xanthinc dchydrogcnasc acLivity in the fat body.
Enzymc aじLivity
(nmollminJg frcsh wt.) (nmül/minJmg prütcin) (nmollminJa凶mal)
Contl・01 -FAO 1)
+FAO")
1) ユ)
(-)B2-V 3) -FAO 1)
+FAO-')
1) 2)
(一)B2-III4) -FAO 1)
+FAO 2)
1) 2)
906.5::t 85.8 18.()l士1.56 888.9::t 37.2 17.72::t 1.60 -17.6::t 117.7 一0.29::t 2.26
933.6::t 92.8 17.2り::t ] .35 961.8士 78.5 17.82::t 1.13 28.2::t 64.5 0.53 ::t 1.15
669.9::t 65.2 15.41士1.06 715.7::t 51.2 16.49::t 1.07
45.8::t 20.2 ト07::t 0.51
The silkwonn used was 4 day old 5th instar malc larva of N 140 x C 145.
Each va1uc reprcscnts thc mcan::t S.O.
1) Enzymc assay was caITied on without FAO.
2) Enzymc assay was じaITicd on with FAO.
115.0::t 21.6 114.7::t 30.8 -0.3::t 14.5
124.R::t 13.7 129.1::t ]8.0 4.3::t 8.42
57.8::t ]0.9 61.8::t 11.7 4.0士2.06
3) Larvac wcrc fcd on ribotlavin-dcL・icient dict 1'rom thc bcgining 01' thc 5th instar.
4) Larvae wcre fcd on ribotlavin-dc1ïcicnt dict 1、1・om thc bcgining 01' thc 31・d instar.
Signi1ïcantly diffrent 1'rom control by t-tcst at 1私IcvcI.
定イl立にiE特はなかった.
FAD
とがi介していない昨ぷ(アポ防ぷ)はほと んどないと結l論される. これは, mI LZにおいて5f,給起蚕から無添加|飼 料で飼育しでもカイコの成長は良好であり, ビタミンB守合量もそのネ111M素型に関しては恒常JIゾ1:があったことと 一致する --方, 3齢起蚕から411日ぷ 加飼料で飼育した幼虫では5齢における防索活性に低下がみられた. こ の場合, 反応液にFADを添加するとMぷ-活性が若|二lfJくなり, アポ防素 が存イ正しているものとィ与えられる. すなわちビタミンB今が充足している 時はキサンチン脱水素醇ぷも充分l12存イ1:し かっFADで飽和されている ものの, ビタミンB勺欠乏状態になると的性のあるキサンチン脱ノk素酵素 量は低下しさらにその一部はFADと結介していない状態にあるものと 結論される Table
]
1にぷノJミしたように3的からリボフラピン欠とする と脂肪体のFAD波皮は村山のがJ 1/3となる• r-�'手ぷ活性は組織豆、!?たりで 対照のがJ 709らであり,ネ"J�f�ミの減少よりも昨素iElqの低下が小さい こ れはFADのプールに余俗がなく,イ災づt (1<]に酵素に結合していることを表 している I同タンパク質の人工飼料で飼育するとカイコの成長が品進さ れ(イ列候・[!Jlþ, ]962)ヲ この時脂肪体における種々の隣友活性がJ-_昇し ていることが械告されている(Ito and Mukaiyama, ] 964). カイコの生以 状態とキサンチン脱水素r�ぷ活性にはll�の相|共jがあり, キサンチン脱水 系酵素活性の低下はカイコの生四状態が必化している兆依であると考え られる.- 83 -
J向 �:
幼虫の脂肪体, 仁1-1)湯, 後部絹糸)J��およびマルピーギ管の各組織の相J山 出液を月jいてキサンチンJ1見ノk素f将来店'[1tを測定したところ, 最も|匂い活 性がみられたのは脂肪休であった)J\1)J)j休について5齢別山111における 活性の変動を追跡したところ, 全活性は組織の発達に作って[-.昇した
ピタミンB2レベルが低ドした状態では, 5前からリボフラビン無添加と した場合, 5齢におけるキサンチン脱ノk玄酵素活性には変化がなかった が, 3 �í令から;nr.添加とした場合は5齢における活性の低下が認められた
さらにFAD添加効果が本パ二認められた.