弘 前 医 学 43:312‑327,1992
鶏腿動脈管の微細構造
第一報 動脈管の組織発生
高 橋 フ亡
抄鼻 筋卵4日か ら20日までの ニ ワ ト1) (白色 レグホン)の動脈管,大動脈及 び肺動脈 を光学 お よび電子顧徴鏡 的に観察 し,動脈管 の組織発生を検討 した. ニ ワ トリの大動脈 は生理的 に右側大動脈 弓であ り,左右 の動脈管 は 発生の全期間を通 じて存続 して肺動脈 と大動脈 を短絡す る.肺動脈 か ら大動脈 までの動脈管 の走行 は極めて長 い が,その遠位 (大動脈側)約半分は発生学的 に背側大動脈 に由来 してお り,大動脈 や肺動脈幹 と同様 な弾性型動 脈 に分化す る. これに対 し動脈管 の近位 (肺動脈側)半分は第6鯉 弓動脈 に由来 し,典型的 な筋性動脈 に分化す る.筋卵9日目には動脈管壁 の弾性線維層 は肺動脈 の分岐部直下 か ら急激 に減少 してお り,動脈管近位部の構造 は隣接す る大血管 とは著 し く異 なる.すでに貯卵14日目 (筋化 6日前)で大動脈や肺動脈 に先 ん じて動脈管近位 部 には良 く分化 した大型 の中膜平滑筋細胞 が発達 してお り,動脈管壁 の構成要素 (内皮細胞 と内皮下層 か ら成 る 内膜,平滑筋細胞 を主体 とした中膜,線維芽細胞 と細胞外基質や線維成分か ら成 る外膜)の形態分化 はほは完成 し,以後僻化時の形態変化 が起 こるまでその構造 に著変 はない.動脈管近位部 には肱発生の全期間を通 じて神経 線維や栄養血管は観察 されない.
弘前医学 43:312‑327,1992 Keywords:ductusarteriosus histogenesis
chick electronmicroscopy
FINESTRtJCTtJREOFTHEDtJCTtJSARTERIOStJSOFTHECHICE l.HLstogenesisof theDuctusArteriosus
GENTAKAHASHI
Abstract Histogenesisoftheductiarteriosiofthechickembryosfrom 4thto20thdayofincubationwas investigatedbylightandelectronmicroscopy.Asearlyas14thdayofincubation,theproximalhalfofthe ductusarteriosusembryologicallyderivedfrom the6thaorticarchhasitsalmostdefinitivestructureofa musculararterycharacterizedbyprecociousdevelopmentofthemedialmusculature,incontrasttothe distalhalfoftheductusderivedfrom thedorsalaorta,whichisathick‑walledarteryoftheelastictype. Theductallumenissurroundedbycuboidaloraattenedendothelialcellswithoutfenestration. Inthe subendotheliallayerasmallnumberofmicrofibrilsandcollagenfibrilsareembeddedinfinelygranularor homogeneousgroundsubstance. Thetunicamediaconsistsoftheinnerandouterzoneswithoutsharp boundarybetweenthem,eachofwhichhas3or4layersofsmoothmusclecellswithvariousamountof connectivetissuesamongthem. Mostofthecytoplasm oflarge,fullydifferentiatedsmoothmusclecells oftheinnercircularzoneisoccupiedbyabundleofmyo丘lamentca60Aindiameter.Smoothmusclecells oftheouterzonearesmallerinsize,lessdifferentiatedthanthoseoftheinnerzone,thecytoplasm which ischaracterizedbyprominentGolgi‑complexandwell‑developedrough‑surfacedendoplasmicreticulum.
Theysecretetheextracellularconnectivetissuecomponents(collagenfibrils,elasticfibersandlnicrofibrils embeddedinthegroundsubstance)inthetunicamediabeforehatchinginpreparationfortheanatomical closureoftheductfollowingtheinitialfunctionalclosureduringhatching.
HirosakiMed.∫.43:312‑327,1992 弘前大学 医学 部解剖学 第二 講座 (主任 加地 隆教
撹)
平成3年12月5日受付
TheSecond DepartmentofAnatomy,Hirosaki UniversitySchoolofMedicine(Director:Prof.T.
KACHI),Hirosaki,Japan
Receivedforpublication,December5,1991
平 成 4年 3月 弘前 医学 43巻 4号
は じ め に
BoTALLO氏動 脈管 は肺動脈 と大動脈 との 間に介在 し,肺循環の未発達 な胎生期 におい て右心、室か らの血流 の大部分を体循環系へ導 く短絡路 としての役割 を果た して,出生後 は 肺 呼 吸 の開 始 と肺 循 環 の発達 に伴 って閉鎖 し,次第 に動脈管索 に退化す る.晴乳類 に於 ける出生後 の動脈管 の閉鎖機構 に関 しては多 くの説が唱 え られて きたが,括約筋様 の中膜ド 平滑筋の収縮,内膜増殖等 の出生前 に設定 さ2) れ た管 壁 自体 の形 態 変化 が重 要 視 され て い る. 1.3,4) 5‑7)
晴乳類や鳥類 の閉鎖前 の動脈管 に共通 した 構造 上 の特 徴 は その 中膜 平 滑 筋 が 良 く発達 し,所謂 「筋性動脈」(muscularartery)で あ る と言 う点であ り, これは動脈管 に連絡す る肺動脈や大動脈 が弾性型動脈 であ るの とは 全 く対照的であ る.
ニ ワ トリ等 の鳥類 の動脈管 は左右二本存在5.7) し, その全経過 が著 しく長 く,栄養血管や神7) 経終末がほ とん ど観察 されず その壁の構築 が 晴乳類 のそれ に比 して単純 であ り,従 って動 脈管 の閉鎖 と言 う現象の基本的機構 を解 明す る上 には興 味あ る好個 の実験材料 であ る と思 われ る.著者 は動脈管 の閉鎖機構 に関す る研 究 の一端 として, ニ ワ トリにおける動脈管, 大動脈 と肺動脈 の組織発生 を光学顕微鏡的 な
らびに電子顕微鏡的 に詳細 に検索 した.
将化時の動脈管 の閉鎖過程 とその後 の変化8) については別報 に報告す る.
研 究 方 法
僻卵4日目か ら約24時間間隔で僻化直前 ま での 白色 レグホンの歴 を使用 した.光学顕微 鏡 (以下 「光顕」 と略す)的観察 には10%中 性 ホル マ リン,susa液,Bouin液 に よ り固定 し,パ ラフ ィン包埋 を行 い,厚 さ約8〝mの 連続切片を作製 し, ‑マ トキシ リン ・エオジ ン(H・E)染色,Mallory・azan染色, レゾ ル シ ン ・フクシンお よびアルデ ヒ ド・フクシ
鶏腫動脈管 の微細構造 313 ンに よる弾性線維染色を施 した. また電子顕 微鏡 (以下 「電顕」 と略す)用 にEpon包埋 した 1〝m切 片を中性 トル イジンブル ー, メ チ レンブル ーアズ ール・II,PAS,アルデ ヒ ド フクシンに よ り染色 し,光顧的 に観察 した.
電顕的観察 には Millonig の燐酸緩衝液 で pH7.2‑7.4に緩衝 した6.25%グル タール ア ルデ ヒ ドで浸潰 固定 あ るいは還流 固定後 に さ らに2時間固定 し,Millonigの1%OsO4溶 液で2時間,後 固定 を行 った. エ タ ノール系9) 列 に よ り脱水後,
L U F
T の方法 に従 い Epon 812に包埋 した.Porter‑Blum MT一Ⅰあ るい は MT一ⅠⅠ型 超 ミク ロ トー ム に よ り厚 さ 400‑800Åの超 薄 切 片 を作製 し,Millonig の鉛溶液 に よる単染色, あ るいは飽和酢酸 ウ ラン水溶液 と鉛溶液 に よる二重染色を施 し, 日立 HU‑11A特型電子顕微鏡 に よ り加速電 圧 50kVあ る い は 75kV で 直 接 倍 率 2,000‑30,000倍で撮影 した.所 見
ニ ワ トリの動脈管 はお よそ次 の5段階の過 程 を経 てその固体発生上 の全経過 を終 える.
1) 血管 (閉鎖循環系)の発生
2)第6鯉 弓動脈 としての動脈管 の発生 3) 動脈管 の発育 と完成
4) 将化後 の動脈管 の閉鎖 5)10動脈管策へ の退化)
成書 の記載 の如 く, ニ ワ トリの動脈管 は僻 卵4日 目に左右 の第 6鯉 弓動 脈 と して発 生 し,以後 の腫発生 の全期間を通 じて左右 とも に存続 し,肺動脈 と大動脈 との短絡路 として の役割を果たす.左右 の動脈管 は肺動脈 か ら 分岐 した後,右 は大動脈 に伴行 して胸腔 を去
り腹大動脈 の前壁 に合流 し,左は食道の後外 側 を下 って右動脈管 の合流部 よ りやや末梢側 で腹大動脈 の前外側壁 に合流す る.
次 に動脈 管 の発 育 と完 成 した管 壁 の光 顕 的,電顕的構造 を記載す る.
Ⅰ) 動脈管 の発育 とその組織学的構造 組織学的 には,すでに将卵9日目には左右
314 高 橋
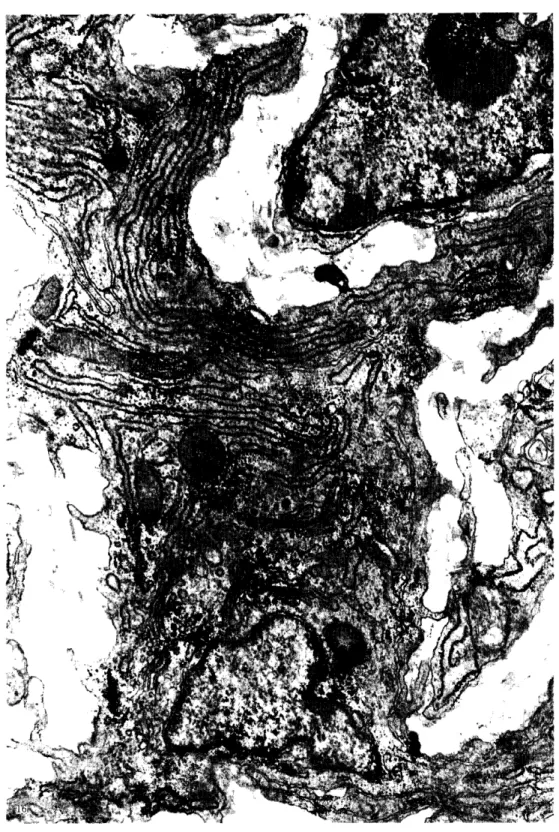
の動脈管 はそれぞれの肺動脈 の分岐部 か ら急 激 にその壁 の厚 さを減 じ,細胞層 は減少 し, 個 々の細胞 は互 いに密 に集 ま り,細胞間層の 厚 さは減 じる. この肺動脈 と動脈管 の壁構造 の違 いは,腫発生の経過 につれて よ り明瞭 と な り,貯卵11日目では動脈管壁全層 の厚 さは 肺動脈 の約 1/3に過 ぎない (図1). この時 期では動脈管壁 を構成す る細胞 は円形 ない し 類 円型で未だ平滑筋 としての形態をほ とん ど 示 していない.僻卵13日目になると肺動脈 と の壁 の厚 さの差はます ます顕著 になるが,動 脈管壁 の内側半分を構成す る細胞 は細長 とな って層状 に配列す る傾 向が見 られ,未熟 なが らも明 らかに平滑筋 としての形態を しめ して いる.僻卵14‑15日目になる と,中膜細胞 の 平滑筋 としての形態 は全 く明瞭で,動脈管 と しての形態分化 ははば完成 し,以後 は貯化時 の形態変化 が起 こるまで,わずかな細胞層の 増加以外 には,管壁 の構造 に著明な質的変化 は起 こらない. このほぼ完成 した動脈管 の大 動脈端か ら肺動脈端 までの全長 を連続切片に よって観察 してみ ると,左右 の動脈管 はその はば中央 よ り遠位部 (大動脈側)では,中膜 の全層 にわた って弾性線維が良 く発達 し,細 胞層 と交互 に配列 して波状 に走 り,その構築 上大動脈 と大差 な く典型的な弾性型動脈 であ る (図2,3). しか しやがて動脈管 のほぼ中 央 か ら中膜 内層 の弾 性線維 が粗 にな り始 め (図4), この弾 性板 の減 少 は動 脈 管 の近 位 (肺動脈側) に進む につれ て急激 に管壁 の全 周に波及 し,管壁 の内側半分 には極 く繊細 な 弾性線維 が少数,平滑筋細胞の間に散在す る のみ とな り,その結果,管壁 の全層の厚 さは 遠位部 に比 して著 し く薄 くその約 1/4に過 ぎない (図5,6). この弾性線維 の粗 な中膜 内層 には2‑4層の大型 の平滑筋細胞が輪状 に配列 し (図6,7),明 らかに典型的 な筋性 動脈 としての構造を有 している. この特徴的 な構造 は,左右 とも肺動脈 か らの分岐部直下 か ら始 ま り,遠位 に対 してははは中央 の短 い 移行部 を境 として,動脈管 の近位約半分に限
March,1992 HirosakiMed.∫.43(4) 局 している.次報 に示す如 く,筋性動脈であ るこの近位半分は僻化後最 も早期 に内腔 の狭 細 と完全閉鎖を起 こす部分であ るのに対 して 遠位約半分は僻化後数 日で もなお内腔 の完全 閉鎖が起 こらず大動脈側 に対 して漏斗状 に開 存 している.未完成 な大動脈 と肺動脈 の各部 分が成熟 した弾性型動脈 としての構造を有す るに至 る過程を連続切片法 に よ り追求 してみ る と,大動脈 のいわば「弾性型動脈化」,即 ち 中膜 における弾性線維 の増加 は,大動脈 の心 臓側 か ら開始 して次第 に末梢 に波及 し,動脈 管 の開 口部 に達す ると, さらに末梢側 に進行 す ると同時に,左右 の動脈管 の大動脈‑の開 口部 か ら逆行 して,動脈管 のほぼ中央の管壁 に まで波及す るが これ よ り近位 (肺動脈側) には達 しない.一方第6鯉 弓動脈 の 「弾性型 動脈化」 も心臓側か ら開始 して次第 に末梢側 に進行す るが,動脈管 と肺動脈 の分岐部では そのまま肺動脈 に波及す るが動脈管 には波及 しない.その結果,左右 の動脈管 の遠位半分 は大動脈 と同様 な弾性型動脈 としての構造を 有す るよ うになるが (図2,3),近位半分に 限 って弾性線維の発達 は貧弱で筋性動脈 とし ての構造を有す るよ うになる (図5‑7).
ⅠⅠ) 閉鎖前の動脈管 の微細構造
鶏腔 の大動脈 の微細構造に関 してはすでに ll)
若干の報告があ り, また動脈管 の遠位半分の 構造は大動脈 のそれ とほは同様 であるか ら, 本論文では貯卵14日か ら僻化直前 までの近位 半分の動脈管 に特徴的 な構造 についてのみ記 載す ることとす る.
近位半分の動脈管壁 は他の大血管 と同様 に 内膜,中膜 お よび外膜 か ら構 成 され て い る (図7).
1) 内膜 (Tunicaintima)
内膜 は一層の内皮細胞 と内弾性板を含む内 皮下腔 か ら成 る.内皮細胞 はその核周囲部 の 高 さが2‑4〟mの比較的小型 の細胞で,核周 囲部以外 は比較的偏平であるが (図8,9), 最 も薄い部分で も約 0.2〃m以上の厚 さを有 し,有 窓 性 毛 細 血 管 で 見 られ る様 な 小 孔
平成 4年 3月 弘前 医学 43巻 4号
(fenestration)は存在せず,動脈管の内面を 完全 に連続的に被 っている.核 は光学顕微鏡 的 には核膜 の深い切れ込みによ り凹凸不整な 輪郭を呈 している (図8,9).内側 および外 側核膜 は核 孔 の部 分で互 いに反転 して連続 し,比較的拡大傾 向のある核槽 を形成す る.
外側核膜 の細胞質面 には リボ ゾー ムが付着 し, しば しば外側核膜が細胞質内に突出 し粗 面小胞体 と連続 し,その内腔内に電子密度の 低 い物質 を入れ る (図9,10).径約500Åの 核孔はporemembrane(diaphragm)によ り 閉 じられてお り, これに よ り核質 と細胞質は 画然 と区別 され る.細胞内小器官 としては, 小胞状 ない し小管状の粗面小胞体が比較的良 く発達 し,時に層状 に配列す る. ゴルジ装置 の発達 は悪 く, ミトコン ドリアは小型で少数 散在す るのみであ る.一層の限界膜で囲まれ た径 500‑1,000Å大 の電子密度の高い頼粒 が散見 され るが, しば しば この近傍 に電子密 度の高い基質を有す る多胞体が観察 され る.
内皮細胞はその管腔側表面か ら少数の徴械毛 様 の細胞質突起 を出 し (図8),飲小胞は内皮 細胞 の管腔側のみな らず,基底側 および外側 の形質膜 に も観察 され る (図8).内皮細胞間 の接触面では形質膜は 100‑200Åの間隔を へだててほぼ平行 に走 るが,密着結合や細隙 結合 な どの junctionalcomplexの形成は著 明ではない.内皮細胞 の基底側表面か ら小 さ な舌状突起が内皮下層に向か って突 出 し, と ころどころで平滑筋細胞 に操す る (図9).内 皮細胞 には遊離 の リボ ゾームが散在 している が,グ リコーゲン頼粒 はほ とん ど存在 しない.
細胞質内には,径約 60Åのマイク ロフィラ
メン トが束 状 を な して不規則 に走 るが (図 10),ときに基底部形質膜 に集中 し,halfdes一 mosome様構造 を形成す る.内皮細胞直下の 基底膜構造 は明瞭ではな く,内皮細胞 と最内 層の平滑筋細胞 との間には巾約 0.3/〃n の内 皮下腔 (subendotheliallayer)が介在 してお り, このなかには波状をな して屈曲 した弾性 線維 の断面や中等度の電子密度を有す る細頼
鶏歴 動 脈管 の微細構 造 315 粒状ない し無定形 の基質が含 まれている (図
8,9).弾性線維 は, ウランと鉛 に染 まらず 電子密度の低 い centralamorphousregion (elastin)と,ウランと鉛の両者 に濃染 し中心 部の電子密度が低 く小管状構造 を呈す る径約 100Åのperipheral丘bril(micro丘bril)か ら 構成 されてお りその微細構造は外膜の弾性線 維のそれ と同様である.
以上の内膜 の構造 は僻化時の中膜平滑筋の 収縮 に よって内皮細胞が受動的に寄せ集め ら れ るまで変化せず,内皮細胞 の増殖や内皮下 結合組織 の肥厚あ るいは ムコイ ド物質の蓄積 な どはみ られない.
2) 中膜 (Tunicamedia)
中膜 はそれぞれ3‑4層の平滑筋細胞か ら 成 る内外二層 の細胞層 と細胞間結合組織層か
ら構成 されている.
中膜内層の平滑筋細胞 は光学顛徴鏡的観察 か らも明 らかな様 に (図7),良 く分化 した大 型 の平滑筋細胞 としての特徴を有 している.
す なわち大型 の長 円形 の核 には平滑筋 に特徴 的 な深 い切れ込みがあ り, このため超薄切片 標本では分葉状を呈す ることもある (図9). 主 として核周囲の細胞質 には小 さい ゴルジ装 置,少数の小型の ミトコン ドリアが観察 され,
また粗面小胞体 の発達 も悪 い.平滑筋細胞の 核周囲部や大 きな細胞質突起の大部分を占め る筋細糸は直径 60‑80Åで, いわゆ るthin 丘lamentのみか ら成 り,thick丘Iamentはほ
とん ど観察 されない.筋細糸は互いに平行に 配列 して密 に集束 し,細胞の長軸におよそ平 行に走 る (図11).所 々において筋細糸が特 に 集合 し,筋細糸間物質の電子密度が増加 して いわゆるdensebody(fusiform density)を 形成 し, また形質膜の内側 に も密に集合 して 電子密度を増 しattachmentplaqueを形成 す る (図11,12).
平 滑 筋 細 胞 の グ リコ ー ゲ ン額 粒 は直 径 300‑400Åの鉛 に濃染 した個 々一つ一つの 細頼粒 (β‑particleglycogen)として観察 さ れ, リボ ゾームとは容易に区別 され る. この
316 高 橋 HirosakiMed.∫Mar.4ch,13(949)2
竃 車 確 撃 撃
平 成 4年 3月
弘前 医学 43巻 4号 鶏旺動脈管 の微細構造 317
図 1 筋卵11日目の左肺動脈(Pm)と動脈管(Da)の分岐部.動脈管 は肺動脈 の分岐部 か ら急激 にそ の壁 の厚 さを減 じ,肺動脈 の約1/3に過 ぎない.迷走神経 と反 回神経 が動脈管 と肺動脈 の分岐 部 に密接 して走行す る.Rln:反 回神経,Vn:迷走神経,Br:左気管支.H・E染色×100. 図 2 僻卵15日目の右動脈管 の遠位 (大動脈側)約1/4と大動脈 の横断面像.動脈管 (Da)の壁構造
は大動脈 (Ao)のそれ と大差 な く弾性型動脈であ る. レゾル シン ・フクシン染色×100. 図 3 僻卵15日目の左動脈管 の遠位約1/4の横 断面.動脈管の壁構造 は大動脈 のそれ と同 じ く弾性型
動脈 である. レゾル シン ・フクシン染色×100.
図 4 僻卵15日目の左動脈管のほぼ中央 の横 断像.管壁の一部が薄 くな り,弾性板 が粗 にな り始め る.
レゾルシン ・フクシン染色×100.
図 5 筋卵15日目の左動脈管 の近位 (肺動脈側)約1/4の横 断面.管壁の厚 さは遠位部 の約1/4に過 ぎず,その中膜内層 には弾性線維 が乏 しい. レゾルシン ・フクシン染色×100.
図 6 図5の強拡大.内弾性板 は薄 く,中険内層 には極 めて繊細 な弾性線維 がわずかに認め られ るのみ で筋性動脈 であ る. レゾル シン ・フクシン染色×500.
図 7 僻卵18日目の左動 脈 管 の近 位 約 1/6の横 断面.中膜内層 には良 く発達 した大型 の平 滑筋細胞,中瞭外層には小型の平滑筋細胞 が層状 に配列 し外膜 には未分化間葉性細胞 が散在 している.Epon包埋,中性 トル イジ
ンブル ー染色×1,000.
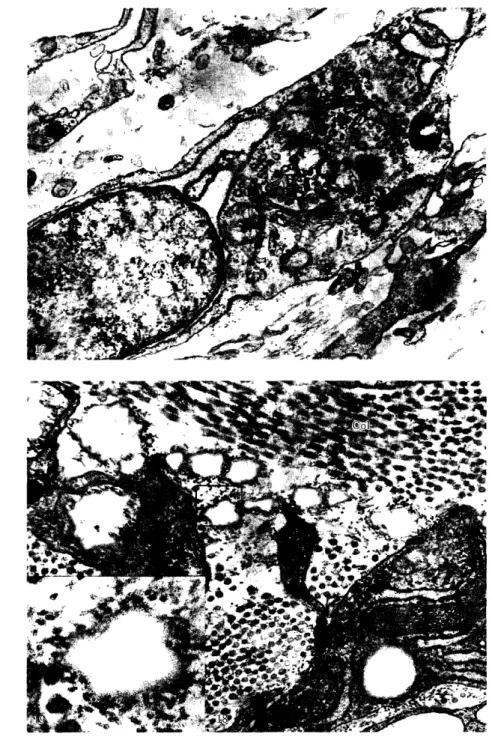
グ 1)コ‑ゲン頼粒は細胞質全体 にかな り豊富 に散在 してお り, また小 さな細胞質突起内に も観察 され る (図8).最内層の平滑筋細胞 に
紘,前述の如 く,内皮細胞 か らの舌状突起が 接す る一方,平滑筋細胞 も小 さな舌状突起を 内皮下腔 に出 し,所 々において内皮細胞 の基 底部 に接す るが,内皮細胞 と平滑筋細胞 との 間には常 に約 100Å以上の細胞間隙があ り, junctionalcomplexの如 き特殊 な形 質膜 の 分化は見 られない (図8,9).
平滑筋細胞 の基底膜は極めて薄 く,不完全 であ って,しば しば弾性線維のmicro丘brilに 類似 した細線維が平滑筋細胞 の表面 に密接す る (図11). 同一層の平滑筋細胞は,end‑to‑ endに も, またside‑by‑sideに も接す る.
平滑筋細胞 は互 いに小突起を もって密 に接 触 し,ときに上皮細胞 におけるintem ediate junction様構造 を形成す るが (図13),他の平 滑筋細胞で見出された細隙結合構造 は明瞭で ない.平滑筋細胞表面の飲小胞 はご く少数で あ り,被覆小胞 も著明でない.
中険外層の平滑筋細胞は,一般 に小型で筋 細糸に乏 しく,平滑筋細胞 としては未熟 な形 態を示 しているが (図14),この中膜外層の平 滑筋細胞のなかには,発達 した細胞 内小器官 を有す る特異 な細胞が混在 している.図15に 示す細胞 は,豊富 な粗面小胞体,ゴルジ装置,
ミトコン ドリアを有 し,線維芽細胞 に類似 し た形態を示す一方,細胞辺縁部 には少量 なが ら筋細糸が発達 し,densebodyや attach‑
mentplaqueの形成 もみ られ,明 らかに平滑
橋高
813 March,1992
HirosakiMed.J.43(4)
図 8 図7と同 じ箇所 の電顕像 (貯卵18日目).内皮細胞 (End)の管腔側 か ら少数の小細胞 質突起 が出る.内皮細胞 の核槽 が拡大 し,外側核膜 が粗面小胞体 と連絡 してい る (小 矢 印).内皮細胞 の基底側 か ら舌状の細胞質突起 (大矢 印)が内皮下腔 を貫 いて平滑筋 細胞(Sm)に接す る. この内皮細胞 の突起間にはグ リコーゲン額粒 を含 んだ平滑筋細 胞 の小突起 (Ⅹ)が介入 してい る.内皮下腔 に内弾性板 の不連続的 な断面 (EI)が見
られ る. ウラソ,鉛二重染色×21,000.
平成 4年 3月
弘前 医学 43巻 4号 鶏腫動 脈管 の微細構造
図 9 図7と同 じ箇所 の電顕像 (僻卵18日日).大型 の平滑筋細胞(Sm)が層状 に配列す る.
平滑筋細胞質 には比較的多数 の グ リコーゲン額粒 が散在 してい る. ウラン,鉛二重染 色×17,000.
図 10 図9の内皮細胞 の高倍率像.電子密 な細額粒 と, マイク ロフ ィラメン トの横 断面 が見 られ る. ×59,000.
図 11 図7と同 じ箇所 の良 く分化 した 平滑 筋細 胞.細 胞 質 内 に は径 60‑80Å の thin 別amentか ら成 る筋細糸が豊富 で あ るが thickmamentは明瞭 で はない.平滑筋細 胞 の基底膜 は薄 く,弾性線維 のperipheral斤bril様 の微細線維 が形 質膜 に密着 してい
る (矢 印). ウラン,鉛二重染 色×44.000.
319
322 高 橋
筋細胞 としての特徴 も有 してい る.中膜外層 の平滑筋細胞 には,時に有糸分裂像が観察 さ れ るが,中膜内層のすでに平滑筋 としての分 化 を終 えた細胞 には有糸分裂像はほ とん ど観 察 されない.
中膜 の最 外 層 (中膜 と外 膜 の境 界部) に は,筋細糸を欠 き,核周囲部の大 きな ゴルジ 装置,比較的多数の ミトコン ドリア,お よび 細胞質全体 に層状 に配列 した粗面小胞体を有 す る大型 の細胞 が散在 してい る (図16).これ らの細胞 は,約150Åの間隔をへだてた接触 面を複雑 に入 り込 ませ互 いに密接 して平滑筋 同様 な細胞配列を とり, この点通常の線維芽 細胞 とは異 なってい る.
中膜の細胞間結合組織層 は,弾性線維,豚 原線維,micro丘brilおよび無定形の基質か ら 成 るが,外層 ほ どその厚 さを増 し,外膜の結 合組織 に移行す る.中膜内層の弾性線維 は, peripheral丘brilが著 明で centralamor‑ phousregionの 占め る割合 は比較的小 さ く (図11,12,14),弾性線維全体 としてほごく繊 細 で あ るが,外層 に行 くにつ れ て central amorphousregionが増加 し (図15,16),弾 性線維全体 としては太 く,平面的 な弾性板が 形成 され る.
3) 外膜 (Tunicaadventitia)
外膜の線維性結合組織 の中には,細胞 内小 器官に乏 しい小型 の未分化間葉性細胞 か ら, よく発達 した粗面小胞体 を有す る線維芽細胞 に至 るまでの各種 の成熟過程 にある細胞が散 在 している.線維芽細胞‑の分化を示す間葉 性細胞では, ときに リボ ゾームの付着 した外 側核膜が細胞質内に深 く突出,延長 し,電子 密度の低 い物質を入れた大 きな槽 を形成 して お り,明 らかに粗面小胞体 との連続を示 して いる (図17).これ らの線維芽細胞 は,中膜外 層の粗面小胞体 の発達 した平滑筋細胞や線維 芽細胞様細胞 と共 に増殖 し,謬原線維,弾性 線 維 お よび細 胞 間 基 質 を 分 泌 す る (図18, 19).
第6鯉 弓神経の迷走神経 とその分枝 の反回
March,1992 HirosakiMed.∫.43(4) 神経が動脈管 の肺動脈か らの分岐部に密着 し て走向す るが (図1),近位半分の動脈管壁 内 には神経線維束や神経終末の分布は観察 され ず, また栄養血管 も観察 されない.
考 察
動脈管開存症のみな らず,大血管完全転移 症や大動脈離 断症 な どのいわゆ る "ductus dependent"な大血管や心臓 の奇形 において
も出生後 に動脈管が正常 に閉鎖す るか,ある いは開存す るか と言 うことが血行動態上重要 な意義 を有す ることは勿論であ り,従 って出 生後 に動脈管が閉鎖す る機構 に関 して多 くの 研究が報告 されて きた.
晴乳類や鳥類の動脈管壁 の構造上の特徴は その中膜平滑筋が良 く発達 し,所謂 「筋性動 脈」であるとい う点であ り, これは動脈管 に 連続す る大動脈や肺動脈が 「弾性型動脈」で あるの とは全 く対照的である.
一般に,動脈管 の閉鎖現象は出生後早期 に 起 こる 「機能的閉鎖」"functionalclosure"と これ に引 き続 いて起 こる 「解 剖学 的 閉鎖」
"anatomicalclosure"の2相か ら成 るとされ 2,12)
てお り,動脈管が 「筋性動脈」であ ることか らその中膜平滑筋細胞 が管腔閉鎖,特 にその
"functionalclosure"において重要 な役割を 果たすであろ うと考 えられ るよ うになった.
この動脈管の平滑筋に対す る最 も有効な刺激 が酸素分圧の上昇であることは,幾多の in v1i3v)oお よび in vitroの実験 で示 され て い る. また 出生 前 に は動 脈 管 壁 の 緊 張 性 は prostaglandinE2,や14) prostaglandin12に よ り調 節 され て お り, そ れ らの 阻 害 剤 (in‑
domethacinや aspirinな ど)が動脈管の収 縮作用を有 していることが明 らかにされてお15) り,動脈管の中膜平滑筋細胞 は大動脈や肺動 脈 のそれ とは異 なる特異 な機能的性質を有 し ている.晴乳類の動脈管 は筋性動脈であると 言 うことの他 に,贋動脈や肺静脈 ほ どではな いに して もかな り多量のmucoid物質が存在 す ること,閉鎖 に際 して内膜増殖が起 こるこ
平 成 4年 3月
弘前 医学 43巻 4号 鶏 旺 動 脈 管 の微 細 構 造 323
図 16 図 7と同 じ箇 所 の中膜 最 外 層 . これ らの細 胞 は筋細 糸 を欠 き, 層 状 に 良 く発 達 した粗 面 小胞 体 , 大 きな ゴル ジ装 置 を有 し一 見 線維 芽 細 胞 様 で あ るが, 互 い に広 い接 触 面 を もって密接 し, 平 滑 筋 細 胞 様 に層 状 に配 列 して い る. ウ ラン,鉛 二 重 染 色×27,000.
324 高 橋 March,1992 HirosakiMed.∫.43(4)
図 17 図 7と同 じ箇所 の外膜 に見 られた未分化間葉性細胞,外側核膜か ら粗面小胞体が発達 しつつあ る. (矢 印) ウラン,鉛二重染色×26,000.
図 18 図 7と同 じ箇所 に外膜の結合組織.弾性線維 (EI) はcentralamorphousregionと peripheral丘brilか ら成 る.col:鯵原線維. ウラン,鉛二重染色×28,000. 図 19 図18の高倍率像.弾性線維 のperipheral丘brilは径約100Å で中心部 の電子密度が低
く中空性管状構造 を呈 してい る. ウラン,鉛二重染色×100,000.
平成 4年 3月 弘前医学 43巻 4号
4) 16)
と, また神経終末や栄養血管が分布 している ことな どのために,その閉鎖過程 には中膜平 滑筋のみな らず上記 の田子 も関与 し,かな り 複雑 な機構を有す るであろ うと考 え られ る.
これに反 し, ニワ トリの動脈管 は左右2本存 在 し,その走 向が著 しく長 く,糖化後最 も早 期 に狭窄を釆たす近位 (肺動脈側)動脈管は 壁 の薄 い筋性動脈 で,比較的単純 な構造を有 しているので動脈管の閉鎖機構の研究のため には好個 の実験材料である.
次に, ニ ワ トリの動脈管 の閉鎖過程 におい て最 も重要 な役割を果たす近位半分の構造 に 関す る所見について若干の考察を加 える.
1) 中膜内層平滑筋 について
動脈管 の近位部 は,酵卵9日目で これ と連 続す る遠位部や肺動脈 とは構造を異 に してお り,既 に酵卵14‑15日目には,大動脈や肺動 脈に先ん じてはば完成 した壁構造,す なわ ち 中膜平滑筋の良 く発達 した筋性動脈 としての 構造を有 している. ニワ トリの動脈管 を最 も 特徴づけるこの良 く発達 した中膜平滑筋細胞 は大型で,その細胞質の大部分は筋細糸によ って占め られ,その他の細胞内小器官 には乏 しい.次報 に示す如 く, この中膜平滑筋細胞 が醇化時の動脈管の機能的閉鎖 に際 し,著明 な形態変化 (収縮)を示す.
2) 中膜外層の平滑筋 について
中喋外層の平滑筋細胞 は筋細糸に乏 しく平 滑筋 としては未熟 な形態を示 している反面, 良 く発達 した粗面小胞体や ゴルジ装置,多数 の ミトコン ドリアを有 している. これ らの未 熟 な平滑筋細胞 は,平滑筋 としての収縮を有 す る一方,外膜 の間葉性細胞 と同様 に,細胞 間の結合組織線維や基質を合成す る能力を兼 ね備 えた筋 線維 芽細胞 (myo丘brobrast)で あ り,動脈硬化巣の平滑筋細胞やエス トロゲ ンで刺激 された子宮筋層の平滑筋細胞 と同様17) な多能性間葉細胞 である と考 えられ る.
3) 弾性線維 の微細構造 について
弾性線維 の電顕的微細構造 については現在 なおその構造の保存や電子染色に満足すべ き
鶏腫動脈管 の微細構造 325 方法が確立 されてお らず不明の点が多いが, 一般 に弾性線維 は ウランと鉛 に濃染す る中空 性管状の径100
Å
のperipheral丘brilと,燐 タングステン酸 に濃染す る cent1r8al a) mor‑ phousregionか ら構成 されてい る.光顕的に も中膜外層や外膜 の弾性線維 は, 動脈管 の遠位半分や大動脈 のそれ と同様 に太 く,弾性板 を形成 してい るが,中膜内層には 極めて繊細 な弾性線維 がわずかに存在す るに す ぎない.電顕的には中膜内層の弾性線維 は, peripheral丘brilが小 さく相対的に central amorphousregionが著明であ り,中膜外層 や外膜の弾性線維 は,それが太 い程 central amorphous region の占め る割合が大 きく peripheral丘brilは相 対的 に少 な い (図16, 17).従 って 弾 性 線 維 は主 と して central amorphouregionの増1加8)に よってその太 さ を増す もの と考 えられ る.
4)動脈管 の発生学的起源
ニワ トリの動脈管 の遠位 (大動脈側)半分 は大動脈 とほぼ同様 な 「弾性型動脈」であ り 次報で示す如 く,醇化後 の動脈管閉鏡 におい て も近位部 の閉鎖 に続発 して二次的に閉鎖す るのみで,酵化後数 日で もなおその管腔は漏
7) 斗状 に開存 して,大動脈 に開 口している.
分枝 もな く血流状態 もほぼ同 じと考 えられ る連続 した一本の動脈 であ る動脈管がその近 位部 と遠位部で構造的に も幾能的に も著 しく 態度を異 にす ると言 う一見奇異 な事実は,動 脈管の遠位部が発生学的には本来背側大動脈
5) に由来 している とす る HUGHESらの説 を考 慮 しなければ容易には説 明 し難 い と思われ る し,括約筋様 の中膜平滑筋が第6鯉 弓動脈 に 由来す る近位部のみに発達 し背側大動脈 に由 来す る遠位部 には発達 しない と言 う現象は,
「大血 管 の構 造 は主 として遺伝 的 に決定 さ れ,末梢血管 の壁構造は主 として局所 の血流 状態 に依存す る」 とす る19) BENNINGHOFFらの 古典的な見解 に も合致 している. さらにすべ ての鯉 弓動脈 の中膜平滑筋細胞や大動脈肺動 脈中隔の間葉性細胞 は,菱脳領域の神経堤細