食用担子菌の成育 に及ぼす
カ ラマ ツ水抽 出物 の影響 に関す る研究
1996と
F
鳥取 大学 大学 院連 合農 学研 究科 生物 生産科学専 攻高畠幸司
Effects of Water Extracts from Larch Wood
on Mycelial Growth of Edible Basidiomycete Fungi 1996
The United GradiSch` of AgriciSci., Tottori Univ. XO,i Takabatake
目 次 緒 言 第
I章
第I章
既往 の研 究 数種食 用担子菌 の菌 糸体成長 に及 ぼすカ ラマ ツ水抽 出物 の影 響1
カ ラマ ツ水抽 出物 の化学組 成1.1
材料お よび方法1.2
結果
2
産地 別 カ ラマ ツの水 抽 出2.1
材料 お よび方法 物 添 加培地 におけ る菌 糸体成 長 の比較 ―…2.2
結果 シベ リア産 カ ラマ ツの水
3.1
材料 お よび方法 抽 出物添 加培地 におけ る菌 糸体成 長3.2
結果
4考
察5結
論 第 Ⅲ章 ヒラタケ菌 によるカラマツ水抽 出物の資化性 と菌体外酵素の産 出 ……1
カ ラマ ツ水抽 出物 の資化 性1.1
材料 お よび方法1.2
結果
2
フ ェノール酸化酵 素 の産 出2.1
材料お よび方法2.2
結果 多糖分解酵素 の産 出
3.1
材料 お よび方法3.2
結果 考 察 結 論 4 5 1 4 9 9 9 10 14 14 15 24 24 25 28 32 34 34 34 35 47 47 48 48 48 51 54 56
第 Ⅳ章 カ ラマ ツ水抽 出物 に よる ヒラタケ菌 の菌糸体成 長 と 菌体外酵素 活性 の経 時変化 との関係 ―… …
1
菌 糸体成長 の変化1.1
材料 お よび方法1.2
結果 フェノール酸化酵素 の経時変化
2.1
材料 お よび方法2.2
結果
3
多糖 分解酵素 の経 時変化3,1
材料 お よび方法3.2
結果 58 58 58 59 59 59 61 66 66 66 70 77 79 80 80 80 83 83 84 95 98 100 107 110 112 124 128 第
V章
カ ラマ ツ水 抽 出物 の ヒラ タケ栽培 へ の利用1
鋸屑培 地 に よる栽 培 へ の利 用 1。1
材料 お よび方法1.2
結果
2
未利 用基材培 地 に よ る栽培 へ の利 用2.1
材料 お よび方法2.2
結果
4考
察5結
論3考
察4結
論 第Ⅵ章 総 合考察 結 言 謝 辞 引用文 献Summary
……… ………… …… ……… …… …… ……… ……… Ⅲ 公表論文 の リス ト …… ……… ……… ……… …… ………緒 _ 我 が国にお いて
,食
用担子菌類,即
ち「 きの こ」は平安時代 の歌集 に「松茸 ・平茸 ・ 滑薄 (なめすす き)」 と詠 まれ,か
な り古 くか ら食 に供 され,親
しまれていた と思われ る (小川,1978)。
また,椎
茸 は安土 ・桃 山時代 には上流社会 の料理 に盛 んに使 われ るよう にな り,江
戸時代 の中期 頃 よ り栽培が始 まった とされている (中村,1983)。
しか し近代 的な栽培形態 である純粋培養 した菌糸体 を接種す る栽培方法 は1940年 代 に確立 され (中村,1982),鋸
屑 と米糠 やフスマ等 の栄養材 を混合 した培地 で栽培す る方法 は1950年 代 か ら長 野県 を中心 に普及 が始 まった (衣川,1982)。
現在,食
用 きの こ類 の総生産額 は1993年 度 実績 で2,732億 円 とな り (林野庁,1995),1975年
(1,265億 円)以
降飛躍的な増加 を示 し, 食用 きの こ類 は農林作物 の中で主要 な位置 を占めるに至 った。 この ように食用 きの こ類 の 生産が大 き く成長 した背景 には,消
費者 の嗜好 が 自然 ・健康食品に対 して高 まってiき
の この食品イメージが消費者のニーズに合致 した ことによ り需要が拡大 された こと,き
の こ 栽培 に も施設 を利用 した生産方式 が導入 されて一年 を通 じて新鮮 な食用 きの こを供給 でき るようになった ことが考 え られ る。 しか し,き
の こ生産農家 を取 り巻 く栽培状況は厳 しく,依
然 としてほ とん どの生産農家 が零細規模 であ り,栽
培技術 は個 々の経験 に依存 しているところが多 く,き
の こ栽培 にお ける生産効率の 目安 とな るホダ木 や培養基 の単位重量当た りのきの こ収量 は未 だに不安定 である。また,1990年
以降:中
国 よ り安価 な生 シイ タケの輸入量が急増 し (林野庁,1995)
消費者 か らは近年のグル メブームを反映 して野生味溢 れ る香 りと歯 ざわ りを備 えたきの こ が求 め られている。 このため,国
際競争 に耐 る ことができ,
しか も消費者 の嗜好 の多様性 に沿 った高品質な きの こを安定 して栽培 できる高位安定化栽培技術 の確立が必要 とされて いる (特用林産 ビジ ョン検討会,1998)。
きの こ栽培 にお いて,菌
糸体 の培養 と子実体形成 は重要 な過程 であ り,
この点 について ホダ木 ・培養基 内におけ る菌糸体 の動態 と腐朽様式 (林 原 ら,1983;中
井,1986;Tuneda
et al,,1987,1989 i Tuneda,1988;Ohira et al.,1991,1992i大
平 ら,1993),菌
糸 体 の栄養生理(Ishikawa,1967i北
木 ら,1978,1980i阪
本 ら,1978;Tokimoto・
Kawai,-1975;杉
森 ら,1971;吉
田 ら,1965i大
山 ら,1976i永
曽・吉川,1975,1978;Niderpr―
uem et al.,1964,Mclaughlin,1988),菌
糸体の培養過程 で生 じるホダ木 ・培地成分の変化 (石川 ら
,1961i沖
ら,1981;川
上,1978;田
中 ら,1984i橋
本 ら,1966;時
木 ら,1982),ホ
ダ木 の腐朽度 や培地 の成熟度 と子実体形成 との関係 (福田 ら,1987,1988;
Ohga et al.,1992a),ホ
ダ木 や培養基 内の菌体量 と子実体形成 との関係 (Tokittoto・Fukuda 1981,Ohga,1990a)に
関 して詳細 に論 じられ,菌
糸体 がほだ木 や培養基 を充分に 腐朽 し,菌
体量が多 くな るに従 い子実体収量の増加が期待 で き,
さ らにホ ダ木 や培養基 内 に速 やかに菌糸体 が蔓延すれば トリコデルマ菌等 の害菌 を防御 で きる こと (大賀 ・近藤11978,1981;日
本 きの こセ ンター,1977)が
明 らかに され た。 したが って,き
の こ栽培 に おいて菌糸体培養 の成否はその後 の子実体収量 に多大 な影響 を及ぼす もの と考 え られ る。 この ような観点か ら,ホ
ダ木 や培養基 の菌糸体の成 育 を促 し,そ
の後の子実体形成 を容 易 にす るきの こ菌糸体への成長促進剤 の開発 は,き
の こ栽培 におけ る今 日的な技術課題 を 解決す る上 で重要 な役割 を果 たす もの思われ る。 これ までに担子菌 の成長促進剤 として, リグニ ンお よび リグニ ン前駆物質 (河村 ら,1983i池
ヶ谷 ・後藤,1988),ウ
ロン酸 (池 ヶ谷 ら,1993),亜
硫酸パルプ廃液成分 (稲葉 ら,1979,1981 i Zadrttil,1974),ネ
ギ煎汁成分 (大賀
:1988),木
酢液蒸留成分 (Yoshimura・Hayakawa,1991,1993, Ohta・
Zhang,1994),植
物 ホルモ ン2, 4-ジ
ク ロロフェノキ シ酢酸 (Tsujiyam et al。,1993),
さらに子実体誘導物質 としてサ イク リックーAMP(宇
野 ・石川:1974),酸
性 プ ロテア ーゼ阻害斉JS P-1(寺
下 ら,198011981;村
尾,1985),セ
レブ ロシ ド(Kawai・ Ikedat1982,1983),安
息香酸 (村 尾,1986),酵
母抽 出物 (松尾 ら,1992),各
種担子菌類 の 抽 出液(Urayama,1969)等
が報告 されてお り,き
の こ菌糸体の栄養成長や子実体形成 を生 理 ・生化学的 に解 明す る上 で貴重な知見 が蓄積 されてきている。 しか し,き
の こ生産農家 が実際の栽培 に使用す るには,ま
ず成長促進剤 は安価 で,安
定 して入手 できる ことが求 め られ るが,
この要件 に関 しては未 だに充分に満たされた状態ではない。 ところで,第
二次世界大戦後 の拡大造林 によ り,特
に北海道 。長野県 を中心 に寒冷地域では
,成
長が早 い ことか らカ ラマツ造林 が積極的に行 われ,現
在,育
成 されたカラマツ材 は構造材な どに利用 されて いる。 また,
日本海狽Jの地域 を中心 にシベ リア産のカ ラマツ材 が毎年多量 に輸入 されている。 これ らカラマツ材 を製材す る際には多量の鋸屑 が生 じてい るが,カ
ラマツ製材鋸屑 は毛羽立 ちが著 しく,ア
ク抜 きが難 しいとの理 由か ら,製
材鋸屑 の一般的な用途 である家畜敷料や きの こ培地基材 として,ほ
とん ど利用 されていな い。 し か し,カ
ラマツ材 中に水溶性多糖類 であるアラ ビノガラクタ ンが多 く合有 している ことに 着 日し,最
近 にな ってカラマツ製材鋸屑 を利用 してアラ ビノガラクタ ンを製造す る ことが 可能 となった (水 本,1991,1994)。
このアラ ビノガラクタ ンを製造す る工程 の中間生成 物 であるカラマツ水抽 出物 は,安
定 して多量 に,
しか も低 コス トで生産 で きる。 この水抽 出物が担子菌の菌糸体成長 に対 して促進作用 を示せば,食
用 きの こ栽培 に利用す る ことが 可能 となる。 そ こで本研究 では,カ
ラマツ水抽 出物 の きの こ菌糸体 の成長 と子実体形成 に及ぼす影響 を明 らかに してカラマツ水抽 出物 を食用 きの こ栽培の成長促進剤 として利用す ることを 目 的に,第
I章
では担子菌類 の成長 に影響 を及ぼす樹木抽 出物,カ
ラマツ材抽 出物 に関す る 既往 の研究 を述 べ,第
Ⅱ章 では,産
地 の異 な るカラマツ材,す
なわち シベ リア産,長
野県 産,北
海道産 のカラマツ材 につ いて,そ
の水抽 出物 の化学組成 を明 らかに し,各
々の水抽 出物 が数種 の食用 きの こ菌糸体 の成長 に及ぼす影響 を比較検討 した。第 Ⅲ章 ではカ ラマツ 水抽 出物の添加 によ り最 も顕著 な菌糸体成長 を示 した ヒラタケ菌 を用 いて,カ
ラマツ水抽 出物 の資化性 と菌体外酵素 の産 出状況 を,第
Ⅳ章では培養経過に伴 う菌糸体成長の変化 と 菌体外酵素活性 の経時変化 を明 らかに して,菌
糸体成長 に及ぼすカラマツ水抽 出物 の促進 作用 とヒラタケ菌 によるカラマツ水抽 出物 の分解様式 を論議 した。第V章
ではカ ラマツ水 抽 出物 を鋸屑培地 お よび鋸屑以外 の未利用基材 の培地 に添加 して ヒラタケ栽培 を試 み,カ
ラマツ水抽 出物 の実用的な利用性 につ いて検討 した。 これ らの結果 に基づ き,第
Ⅵ章 では カラマツ水抽出物の食用担子菌類 に及ぼす生理作用 と食用 きの こ栽培への利用性 について, 今後 の展望 を合 めて総合的 に論議 した。-3-第 二章 既往の研究 担子菌類 の成長 に及ぼす樹木抽 出物 の研究 は
,木
材腐朽菌 に耐久性 を有す る樹木抽 出物 の解明,即
ち担子菌類の成長 に阻害作用 を示す成分の検索 を中心 に展 開 されて きた。イス ノキ,カ
ラマツ,ベ
イマツの タキ シフ ォ リン,
ヒノキの トロポ ロン類,ベ
イ ヒバ の ヌー ト カチ ン,ユ
ーカ リの スチルベ ン類,イ
ンセスシーダのカルバ クロール,パ
ドクの没食子酸, ホ ワイ トオークのエ ラグタ ンニ ン,ベ
イスギの ツヤプ リシ ン類,ク
リのタ ンニ ン (善本,1983)と
樹木抽 出物 由来 の多数の抗菌成分が報告 されている。 これ ら抗菌成分のバ イオア ッセイには,木
材腐朽菌 を対象 としているため,カ
ワラタケ 釣rrοryS ttrdlicο′θ4オ
オ ウズフタケ 物r伽/じθS pβ′′strFs
ワタグサ レタケ 乃rf2 7ppο″ デβ,
ヒイ ロタケ乃′
/stuictys sβpglr'コθ
ysが
供試され
,いずれも硬質菌類が用いられた。
食用 きの こ栽培 が盛んにな るにつれて,原
木栽培 のホダ木適性 や菌床栽培 におけ る培地適 性 に関す る研究が進 め られ るようにな り,樹
木 の抽 出成分 と食用担子菌類 の菌糸体成長 と の関係,特
に針菜樹材抽 出成分中の阻害成分について論議 され るようになった。大賀 ら(1977)は
,シ
イタケ と餌tJipps θヵ胞sに
阻害作用 を示 すアカマツ,ヤ
マモモの抽 出成 分 を検索 した結果,ア
カマツでは脂肪酸 ・フ ェノール類,ヤ
マモモでは ミリシ トリンが各 々阻害物質 であることを明 らかに した。金城 ・近藤(1979)は
,針
葉樹材 (ヒ ノキ,ア
カ マツ,カ
ラマツ,ス
ギの心辺材)の
エタノール・ベ ンゼ ン抽出物 には,シ
イタケ,
ヒラタ ケ P′θEFrOιys οδιrθβιys
エ ノキ タケ Fこmp′
力2確
′′ιuipθsナ
メコ 励0"OιβЛ潟〃机dこ対す る阻害作用があ り
,金
城 ・屋我(1986)は
,特
に阻害性 の著 しい ヒノキ をと りあげ,阻害成分 として特muurolol, α―
cadinolが
含 まれていることを明 らかに した。中島 ら (1980)は
,ス
ギ材 ではシイタケ菌の成育が劣 るので,ス
ギ 内皮お よび辺材 中の シイ タケ菌に対す る阻害成分 を検討 した ところ
,多
量 のフ ェルギ ノールの存在 を確認 した。米 山 ら (1989)は
,
トドマツ材 のアル コール抽 出物 中のヘキサ ン可溶部 にはシイ タケ,エ
ノキ タケ,ヒラタケ
iナ
メコ,マ
イタケ σrfroFa rr伽 ぬa
タモギ タケ P′θ′rοιps cοr口μo9p力 θ,ηr.c」ιrfコ叩 ′′θβ
tysに
対す る阻害作用が あ り,Yoneyama et al。(1990)は
セ スキテルペ ン類のジュバ ビオ ンが原 因物質 であることを明 らかに した。三浦 ら(1984)は
,タ
モギ タケに対す るエゾヤマザ クラ抽 出物 の影響 を検討 し
,フ
ラボノイ ド類 が菌糸伸長に阻害作用 を示す ことを明 らかに した。高 畠(1987.1989)は
I針
葉樹2樹種 ・広葉樹13樹種 の辺心材 木粉 におけるシイ タケiナ
メコi
ヒラタケの培地適性 を検討 し,
シイタケに対 してホオ ノ キ ・ ヒノキの辺心材i ウワミズザ クラ・スギ ・ニセアカシア・コシアブラの心材,ナ
メコ にはク リ・ ウワミズザクラ・ホオ ノキ ・ニセアカシアの心材: ヒラタケにはホオ ノキ辺心 材〕ニセアカ シア ・ク リ・コシアブラの心材 の各温水抽 出物 が成育阻害作用 を示す ことを 指摘 した。 また,担
子菌以外 の菌類 に関 して,善
本 ら(1984)は
,コ
ナラ属 (θ口θrじμs)樹
木 (コ ナラ,
ミズナ ラ,ク
ヌギ)の
樹皮温水抽 出物 中に存在す るポ リフラバ ノール,ポ
リ ガ ロイルエステル様化合物が食用 きの こ栽培の害菌である トリコデルマ菌類の成長 を阻害 す る ことを明 らかに してお り,鮫
島 ・善本(1984)は
,ス
ギ,
ヒノキ,ア
カマツ,カ
ラマ ッの針葉樹温水抽 出物 に も トリコデルマ菌や ヒポク レア菌の成育 を阻害す る成分が存在す る ことを報告 している。 一方,担
子菌類 の成育に促進作用 を示す樹木抽出物 に関す る研究は乏 し く,ス
ギ,
ヒノ キ,ア
カマツ材の内樹皮温水抽出物がシイタケ,キ
クラゲの菌糸体成 育に (鮫 島 ・善本:1984),シ
ナノキ辺材部 や コナラ,
ミズナラ・ クヌギの温水抽出物がシイタケの菌糸体成 長 に (松尾,1957i善
本 ら]1984),イ
チ ョウ生葉 の酢酸 エチル抽 出物 がシイタケ・ エノ キ タケの菌糸体成長 と子実体形成 に促進作用 を示す (沖・橘,1994,1995)こ
とが報告 さ れている程度 である。また,西
田 ら(1992,1993,1994,1995)は
担子菌類 の宿主選択性 に注 目して,サ
クラ (Pr2rp′s)属
の樹木 に発生 しているカ ワウソタケ れ,Oι "d〃rねヵ ′ゃ エゴノキにのみ発生するエゴノキ タケ(肋
θ胞 ′θttsrs st/racゴ Л2)は
,そ
れぞれサ クラと エゴノキの温水抽 出物,特
にフラボノイ ド系 の配糖体や リグナ ン系の配糖体 によって菌糸 体成長 と胞子 の発芽 が促 され る ことを明 らかに して いる。 しか し・ いずれの樹木抽出物 も 促進作用 を示す担子菌の種類 は,一
な い し二種類 に限定 されお り,広
範 囲な菌種 に対す る 作用は明 らかに されていな い。 カラマツ属 (ιβrゴχ)の
樹木 は世界的 に広 く分布 してお り,カ
ラマツ材 いわゆ るlarchwoodに は, European larch (とβr′χどθθ′J2)i Dahurian larch(と ,ど〃θ′√盟rr), Kurili一
-5-an larch(と .=〃θ′ゴコゴF, 7β r.プβρOIPrca), Tamarack larch(と ,′βrrじゴЛβ
),
」apaneselarch(と,feptο′
Qpsrs),sabalpine(mountain)larch(と
.′/β′′デr),western larch
(と,οじじ
'Jepι
βr′
d),
と,Pοιβ刀′河デザ, Siberian larch(と,drヵθr,c溜),
と,s21色2じzθyゴゴが ある。一般 に
,針
葉樹 材 には1%未
満 のア ラ ビノガ ラク タ ンを合 ん で いるが,larch woodの
心 材部 に限 っては,多
量 のア ラ ビノガ ラク タ ンが仮導管 や放射 組織 の細胞 内腔 に沈着 して合まれており,そ の合有量は
5∼30%で
ある
(C6tёet al.i1966)。
アラビノガラクタンの
合有量と樹幹内での分布は
,C6tёet al.(1966,1967),C6tё
・TimeH(1967)に より
,Tamarack larch,European larch,Westem larch
につ いて詳細 に検討 され ている。その 結果,ア
ラ ビノガラクタ ンの合有量は,樹
種 によって異 な り,年
輪幅が狭 く,樹
高が高 く, 樹齢が低 い部位 で多 くな り,樹
幹 よ りも枝 の方 が,ア
テ材 よ り正常材 の方 が多 くな ること, また,ア
ラ ビノガ ラクタ ンの水平分布 は,髄
周辺 よ り心辺材 の境界付近 にかけて合有量 が 多 くな り,心
辺材 の境界部位 で最大合有量 を示 し,そ
こか ら辺材部 にかけてアラ ビノガラ クタ ンの合有量 は急激 に減少 し,辺
材部分では低 い合有量 で推移 す る ことが明 らかに され た。 このような水平分布 の傾 向は,橋
爪 ・高橋(1974)の
日本カラマツや水本 ら (1994) のシベ リア産カラマツにおいて も同様 な傾 向が認 め られている。 さらに,Western larch
において,樹
齢が高 くなるに従 い,材
中の有機酸濃度 が高 くな リアラ ビノガラクタ ンが加 水分解 を受 けて低分子化 され ていることが提示 されている。カラマツ材 由来 のア ラ ビノガ ラクタ ンの構造 は,Tamarack larchの
ア ラ ビノガラクタ ンに関 して Fu・TimeH(1972),
Lynch et al.,(1968)に
よ り,
」apanese larchに
関 してAspinall et al.,(1968),
Teratani et al.,(1987)に
よ り検討 されてお り,い
ずれのアラ ビノガラクタ ンも β―D
(1→3)結
合 したポ リガ ラク トースを主鎖 として,
これにO-6位 でD―ガラク トース,L一
ア ラ ビノース,グ
ルク ロン酸か らなる側鎖が分岐 していることが明 らかにされた。 しか し, 樹種 によ り狽J鎖の分岐の仕方が異な り]微
妙 に分子構造 が異な ってお り,カ
ラマツ材 由来 のアラ ビノガラクタ ンの詳細な構造は明 らかにされていない。 カラマツ材,」apanese larchの
フ ェノール成分 に関 しては,出
町 ら(1968)に
よ り心材部のフェノール成分 につ いて
,竹
原 ・笹谷(1979)に
よ り辺材部 のフ ェノール成分 につ い て,笹
谷 ら(1980,1987)に
よ り樹幹 内のフ ェノール成分の分布 につ いて詳細 に検討 され ている。すなわち,材
中の主要 なフェノール成分は タキ ンフ ォリン,カ
ツラニ ンのフラバ ノール類 であ り,ケ
ルセチ ン,ケ
ンフ ェロールのフラボノール類 が少量存在 している。 こ れ らのフラボノイ ド類の樹幹 内での分布 は,半
径方 向で形成層隣接 の辺材 か ら心材へ と緩 やかに増加 し,辺
心材境界 の心材狽]で急激 に増加 し,髄
へ と徐 々に減少 してい く傾 向が認 め られている。フラボノイ ド類以外のフ ェノール成分 として リグナ ン類 が確認 されている。 このフラボノイ ド類の分布傾 向は,ア
ラ ビノガラクタ ンの分布傾 向 と類似 しているが,両
者 の相互間系 につ いては明 らかに されていな い。 これまでのカ ラマツ材 に関連 した担子菌類の研究は,主
に材質腐朽菌 について検討 され てきた。近年,カ
ラマツ材 の良質材生産のため大径木施業 が行われているが,カ
ラマッは 病害 に弱 い樹種 であ り,生
立木 の材質腐朽病が多 く,腐
朽病 の被害 は樹齢が増加す るとと もに状大 している。特 に根株心腐病 は深亥Jな被害 をもた らしている。カラマツ根株心腐病 に関す る研究 は,Ohsawa et al.(1987,1992,1994)に
よ り,八
ヶ岳東 山麓!富
士山麓 で 被害実態i病
原菌 の分離 とその遷移,羅
病木 に成育す る菌類 による木材成分の資化能力に ついて詳細 に検討 された。カラマツ根株心腐病菌 として,カ
イメ ンタケ P/Paθο′ws scIP餌一 」ιzガ),
レンゲタケ 乃sιデβ力β′sa〃θaハ
ナ ビラタケ 勲βttSSrs crデ甲2等
が分離 され, いずれ も褐色腐朽 を起 こす担子菌類であ り,単
糖類,ア
ラ ビノガラクタ ン,ヘ
ミセル ロー スを資化 していること,生
立木 の腐朽 は腐朽菌のみによって生 じるのではな く,非
担子糸 状菌の遷移 によって生 じる こと,
さらに,
これ ら糸状菌はカラマツ冷水抽 出物や腐朽材の 冷水抽 出物 を主 に資化 して いる ことが明 らかに された。 エノキタケ,
ヒラタケ,ヤ
ナギマツタケ 1『οθ力θじ/′Fだ用じθβの菌床栽培 では,培
地 基材 に針葉樹鋸屑 が用 い られ,い
ずれ もスギ材 の使用が奨 め られている (農耕 と園芸編集 部,1988)。
しか し,最
近 の鋸屑流通 の事情 は,各
地 での きの こ栽培 が振興 して きた こと によ り良質 の スギ鋸屑 の入手が困難 とな り,他
の針葉樹鋸屑 に依存せ ざるを得ない状況 で ある。 しか し:カ
ラマツ材 の鋸屑 を使用す るには,散
水堆積 の期 間 を十三 分に とる ことが-7-求め られ (柿木
,1993),カ
ラマツ鋸屑 はエプマツ・ トドマツに比べ散水堆積 の影響 を受 けに くく,鋸
屑 の改善が遅 れ る ことが指摘 されている (沓沢 ら,1980)。
このため,他
の 針葉樹鋸屑 に比べて,き
の こ菌床培地基材 としての適性は低 く評価 されている。カラマツ 鋸屑 が きの こ栽培 の培地基材 として適 さな いのは抗菌作用 を有す るフ ェノール性成分等 の 阻害成分 (いわゆ るアク)が
材 中に存在 し,ア
ク抜 きが難 しいためであるとされて いる。 カラマツ鋸屑 よ リアラ ビノガラクタ ンを製造するには,鋸
屑 を多量 の水 で抽 出処理 を行 うが,そ
の際に副次的に生 じるカラマツ鋸屑の水抽 出残造は,ア
ラ ビノガ ラクタ ンと共 に 水溶性 フェノール成分 も同時 に溶脱 されている。そ こで高 畠 らは,カ
ラマ ッ鋸屑 の水抽 出 残澄は1屋
外 で散水堆積処理 した鋸屑,い
わゆ るアク抜 き処理 した鋸屑 と同様 にきの こ菌 床栽培 の培地基材 として利用 できるのではな いか と想定 し,カ
ラマツ鋸屑 の水抽 出残澄 に つ いてエノキ タケ】 ヒラタケ,ヤ
ナギ マツタケに対 す る培地適性 を検討 した。その結果, カラマツ鋸屑の水抽出残澄 は未処理の鋸屑 よ りも子実体収量が多 くな り,ま
た,従
来 よ り 培地基材 として利用 されて いるスギ材 (辺 材)鋸
屑 に比べて・ 同等 もしくはそれ以上の子 実体収量 を示 した (高 畠 ら,1991;高
畠,1992)。
この ことか らiカ
ラマツ鋸屑 の水抽 出 残澄 は培地基材 として充分 に利用可能 な ことが明 らか とな ったが,同
時にカラマツ材のア クに相 当す るカラマツ水抽 出物 には抗菌成分の存在が推察 された。次 いで,カ
ラマツ水抽 出物 の抗菌性 を検討 した ところ,き
の こ栽培 の害菌である トリコデルマ菌類の成長 に対 し ては強 い阻害作用 を示 した。一方,食
用担子菌 の成長 に対 しては阻害作用が認 め られず, む しろ菌糸体成長 を促進 させ る傾 向が認 め られ た (高畠,1991)。
そ こで,
これ らの結果 に基づ いて,本
論文 では食用 きの こ栽培 にカ ラマツ水抽 出物 を成 長促進剤 として利用す る観点 か ら,食
用担子菌の菌糸体成長,子
実体形成 に及ぼすカラマ ツ水抽 出物 の影響 に関す る検討 を展開 した。第 Ⅱ草
数種食用担子菌の菌糸体成長 に及 ぼすカラマツ水抽 出物 の影響 これ までにア ラ ビノガラクタ ンの代謝 につ いて
,
ヒ ト糞便 によ リア ラ ビノガラクタ ンが 代謝 され低級脂肪酸 が産 出 され る こと(Englyst et al.,1987),
ヒ ト腸 内優先菌 である 肋cιb翔デJes οttι口dや
ラ ッ ト・ ヒ トの腸 内細菌であるβrr′J9肋ctθrデ閉 ′ongumん
ゞアラ ビノガラクタ ンを資化 して著 し く増殖す る こと (MaCfarlane・Cibson,1991i今
村 ら,19
92)が
報告 されている。 しか し,食
用担子菌類 に対 す るアラ ビノガラクタ ンの作用 は明 ら かにされていな い。 カラマツ材 由来 のアラ ビノガラクタ ンは材 の水抽 出液 を脱 フェノール処理 して得 られ る (水本 ・高橋,1987)の
で,水
抽 出物 にはアラ ビノガラクタ ンが多量 に合 まれている。本 章 では,本
邦産 (北海道産:長
野県産),シ
ベ リア産 と産地の異なるカラマツ材 の水抽出 物 につ いて,そ
の化学組成 とヒラタケ,エ
ノキ タケ,シ
イタケの食用担子菌の成育 に及ぼ す影響 を比較検討 した (高 畠・水本,1994)。
さらに,三
産地間で最 も水抽 出率 が高 いた め,水
抽 出物 の調製 が容易 な シベ リア産カラマツ材 の水抽 出物 につ いて前述の食用担子菌 三菌種 の外 にヤナギマツタケ:ナ
メコ,マ
イタケの三菌種 を追加 し1液
体 ・寒天 ・木粉培 地 と様 々な培地条件 で食用担子菌の成育 に及ばす影響 を検討 した (高 畠 ら11994)。
1.カ
ラマツ水抽 出物の化学組成 1。1
材料 お よび方法(1)カ
ラマツ水抽 出物 シベ リア産カ ラマツ (とβttχdP.)お
よび本邦産 カラマツ (とβr放 ′9pιOFeprs i】ヒ海道産 ・長野県産)の
各製材鋸屑 (気乾)に
,そ
れぞれ重量比で5倍量 の水 を加 え,室
温 にて撹拌 しなが ら30分 問抽 出 した。濾別 した抽 出液 を室温 にて減圧濃縮 し,130°Cの
ドラムで数分間 乾燥 して,粉
末状のカラマツ水抽 出物 を得 た。 (なお,以
下 では,シ
ベ リア,北
海道 お よ び長野県の各産地 のカラマツ材 よ り得 られた水抽出物 をそれぞれシベ リア産,北
海道産, 長野県産 と略称 す る。)製
材鋸屑 の気乾重量 に対す る水抽 出物 の収率 は,シ
ベ リア産で約-9-10%,北
海道産,長
野県産 は各 々約5%で
あった。(2)カ
ラマツ水抽 出物 の化学組成分析シベ リア
,北
海道,長
野県産 の各カ ラマツ水抽 出物 につ いて全糖量,中
性還 元糖 の組成,ウロン酸量
,全
フ ェノール量i分
子量分布 お よび重量平均分子量 を測定 した。全糖量はSomogyi一
Nelson法 (SomOgyi,1952)に
よって測定 し,全
還元糖量 (重 量比Calactose:Arabinose=3:22
基準)に
0,9を乗 じて求 めた。 ウロ ン酸量はカルバ ゾール ・硫酸法 (福井11969)に
よって,全
フ ェノール量 はFolin・Denis法
(Folin・ Denis,1915 i Swain・
Hills,1959)(Taxifolin基
準)に
よって測定 した。中性還 元糖 は,ポ
ス ト カラム方式 のHPLC(東
ソーKK製
,還
元糖分析 システ ム)に
よ り分析 した。すなわち,各単糖 を糖 ―ホ ウ酸 アニオ ン性錯化合物 としてか ら
,陰
イオ ン交換 カ ラム (東ソーKK製
,TSKgel, Sugar AXI)で
分離(Sinner et al.,1975)後
,α
―シアノアセトア ミ ドを反応 させ (Honda et al。
,1981),UV検
出器 (波長275nm)で
検 出 し,定
量 し た。分子量分布お よび重量平均分子量は
,GFCカ
ラム (東ソーKK製 ,TSKgeli G
4000P WX,G3000P WX L)を
用 い,50mMNa C l溶
液 を分離溶液 として試料 を溶 出 分離 し,RIお
よびUV(波
長280nm)検
出 して測定 した。なお,標
準物質 には,PuHul―
an(昭
和電工KK製
,Mw850000,380000,186000,100000,48000,23700,12200,580
0),Ma比
oheptaose(KK林
原製,Mwl153),Mattose(KK林
原製,Mw360),α
―Dextrose(和
光純薬KK製 ,Mw180),卜
Arabinose(和
光純薬KK製 ,Mw150)を
用 い た。1.2
結果 各産地のカラマ ツ水抽 出物 の全糖量
,中
性還 元糖量の組成 (以下,糖
組成 とす る。 ), ウロン酸量,全
フ ェノール量 を測定 し,そ
の結果 をTable Ⅱ-1に示す。 全糖量ではシベ リア産は,91.5%と
な り,北
海道産,長
野県産 の85∼
86%に
比べて若千 高 い値 を示 した。糖組成 では,北
海道産 でマ ンノースが微量認 め られ たが,シ
ベ リア産やTable Ⅱ一-1. The chemical composition of Karalllastu water extracts.
Sample T.Sugarl) Sugar Compo。 2) urOnic Acid3) T,pheno14)
(%)5) Man Ara Gal(%) (%)5) (%)5)
Siberia 91.5 -- 11,6 88.4 0.84 0.47
Hokkaido 85.8 0.8 10.7 88.5 0。 79 1.50 Nagano 85.1 -- 13.1 86.9 0.80 コ 1.58
1)Somogyi―Nelson method(Ara/Cal=3/22, 団OleCular weight ratio used as
the standard.)T.Sugar=T.reducing sugar X O.9.
2)IEC(colunn:TSKgel SugarAXl)
3)CarbazoletSulferic acid method(Glucronic Acid used as the standard.) 4)Folin・ denis lnethod(Taxifolin used as the standard.)
5)Percentage of the oven― dried veight.
-8.00
7.00
6.00
5.00
4.00
3.00
2.00
1.00
0.00
600
400
200
Fig. Ⅱ-1.
門 D E ] 田 E り 0 コ ︼ [ う と ] 29.0[TIHE]
39口 040.0
40山 0600
400
200
600
400
200
﹃ ⊃ E ]20,0 39.0
[TIHE]
GPC―chromatography of water extracts from 」apanese Siberian larch wood by RI.
Or
1、
、 Hv 2L江 00 Siberia
20.0
[TIHE]
30.0
︻ 〓 り O J ] [ ⊃ 〓 ︺8.00
7.00
6.00
5.00
4.00
3.00
2.00
1,00
0.00
120 100 80 60 Fig. Ⅱ-2.
Notes:20.0 3o.0
[TIHE]
ユ20 100 80 Sa 120 100 89 60 」apanese or reglons 40.0 4日.0 ﹃ ∋ 〓 ︺ [ ⊃ 〓 ]GPC―chromatography of water extracts from Siberian larch wood by UV.
Arrowheads indicate high molecular veight of phenolic compounds
Siberia
‖okkaido
-長野県産では認 め られなかった。 しか し, いずれの産地 もアラ ビノー スが11∼
13%,ガ
ラ ク トースが87∼89%と
な り,ほ
ぼ 同様 な糖組成 とな った。 ウロン酸量 は,
いずれの産地 も0,8%前
後 とな った。 この よ うに全糖量,糖
組成!
ウロン酸量につ いては,各
産地 間で顕著 な相違 は認 め られなかった。 しか し,全
フ ェノール量は, シベ リア産が0,47%で
あったの に対 し,北
海道産が1.50%:長
野県産 が1.58%と
な り,北
海道,長
野県産 は シベ リア産の 3.5倍余 りのフ ェノール合量 を示 した。 重量平均分子量,分
子量分布 を検討 した ところ,重
量平均分子量 は,シ
ベ リア産は18400,北
海道 産は18700と な りほぼ 同 じ分子量 であった。長野県産は21400と な り,シ
ベ リア産 や北海道産 よ りやや大 きな分子量 を示 した。分子量分布 は1い
ずれの産地 もほぼ 同様な分 布状態 を示 した (Fig.Ⅱ-1)。
重量平均分子量,全
糖量,糖
組成 の結果 か ら1い
ずれの産 地のカラマツ水抽 出 も主要構成成分は水溶性多糖類 のアラ ビノガラクタ ンであると考 え ら れ る。 水溶性 フ ェノール成分の分子量分布 を測定 した結果 をFig.コ ー2に
示 す。 シベ リア産 は, 本邦産 よ りも全分子量域 で波長280nmの 吸収が低 く,フ
ェノール成分の合有量が少 ないこと が示唆 され,Table
Ⅱ-1の
全 フ ェノール量の測定結果 と符合 した。北海道産 と長野県産は, 概ね 同様な分布傾 向を示 したが,北
海道産 では,高
分子量領域 での吸収が認 め られ,フ
ェ ノール類 の縮重合 した成分の存在が示唆 され た。2.産
地別 カ ラマ ツの水抽 出物 添 加培 地 におけ る菌 糸体成 長 の比較2.1
材料 及 び方法(1)カ
ラマ ツ水 抽 出物 前節「 カ ラマ ツ水抽 出物 の化学 組成 」 にお いて,調
製,供
試 した シベ リア産,北
海道 産, 長野県産 の各 カ ラマ ツ材 の水抽 出物 を供 試 した。(2)供
試菌 当セ ンター保存 菌株 で あ る以下 の3菌 株 を供試 した。ヒラタケ P′θ口rοι
"δ
οstrθ2をμs(」acq:Fr。
)Kummer.(Po-03)
エ ノ キ タ ケ 周 胸町pFrη β 79′μιや θ
S(Curt,:Fr.)Sing。 (Fv-4)
シイタケ と伊元JipEFd θ」9ぬs(Berk。
).(Le-01)
(3)供
試培地培地 には
,ポ
テ トデキ ス トロース培地(PD培
地,Difco製 )お
よび同寒天培地(PDA
培地,
日水製)を
供試 した。カラマツ水抽 出物 の添加量 はIW/V比
で1,2,3,5,10,
15%と
した。なお,接
種源 には,供
試菌 を麦芽 エキ ス・酵母 エキ ス寒天培地(MYA培
地;Glucose 4g, Malt extracts 10gi Veast extracts 4g, Agar 15gi Dis,Water lL)で 24 °
C,6∼
10日間平面培養 した後:コ
ルクボーラーで直径5mmに打ち抜 いたデ ィスクを用 いた。(4)菌
糸体成長量 菌糸体成長量 として,培
養菌糸体 の伸長量 と乾燥重量 を測定 した。伸長量は善本 らの方 法 (善本 ら,1984)に
準 じて測定 した。すなわち,直
径12mm,長
さ160mmの ライア ン管 にP
DA培
地 を1.8ml分 注 して菌糸体仲長量測定用培地 を調製 し,そ
の一端 に接種源 を接種 した 後,24℃
にて7日 間培養 して菌糸体仲長量 を測定 した。 菌糸体乾燥重量は,次
の方法 で求めた。すなわち,50ml容
三角 フラスコにPD培
地 を5 ml分注 して菌糸体乾燥重量測定用培地 を調製 し,120°C,15分
間の条件 で殺菌 した後,各
供 試菌 を接種 し,24℃
にて7日 間静置培養 した。その後,菌
糸体 を濾別 し,温
水,冷
水 で順次 洗浄後:菌
糸体 の乾燥重量 を測定 した。各試験 区 とも5検体づつ供試 し,繰
り返 しを行 った。2.2
結果
(1)菌
糸体伸長成長 各産地のカラマツ水抽 出物 にていて,水
抽 出物濃度 の異 な るPDA培
地 で ヒラタケ,エ
ノキ タケ,シ
イタケの菌糸体 を培養 しi菌
糸体仲長量 を測定 した結果 をFig.Ⅱ-3∼
5に示 す。 ヒラタケに関 して,シ
ベ リア産 では添加濃度 を1∼3%と
した場合,対
照 区よ り若千良好 な伸長量 を示 した。2%の
添加濃度 で最大値 とな り,対
照 区の1.2倍 の伸長量 を示 した。 し15
-P.oStreatus
40
20
︵ 沢 ︶ ∽ や o 、 L や ※ 口 ︼ ① や 嶋 避 ″ O ① ℃ 0 く Siberia Hokkaido Nagano吻
国
圏
40
Growth(mm)
Relationships betveen added amount of water extracts from Japanese or Siberian larch wood in PDA medium and mycelial growth of P′ θEPrOtWs Ostrθaをvs incubated for seven days.
: The horizonal bars represent standard deviations.
20
Mycelial60
Fig. Ⅱ-3.
:・ :・ :・ !・::::::::::・・ :・..:・ . NotesF.vetutipes
40
20
︵ 沢 ︶ ∽ や り ヽ 彎 や x 口 L ① や ∝ 鮮 O ① 巧 ▼ < 物 i Sibctta 圏 :‖ 。kka閲。 ピ【径田: Nagano20
Mycelial40
Crowth(mm)
Fig.Ⅱ
-4
Relationships between added aⅢ ount of water extracts from Japanese or Siberian larch wood in PDA medium and mycelial growth of F′ 曖□nE「′丁Л2 7θ′どをIPθS incubated for seven days.The horizonal bars represent standard deviations。
・ :・ :・ :・ !・ :・ !・ :・ :・ :・ !・:・:・ !・ :・ :・ :・ :・ !・ :・・ ...● : :・ :・ :・:c!・!:!::::::::::::i::::::::::::° .。 .. ・ :・ :・ !・ :・ :・ :・ ..。 ..: !・ :・ !・ :・ !・ :・ !・ :・・ .. Notes:
17
-10
Mycelial
L.edodes
20 30
Growth(mm)
20
︵ 誦氏 ︶ 2 3 5 ︲0 ∽ 中 υ d 監 中 瑣 口 噴 0 中 § > 巧 ① つ づ く物
囲
Siber,a ‖okkaido 足電臣]: Nagano
15Fig, Ⅱ
-5.
Relationships between added amount of water extracts froШ 」apanese or Siberian larch wood in PDA medium and mycelial growth of とentFコys θ」oJ9s incubated for seven days. The horizonal bars represent standard deviations.::!士:!::::卜:・ :・ :・ :・ :・ :・ :・ :・ :・ :・ :・ :・
静
:・:i:!・・ .・.・ :・ i・ :・ :・ :・ :・ :・ :・ :・ :・ :・ :・ :・ :・ :・ :・ I・
か し
,添
加濃度 を10%以
上 にす ると,伸
長量 は対照 区よ りも低下 し,15%濃
度 では対照 区 のO.9倍 とな った。北海道産 お よび長野県産 は,
ともに添加量が増加す るに従 い!伸
長量は 低下す る傾 向を示 し,低
下 の程度 は北海道産が顕著 であった。15%の
添加濃度 では,北
海 道産は対照 区のO.5倍 とな り,長
野県産 は対照 区のO.8倍 とな った (Fig.Ⅱ-3)。
エノキタケに関 して
,シ
ベ リア産,長
野県産 ともに1∼5%の
添加濃度 で伸長量は1対
照 区に比べて良好 とな った。伸長量 の最大値 を示す添加濃度 はIシ
ベ リア産 では2∼5%で
あ り,対
照 区の1.3倍 とな った。長野県産 では1%で
あ り,対
照 区の1.5倍 とな った。 しか し,10%あ
るいは15%の
添加濃度 では,対
照 区 と同等 もし くは,そ
れ よ り劣 った伸長量 とな っ た。北海道産 は,添
加量が増加す るに従 い伸長量は著 し く低下 し,10%の
添加濃度 で対照 区のO.5倍,15%の
添加濃度 で0。4倍 とな った (Fig.Ⅱ-4)。
シイ タケに関 して,シ
ベ リア産は,1%の
添加濃度 で対照 区よ りやや良好 とな ったがi添
加濃度 を2%以
上 とした場合,添
加量が増加す るに従 い,伸
長量 は低下 し,15%の
添加濃度 では対照 区の0.6倍 とな った。本邦産の北海道,長
野県産 は,添
加量 が増加す るに従 い,伸
長量は低下 し,低
下 の度合 いは北海道産 が著 し く,10%の
添加濃度 で対照 区の0.9倍,15%
の添加濃度 で0。85倍 とな った (Fig.Ⅱ-5)。
菌糸体伸長量 においてiシ
ベ リア産は,い
ずれの供試菌 に対 して も1∼2%の
添加濃度 で, 対照 区の1,1∼ 1.3倍 の伸長量 とな り,若
千 の促進作用 を示 したが,10%以
上の添加濃度 で は,対
照 区 と同程度以下 の伸長量 とな った。北海道産は,供
試 した全 ての菌種 で添加濃度 が高 くなるに従 い,伸
長量 は低下 し,10%あ
るいは15%の
添加濃度 では,対
照 区の1/2∼ 1/3の伸長量 に抑制 された。長野県産 は,エ
ノキ タケにお いて1∼5%の
添加濃度 で若干の促 進作用 を示 したが,10%以
上 の添加濃度 では,対
照 区よ り低下 した。 ヒラタケ,シ
イタケ では,北
海道産 と同様 に添加量が増加す るに従 い,伸
長量は低下 し,い
ずれ も15%の
添加 濃度 では対照 区のO.8倍 前後 とな った。(2)菌
糸体重量成長 各産地のカラマツ水抽 出物 につ いて,添
加濃度の異な るPD培
地 で ヒラタケ,エ
ノキ タ-19-ケンイタケ菌糸体 を培養 し
,菌
糸体重量 を沢J定 した結果 を Fig.Ⅱ-6∼
8に示 す。 ヒラタケに関 して,い
ずれの産地 のカ ラマツ水抽 出物 も添加量 が増加す るに従 い,菌
糸 体重量は顕者 に増加 した。1∼3%の
添加濃度 では,産
地間で差が生ぜず,3%の
添加濃度 で, いずれの産地 も対照 区の2倍程度 の菌糸体重量 を示 した。添加濃度 を5%以
上 とした場合, 添加濃度が高 くな るに従 い,産
地 間で差 が認 め られ,15%の
添加濃度 では,シ
ベ リア産は 対照 区の7.5倍i北
海道産,長
野県産 は4.5倍 前後 とな った (Fig.Ⅱ-6)。
エノキタケに関 して,
ヒラタケ同様 に,い
ずれの産地 も添加濃度が高 くなるに従 い,菌
糸体重量は増加 した。1∼3%の
添加濃度 では,産
地 間で差が生ぜず,3%の
添加濃度 で対照 区の1.5∼1.9倍 の菌糸体重量 を示 した。5%以
上の添加濃度 では1産
地 間で差 が生 じ,そ
の 差 は,添
加濃度 が高 くな るに従 い大 き くな った。15%の
添加濃度 では,シ
ベ リア産,長
野 県産は対照 区の4.6倍 の菌糸体重量 を示 したのに対 し,北
海道産は対照 区の2倍程度 に とど まった (Fig.Ⅱ-7)。
シイタケに関 して,い
ずれの産地 も添加量が増加す るに従 い菌糸体重量は増加 した。1∼3%の
添加濃度 では産地 間で差 が生ぜず,3%の
添加濃度 で対照 区の3∼4倍 の菌糸体重量 を 示 した。 しか し,5%以
上の添加濃度 では,産
地間に差 が生 じ:そ
の差 は濃度が高 くなるに 従 い大 き くな った。15%の
添加濃度 では長野県産が対照 区の約9倍の菌糸体重量 を示 したの に対 し,シ
ベ リア産,北
海道産 は5倍余 りとな った (Fig.コ ー8)。 供試 した3菌種全 てに対 して,シ
ベ リア産および本邦産 ともに添加濃度が高 くな るに従 い 菌糸体重量 は顕著 に増加 した。1∼3%の
範 囲ではI産
地 間に差 は生 じなか ったが,5%以
上 の添加濃度 では菌種 に応 じて産地間に差が生 じた。すなわち,
ヒラタケではシベ リア産, エノキタケではシベ リア産および長野県産,シ
イタケでは長野県産 のカ ラマツ水抽 出物 は 各 々の菌糸体の重量成長 に対 して顕著 な促進作用 を示 した。P.ost■
eatus
100
150
0 1 ︵ で 代 ︶%
□
鯉
Siberia ‖okkaido Nagano 2 3 5 ︲0 ∽ や O 礎 L ゛ X 口 L ① や 、 垂 r づ ① つ ℃ < 1550
Mycelial100 150 200
Dry Veight(mg/fiask)
250
Fig. Ⅱ
-6.
Relationships between added alllount of water extracts froⅢ 」apanese or Siberian larch wood in PD llledium and mycelial dry weight of P′ θErrOιrrs οstrθβιE「S inCubated for seven days.: The horizonal bars represent standard deviations.
・ :・ :・ :・ :・ :・ :・ :・ :・ !・ :・ !・::::::!:!:::!:::::!:!:::!:::・ :・ :・:
・ :・ :・ :・ :・ :・ :・:・:・ :・ :・ :・ :・ :・ :・ :・ !・・
・ :・・・・ .・ .・ :・ :・ :・:
Notes
-F.velutipes
60
40
1 2 3 5 10 ︵ 沢 ︶ ∽ 中 ω 僣 L や x 口 L O や ヽ 甚 ガ O ① 巧 巧 <15
20
Mycetial
n u R U40
80 100
(mg/flask)
Fig. Ⅱ―-7.Dry Weight
Relationships betveen added amount of water extracts from 」apanese or Siberian larch wood in PD medium and mycelial dry weight of F′ じこ騒rr′デコβ 7θ′rrtlipθs incubated for seven days.
The horizonal bars represent standard deviations.
%i Sibetta
□ :HOkkaid。
EΣ憂】: Nagano
L.edodes
20 30 40
1 2 3 5 ︲0 ︵ 沢 ︶ ∽ ↓ o § ﹄ 中 X 国 ﹄ 0 や 僣 善 芍 ω O つ く 15 Mycelial30 40 50
Dry weight(mg/fiask)
Fig. Ⅱ
-8.
Relationships between added amount of water extracts from Japanese or Siberian larch wood in PD medium and mycelial dry weight of とentFコEPs θ」OJOS incubated for seven days.The horizOnal bars represent standard deviations.
囲 :‖。kkaid。
Notes I
-3.シ
ベ リア産 カ ラマツの水抽 出物添加培地 におけ る菌糸体成長3.1
材料及 び方法(1)シ
ベ リアカ ラマツ水抽 出物 シベ リア カ ラマ ツ (とβrrχsP,)の
製材 鋸屑 (気乾 状 態)に
重 量比 で5倍 量 の水 を加 え, 室温 にて撹 拌 しなが ら30分間抽 出 した。濾別 した抽 出液 を室温 で減圧 濃縮 し,130℃
の ドラ ム中で数分 間乾燥 して粉 末状 の カ ラマ ツ水抽 出物 (以下,KWEと
略 す る。)を
得 た。使 用 した鋸屑 の粒度 分布 はi気
乾 重量比 で0∼12メ ッシュの ものが6%,12-48メ
ッシュが72%,48-100メ
ッシュが19%,100メ
ッシュ以■ が3%で
あ った。(2)供
試菌 当セ ンター保存 菌株 で あ る以下 の6菌 株 を供試 した。 ヒラタケ P′ θprοtps θdιrθttμs(」acq:Fr.)Kummer。(Po-03) _
エノキタケ デ【鞭 w′Fpa yθ力ιゎθs(Curt.:Fr。 )Sing。(Fv-4)
ヤナギマツタケ 】grοcヵθじ/′Jir2Jr2cθβθ(DC.:Fr。
)Maire.(Ac-01)
シイタケ と94tゴ刀胚 θど0」9(Berk.)。(Le-01)
ナメコpヵ
rο tβ コ溜〃θ々0(T.Ito)S.Ito et lmai。(Pn-03)
マイタケ fttο乃翔 rrOr2的s2(Dicks,:Fr.)S,F,Gray.(Gf-01)
(3)供
試培地 培地 には,ポ
テ トデキ ス トロース液体培地(PD培
地 ;馬鈴薯200g,グ
ル コース20g,
蒸留水lL):20m1/100ml容
三角 フラスコ,同
寒天培地(PDA培
地 ;日水製):20m1/直
径90mmペ トリ皿お よび木粉培地 :16g/直径18mm試 験管 を用 いた。木粉培地 は木粉 :米糠=3:1(全
乾重量比)と
し,合
水率65%(湿
量基準)に
調整 した。なお ヒラタケ,エ
ノキ タケ, ヤナギマツタケにはスギ辺材 を:シ
イタケ,ナ
メコ,マ
イタケにはブナ材 の木粉 をそれぞ れ用 いた。KWEの
添加量はPD,PDA培
地 ではW/V比
で,木
粉培地 にお いては,培
地 の全乾重量当た りのW/V比
で各 々1,2,3,5,10,15%と
した。 なお,接
種源 には,供
試菌 を酵母 エキ ス・麦芽エキ ス寒天培地(MYA培
地)で
24°C,6∼14日 間平面培養 した後
,コ
ル クボーラーで直径5mmに打 ち抜 いたデ ィスクを用 いた。(4)菌
糸体成長量 培養はすべて 24°C暗
黒下 で行 った。PD培
地 では,10日
間培養 した後 菌糸体 を濾別 し, 温水,冷
水 で順次洗浄後,菌
糸体 の乾燥重量 を測定 した。PDA培
地 では,6日
間培養 した 後 コロニー直径 を沢!定した。 さ らに,4日
間培養後,電
子 レンジを用 いて培地 を溶解 し菌糸 体 を濾別,温
水冷水 で1贋次洗浄後,菌
糸体の乾燥重量 を測定 した。木粉培地 では2∼5日 間 の前培養 を行 い,そ
の後20日 間の菌糸体伸長量 を沢」定 した。各試験 とも5検体づつ供試 し, 繰 り返 しを行 った。3.2
結果
(1)菌
糸体仲長量KWE添
加濃度 の異な るPDA培
地 で培養 した各種供試菌 の菌糸体伸長量 をFig.コ ー9に 示す。KWEの
添加 によ り,供
試 した6菌種全 てにお いて菌糸体伸長量 は対照 区に比べて若 千増加 し,1∼3%の
添加濃度 で最大伸長量 を示 した。最 も菌糸体伸長量が大 きか ったのは ヒラタケで,2%の
添加濃度 で最大伸長量(81.4mm)を
示 し,対
照 区(69.6mm)の
1.17倍 で あった。 しか し,い
ずれの供試菌 も添加濃度が高 まるにつれて菌糸体伸長量は低下 した。 添加濃度が15%の
場合,マ
イ タケ を除 いた全 ての供試菌 にお いて対照 区よ りも低下 し,
ヒ ラタケ,ナ
メコ,ヤ
ナギマ ツタケでは対照 区のO.8∼ 0.85倍,さ
らにエノキ タケ とシイタケ では0.5∼0。6倍 の菌糸体伸長量 となった。KWEの
添加濃度 の異 な る木粉培地 で培養 した各種供試菌の菌糸体伸長量 をFig.Ⅱ-10
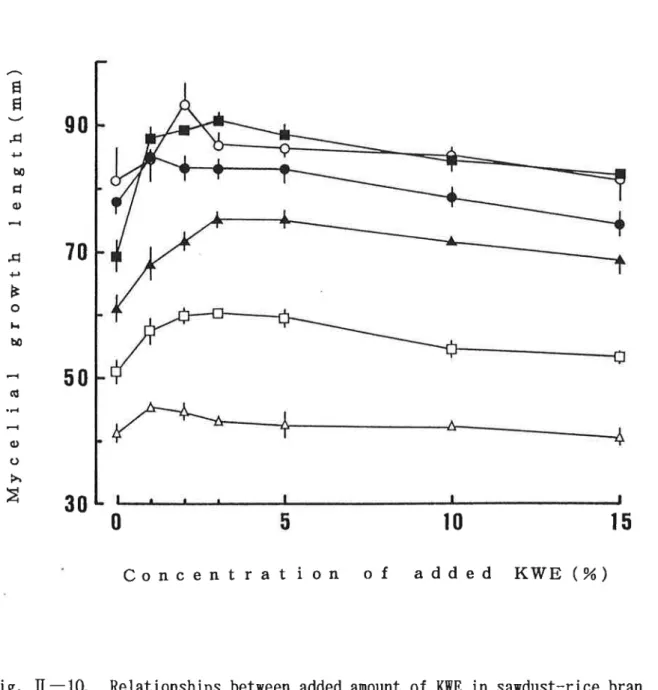
に示す。木粉培地 での菌糸体伸長量は供試 した6菌種全 てにおいて1∼3%の
添加濃度 で最大 伸長量 を示 し,添
加濃度が高 くな るにつれて漸減 したが,15%の
添加濃度 で も対照 区 とほ ぼ同程度 またはそれ以上の菌糸体伸長が認 め られた。(2)菌
糸体重量KWE添
加濃度 の異な るPDA培
地 で培養 した各種供試菌の菌糸体重量 をFig.Ⅱ ―llに 示す。供試 した全 ての菌種 でKWEを
添加す る ことによ り菌糸体重量が増加 した。すなわ-90
0 6 ︵ 日 日 ︶ h O 一 0 日 “ ︼ 巧 ヽ 日 0 ﹁ O O30
Concentration of
10 15
added KWE(%)
Figo Ⅱ-9. Relationships between added aⅢ ount of KWE(water extracts from Siberian larch wood)in PDA medium and colony diameter of edible
mushrooms incubated incubated for six days.
Legends:():HIRATAKE (P.Ostrθ βιps)・ ● : ENOKITAKE (F.79Frrtlipθ
s),
△ :YANAGIMATSUTAKE(ズ.σ/FデηJr2じθコθ
), A :SHIITAKE(と
.θ」οJ9s),□ :NAMEKO(P.″β〃94o)・ ││ :MAITAKE(f。 こ
{機
沈石』).
︵ 日 日 ︶ 目 , ] 目 0 引 劇 , 卜 o ﹄ ︼ 引 ω ︼ 引 o o 、 Σ
90
n u η f50
30
0
10
15
Concentration of added KWE(%)
Fig. Ⅱ―-10. Relationships between added amount of KWE in sawdust― rice bran medium and lllycelial growth length of edible mushrooms incubated
incubated for twenty days.
Legends:O :HIRATAKE (P,Ostrθ βιErS)・ ● : ENOKITAKE (F.7θ ′ErtFpθ
s),
△ :YANAGIMATSUTAKE(】.σ /′ fコJraθθ』θ
), A:SHIITAKE(と
.θ」οJ9s),□ :NAMEKO(P.,β〃
eko),
日│ :MAITAKE(♂.rr92」οsa).
Note: The vertical bars represent standard deviation.
-ち
,KWE添
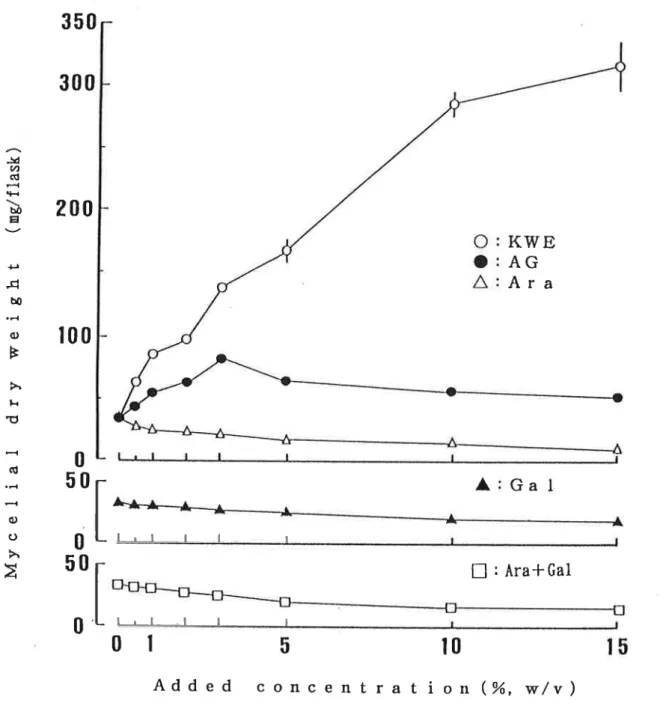
加濃度 と菌糸体重量 の増加 との関係は,菌
種 によって異 な っているが,5∼
10%の
添加 によ り,ナ
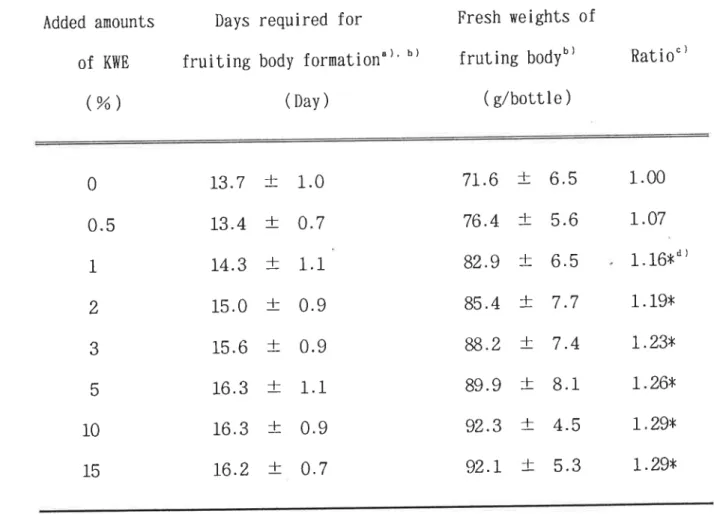
メコ以外 のすべての菌種 で対照 区の2倍 以上の菌糸体重量 を示 した。 ヒラタケでは10%の
添加濃度 で最高値(470.9mg/plate)を 示 し,対
照 区 (137.Omg/plate) の3,4倍 の成長量 を示 した。 しか し,15%の
添加濃度 では,対
照 区のO.7倍 に激減 した。ナ メコではKWE添
加 による菌糸体重量は,最
大 で対照 区の1.3倍に とどま り,
ヒラタケ同様15%の
添加では,対
照区よ り低下 した。KWE添
加濃度 の異な るPD培
地 で培養 した各種供試菌の菌糸体重量 をFig.Ⅱ-12に
示す。 いずれの供試菌 において も,KWEを
添加す ることに よ り菌糸体重量は増加 した。KWE
添加濃度 と菌糸体重量増加 との関係は,菌
種 によって異 な った。 しか し:全
ての供試菌で 5∼15%の
添加濃度が最適濃度 とな り]対
照 区の2倍 以上の菌糸体重量 を示 した。 ヒラタケ ではl PDA培
地 と異な りKWEの
添加濃度 が15%(185皿 g/flask)で
あって も10%の
添加 濃度 (187mg/flask)と 同程度 の菌糸休重量 を示 した。 また,シ
イ タケではPD培
地 におい て15%が
最適添加濃度 とな り,そ
の他 の供試菌 においてもPD培
地 ではPDA培
地 と異 な り15%の
添加濃度 であって も菌糸体重量 は著 し く減少 しなか った。 これ らの結果 は,KW
Eの
飽和濃度 に近 い15%の
添加濃度 ではI寒
天の有無 による培地 の物性条件 の相違 が菌糸 体成長 に影響す る ことを示唆 して いる。4.
考 察 シベ リア産お よび本邦産 (北海道産,長
野県産)と
供試 した産地 の異 な るカラマツ材 の 水抽 出物 にはI共
通 してア ラ ビノガラクタ ンが主要構成成分 として合 まれ ていた。各 々の 水抽 出物 におけ る全糖量i中
性還 元糖 の糖組成,ウ
ロン酸量に著 しい相違が認 め られなか った ことか ら,水
抽 出物 に 占めるアラ ビノガ ラクタ ンの割合は,産
地 間で大 きな相違 はな いと考 えられ る。 しか し,水
抽 出物 の収率が シベ リア産 で約10%,本
邦産が約5%で
あるこ とか ら,シ
ベ リア産カラマツ材 におけるアラ ビノガラクタ ンの合有率はI本
邦産 カ ラマツ 材 の2倍以上であると考 え られ る。カ ラマツ材 のアラ ビノガラクタ ンの合有率 は,カ
ラマツC 駕 ■ ヽ こ P 声 ] ︼ o 卜 ゝ H 巧 引 ω ︼ 引 o o ヽ 周
400
300
200
100
ConcentratiOn of added KWE(%)
Fig. Ⅱ―-11. Relationships between added amount of KWE in PDA medium and Шycelial dry weight of edible mushrooms incubated incubated for ten dayst
Legends i O:HIRATAKE(P.Ostr92ι
EPs),
● :ENOKITAKE(F.79′Ertwipes)△ :YANAGIMATSUTAKE(Л .りFFЛJr2θθβθ), A I SHIITAKE (と .θ」οJ9♂), □ :NAMEKO(P.コβ〃
94o), ││:MAITAKE(♂
.i〔饂 」9sコ).
Note: The vertical bars represent standard deviation.-40o
奮 器 E ヽ こ “ 目 ∞ ・出 0 卜 ゝ H 巧 引 ω ・H ﹁ o o ゝ Σ300
200
100
Concentr
10 15
ion Of added KWE(%)
Fig. Ⅱ―-12. Relationships between added amount of KWE in PD medium and mycelial dry weight of edible mushrooms incubated incubated for ten days.
Legends:O:HIRATAKE(P.Os材
留 ιrd)・●
:ENOKITAKE(F.79カ
ιlipθs)△ :Y△NAGIMATSUTAKE(】.θ /′ fЛJraσθβθ
), A :SHIITAKE(と
.θ」οJ9s),□ :N△MEKO(P.コβ〃θb), 1日 :MAITAKE(fぅ とて甑 」ο
s2).
Note l The vertical bars represent standard deviation.
0 ´ R v t a
材の樹種
,樹
齢,年
輸幅,樹
高 に よって異 な る (C6tёet al.11966,1967;C6tё
・Time―H,1967)が ,本
実験 におけ るアラ ビノガラクタ ンの合有率の相違 は,樹
種 による違 いが 大 きな要因であると推察 され る。 カラマツ水抽 出物 中のフェノール成分の合有量は,シ
ベ リア産 と本邦産では大 き く異な り,本
邦産 にはシベ リア産の3.5倍 余 りのフ ェノール成分が合 まれていた。 また,北
海道産 のフ ェノール成分 には長野県産 には見 られない高分子量の成分が存在 していた。一般 に樹 木抽 出成分 には,そ
の樹種 固有の代謝産物が合 まれてお り (今村 ら,1983),特
に樹皮や 材の温水抽 出物 ・酢酸エチル抽 出物 ・タ ンエ ン類 は,材
を識別す る上 で指標 とな る ことが 報告 されている (岸 本 ら,1979,1981)。
したが って,カ
ラマツ材 の樹種,生
育立地条件 の違 いは,微
量成分 である水溶性 フェノール成分の形成 に影響 を及ば して いるもの と考 え られ る。フ シベ リア産
,北
海道産,長
野県産の いずれの産地 にお いて も,
ヒラタケiエ
ノキ タケ〕 シイタケ と供試 した3菌種 に対 してカラマツ水抽 出物 を添加す るに したが い,菌
糸体重量は 増加 し,特
に5∼15%の
添加濃度 で顕著 な増加 を示 した。一方:菌
糸体伸長量 は10%以
上の 添加濃度では,供
試 した3菌種すべてにおいて抑制 された。 この ように10%以
上の添加濃度 では,菌
糸体 の伸長成長 と重量成長に対 して全 く逆 の作用 を示 した。食用 きの この子実体 発生 には,ホ
ダ木 や培養基 内の菌体量が関与 し,菌
体量が多 いほ ど子実体発生量 が増 える ことが知 られている (Tokimoto・Fukuda,1981,Ohga,1990a)。
また,木
粉培地 の伸長 試験 と子実体形成試験 では,両
者 の結果 は必 ず しも一致 しな い ことが指摘 されて いる (有 田 ら,1969)。
これ らの ことは,菌
糸体 の総成長量 を現す菌糸体重量 は伸長成長量 よ りも 子実体形成 の状況 を反映 している ことを示唆 している。 したが って,い
ずれの産地 のカラ マツ水抽出物 も子実体形成 を促す ことが期待 できる。 この点 については1第
V章
で検討 を イ〒う。 菌糸体重量成長 に対 して,1∼3%の
添加濃度 では産地間で成長量 に差 が生 じなか ったが,31
-5%以
上の添加濃度 では産地間で著 しい差 が生 じた。 いずれのカラマツ水抽 出物 も主要構成 成分がアラ ビノガ ラクタ ンであ り,抽
出物 中の糖関連成分はほぼ 同様 な傾 向を示 した こと か ら,産
地 間におけ る添加効果 の差 には,糖
関連成分以外 の物質 の関与 が推察 され る。 シベ リア産カ ラマツ水抽 出物 は,
ヒラタケiエ
ノキ タケiヤ
ナギマツタケ,シ
イタケ! ナメコ,マ
イタケ と供試 した6菌 種全 ての食用担子菌 に対 して, PD培
地,PDA培
地のい ずれの培地 において も菌糸体重量成長 に顕著 な促進作用 を示 した。カ ラマツ内樹皮 の温水 抽 出物 は,エ
ノキ タケ,ナ
メコ,
ヒラタケ,キ
クラゲ,ア
ラゲキ クラゲに対 して顕著 な成 育抑制作用 を示す こと (善木 ら,1984)か
ら,カ
ラマツ材 の水抽 出物 は, 内樹皮温水抽 出 物 とは化学組成が異 な ることが示唆 され る。 スギ,
ヒノキ,ア
カマツの 内樹皮温水抽 出物 は,キ
クラゲ,シ
イタケの成育には促進作 用 を示すが,
ヒラタケに対 しては成育抑制作用 を示 している (善本,1984)。 。 また,
シイ ノキ辺材部 の熱水抽 出物 はシイタケには成育促進作用 を示すが,ナ
メコ,
ヒラタケに対 し ては成育抑制作用 を示 している (松尾,1957)。
一般 に,樹
木心材 の温水抽 出物 は,食
用 担子菌の成長 に対 して抑制作用 を示す場合が多 く,例
外的にブナ心材がナメコ,
ヒラタケ に対 して,
ミズナラ心材が シイ タケに促進作用 を示 す ことが報告 されて いる (高畠,1987, 1989)。 カラマツ材 の製材鋸屑 は大部分が心材か ら生 じていることを考慮 に入れ ると, シ ベ リア産カラマツ水抽出物 が供試 した6菌種全 ての食用担子菌に対 して顕者な促進作用 を示 す ことは,樹
木心材 の水抽 出物 として極 めて特異的 であ り,カ
ラマツ水抽 出物 には[特
定 菌種 に限定 され る ことのな い,広
範 囲な担子菌 の菌種 に共通 して作用す る成育促進成分の 存在が示唆 され る。 5。結 論 シベ リア産
,北
海道産,長
野県産 と産地 の異 な るカ ラマツ材水抽 出物 につ いて,化
学組 成 を検討 した ところ,い
ずれ も主要構成成分 は共通 してアラ ビノガラクタ ンであ り,糖
関 連成分 は産地間で顕著な差 が認 め られなか った。 しか し,全
フェノール量 は,本
邦産 (北海道産
,長
野県産)で
は,シ
ベ リア産の約3.5倍 の合有量 を示 した。三産地のカラマツ水抽 出物 につ いて:
ヒラタケ,エ
ノキ タケ,シ
イタケと三菌種 の食用担子菌の菌糸体成長 に及 ぼす影響 を検討 した ところ,い
ずれのカラマツ水抽 出物 も供試 した三菌種全 てに対 して添 加量が増加す るに したが い菌糸体重量は顕者 に増加 した。1∼3%の
添加濃度 では産地 間で 添加効果 に差 が生 じなか ったが,5%以
上の添加濃度 では菌種 に応 じて産地間で差 が生 じた。 シベ リア産カラマツ水抽 出物 につ いて,
さ らにヤナギマツタケ,ナ
メコ,マ
イタケ と対象 菌種 を増や して食用担子菌 の菌糸体成長 に及ぼす影響 を検討 した ところ,全
ての菌種 で寒 天,液
体,木
粉 と物理条件 の異 な るいずれの培地 にお いて も,菌
糸体成長 に顕著 な促進作 用 を示 した。第 Ⅲ草 ヒラタケ菌 によるカ ラマ ツ水抽 出物 の資化性 と菌体外 酵素 の産 出
前章 にお いて
,カ
ラマ ツ水抽 出物 は ヒラタケ,エ
ノキ タケ,シ
イ タケ等 の有用 な食 用担 子 菌 の菌糸体成 長 に顕著 な促進作 用 を示 す ことを見 いだ した。 また,カ
ラマ ツ水抽 出物 の 主要 構成成 分 は,水
溶性 多糖 類 の ア ラ ビノガ ラク タ ンで あ る ことを明 らか に した。シイ タケ (吉田 ら