タイ伝統発酵食品中の乳酸菌の多様性について
宮下 美香 *、中川 恭好、鈴木 健一朗
独立行政法人製品評価技術基盤機構
バイオテクノロジーセンター(NBRC)
年間を通じて高い気温と湿度が保たれるタイ王国では食材が豊富なこともあり、様々な発酵食品が存在 しており、タイ料理の独特の風味や旨味に欠かせないものとなっている。これら発酵食品には乳酸菌が生 息することが知られていて、風味や旨味の付与、また高温多湿における食品の保存に重要な役割を担って いる。本稿では、様々なタイの発酵食品とそこに生息する乳酸菌の多様性について、著者のこれまでの調 査結果を交えて概説する。Key words:traditional fermented food, Thailand, microbial diversity, lactic acid bacteria
* To whom correspondence should be addressed. Phone : +81-438-20-5763 Fax : +81-438-52-2329 E-mail : [email protected] 1.タイの多様な発酵食品 比較的穏やかな熱帯モンスーン気候に属するタイ王国で は、様々な果物や野菜が栽培されており種類が豊富であ る。また肥沃な大穀倉地帯を抱えるタイ中央部では米の生 産が盛んで、日本と同じく米が主食である。タイ湾に面し た海岸線を有していて魚介類も一般的によく食べられてい るが、メコン川やチャオプラヤー川といった大河が流れて おり、生息する淡水魚の種類も豊富であるため、海岸部以 外では川魚が中心に食されている。これら果物や野菜、穀 物、魚介類に加えて鶏肉や豚肉を中心とした肉類や香辛料・ 香草類など食材は多種多様で、豊富な食材を活かしたタイ 料理は辛み・酸味・甘みなどが複雑に組み合わされた独特 の味付けが特徴となっている。その味は世界的にも認めら れており、日本でも多くのタイ料理レストランを見つける ことができる。このタイ料理独特の風味や旨味を生み出す 要因のひとつとして、発酵食品が欠かせない。Nam-pla(魚 醤)をはじめとして、Kapi(エビ味噌)や Tuaw jaew (Tao jiao, Thua nao)(発酵大豆)のように日本の味噌や醤油に よく似た、タイ料理の味付けのベースとなるような発酵食 品や、その他にも食材の多様性に比例してタイの発酵食品 には様々な種類がある。
肉の発酵食品としては、豚肉や牛肉に蒸した米や香辛料 などを混ぜ合わせて発酵させた酸味の強いソーセージであ る Sai krong prieo や Nham などがあり、タイ北部や東北 部の料理として知られている。 漬物など植物由来の発酵食品に用いられる食材は、カラ シナやナス、ダイコン、タマネギ、タケノコなど日本でも なじみ深いものから、バナナの花やフウチョウソウ科の植 物、マンゴー、タマリンドなど熱帯特有のものまで様々で ある。これらは比較的低い塩濃度(2 ~ 6%)で漬けられ て、少量の炊いた米が加えられる場合が多いが、Noa mai (タケノコの漬物)や Miang(発酵茶葉)などのように無 塩のものもある。主食である米を原料とする発酵食品も多 様で、Khanom jeen(発酵米麺)や、麹を加えて発酵させ
る Khao mak(甘酒)や Sato(米酒)などがある1-2)。年
間を通じて気温が高いため、日本の漬物製造のように寒い 時期に仕込んで長期熟成をさせるといった方法ではなく、 数日間だけ漬ける浅漬けが主流である。しかし日本の味噌 に似た大豆の発酵食品である Tuaw jaew (Tao jiao, Thua nao)は、日本の味噌同様、大量の塩が加えられ、比較的 長期間(数ヶ月~数年間)熟成させる。 魚介類の発酵食品においても数日から数週間の発酵期間 で食されるものが多い。主に川魚を原料とし、植物由来の 発酵食品よりも比較的高い塩濃度で漬けられ、炒った生米 を粉砕して粉状にした Kao kouwa または蒸米、餅米、さ らに種類によっては香辛料や水などを加えて発酵させる。 スターターとして米麹や発酵が終わった残りを加える場合 もある。また、加える食塩の量を増やして数ヶ月から数年
総
説
間の発酵期間をもうけるものもある。タイ語で「魚(pla) の水(nam)」と呼ばれる Nam-pla(魚醤)は、日本のしょっ つる(塩汁・塩魚汁)と名前の成り立ちも非常に似ている が、作り方も同じく大量の塩と共に魚を発酵させた液体調 味料で、長期間の発酵で熟成させる3)。 2.タイ発酵食品中の乳酸菌 東南アジア各国の発酵食品には共通点が多い。日本とタ イの発酵食品においても例外ではなく、両国が主食とする 米を原料とした発酵食品や、醤油や大豆の発酵食品、漬物 など日本と共通するものが多い。これらの食品に生息する 微生物については、タイはもちろん日本の研究者によっ ても研究報告がされている。内村ら(1991)は、タイ各 地の製造工場から合計 37 サンプルの発酵米麺(Khanom jeen)を収集し、GYP 白亜寒天培地を用いて乳酸菌を分 離した。表現性状による同定試験の結果、Lactobacillus (Lb.)
acidophilus、Lb. plantarum、Lb. reuteri、Pediococcus (P.) acidilactici、Streptococcus (Str.) lactis、Str. thermophilus
が存在することを報告した1)。岡田ら(1986)は、タンニ
ン酸を含む無塩の発酵茶葉(Miang)の乳酸菌叢を調べる ため、発酵期間の異なる 5 つの Miang から GYP 白亜寒天
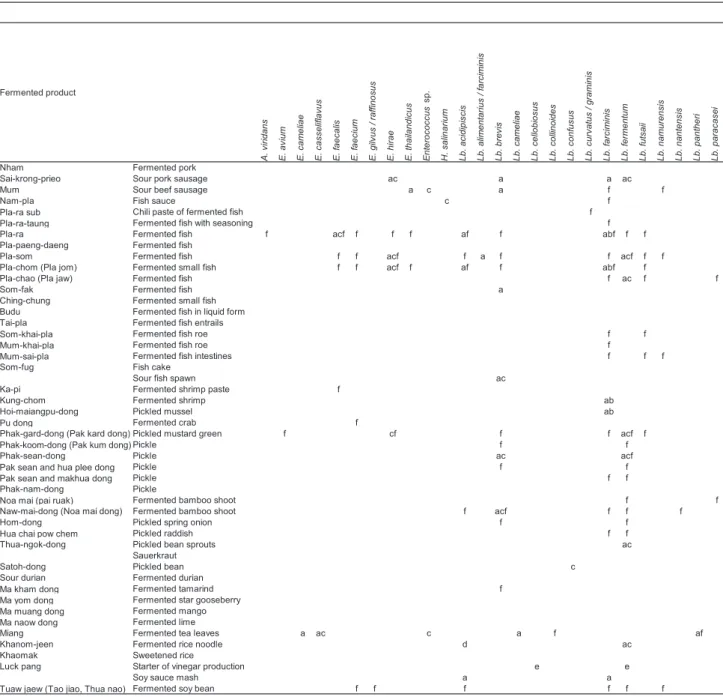
Table 1. Distribution of LAB and Staphylococcus species in Thai fermented foods.*(※次の P も)
* Species isolated from Thai fermented foods shown by a to f were reported by a, Tanasupawat, 2009 14); b, Tanasupawat et al., 1998 6); §A., Aerococcus; E., Enterococcus; H., Halolactibacillus; Lb., Lactobacillus; Lc., Lactococcus; P., Pediococcus; Str., Streptococcus; T.,
培地及び茶葉抽出液を加えた同培地を用いて乳酸菌の分離 を行った。その結果、Lb. plantarum が最優性種であるこ
とを報告した4)。Tanasupawat ら(2002)は、タイの工
場から収集した発酵段階の異なる醤油もろみ(Soy sauce mash)14 サンプルから、5% NaCl を加えた MRS 培地を 用 い てTetragenococcus (T.) halophilus、Lb. acidipiscis、 Lb. farciminis、Lb. pentosus、Lb. plantarum を分離し、T. halophilus は醤油発酵初期から後期にわたる全てのサンプ
ルから、Lb. acidipiscis は発酵中期から後期のサンプルか
ら分離されたことを報告した5)。また 4 種類の魚の発酵食
品(Pla-ra, Pla-chom, Kung-chom, Hoi-dong)からは、5%
NaCl を加えた MRS 培地および 2 倍希釈した MRS 培地を
用いて 52 株を分離し、同定の結果それら分離株はLb.
far-ciminis、Lb. pentosus、Lb. plantarum、Leuconostoc sp. に
分類され、10% 以上の食塩を含む Pla-ra からLb.
farcimi-nis は分離されたが、Lb. pentosus と Lb. plantarum は分 離されなかったことを報告している(Tanasupawat ら、
1998)6)。Soy sauce mash および数種類の魚の発酵食品か
らはStaphylococcus 属細菌が分離されている7)。
Staphy-lococcus piscifermentans はタイの Kung-chom(発酵エビ) を起源として新種提案された。また Pla-ra、Bu-du からも
同種が分離されたと報告されている8)。
Table 1. (※続き)
c, Tanasupawat et al., 1995 15); d, 小崎 , 1991 16); e, 内村ら , 1991 17); f, Miyashita et al., 2012 18).
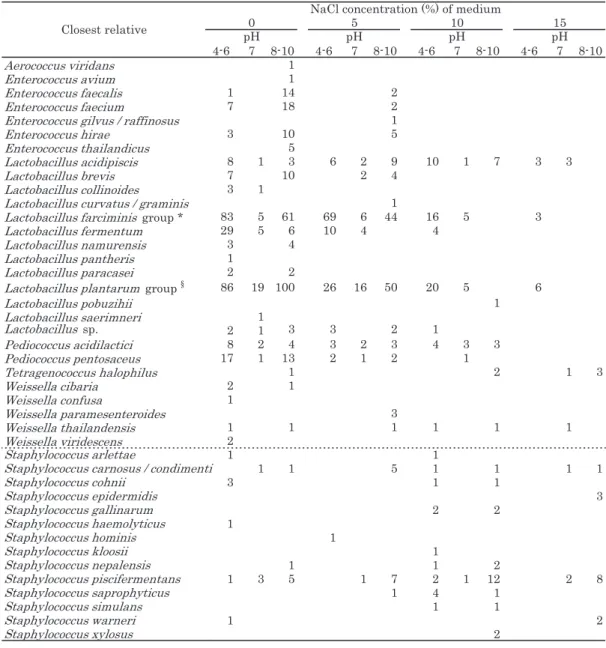
これらの報告にもあるように、これまで、乳製品を除 き、肉・魚・植物の発酵食品からの乳酸菌の分離には、一 般的に MRS や GYP などグルコースを糖源とする培地を 中性から弱酸性に調整し、分離源の食塩濃度によって 5% 程度の NaCl を加えた培地が用いられてきた。一方、石川 らは 7% NaCl と pH 9.5 または pH 10 を組み合わせた条件 で海洋環境からの好塩・好アルカリ性の乳酸菌の分離に 成功し、さらにこの条件を食品にも適用して Kapi や熟成 チーズにおける好塩・好アルカリ性乳酸菌の存在を明らか にした9-13)。タイ発酵食品には高濃度の食塩を含む発酵食 品が数多いため、好塩性或いは耐塩性乳酸菌が生息する 可能性は高いと推測できる。また高塩濃度を好む乳酸菌 は、同時にアルカリ性を好む種が多いことから、NaCl 濃 度だけでなく pH も組み合わせて変化させる事が必要と なる9, 10, 12, 23)。 筆者らは 2008 年度から 2010 年度にかけて、タイ東北部、 中部、及び北部南寄りの地域から 114 の様々な発酵食品を 収集し、内 14 サンプルには NaCl 濃度が 0, 5, 10, 15% と pH 4.0 ~ 10.0 を組み合わせた条件に調整した MRS 培地、 その他のサンプルには NaCl 濃度 0, 5 または 10% と pH 6.0, 8.0 または 10.0 を組み合わせた条件に調整した MRS 培地 を用いて乳酸菌の分離を行った。得られた 1032 株の分離 株のうち、16S rRNA 遺伝子配列に基づく解析により 945 株 はAerococcus viridans、Enterococcus (E.) avium、E. faecalis、E. faecium、E. gilvus/E. raffinosus、E. hirae、 E. thailandicus、Lactobacillus acidipiscis、Lb. brevis、 Lb. collinoides、Lb. curvatus、Lb. farciminis、Lb. fer-mentum、Lb. futsaii、Lb. namurensis、Lb. nantensis、Lb. paracasei、Lb. paralimentarius、Lb. plantarum group、 Lb. pobuzihii、Lb. saerimneri、Pediococcus acidilactici、P. pentosaceus、Tetragenococcus halophilus、Weissella (W.) cibaria、W. confusa、W. paramesenteroides、W. thai-landensis、W. viridescens の 6 属 29 種と幾つかの新規なグ
ループに分かれた。その他の 87 株はStaphylococcus (S.)
属に含まれ、S. arlettae、S. carnosus/S. condimenti、S. cohnii、S. epidermidis、S. gallinarum、S. haemolyticus、 S. hominis、S. kloosii、S. nepalensis、S. piscifermentans、 S. saprophyticus、S. simulans、S. warneri、S. xylosus の
14 種に分かれた。これらの種には、様々なタイの発酵食 品から分離された乳酸菌の多様性がまとめられた Tanasu-pawat による報告や個々の発酵食品を詳細に調べた研究報 告では見られなかった種が含まれる(Table 1)6, 14-18)。 3.NaCl 濃度と pH による検出種の変動 分離した種を、分離培地の NaCl 濃度と pH の条件ごと にみてみると、一般的に乳酸菌の分離に用いられる無塩・ 弱酸性の条件において最も多様な種が分離されたことがわ かる(Table 2)。一方、NaCl 10% 以上の条件下ではLb.
plantarum group と Lb. fermentum 以 外 は Lb. acidipis-cis、Lb. farciminis group、Lb. pobuzihii、P. acidilactici、P. pentosaceus、T. halophilus、W. thailandensis などの 10%
以上の耐塩性が報告されている種19-23)が分離され、NaCl 濃度 15%, pH 8 以上の条件ではT. halophilus と Staphylo-coccus 属の種のみが分離された。NaCl 濃度 10% 以上の分 離培地では分離されなかったEnterococcus 属は、5% 以 下の NaCl 濃度においては主に pH 8 ~ 10 のアルカリ域で 分離された。 ひとつのサンプルに対して、NaCl 濃度と pH の条件を 組み合わせた 20 種類の分離培地を用いて分離した結果か らも、条件の違いで分離される種に違いがあることが確認 出来る。低い塩濃度の分離源からは、分離条件においても 低塩濃度の培地における菌株の出現頻度が高かったが、分 離源の塩濃度が高くなれば、比例して高塩濃度培地におけ る菌株の出現頻度も上がり、分離株も耐塩性の高い種の 分離頻度が高くなった。このことから、分離源の塩濃度 に合わせた分離条件の設定は有効で、効率的であるとい える。しかし一方で、分離源は全て酸性だったが、酸性 域のみならずアルカリ域の分離培地からも分離株が得ら れた(Fig.1)。高塩濃度の発酵食品からも、Tanasupawat ら(1998)6)の報告では分離されなかった好塩性でない Lb. plantarum が分離された。高塩濃度存在下で死滅はし ないが増殖が弱い菌は、分離培地に増殖に適さない濃度 の NaCl が含まれていると好塩性またはより耐塩性の強い 他の種に抑制されてしまう。しかし、生育に適した NaCl 無添加の培地を用いることにより強く速い増殖が促され、 また同時に好塩性の種の抑制に繋がったと推測される。 Enterococcus 属種が分離されたサンプルでは 5% 以下の NaCl 濃度条件下において、pH 8 ~ 10 でEnterococcus 属 が分離された一方、中性から酸性条件下ではLb.
farcimi-nis group、Lb. fermentum、Lb. plantarum group などが 分離された(Fig. 1 d, i, k)。耐塩性は弱いが高いアルカ リ域で生育可能なため24)、弱酸性から中性域を好む多く の種を抑えてアルカリ域で優先的に分離されたと考えられ る。Tetragenococcus halophilus は高塩濃度のサンプルか ら、15% NaCl と pH 7 ~ 10 の条件で分離された(Fig. 1 n)。好塩性であるT. halophilus は、その生育に高い塩濃 度と同時に高い pH を要求する23)。この種を、他の種が優 性種として存在している発酵食品から分離する場合、アル カリ域の分離培地を用いることによって他の種の増殖を抑 え、分離が容易になると考えられる。 また同じ種でも、同時に存在する他種によって、分離さ れる条件が異なる事が確認出来る。Lb. plantarum group、 P. acidilactici、Lb. acidipiscis、Staphylococcus 属 の 耐 塩 性はそれぞれ 10% 未満、10%、10 ~ 12%、10 ~ 15% であ ると報告されている20, 22, 25)。P. acidilactici と Lb. acidipi-scis が分離された条件に着目すると、NaCl 濃度が低い分 離条件でLb. plantarum group が優勢に分離されているサ
ンプルでは、P. acidilactici と Lb. acidipiscis は NaCl が低 濃度の条件では分離されずに高濃度条件下で分離された (Fig. 1 a, g, j)。一方、Staphylococcus 属が NaCl が高濃
度の条件下で優位に分離されたサンプルでは、P.
acidilac-tici と Lb. acidipiscis は NaCl が低濃度の条件下で分離さ
れた(Fig. 1 k, m)。低塩濃度にLb. plantarum group, 高
塩濃度にStaphylococcus 属が出現したサンプルでは、P. acidilactici と Lb. acidipiscis はその間の条件に挟まれるよ うに出現している(Fig. 1 l)。このように、必ずしも分離 源とする発酵食品の条件に合わせた分離条件だけではな く、分離源に合わせた条件を軸としながらも、ある程度分 離に用いる条件を変えることは、発酵食品の微生物叢を構 成する優性種以外の種の存在を明らかにするためには有効 であると考えられる。 4.タイ発酵食品中の乳酸菌の多様性 タイ発酵食品には、これまでに少なくとも 10 属 49 種の 乳酸菌が生息することが確認されている(Table 1)。多様 な食材や塩濃度、副原料の様々な組み合わせがあるため、
種は、Pla-ra(魚の発酵食品)から分離されたLb.
acidipi-scis と W. thailandensis 20)、Miang(発酵茶葉)から分離
されたE. camelliae、Lb. camelliae、Lb. thailandensis、P.
siamensis 28-29)、Mum(発酵ソーセージ)から分離され たE. thailandicus 30)、Kapi(エビ味噌)から分離された Alkalibacterium kapii 12)などがある。近年、乳酸菌の新 種探索や提案には、表現性状や 16S rRNA 遺伝子塩基配 列に基づく解析に加えて、ハウスキーピング遺伝子塩基配 列の解析や DNA フィンガープリンティング法など、より 解像度の高い手法が必須となってきている。最近新種報告 された、台湾の漬物(Fu-tsai)から分離されたLb. futsaii や中国の漬物から分離されたLb. xiangfangensis において
も、pheS 遺伝子や rpoA 遺伝子、dnaK 遺伝子に基づく解
析によって明確に種の識別を行っている31-32)。これらの種 タイの発酵食品は非常に多様であり、これまで用いられて きた一般的な分離条件だけでは、タイ発酵食品に生息す る乳酸菌を捕捉しきれるものではない。著者らの研究で は NaCl 濃度と pH の条件を変化させる事により、タイ発 酵食品に生息する乳酸菌種の多様性の一端を捉えた。しか し、例えば糖源など、検討すべき要素は多い。タイには熱 帯特有の果実やバナナの花などを用いた発酵食品も豊富で ある。糖源をグルコースからフルクトースに置き換えるこ とによってフルクトフィリック乳酸菌の存在を明らかにし た報告のように26-27)、グルコースを主体とした分離条件に 加えて異なる糖源の分離条件への適用は、タイ発酵食品中 の乳酸菌叢のさらなる多様性解明へも繋がることが期待さ れる。 これまでにタイの発酵食品を起源として新種報告された Fig. 1 (※続き)
参 考 文 献
1) 内村泰,高尾哲也,菊池孝治 , 新村洋一,岡田早苗,小原直弘,
Daengsubha, W., 小崎道雄 : タイ国発酵米麺(Khanom Jeen)
中の乳酸菌の同定,日本食品工業学会誌,38, 465-475 (1991). 2) 小崎道雄,岡田早苗,関達治 : タイの米酒-籾穀混合酒オウ
と精米酒サトー-,日本醸造協会誌,97,46-61 (2002). 3) 野田文雄 :
東南アジアの魚醤油,日本醸造協会誌,88,531-536 (1993).
4) Okada, S., Daengsubha, W., Uchimura, T., Ohara, N. and
Kozaki, M.: Flora of lactic acid bacteria in Miang produced
in Northern Thailand, J. Gen. Appl. Microbiol., 32, 57-65 (1986). 5) Tanasupawat, S., Thongsanit, J., Okada, S. and Komagata,
K.: Lactic acid bacteria isolated from soy sauce mash in
Thailand, J. Gen. Appl. Microbiol., 48, 201–209 (2002).
6) Tanasupawat, S., Okada, S. and Komagata, K.: Lactic acid bacteria found in fermented fish in Thailand, J. Gen. Appl. Microbiol., 44, 193–200 (1998).
7) Tanasupawat, S., Hashimoto, Y., Ezaki, T., Kozaki, M. and
Komagata, K.: Identification of Staphylococcus carnosus strains from fermented fish and soy sauce mash, J. Gen. Appl. Microbiol., 37, 479-494 (1991).
8) Tanasupawat, S., Hashimoto, Y., Ezaki, T., Kozaki, M. and
Komagata, K.: Staphylococcus piscifemzentans sp. nov., from fermented fish in Thailand, Int. J. Syst. Bacteriol., 42, 577-581 (1992).
9) Ishikawa, M., Nakajima, K., Yanagi, M., Yamamoto, Y. and
Yamasato, K.: Marinilactibacillus psychrotolerans gen. nov., sp. nov., a halophilic and alkaliphilic marine lactic acid bacte-rium isolated from marine organisms in temperate and sub-tropical areas of Japan, Int. J. Syst. Evol. Microbiol., 53, 711-720 (2003).
10) Ishikawa, M., Nakajima, K., Itamiya, Y., Furukawa, S.,
Ya-mamoto, Y. and Yamasato, K.: Halolactibacillus halophilus gen. nov., sp. nov. and Halolactibacillus miurensis sp. nov., halophilic and alkaliphilic marine lactic acid bacteria consti-tuting a phylogenetic lineage in Bacillus rRNA group 1, Int. J. Syst. Evol. Microbiol., 55, 2427-2439 (2005).
11) Ishikawa, M., Kodama, K., Yasuda, H., Okamoto-Kainuma,
A., Koizumi, Y. and Yamasato, K.: Presence of halophilic
and alkaliphilic lactic acid bacteria in various cheeses, Lett. Appl. Microbiol., 44, 308-313 (2007).
12) Ishikawa, M., Tanasupawat, S., Nakajima, K., Kanamori,
H., Ishizaki, S., Kodama, K., Okamoto-Kainuma, A., Koi-zumi, Y., Yamamoto, Y. and Yamasato, K.: Alkalibacterium thalassium sp. nov., Alkalibacterium pelagium sp. nov., Alka-libacterium putridalgicola sp. nov. and AlkaAlka-libacterium kapii sp. nov., slightly halophilic and alkaliphilic marine lactic acid
bacteria isolated from marine organisms and salted foods collected in Japan and Thailand, Int. J. Syst. Evol. Microbiol.,
59, 1215-1226 (2009).
13) 石川森夫 : 好塩性 ・ 好アルカリ性乳酸菌の多様性と特性,日 本食品微生物学会雑誌,26,49-59 (2009).
14) Tanasupawat, S.: Thai lactic acid bacteria: diversity and ap-plications, SWU. Sci. J., 25, 1-13 (2009).
15) Tanasupawat, S. and Komagata, K.: Lactic acid bacteria in fermented foods in Thailand, World J. Microbiol. Biotechnol.
11, 253-256 (1995). 16) 小崎道雄 : 東南アジアの伝統発酵食品に関する微生物学的研 究,日本食品工業学会誌,38,651-661 (1991). 17) 内村泰,新村洋一,小原直弘,小崎道雄 : タイ国産食酢醸造 に用いる餅麹ルクパンの微生物相,日本醸造協会誌,86,62-67 (1991).
18) Miyashita, M., Yukphan, P., Chaipitakchonlatarn, W.,
Mali-mas, T., Sugimoto, M., Yoshino, M., Potacharoen, W., Tana-supawat, S., Nakagawa, Y., Kirtikara, K., Tanticharoen, M. and Suzuki, K.: 16S rRNA gene sequence analysis of lactic
acid bacteria isolated from fermented foods in Thailand, Mi-crobiol. Cult. Coll., 28, 1-9 (2012).
19) Tanasupawat, S., Shida, O., Okada, S. and Komagata, K.: Lactobacillus acidipiscis sp. nov. and Weissella thailandensis sp. nov., isolated from fermented fish in Thailand, Int. J. Syst. Evol. Microbiol., 50, 1479-1485 (2000).
20) Hammes, W.P. and Hertel, C.: Genus I. Lactobacillus, p.465-511, Bergey’s Manual of Systematic Bacteriology Second Edition vol. 3, ed. by De Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.-H. & Whit-man, W.B., Springer, New York (2009).
21) Chen, Y.S., Miyashita, M., Suzuki, K., Sato, H., Hsu, J.S. &
Yanagida, F.: Lactobacillus pobuzihii sp. nov., isolated from pobuzihi (fermented cummingcordia), Int. J. Syst. Evol. Mi-crobiol., 60, 1914-1917 (2010).
22) Holzapfel, W.H., Franz, C.M.A.P., Ludwig, W. and Dicks,
L.M.T.: Genus III. Pediococcus, p.513-532, Bergey’s Manual of Systematic Bacteriology Second Edition vol. 3, ed. by De Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rain-ey, F.A., Schleifer, K.-H. & Whitman, W.B., Springer, New York (2009).
23) Dicks, L.M.T., Holzapfel, W.H., Satomi, M., Kimura, B. and
Fujii, T.: Genus III. Tetragenococcus, p.611-616, Bergey’s Manual of Systematic Bacteriology Second Edition vol. 3, ed. by De Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.-H. & Whitman, W.B., Springer, New York (2009).
24) Svec, P. and Devriese, L.A.: Genus I. Enterococcus, p.594-607.,
が含まれるLb. alimentarius-crustorum group や Lb. plan-tarum group のように、16S rRNA 遺伝子配列の類似度が 高いグループにおいても、これら解像度の高い遺伝子を用 いることで種の識別が可能となり、より詳細な微生物叢の 解明にも有効であると考えられる。 5.おわりに タイで伝統的に食されている発酵食品に生息する乳酸菌 は、年間を通じて高温多湿なタイにおける食品の保存性に 重要な役割を担っており、生み出される独特の風味や旨味 は、食品製造への応用が期待できる。また分離条件による 出現パターンの変化などの情報は、食品製造での安定した 品質の確保における微生物制御に利用可能な情報のひとつ と考えられる。伝統的に食されているタイ発酵食品中の乳 酸菌の多様性やその役割を解明することにより、有用な乳 酸菌を見いだし、情報を得るために、さらなる解析が必要 であると考える。
Bergey’s Manual of Systematic Bacteriology Second Edition vol. 3, ed. by De Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.-H. & Whitman, W.B., Springer, New York (2009).
25) Schleifer, K.-H. and Bell, J.A.: Genus I. Staphylococcus, p.392-421., Bergey’s Manual of Systematic Bacteriology Sec-ond Edition vol. 3, ed. by De Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.-H. & Whit-man, W.B., Springer, New York (2009).
26) Endo, A. and Okada, S.: Reclassification of the genus Leuco-nostoc and proposals of Fructobacillus fructosus gen. nov., comb. nov., Fructobacillus durionis comb. nov., Fructobacil-lus ficulneus comb. nov. and FructobacilFructobacil-lus pseudoficulneus comb. nov., Int. J. Syst. Evol. Microbiol., 58, 2195-2205 (2008). 27) Endo, A., Irisawa, T., Futagawa-Endo, Y., Sonomoto, K.,
Itoh, K., Takano, K., Okada, S. and Dicks, L.M.T.: Fructo-bacillus tropaeoli sp. nov., a fructophilic lactic acid bacterium isolated from a flower, Int. J. Syst. Evol. Microbiol., 61, 898-902 (2011).
28) Sukontasing, S., Tanasupawat, S., Moonmangmee, S., Lee,
J.S. and Suzuki, K.: Enterococcus camelliae sp. nov., isolated from fermented tea leaves in Thailand, Int. J. Syst. Evol. Mi-crobiol., 57, 2151-2154 (2007).
29) Tanasupawat, S., Pakdeeto, A., Thawai, C., Yukphan, P.
and Okada, S.: Identification of lactic acid bacteria from
fermented tea leaves (miang) in Thailand and proposals of Lactobacillus thailandensis sp. nov., Lactobacillus camelliae sp. nov., and Pediococcus siamensis sp. nov., J. Gen. Appl. Mi-crobiol., 53, 7-15 (2007).
30) Tanasupawat, S., Sukontasing, S. and Lee, J.S.: Enterococ-cus thailandiEnterococ-cus sp. nov., isolated from fermented sausage (‘mum’) in Thailand, Int. J. Syst. Evol. Microbiol., 58, 1630-1634 (2008).
31) Chao, S.H., Kudo, Y., Tsai, Y.C. and Watanabe, K.: Lactoba-cillus futsaii sp. nov., isolated from fu-tsai and suan-tsai, tra-ditional Taiwanese fermented mustard products, Int. J. Syst. Evol. Microbiol., 62, 489-494 (2012).
32) Gu, C.T., Wang, F., Li, C.Y., Liu, F. and Huo, G.C.: Lacto-bacillus xiangfangensis sp. nov., isolated from Chinese pickle, Int. J. Syst. Evol. Microbiol., 62, 860-863 (2012).
Diversity of lactic acid bacteria in fermented foods in Thailand.
Mika Miyashita, Yasuyoshi Nakagawa and Ken-ichiro Suzuki
NBRC Culture Collection, Biological Resource Center,
National Institute of Technology and Evaluation
Abstract
Various fermented foods are present in the Thailand where is kept at high temperature and humidity throughout the year, and these fermented foods give unique flavor and taste to Thai cuisine. Lactic acid bacteria are known to exist in these fermented foods, and it is conceivable that they produce unique flavor and play the role for bio-preservation in a climate of high temperature and high humidity. This paper reviews the diversity of lactic acid bacteria found in various Thai fermented foods.