目次
序論 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 1 第一章 過食による全身性のインスリン抵抗性と骨格筋におけるインスリン作 用減弱との関連 ・・・・・・・・・・・・・・・・・・・・・・・・・ 4 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 4 第一節 メタボリックシンドロームモデル SHR/NDmc-cp ラットにおける インスリン抵抗性と骨格筋における異所性脂肪細胞の肥大化と血管 内皮細胞の機能障害との関連 ・・・・・・・・・・・・・・・・・ 6 1 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 6 2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 8 2-1 実験動物 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 8 2-2 生化学分析 ・・・・・・・・・・・・・・・・・・・・・・・・・・ 8 2-3 組織化学分析 ・・・・・・・・・・・・・・・・・・・・・・・・・ 8 2-4 ウエスタンブロッティング ・・・・・・・・・・・・・・・・・・・ 8 2-5 その他計測 ・・・・・・・・・・・・・・・・・・・・・・・・・・ 9 2-6 統計処理 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 9 3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 10 3-1 身体および血液生化学パラメータ ・・・・・・・・・・・・・・・・ 10 3-2 腓腹筋内の異所性脂肪細胞 ・・・・・・・・・・・・・・・・・・・ 10 3-3 腓腹筋内の異所性脂肪細胞の特徴 ・・・・・・・・・・・・・・・・ 10 3-4 腓腹筋における血管内皮細胞のインスリン感受性関連タンパク質レベル 10 4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 15 第二節 メタボリックシンドロームモデル SHR/NDmc-cp ラットの骨格筋 におけるインスリン抵抗性とペルオキシソーム増殖因子活性化受容 体(PPAR)の標的であると推定される遺伝子発現との関連 ・・ 18 1 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 18ii 3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 24 3-1 身体および血液生化学パラメータ ・・・・・・・・・・・・・・・・ 24 3-2 WKY ラットおよび SHR/NDmc-cp ラットの腓腹筋における遺伝子発現解 析 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 24 3-3 SHR/NDmc-cp ラットの腓腹筋において発現上昇が観察された遺伝子の リアルタイム RT-PCR およびタンパク質レベルの解析 ・・・・・・・ 24 3-4 発現が増大していた遺伝子における PPRE 探索 ・・・・・・・・・・ 24 4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 32 第二章 第一節 マウスにおける DPP-4 阻害薬アナグリプチンによる軽度イン スリン抵抗性改善と肝臓における代謝変化 ・・・・・・・・ 34 1 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 34 2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 36 2-1 実験動物 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 36 2-2 生化学分析 ・・・・・・・・・・・・・・・・・・・・・・・・・・ 36 2-3 肝臓の総胆汁酸抽出および総コレステロール、総胆汁酸の測定 ・・・ 36 2-4 RNA 解析 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 37 2-5 ウエスタンブロッティング ・・・・・・・・・・・・・・・・・・・ 37 2-6 マイクロアレイ解析 ・・・・・・・・・・・・・・・・・・・・・・ 37 2-8 統計処理 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 37 3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 42 3-1 解剖時の身体および血液生化学パラメータ ・・・・・・・・・・・・ 42 3-2 インスリン負荷試験(ITT) ・・・・・・・・・・・・・・・・・・・ 42 3-3 肝臓の糖およびエネルギー代謝、インスリンシグナル関連遺伝子の発現 42 3-4 肝臓の脂質代謝関連遺伝子の発現 ・・・・・・・・・・・・・・・・ 42 3-5 マイクロアレイによる発現遺伝子の網羅的解析 ・・・・・・・・・・ 43 3-6 コレステロール代謝および胆汁酸代謝関連遺伝子の発現 ・・・・・・・ 43 4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 53 第二章 第一節 DPP-4 阻害薬アナグリプチンによるマウス腓腹筋におけるイ ンスリン抵抗性改善作用 ・・・・・・・・・・・・・・・・・ 55 1 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 55 2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 56 2-1 実験動物 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 56 2-2 RNA 解析 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 56 2-3 ウエスタンブロッティング ・・・・・・・・・・・・・・・・・・・ 56 2-4 統計処理 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 56
3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 58 3-1 解剖時の身体および血液生化学パラメータ ・・・・・・・・・・・・ 58 3-2 腓腹筋における糖および脂質代謝に関連する遺伝子およびタンパク質の 発現 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 58 3-3 腓腹筋におけるエネルギー代謝、インスリンシグナル、血管内皮機能に関 連する遺伝子の発現 ・・・・・・・・・・・・・・・・・・・・・・ 58 4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 61 第二章 第三節 レスベラトロールによるマウス末梢血白血球の酸化ストレス 低減効果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 62 1 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 62 2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 64 2-1 実験動物 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 64 2-2 生化学分析 ・・・・・・・・・・・・・・・・・・・・・・・・・・ 64 2-3 RNA 解析 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 64 2-4 血漿中の動脈硬化リスク因子タンパク質の測定 ・・・・・・・・・・ 64 2-5 統計処理 ・・・・・・・・・・・・・・・・・・・・・・・・・・・ 64 3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 67 3-1 解剖時の身体および血液生化学パラメータ ・・・・・・・・・・・・ 67 3-2 インスリン負荷試験(ITT) ・・・・・・・・・・・・・・・・・・・ 67 3-3 炎症および活性酸素種(ROS)代謝関連遺伝子の発現量 ・・・・・・ 67 3-4 血漿中の動脈硬化リスク因子タンパク質濃度 ・・・・・・・・・・・ 67 4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 71 第三章 2 型糖尿病患者におけるα-グルコシダーゼ阻害薬の先行薬から後発薬 への変更に伴う血糖振幅と血中の心血管疾患リスク因子濃度の低下と の関連 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 74 1 はじめに ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 74 2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 75
iv
4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 82
総括 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 85
引用文献 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 87
序論
ヒトは生命活動のため食事から栄養素を摂取する。この中で糖質は、消化管において α アミラーゼや二糖類水解酵素による分解を受けてグルコースとなり、小腸より吸収される。 小腸から門脈を経て肝臓へ入ると、グルコースはグリコーゲン合成および脂肪合成に利用 されるとともに、必要に応じて血糖として他の組織へ供給される。食事摂取によって上昇 した血糖はインスリンの作用を受けて、脂肪組織や骨格筋などのインスリン標的組織にお いて速やかに取り込まれ、正常値まで低下する。一方で、空腹時では糖新生によって正常 血糖が保たれる。肝臓は、摂食時に蓄積したグリコーゲンや、骨格筋から供給されるアラ ニンを基質として糖新生を行い、グルコースを供給する。この糖新生が亢進しすぎないよ う、インスリンは空腹時でも低量が分泌されている。したがって、糖代謝が正常に行なわ れるためには、インスリンの産生および分泌を行なう膵臓ランゲルハンス島β 細胞の機能 および、インスリン標的組織におけるインスリン感受性の維持が重要となる。 特に生活習慣の異常は、インスリン標的組織においてインスリン作用の減弱すなわち、 インスリン抵抗性を誘導する。インスリン抵抗性はメタボリックシンドロームおよび2 型 糖尿病の主な病態である。インスリン抵抗性の発症要因の1 つに肥満が挙げられる。肥満 は遺伝素因に過食や運動不足といった環境要因も加わり、摂取したエネルギーが脂肪とし て過剰に蓄積される状態のことである。脂肪組織では、脂肪細胞の肥大化および細胞数の 増大により組織の肥大化を引き起こす。肥大化した脂肪組織は、インスリン感受性やエネ ルギー代謝を促進するインスリン感受性ホルモンのアディポネクチンの産生および分泌が 低下する一方で、インスリン抵抗性や炎症を惹起するインスリン抵抗性惹起因子(TNF-α、 レジスチンなど)の産生および分泌が亢進する[1,2]。肝臓や骨格筋などの非脂肪組織では、 過食に伴い実質細胞内および組織周辺に細胞内脂肪滴もしくは脂肪細胞として蓄積する。 これらの脂肪組織以外における脂肪を総称して異所性脂肪という。これまでは、肥大化し た内臓脂肪組織から分泌されたインスリン抵抗性惹起因子が、末梢のインスリン感受性を 減弱すると考えられてきた。一方でこれら因子の血中濃度は、脂肪組織間質と比較して100 倍も低く[3]、末梢において作用するとは考えにくい。このため、肥大化した異所性脂肪細 胞より分泌される因子が末梢のインスリン抵抗性惹起に関与している可能性が考えられる。 インスリン抵抗性を惹起してもしばらくは膵β 細胞量の増加やインスリン分泌の亢進に よって正常血糖を保つことができる。しかしながら、この状態が続くと膵臓は次第に疲弊 し、β 細胞量やインスリン分泌能が低下して 2 型糖尿病を発症する。2 型糖尿病発症前で あっても、インスリン感受性の減弱による肝臓の糖新生の増大、骨格筋や脂肪組織のグル コース取り込みの低下により食後の高血糖状態が長引く、いわゆる耐糖能異常を引き起こ される。さらに、耐糖能異常は心血管疾患リスクを増大させることが大規模疫学調査によ要もある。 2 型糖尿病の薬物療法に用いられる薬剤は、注射によってインスリンを補うインスリン 製剤、インスリン分泌を促進するスルホニル尿素薬(SU 薬)やグリニド薬、糖新生を抑 制するビグアナイド薬(BG 薬)、インスリン感受性を増大させるチアゾリジン薬(TZD 薬)、食後高血糖を抑制する α-グルコシダーゼ阻害薬(α-GI)および DPP-4(dipeptidyl peptidase-4)阻害薬、尿からのグルコース排泄を促進する SGLT2 阻害薬などがある。イ ンスリン注射や SU 薬、グリニド薬は、血糖低下作用が強力である反面、低血糖のリスク が生じる。さらに、インスリンは脂肪組織における脂肪の合成および蓄積を促進するため 肥満を助長させる。加えて、インスリン製剤、インスリン分泌促進薬を長期にわたって使 用すると二次無効が生じやすくなる。肥満を助長させる薬剤としてはTZD 薬も含まれる。 最新の治療薬である SGLT2 阻害薬は、腎臓の近位尿細管においてグルコースを再吸収す る糖輸送単体 SGLT2 の作用を阻害し、グルコースを尿から排泄させる薬剤である。本薬 剤はインスリン非依存的に血糖を低下させるだけではなく、体重減少や脂質異常症の改善 などの副次的な効果もある。その一方で、多尿による脱水が脳梗塞や尿路感染症など引き 起こす副作用も報告されており、腎機能の低下した患者や高齢者に対しての適用は特に注 意が必要である。また SGLT2 阻害薬療法は、食事療法による食習慣の改善といった基本 的なことがおろそかになる可能性も考えられる。α-GI と DPP-4 阻害薬は異なる機序で食 後高血糖を抑制する。α-GI は小腸上皮細胞の二糖類水解酵素を競合阻害することにより、 でんぷんなどの多糖類の消化吸収を遅延させて食後高血糖を抑制する。米や小麦などので んぷんを多く摂取する日本人にとって有用な薬剤であると考えられる。一方でDPP-4 阻害 薬は、インクレチンの半減期を延長させることによって食後高血糖を抑制する。インクレ チンとは、食事摂取に伴い消化管から分泌されるペプチドで、膵β 細胞に作用し血糖依存 的 に イ ン ス リ ン 分 泌 を 促 進 す る ホ ル モ ン の 総 称 で あ る 。GIP ( glucose-dependent
insulinotripic polypeptide)および GLP-1(glucagon-like peptide-1)がインクレチンと して機能することが確認されている。これらインクレチンは、セリンプロテアーゼである DPP-4 により速やかに不活化されるため、半減期は数分といわれている。この DPP-4 を 選択的に阻害するのがDPP-4 阻害薬である。食後のインスリン分泌応答の 3 分の 2 はイ ンクレチン効果であることから、DPP-4 阻害薬による食後高血糖の抑制は 2 型糖尿病の治 療には重要といえる。さらにDPP-4 の作用を受けにくくした GLP-1 受容体作動薬(GLP-1 アナログ)も治療薬の1 つである。GLP-1 受容体作動薬は、平均血糖の指標であるヘモグ ロビンA1c(HbA1c)および体重を効果的に低下させる作用を有するが、注射による投与が 必要であることが難点である。 好気性細胞において、ROS やフリーラジカルが産生されるのは特別なことではない。特 にROS は、ミトコンドリアの呼吸鎖において産生され、細胞質へ漏出した電子により ROS が産生される。しかしながら、カタラーゼや SOD などの抗酸化酵素により速やかに除去 され、呼吸に伴い産生されるROS の毒性から守られている。一方で ROS は細胞内シグナ ル分子としての役割も持ち、遺伝子発現や免疫応答などと関わりがある[7]。このため、ROS 濃度が過剰に上昇しないよう、適度に保たれることが健康において望ましい。しかしなが ら、2 型糖尿病などの高血糖では ROS 産生が亢進する。この酸化ストレスの増大によって 各種酵素や細胞膜、LDL コレステロールなどが酸化され、タンパク質の機能低下、酸化 LDL コレステロールの蓄積により動脈硬化および関連する合併症の発症が促進されると 考えられている。抗酸化食品成分は、ROS による生体物質の酸化を抑制すると考えられて いる。したがって、2 型糖尿病発症の予防および進行抑制のためには、食事から積極的に 抗酸化食品成分を摂取することが望ましい。食品に含まれる抗酸化成分はトコフェノール 類、アスコルビン酸、カロテノイド類、ポリフェノール類などがある。この中でポリフェ
− −3 ノール類は緑茶やコーヒーなどの嗜好飲料、野菜や果物などに含まれている。日本人は野 菜や果物の摂取量が目標量に達していない、もしくは減少しているため[8]、ポリフェノー ル類の健康に対する有用性を研究することは、野菜や果物の積極摂取の動機付けとなると 考えられる。抗酸化食品成分の中でもレスベラトロールは、ワインに含まれることが知ら れており、フランス人において飽和脂肪酸を豊富に含む食事を摂取しているのにも関わら ず心血管疾患の罹患率が比較的低い[9]、いわゆる「フレンチパラドックス」との関連が指 摘されている。レスベラトロールが高血糖における酸化ストレスを低減させ、炎症[10]や 血糖値[11]を改善することが報告されているが、これらは主に膵 β 細胞や血管内皮細胞、 メサンギウム細胞などにおける作用であることが考察されており、末梢血白血球における 作用については未解明である。 以上の知見より、本研究では以下の仮説を立てた。 1 過食によるインスリン抵抗性の発症には、インスリン標的組織近傍から分泌される 異所性脂肪組織由来の液性因子が関与している。 2 インクレチン効果増大は、軽度インスリン抵抗性を改善する 3 抗酸化食品成分の摂取は、軽度インスリン抵抗性状態における末梢血白血球の酸化 ストレスを低減する 4 2 型糖尿病における食後高血糖の抑制は、動脈硬化および合併症の発症進展を抑制す る これらを検証するために、メタボリックシンドロームモデルSHR/NDmc-cp ラット、高 脂肪食誘導による軽度インスリン抵抗性マウス、2 型糖尿病患者における臨床介入試験を 行なった。 2.4. 非臨床試験の概括評価 アナグリプチン,代謝物及び類縁物質の構造一覧表 略称・略号 構造式 由来 アナグリプチン (未変化体,S 体) 原薬 SKL-12250 (プロリンアミド体) 代謝物 SKL-12277 (カルボン酸体) 代謝物 SKL-12320 (プロリン体) 代謝物 SKL-12339 (一水酸化体:2-ヒドロキ シメチル体) 代謝物 SKL-13776 (一水酸化体:3-ヒドロキ シメチル体) 代謝物 SKL-14687 1)ナトリウム塩 2)フリー体 代謝物 N H N N H O CH3 H3C O N N CN H N H3C N H N N H O CH3 H3C O N N CONH2 H N H3C OH H N N H O CH3 H3C O N N N H3C N H N N H O CH3 H3C O N N CO2H H N H3C N H N N H O CH3 H3C O N N CN H N HO O O CN H H N N N H CH3 H3C N N N HO H3C
Figure Structure of anagliptin.
Chemical name:
N-[2-({2-[(2S)-2-Cyanopyrrolidin-1-yl]-2-oxoethyl}amino)-2-methylpropyl]-2-methyl pyrazolo{1,5-α}pyramidine-6-carboxamide
Molecular formula: C19H25N7O2
第一章
過食による全身性のインスリン抵抗性と
骨格筋におけるインスリン作用減弱との関連
はじめに 糖尿病患者数は世界的に増加しており、20 歳から 79 歳の有病者数は 2013 年時点で 3 億 8 千万人を超え、2030 年には 6 億人に達すると予想されている。有病者数は低所得お よび中所得の国に集中している一方で、耐糖能異常者数は日本を含める先進国と経済成長 が著しい発展途上国の両方で増加している[12]。わが国においても有病者数は 720 万人で 世界第10 位、耐糖能異常者数は 1519 万人と推定されている。したがって、糖尿病予防は 世界的な健康課題であり、その発症要因を明らかにすることは予防策の発展につながると 考えられる。 糖尿病は、インスリン不足に基づく慢性の高血糖を主徴とする代謝疾患群であり、1型、 2 型、妊娠性、その他に分類される。わが国において糖尿病患者の 95 %以上は 2 型である。 1 型糖尿病は膵 β 細胞の破壊による絶対的インスリン欠乏が特徴であるが、2 型糖尿病は 膵β 細胞の機能低下およびインスリン標的組織におけるインスリン作用の低下、いわゆる インスリン抵抗性により、相対的なインスリン不足に陥る。2 型糖尿病の発症には、食事 や運動などの生活習慣を含む環境要因が大きく関与して肥満を引き起こし、そこに遺伝子 多型などによる遺伝素因が加わることでインスリン抵抗性が惹起されると考えられている。 2 型糖尿病発症との関連が示唆されている遺伝子多型はゲノムワイド相関解析(GWAS) により明らかになってきたが、その寄与に関しては個人差が大きく、環境因子との相互作 用についてもいまだ不明な点が多い。一方で食事や運動といった環境因子が2 型糖尿病発 症に関与することが疫学研究によって明らになっている[13,14,15]。 肥満は、エネルギーの摂取量が消費量を上回って過剰に蓄積された状態であり、運動不 足や過食により引き起こされる。食欲を制御する因子は血液中のグルコース、アミノ酸、 脂質の濃度だけではなく、ホルモンや神経伝達物質など複数が存在し、短期的および長期 的な調節を行う。これら因子の一つであるレプチンは、脂肪細胞で産生されるアディポカ インであり、視床下部に受容されて摂食を抑制する[16]。レプチン遺伝子あるいはレプチ ン受容体遺伝子の変異のある動物は、過食による肥満を呈するため、インスリン抵抗性お よび2 型糖尿病の自然発症モデルとして広く利用されている。これらの動物は肥満、イン スリン抵抗性、高血糖、脂質異常症などを発症するが、中でも SHR/N 系統ラットはイン スリン分泌能が障害されることなく食後高血糖を呈する。さらに、脂質異常症ならびに高 血圧を併発する。それゆえ、ヒトにおいて2 型糖尿病およびメタボリックシンドローム発 症前の耐糖能異常の病態に近いと考えられる[17]。一方で、レプチン遺伝子あるいはレプ チン受容体遺伝子の変異自体が糖尿病発症の直接的な要因ではないことも知られており [18]、インスリン抵抗性に与える過食の影響をみる上で、SHR/NDmc-cp ラットは適した 系統である。 村松らは、この SHR/NDmc-cp ラットの肝臓において、脂肪酸合成酵素(FASN)や、FASN の 発 現 を 制 御 す る 転 写 因 子 SREBP ( sterol regulatory element-binding transcription factor)1 の遺伝子発現およびタンパク質レベルが、対照の Wistar-Kyoto
(WKY)ラットおよびストレプトゾトシン(STZ)投与 WKY ラットと比較して、顕著に 高いことを見出した[19]。SHR/NDmc-cp ラットは高血糖および脂質異常症に加え、高イ ンスリン血症を呈したことから、同ラットにおける肝臓における脂肪酸合成関連遺伝子の 発現は、インスリン抵抗性に伴う血糖上昇およびインスリンの過剰分泌によって誘導され たと考えられる。このように、インスリン抵抗性に伴い肝臓で合成された脂肪酸は、中性 脂肪として骨格筋に運搬される。さらに、骨格筋においてもインスリン抵抗性時には脂肪 合成が誘導される。よってSHR/NDmc-cp ラットの骨格筋における脂質代謝の異常が、イ ンスリン抵抗性および2 型糖尿病の進展に関与している可能性が考えられる。そこで本章 では、インスリン抵抗性と骨格筋における脂質代謝との関連および、これらとインスリン 抵抗性誘導因子との関連を検討した。
第一章 第一節
メタボリックシンドロームモデル
SHR/NDmc-cp ラットにおける
インスリン抵抗性と骨格筋における異所性脂肪細胞の肥大化と
血管内皮細胞の機能障害との関連
1 はじめに 過食や高脂肪食などの食習慣は、インスリン抵抗性およびインスリン抵抗性を発症要因 とする疾患(2 型糖尿病、高血圧およびメタボリックシンドローム)の発症進展を促進す る。特に、骨格筋はインスリン分泌に伴う各組織のグルコースの取り込みの約75%を担う。 それゆえ骨格筋におけるグルコース取込み能の低下は、空腹時および食後の血糖値の上昇 をまねき、2 型糖尿病の発症進展を促進する[20]。したがって、健康的な食習慣によって 骨格筋のインスリン感受性を維持することは、2 型糖尿病の発症および進展を抑制する上 で重要である。 食事の摂取に伴いインスリンが分泌されると、骨格筋細胞内小胞にあるグルコース輸送 体 GLUT4 が細胞膜へ移行することによってグルコースが取り込まれる[21]。インスリン によるGLUT4 の膜移行は、インスリン受容体基質(IRS)-ホスファチジルイノシトール3-キナーゼ(PI3K)経路および AMP 活性化プロテインキナーゼ(AMPK)の経路によっ てシグナル伝達される[22]。最近の報告により、骨格筋内の血管内皮細胞におけるインス リン作用の減弱が、インスリン抵抗性発症の第一段階であることが示された。具体的には、 筋細胞へのインスリンの移行には、内皮型一酸化窒素合成酵素(eNOS)による NO 産生 による血管拡張が関与すること、高脂肪食摂取は、eNOS の活性を低下させることによっ て筋細胞へのインスリンの移行を抑制することが明らかにされた[23]。筋組織間質へのイ ンスリン移行の減少は、筋細胞においてインスリン依存的なシグナルカスケードを抑制し、 GLUT4 の膜移行および IRS-PI3K 活性を抑制する。しかしながら、インスリン抵抗性の 発症進展において、骨格筋組織の血管内皮細胞のインスリンシグナルの障害がどのような 因子によって誘導されるかはいまだ明らかになっていない。 これまでに内蔵脂肪組織の機能障害は、インスリン抵抗性発症の主要因であることが示 唆されている[24]。具体的には、インスリン作用が減弱した内臓脂肪組織からは腫瘍壊死 因子(TNF)-αおよびインターロイキン(IL)-6、レジスチンなどのアディポカインが分 泌されることが知られており、これらサイトカインの血中濃度は、インスリン抵抗性およ び 2 型糖尿病の発症進展と密接に関連することがヒト試験により報告されている[25,26]。 培養細胞を用いたin vitro 試験では、インスリン抵抗性を惹起させる IL-6 濃度は、脂肪細 胞において20-100 ng/mL[27,28]、肝細胞において 20 ng/mL[29]であることが報告されて いる。その一方で、IL-6 のヒト血液中の濃度は 40 pg/mL 未満である[3,30,31,32,33]。し たがって、インスリン標的組織のインスリン抵抗性は、内臓脂肪組織から分泌されるサイ トカインより、末梢組織近傍から分泌されるサイトカインによって誘導される可能性が高 いと考えられる。実際に、それぞれの代謝組織周囲には脂肪組織(異所性脂肪)が存在す ることがわかっている。これまでに、異所性脂肪は、心臓、冠動脈、他の動脈周囲に存在
することがヒトおよび動物モデルにおいて明らかとなっている[34,35,36,37,38]。さらに、 大動脈周囲の脂肪組織の除去が動脈硬化の進行を促進すること、大動脈周囲にインスリン 抵抗性を惹起した脂肪組織を移植すると動脈硬化が進行することが、動物モデルにおいて 明らかとなっている。この動脈硬化の進行には、血管周囲脂肪組織から分泌される単球遊 走促進因子(MCP)-1 などのインスリン抵抗性アディポカインや、アディポネクチンの分 泌不足などが関与していると考えられる[36,39]。それゆえ、インスリン抵抗性を惹起した 異所性脂肪組織は骨格筋においても、その近傍の筋組織のインスリン抵抗性を誘導する可 能性が高いと考えられる。ヒト骨格筋における異所性脂肪は、筋束間の脂肪細胞および筋 実質細胞内に蓄積しているが、この蓄積量とインスリン抵抗性には正の関連が観察される [40,41,42]。しかしながら、骨格筋内の血管周囲脂肪細胞がヒトあるいは動物モデルにお いてインスリン抵抗性の発症に直接関連しているかどうかは明らかではない。 そこで本節では、過食によるインスリン抵抗性により肥満、高血圧、脂質異常症、高血 糖を呈するメタボリックシンドロームモデルの SHR/NDmc-cp ラットとその対照である Wistar-Kyoto(WKY)ラット、ならびに WKY ラットにストレプトゾトシンを投与して 膵β 細胞を破壊し、インスリン不足による高血糖を呈するストレプトゾトシン(STZ)投 与ラットを用い、以下の仮説を検証した。 (1) 腓腹筋の異所性脂肪組織の存在 (2) SHR/NDmc-cp ラットでは、他のモデルと比較し腓腹筋内の異所性脂肪細胞が肥大 化している。 (3) SHR/NDmc-cp ラット腓腹筋内の異所性脂肪組織から分泌されるアディポカインの タンパク質レベルと血管内皮細胞の機能(eNOS の活性型レベル)は関連する。
2 材料と方法 2-1 実験動物 5 週齢雄性の SHR/NDmc-cp ラットおよび Wistar-Kyoto(WKY)ラットを日本エスエ ルシー株式会社より購入し、温度23 ± 2℃湿度 55 ± 5 %、明暗サイクル 12 時間の条件下 で飼育した。飼料は標準固形飼料(MF;オリエンタル酵母工業株式会社)を水とともに 自由摂取させた。16 週齢時に WKY ラットを 2 群に分け、一方を対照、もう一方を STZ 投与群とした。STZ 投与群には生理食塩水[0.9 %(w/v)食塩水]に溶解した STZ 40 mg / kg 体重を尾静脈より注入した。対照ラットおよび SHR/NDmc-cp ラットには溶媒のみを 注入した。5 日後の血糖値測定により、STZ 投与群では血糖値の上昇がみられなかったた め、STZ 60 mg / BW 体重を再び注入し、他の 2 群には溶媒のみを注入した。この 5 日後 の血糖値測定により、STZ 投与群では顕著な血糖上昇がみられた。18 週齢時に非空腹時 の血漿をヘパリンリチウム含有採血管(テルモ株式会社)にて尾静脈より午前9 時半から 10 時の間に採取した。ラットの解剖は午前 10 時から正午までの間に行い、腓腹筋を摘出 した。本研究は静岡県立大学の動物実験倫理員会のガイドラインに従って執り行われた。 2-2 生化学分析 血漿グルコースはグルコース CⅡ-テストワコー、血漿中性脂肪はトリグリセライド E-テストワコーを用いてそれぞれ濃度を測定した(和光純薬工業株式会社)。血漿インスリン 濃度は高感度ラットインスリン測定キット(株式会社森永生科学研究所)を用いて測定し た。 2-3 組織化学分析 腓腹筋を固定後にパラフィン包埋し、3-4 µm の厚さに切削し、スライドグラスにのせた。 脱パラフィン後に水洗し、カラッチヘマトキシリン(武藤化学株式会社)で 20 分間染色 し、流水で水洗した。1 % 塩酸アルコールに 30 秒間浸漬し、流水で水洗した。0.05 % 炭酸リチウムに 30 秒間浸漬し、流水で水洗の後に 70 % エタノールに浸し、1 % エ オジン Y 溶液(武藤化学株式会社)で 5 分間染色した。100 % エタノールに浸した後に 封入剤マリノール(武藤化学株式会社)にて封入した。これらの作業および撮影は株式会 社新組織科学研究所に委託した。 2-4 ウエスタンブロッティング タンパク質画分はRIPA バッファー〔1 % ノニデット P-40、0.1 % ラウリル硫酸ナト リウム、0.1 % デオキシコール酸ナトリウム、20 mM Tris-HCl pH 8.0、5 mM エチレン ジアミン四酢酸、150 mM NaCl、プロテアーゼ阻害剤タブレット(cOmplete, Mini, ロシ ュ・ダイアグノスティックス株式会社)/ 10 mL、ホスファターゼ阻害剤(1 mM NaMoO4、 50 mM NaF、1 mM Na3VO4)〕を用いて腓腹筋から抽出した。この抽出液を17,750×g で 30 分間、25 ℃で遠心分離した。上清のタンパク質濃度を Lowry 法[43]にて測定し、タン パク質濃度を揃え、解析に使用するまで−30 ℃で保管した。タンパク質は 10 % ドデシル 硫酸ナトリウムポリアクリルアミドゲル電気泳動で分子量別に分離し、トリス/グリシン /メタノール トランスバッファー内で Immobilon トランスファーメンブレン(メルク株 式会社)へ80 mA、90 分間転写した。メンブレンは 5 %スキムミルク(雪印メグミルク 株式会社)および 0.05 % Tween 20 を含むリン酸緩衝生理食塩水(PBS-Tween、pH 7.4) もしくは Blocking One-P(ナカライテスク株式会社)に室温で 1 時間以上浸してブロッ キングした。これに一次抗体を加えて4 ℃で 7 時間以上インキュベートした。一次抗体は
レジスチン(CST ジャパン株式会社)、α-Tubulin(シグマアルドリッチジャパン合同会社)、 eNOS(サンタクルーズバイオテクノロジー)、セリン 1177 リン酸化 eNOS(p-eNOS; サンタクルーズバイオテクノロジー)、AMPK α-サブユニット(AMPKα;CST ジャパン 株式会社)、スレオニン172 リン酸化 AMPKα(p-AMPK;CST ジャパン株式会社)を用 いた。PBT-Tween で洗浄後、ビオチン化抗ウサギ IgG もしくはビオチン化抗マウス抗体 (GE ヘルスケア・ジャパン株式会社)、西洋わさびペルオキシダーゼ(HRP)結合抗ヤギ IgG(サンタクルーズバイオテクノロジー)を 3 % スキムミルク PBS-Tween に加えて 4 ℃ にて7 時間以上インキュベートした。PBS-Tween で洗浄後、ビオチン化抗ウサギ IgG も しくはビオチン化抗マウス抗体でインキュベートしたメンブレンは HRP 結合抗ビオチン 抗体(CST ジャパン株式会社)を 3 % スキムミルク PBS-Tween に加えて 4 ℃にて 7 時 間以上インキュベートした。検出は化学発光(ウエスタンライトニング ECL プロ; PerkinElmer)を用いてルミノ・イメージアナライザー(Las 3000-mini、富士フィルム) にて視覚化した。 2-5 その他計測 体重は 2-3 日おきに測定した。腓腹筋のアディポネクチン濃度はラット ELISA アッセ イキット(CircuLex)を用いて測定し、タンパク質濃度で補正した。血管周囲脂肪細胞面 積は次式で算出した:(長径/2)×(短径/2)×3.14。 2-6 統計処理 結果は平均値 ± 標準誤差(SEM)で示した。検定はフィッシャーの最小有意差法によ る多重比較を行なった。有意水準5 % 未満を統計学的に有意とした。
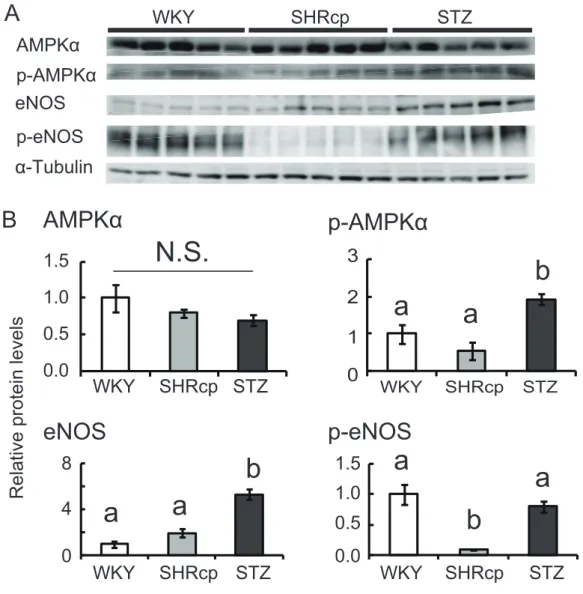
3 結果 3-1 身体および血液生化学パラメータ SHR/NDmc-cp 群の体重ならびに腸間膜脂肪重量、血漿中性脂肪およびインスリン濃度 は、対照群および STZ 投与群と比較して顕著に高かった。血漿グルコース濃度は、STZ 投与群が最も高く、続いてSHR/NDmc-cp 群、対照群の順となった(Table 1)。 3-2 腓腹筋内の異所性脂肪細胞 各群において血管の周囲に脂肪細胞が存在したが、筋束間には存在しなかった。 SHR/NDmc-cp 群における血管周囲脂肪細胞の面積は、他の 2 群よりも有意に大きかった。 腓腹筋の中性脂肪濃度も、SHR/NDmc-cp 群において他の 2 群よりも有意に高かった(Fig. 1)。 3-3 腓腹筋内の異所性脂肪細胞の特徴 腓腹筋内に存在する異所性脂肪細胞の性質を調べるため、インスリン抵抗性アディポカ インのレジスチンおよびインスリン感受性アディポカインのアディポネクチンの腓腹筋内 のタンパク質レベルを測定した。レジスチンのタンパク質レベルは、SHR/NDmc-cp 群に おいて他の 2 群よりも有意に高かった。一方でアディポネクチンのタンパク質レベルは 3 群間で差はみられなかった(Fig. 2)。 3-4 腓腹筋における血管内皮細胞のインスリン感受性関連タンパク質レベル 筋細胞および内皮細胞のインスリン感受性関連タンパク質の発現レベルを測定した。骨 格筋のインスリン感受性を調節するAMPKα のタンパク質レベルは 3 群間で差はみられな かった。活性型AMPKα である p-AMPKα のタンパク質レベル、は SHR/NDmc-cp 群と対 照群の間では差はなかったが、STZ 投与群のタンパク質レベルは、他の 2 群よりも有意に 高かった。内皮細胞特異的に発現するeNOS のタンパク質レベルは、3 群間で差はみられ なかった。活性型eNOS である p-eNOS のタンパク質レベルは、SHR/NDmc-cp 群におい て他の2 群よりも有意に低かった(Fig. 3)。
Body weight (g) 402 ± 3.5b 603 ± 9.4c 346 ± 5.0a
Plasma glucose (mg/dL) 172.5 ± 7.9a 326.0 ± 40.8b 532.1 ± 21.8c
Plasma triglyceride (mg/dL) 63.7 ± 6.4a 946.5 ± 94.0c 174.5 ± 29.3b
Plasma insulin (ng/mL) 2.10 ± 0.34b 14.01 ± 0.12c 0.61 ± 0.04a
Values are expressed as means ± SEM for 5–6 animals. a, b, c: Values not sharing a common superscript are significantly different (P <0.05) from one another.
Table 1
Body weight, liver weight, adipose tissue weight and blood biochemical parameters in normal WKY rats, SHR/NDmc-cp rats and STZ-treated WKY rats
WKY SHR/NDmc-cp STZ
Mesenteric adipose tissue
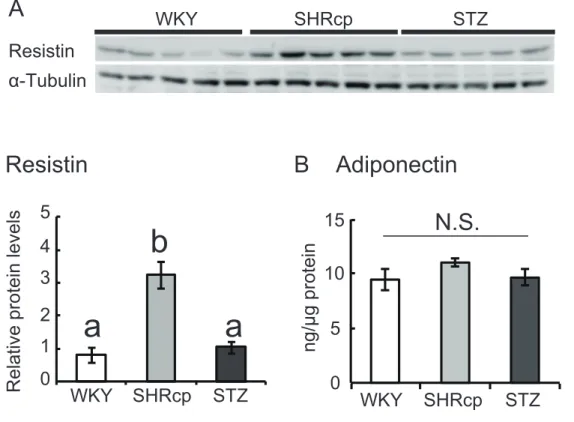
Fig. 1. Adipocyte morphology, adipocyte area, and triacylglycerol content in the gastrocnemius.
(A) Tissue sections stained with hematoxylin and eosin. Perivascular adipocytes are labeled with arrows. # = blood vessels. (B) Area of perivascular adipocytes. (C) Intramuscular triacylglycerol levels. Values in B and C are means ± SEM (n = 5–6). Values not sharing a common superscript are significantly different from one another (P < 0.05). WKY SHR/NDmc-cp STZ A B WKY SHR/ND STZ 500 1000 1500 Adipocytes area WKY STZ 0.3 0.2 0.1 0.0 C Triacylglycerol level g/g protein
a
a
a
a
b
b
2 # # # # # # # # # # # # # # # # # # # # # # # #Fig. 1
mc-cp SHR/NDmc-cpFig. 2. Adipocytokine protein expression in the gastrocnemius.
(A) Protein levels of resistin. Values were normalized for α-tubulin expression. (B) Protein levels of adiponectin. Values were normalized for those in control WKY rat. Values are means ± SEM (n = 5). Values not sharing a common superscript are significantly different from one another (P < 0.05). N.S.: not significant.
Resistin
3 1 2 4 5 0 WKY SHRcp STZRelative protein levels
Adiponectin
15 10 5 0 WKY SHRcp STZB
ng/ μg proteinN.S.
WKY SHRcp STZ Resistin α-TubulinA
a
a
b
Fig. 3. Expression of proteins associated with insulin sensitivity in the gastrocnemius.
(A) Protein levels of AMPKα, p-AMPKα (Thr172), eNOS, and p-eNOS (Ser1177). (B) Quantification of protein levels after normalization for α-tubulin. Values are means ± SEM (n = 5). Values not sharing a common superscript are significantly different from one another (P < 0.05)
AMPKα
WKY SHRcp STZ 1.5 1.0 0.5 0.0p-AMPKα
WKY SHRcp STZ 1 2 3N.S.
a
a
b
a
a
b
WKY SHRcp STZ 0 4 8eNOS
WKY SHRcp STZp-eNOS
1.5 1.0 0.5 0.0a
a
b
0Relative protein levels
AMPKα p-AMPKα eNOS p-eNOS WKY SHRcp STZ
B
A
α-Tubulin4 考察 本研究において、すべての群の腓腹筋において血管周囲に脂肪細胞が観察された。特に、 SHR/NDmc-cp 群で観察された血管周囲脂肪細胞は、対照群および STZ 投与群と比較して 顕著に肥大化していた。加えて、腓腹筋および血漿の中性脂肪レベルも、対照群およびSTZ 投与群と比較してSHR/NDmc-cp 群において有意に高かった。STZ 投与群の腓腹筋内の血 管周囲脂肪細胞がほとんど萎縮していたことをふまえると、SHR/NDmc-cp 群の血管周囲 脂肪細胞は、過剰に分泌されているインスリンが肥大化を促していると考えられた。イン スリン受容体と相互作用してインスリンシグナルを阻害するアディポカイン[44,45,46]で あるレジスチンのタンパク質レベルは SHR/NDmc-cp 群において顕著に高かった。一方、 インスリン感受性と関連のあるアディポカインのアディポネクチンのタンパク質レベルは 3 群間に差がみられなかったことは興味深い。レジスチンは内皮機能の不活化[47,48]、筋 細胞および脂肪細胞におけるグルコース代謝阻害[48,49]にも関与する。さらに、レジスチ ンは、インスリン刺激による血管拡張をインスリンの容量依存的に減弱させる[44]。した がって、骨格筋内の血管周囲脂肪細胞からレジスチンが分泌されることが、骨格筋のイン スリン抵抗性の発症に関与していると考えられる。 骨格筋におけるグルコースの取り込みは、インスリンが血管から骨格筋組織の間質へ移 行することが律速であることが近年報告された[50]。インスリンは骨格筋において一酸化 窒素(NO)依存的に血管拡張を誘導するが[51]、これは内皮細胞特異的に発現する eNOS の1177 番目のセリン残基が活性型 IRS-2 によってリン酸化されることによる[52]。IRS-2 欠損マウスでは、eNOS の不活化が引き起こり、骨格筋内の血管拡張およびグルコース取 り込み低下が観察される[23]。したがって、血管内皮細胞のインスリン感受性は、骨格筋 におけるインスリン依存的なグルコース取り込みの重要な因子であるといえる。本研究に おいては、SHR/NDmc-cp 群の腓腹筋におけるリン酸化 eNOS のタンパク質レベルが、他 の2 群と比較して顕著に低下していた。レジスチンは、内皮細胞においてインスリン誘導 性のeNOS のリン酸化を減少させることが報告されている[44]。加えて、ラットへのレジ スチンの投与は、eNOS のタンパク質レベルおよび一酸化窒素の産生を減少させることに より血管内皮の機能障害を誘導する[53]。したがって、脂肪細胞の肥大化およびレジスチ ンのタンパク質レベルの増大は、SHR/NDmc-cp ラットの骨格筋において eNOS の不活化 およびインスリン抵抗性惹起に関与している可能性が考えられる。レジスチンを含むイン スリン抵抗性を惹起するアディポカインは脂肪組織からも分泌されており、特に内臓脂肪 組織の関与についても考慮する必要がある[54]。しかしながら、これらのアディポカイン の濃度は脂肪組織においては2-10 ng / mL であるが、血液においては 40 pg / mL 未満で ある[3,33]。したがって、レジスチンは脂肪組織からではなく骨格筋の血管周囲脂肪細胞 から分泌され、骨格筋の eNOS 不活化およびインスリン抵抗性を誘導すると考えられる。 血管周囲脂肪細胞より分泌された TNF-α が大血管障害を引き起こし、動脈硬化に由来す
かった。この特性はSHR/NDmc-cp ラット特有である可能性があり、他の動物モデルにお いても調べる必要がある。しかしながら本研究においては血管周囲脂肪細胞の顕著な肥大 化が観察され、このことがインスリン抵抗性および血管内皮の機能障害と密接および正に 関連していた。ヒトを対象とした研究において、骨格筋のインスリン抵抗性は筋細胞外に 蓄積した脂質よりも筋細胞内に蓄積した脂質とより強い関連があることが報告されている [56]。さらに、持久トレーニングをしているアスリートにおける筋細胞内の脂質蓄積量は、 健常者および肥満者よりも高いことが報告されている[42]。骨格筋組織内の血管周囲脂肪 細胞が、骨格筋のインスリン抵抗性と関連があるかどうかについてヒトを対象とした研究 による報告は今のところない。したがって、インスリン抵抗性が血管周囲脂肪細胞の面積 やレジスチンのタンパク質レベルと関連があるかどうかを、生検などでヒトの骨格筋を採 取し調べる必要がある。 本研究では、インスリン抵抗性を発症したSHR/NDmc-cp ラットの腓腹筋では、対照の WKY ラットと比較し、肥大化した血管周囲脂肪細胞、高インスリン血症、血管内皮機能 (eNOS 活性化)の低下が観察された。SHR/NDmc-cp ラットは、レプチン受容体遺伝子 の突然変異を持つ動物である[57]。レプチン受容体の遺伝的変異が、直接的に血管周囲脂 肪細胞の肥大化およびレジスチンレベルの増大および血管内皮機能障害を引き起こしてい る可能性も考えられる。骨格筋内の内皮細胞におけるeNOS リン酸化は、インスリン刺激 に依存していることが報告されている[23]。したがって肥大化した血管周囲脂肪細胞は、 過食および高脂肪食によって誘導されていると考えることが妥当である。加えて、内臓脂 肪組織の肥大化した脂肪細胞はインスリン抵抗性の性質を呈し、この脂肪細胞から大量の レジスチンが分泌されることが報告されている[54]。これらのことより、過食によるイン スリンの過剰分泌は、骨格筋の血管周囲脂肪細胞の肥大化およびこの脂肪細胞のインスリ ン抵抗性惹起を誘導している可能性が考えられえる。インスリン抵抗性および肥大化を伴 った血管周囲脂肪細胞は、高レベルのレジスチン分泌能を有し、骨格筋の血管内皮のeNOS 活性化を阻害すると考えられる。高脂肪食の摂取あるいは過食が、骨格筋内の血管周囲脂 肪細胞の肥大化およびレジスチンの高発現を誘導するかどうかは新たな研究において解明 する必要がある。さらに、カロリー制限や、食物繊維や抗酸化食品成分などの機能性食品 成分の摂取が、骨格筋のインスリン抵抗性や血管脂肪細胞の肥大化、レジスチンレベルを 抑制し、リン酸化eNOS レベルを増大させるかどうかを肥満、インスリン抵抗性、2 型糖 尿病、メタボリックシンドロームなどの動物モデルを用いて検証する必要がある。 血管周囲脂肪細胞から分泌されたレジスチンは、骨格筋のインスリン抵抗性を直接的に 誘導する可能性が考えられる。レジスチンの投与[58]もしくは培養筋細胞へのレジスチン 発現プラスミド導入[59]は、グルコース取込みを低下させることが報告されている。これ 以外にも、IRS-2 欠損は、骨格筋中の末梢血管において eNOS の不活化を誘導することが 報告されている[23]。したがって、レジスチンは骨格筋において、血管内皮細胞と筋細胞 の両方のインスリン抵抗性を誘導していると考えられる。骨格筋のインスリン抵抗性の発 症進展において、血管内皮細胞および筋細胞のどちらが大きく寄与しているのかを調べる 必要がある。 腓腹筋組織の免疫染色では、レジスチンタンパク質を検出することができなかった。分 泌タンパク質は、固定や洗浄など組織化学染色工程において溶出しやすいためと考えられ る。レジスチン抗体が、組織切片と反応しなかった可能性も考えられる。レジスチンタン パク質が腓腹筋の血管周囲脂肪細胞で発現しているかどうか、また、SHR/NDmc-cp ラッ トの血管周囲脂肪細胞のレジスチンの発現が対照ラットと比較して高いかを、分泌タンパ ク質を除去しないような他の実験方法、例えばレーザーマイクロダイセクション法などを 用いて調べる必要がある。
STZ 投与 WKY ラットにおける AMPKα および eNOS のリン酸化レベルが、 SHR/NDcm-cp ラットおよび対照の WKY ラットと比較して顕著に高かった。STZ 投与 WKY ラットは膵 β 細胞からのインスリン分泌能が極めて低いことから、血中インスリン 濃度低下の代償としてAMPKα および eNOS タンパク質のリン酸化レベルが増大したと考 えられる。 以上をまとめると、本研究においてインスリン抵抗性 SHR/NDmc-cp ラット腓腹筋は、 肥大化した血管周囲脂肪細胞、レジスチンタンパク質の高発現、リン酸化eNOS の低発現 という特徴を呈していた。これらの結果より、インスリン抵抗性を惹起した血管周囲脂肪 細胞は、骨格筋において血管内皮機能障害を引き起こす重要な役割を担っている可能性が 示唆された。
第一章 第二節
メタボリックシンドロームモデル
SHR/NDmc-cp ラットの骨格筋にお
けるインスリン抵抗性とペルオキシソーム増殖因子活性化受容体
(PPAR)の標的と推定される遺伝子発現との関連
1 はじめに 骨格筋は、インスリン依存的な糖取り込みの約75%に寄与している。したがって、骨格 筋におけるグルコース取込み能の低下は、インスリン抵抗性および2 型糖尿病のような代 謝性疾患の発症進展を促進することとなる[20]。ヒトにおいて肥満をともなうインスリン 抵抗性は、高血糖だけではなく、高血圧や脂質異常症の発症とも関連する。肥満に加えて 空腹時血糖、空腹時血中脂質、血圧のうち、2 つ以上に異常がみられる場合は、メタボリ ックシンドローム(MetS)とよばれる。MetS を有する対象者は、心血管疾患(CVD)お よびCVD 関連死の発生が高くなることが報告されている[60]。したがって食習慣を管理す ることによって骨格筋のインスリン感受性を維持することは、MetS および MetS 関連疾 患の発症進展を予防する上で重要である。 骨格筋におけるグルコース取り込みは、グルコース輸送体GLUT4 を介して行なわれる。 GLUT4 は、細胞内小胞から細胞膜上へインスリン依存的に移行する[21]。インスリン誘 導性のGLUT4 の膜移行シグナルは、インスリン受容体基質-ホスファチジルイノシトール 3-キナーゼ経路および AMP 活性化プロテインキナーゼ経路を介することが報告されてい る[22]。インスリンシグナルは、骨格筋のグリコーゲン合成酵素の脱リン酸化を促進し、 グルコースをグリコーゲンに変換する。それゆえ、骨格筋におけるインスリンを介したグ ルコースの取り込みは、グリコーゲン合成および脂肪酸合成に関連した代謝関連遺伝子の 発現変化とは独立していると考えられる。 骨格筋における脂肪酸酸化は、筋肉自体のインスリン抵抗性と関連があることが多くの 研究によって示されている。実際に、中程度のインスリン抵抗性を有する対象者が運動す ると、インスリン抵抗性が抑制されるとともに、骨格筋においてカルニチンパルミトイル 転移酵素-1b(CPT1B)やペルオキシソーム増殖因子活性化受容体 γ コアクチベータ 1α (PPARGC1A)といった脂肪酸酸化に関連する遺伝子の発現が減少することが報告され ている[61]。さらに、PPARGC1A および CPT1B、脂肪酸酸化を活性化する核内受容体 PPARA などの脂肪酸酸化関連遺伝子の発現が、中程度のインスリン抵抗性と正に関連す るが、重度のインスリン抵抗性ではこの関連性がみられないことがMetS 対象者による研 究によって近年報告された[62]。これらの結果は、中程度のインスリン抵抗性時には、骨 格筋は脂肪酸をエネルギー源として用いるために脂肪酸酸化が亢進していることを示唆し ている。一方で、重度のインスリン抵抗性および中程度のインスリン抵抗性から重度のイ ンスリン抵抗性へ移行した2 型糖尿病患者の骨格筋では、脂肪酸酸化関連遺伝子の発現は 抑制されていると考えられる。動物を用いた研究により、インスリン抵抗性から2 型糖尿 病をすみやかに発症するZucker 糖尿病肥満ラット(ZDF)の骨格筋では、脂肪酸酸化関 連遺伝子の発現が対照ラットと比較して差が観察されないことが報告されている[63,64]。加えて、高脂肪食を8 週間与えて中程度のインスリン抵抗性を発症した C57BL/6J マウス の骨格筋においても、脂肪酸酸化関連遺伝子の発現は対照マウスと比較して差がないこと が報告された[65]。これらのように動物を用いた研究結果とヒト研究の結果は一致してい ない。それゆえ、ヒトの病態に近い動物モデルで、インスリン抵抗性の発症進展と骨格筋 における脂肪酸酸化関連および他の代謝関連遺伝子の発現との関連を検証する必要がある。 本節では、過食より肥満を呈し、脂質異常症、高血糖、高血圧を自然発症してMetS へ と進行するSHR/NDmc-cp ラット[66]の骨格筋においてマイクロアレイ解析を行い、脂肪 酸酸化関連遺伝子の発現亢進が観察されるかどうかを、対照の Wistar-Kyoto(WKY)ラ ットと比較することによって調べた。さらに、SHR/NDmc-cp ラット腓腹筋において発現 亢進が観察された遺伝子のプロモーター/エンハンサー領域に、脂肪酸の酸化や代謝を制 御する転写因子であるペルオキシソーム増殖因子活性化受容体(PPAR)の応答領域 (PPRE)が存在するかどうかを調べた。
2 材料と方法 2-1 実験動物 実験に用いた動物、飼育環境、与えた飼料、解剖および採血方法については第一章第一 節にて上述したとおりである。本研究は静岡県立大学の動物実験倫理員会のガイドライン に従って執り行われた。 2-2 マイクロアレイ解析 総RNA を Chomczynski らの方法[67]にて酸性グアニジンチオシアネートを用いて抽出 した。マイクロアレイ解析のために各群から3 匹ずつの総 RNA を用いた。総 RNA(300 ng)
の相補的RNA(cRNA)への変換およびビオチンラベル化は、Ambion WT Expression kit
(ライフテクノロジーズジャパン株式会社)およびGeneChip WT terminal labeling kit、
Eukaryotic poly-A RNA Control kit(アフィメトリクス・ジャパン株式会社)を用いた。
ビオチンラベル化cRNA(10 µg)をラット 1.0 ST arrays(アフィメトリクス・ジャパン 株式会社)にてハイブリダイゼーションし、染色および洗浄の後にGeneChip スキャナー 3000(アフィメトリクス・ジャパン株式会社)にてスポット画像を取り込んだ[68]。Partek Software(株式会社菱化システム)を用いて取り込んだスポットシグナルを数値化した。 この数値化シグナルを Partek のアルゴリズムに従い、グローバル正規化によって標準化 した。データは底が 2 の自然対数(ln)へ変換し、2 群間でこの比が 2 倍以上もしくは 2 分の 1 以下の遺伝子データのみを抽出した。これらの遺伝子は Student の t 検定を行い、 有意水準0.05 未満(P < 0.05)を統計学的に有意と見なし、有意データのみその後の解析 に使用した。シグナルは1,477 検出されたが、SHR/NDmc-cp ラットにおいて対照の WKY ラットと比較して発現が2 倍以上に増大していた遺伝子数が 42、2 分の 1 以下に発現が減 少していた遺伝子数が 33 であった。SHR/NDmc-cp ラットにおいて発現が上昇していた 遺伝子を以下の 9 つのグループに分けた:energy/metabolism、signal transduction、
transcription/chromatin 、 structural proteins 、 immune response 、 transport of nutrients/ions、stress、others、unknown。これらは、Gene Ontology(GO)annotation
および PubMed における遺伝子機能によって分類した。Table 3 に GO annotation と
PubMed ID を示した。栄養素の代謝と関連のある遺伝子は energy/metabolism へ分類し た。G タンパク質共役受容体、細胞膜受容体、リン酸化シグナルカスケードに関わる遺伝 子 は signal transduction へ 分 類 し た 。 転 写 因 子 お よ び 転 写 共 役 因 子 は transcription/chromatin へ分類した。栄養素およびイオンの細胞内への輸送に関わる遺伝 子はtransport of nutrients/ions へ分類した。活性酸素種および酸化物質の消去に関わる 遺伝子はstress へ分類した。 2-3 RNA 解析 SHR/NDmc-cp ラット(n = 6)および WKY ラット(n = 5)から抽出した総 RNA サン
プル(250 ng)を、SupseScriptTM Ⅲ Reverse transcriptase(ライフテクノロジーズジ
ャパン株式会社)を用いて添付の説明書に従い cDNA へと変換した。RNA レベルの定量
はLight Cycler 480 システム(ロシュ・ダイアグノスティックス株式会社)および SYBER
Green Ⅰ(ロシュ・ダイアグノスティックス株式会社)にて RNA を PCR 増幅させ、内 部標準および解析遺伝子の閾値サイクル数(CT 値)を測定した。得られた値はデルタ-デ ルタ法[69]を用い、内部標準遺伝子に対する解析遺伝子の相対発現量を算出した。リアル
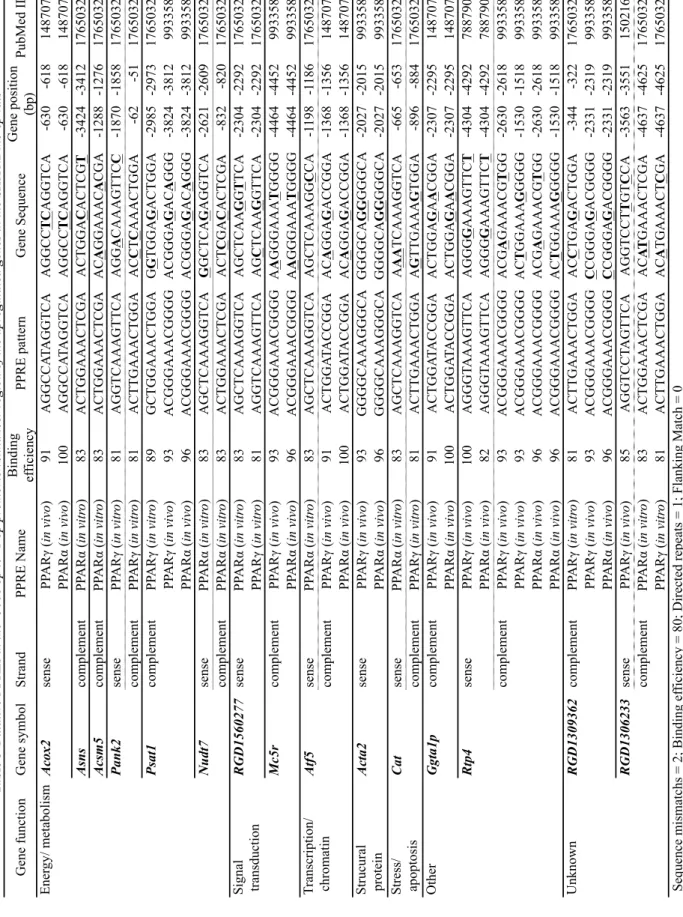
2-4 ウエスタンブロッティング 第一章第一節と同じ方法を用いた。一次抗体はA-TUBULIN(シグマアルドリッチジャ パン合同会社)、ACOX2(ProteinTech)、PPARA(サンタクルーズバイオテクノロジー)、 PPARD(サンタクルーズバイオテクノロジー)、PPARG(CST ジャパン株式会社)を用 いた。 2-5 高発現遺伝子の上流域における PPAR 応答領域(PPRE)の探索 マイクロアレイ解析より、SHR/NDmc-cpラットにおいてWKYラットよりも発現が高い遺 伝子のプロモーター/エンハンサー(-5000 bpから-1 bp)領域におけるPPREの探索は、 「PPRESeach(http://www.classicrus.com/PPRE/)」プログラム[70,71]を利用した。解析 基準は、結合有効性(binding efficacy)を80 %以上、ダイレクトリピート (AGGTCA-N-AGGTCA)が1つ、かつダイレクトリピートの5’側隣接領域の7塩基配列 〔C(A/G)(A/G)A(A/T)CT〕を考慮しない(zero flanking match)とした。5’側隣接領域の塩
基配列によりPPARサブタイプのDNAへの結合度が変化する[72]ことが知られているが、本
研究においてはPPREのみを探索の対象とした。
2-6 統計処理
結果は平均値 ± 標準誤差(SEM)で示した。異なる2群間の比較は、Studentのt検定で
Gene Sequence 5'-TGTGAGCTATGCCTTCCACTT-3' 5'-GCCTTCATACCAGTGCTCAAT-3' 5'-TTCTGGCTGTGTGTTCAGAAG-3' 5'-GGGCTCCACTTTTAGGAAGG-3' 5'-CTTGCGTTTTTGTGCATGAG-3' 5'-TTATTGGGAACCTGCAGAGAG-3' 5'-GACCTTGCCAGAGCAACTTT-3' 5'-TTGCCGACAAATACAACCTG-3' 5'-TTGAAGAAGCCGTGAACTACC-3' 5'-GCCATTGATGACAACAGCAG-3' 5'-GGAATGGATCAAGAACAATGG-3' 5'-CGGAATGTTCATCCTGCTTC-3' 5'-TCCCACTTGTCCCCTAGTAAGT-3' 5'-CAGGTGCCCTTCTCAGCTT-3' 5'-CTAGGATATCCAACTCAGGTTGC-3' 5'-CGTAGACAAAGTCGGGGATG-3' 5'-TTCGACTCCATGATCTGCAT-3' 5'-ATGTACCTGTCCACCGCAAT-3' 5'-TCAAGCCCTAAGTGAAGACCA-3' 5'-TGGAATTCGGCAGAGTTGAT-3' 5'-CTGCTCACACCGACTCTTCA-3' 5'-GTGAAGACAGGCACCAAGG-3' 5'-GCCAGGAGTGTAAGAAGACCA-3' 5'-CCTTAGGTATGAAGCTCTTGGTTC-3' 5'-CTTTGCTGAAATGCCACGTA-3' 5'-GGATCACGAGAGCCATAAAGA-3' 5'-TTCATCAGGGATGCCATGT-3' 5'-TGATGGAGAGACTCTGGACAAA-3' 5'-GCTCTGGGACTGGTTCAATC-3' 5'-AAGTGCCTTCCCACACAATC-3' 5'-GGACCCTGCACTTGGATAAG-3' 5'-CATCTGGAACACTGGAACCTG-3' 5'-TACACCGCTCAATGTTGCTG-3' 5'-TCTCCACCACCCCAATTTTA-3' 5'-AAGAAAACCCCCAAGAGTGC-3' 5'-TTTCTCTGGGGCAGGCTTA-3' 5'-GCAGGCCGACTGTCTTTTT-3' 5'-TGACCCCAAAATAGGAGAGC-3' 5'-TCTTTTCAGCTGTGGACATCA-3' 5'-TGTTTTCCACAAGAGCGATTC-3' 5'-TCTACCAACCAGATGCTGACA-3' 5'-CCAGGTGGTAGTTCCCAAAA-3' Acyl-CoA thioesterase 2 (Acot2)
Table 2 Sequences of oligonucleotide primers used for real-time RT-PCR Acyl-CoA oxidase 2, branched chain (Acox2)
Asparagine synthetase (glutamine-hydrolyzing) (Asns)
Acyl-CoA synthetase medium-chain family member 5 (Acsm5)
Pantothenate kinase 2 (Pank2)
Similar to interferon-inducible GTPase (RGD1309362)
Phosphoserine aminotransferase 1 (Psat1)
Nudix (nucleoside diphosphate linked moiety X)-type motif 7 (Nudt7)
GDP-D-glucose phosphorylase 1 (RDG1560277) Melanocortin 5 receptor (Mc5r)
Phospholipase A2, group VII (platelet-activating factor acetylhydrolase, plasma) (Pla2g7)
Activating transcription factor 5 (Atf5) Smooth muscle alpha-actin (Acta2)
Interferon-induced protein with tetratricopeptide repeats 1 (Ifit1)
Catalase (Cat)
Glycoprotein, alpha-galactosyltransferase 1,3 (Ggta1p)
Receptor (chemosensory) transporter protein 4 (Rtp4)
Similar to hypothetical protein MGC29761 (RGD1306233)
Retinol dehydrogenase 16 (all-trans) (Rdh16) Phospholipase D family, member 5 (Pld5) Solute carrier family 7 (amino acid transporter light chain, L system), member 5 (Slc7a5)
Gene Sequence 5'-CACCTTCGAGGGAGAGAACA-3' 5'-CCGCACCTGGTCATAGATTT-3' 5'-GCTGCTTATGGATGACCTCAA-3' 5'-TCACTGACGTGTTTGCTTGTC-3' 5'-ACTGTTACAACCCTGATGACGA-3' 5'-AAAGATTTGTCGAACCACCTG-3' 5'-CCTGGGAAAATTCTGTCACTG-3' 5'-TGGTGTTTAGCATTGCCTGTC-3' 5'-CGCAGGAAAGACTAGCAACA-3' 5'-TGGCAGCAGTGGAAGAATC-3' 5'-ATAACGCACCCTTCATCATC-3' 5'-GAGGAAGAGGCTGCTGAAGT-3' 5'-AATGCGATCTCTGTGTCAACC-3' 5'-GCTGCAGCGCTAAATTCATCT-3' 5'-ATGGGTCAAACTCTGGCACAT-3' 5'-AATGGCATCTCTGTGTCAACC-3' 5'-TACAGGAGCGGCAAGTTTTGA-3' 5'-ATTGCGTGGGAGGTTGATTCT-3' General transcription factor IIB (Gtf2b)
Acyl-CoA oxidase 1, palmitoyl (Acox1)
Acyl-CoA synthetase long-chain family member 1 (Acsl1)
Carnitine palmitoyltransferase 1b, muscle (Cpt1b) Cytochrom c, somatic (Cycs)
Peroxisome proliferator activated receptor alpha (Ppara)
Peroxisome proliferator activated receptor delta (Ppard)
Peroxisome proliferator activated receptor gamma 1 (Pparg1)
Peroxisome proliferator activated receptor gamma 2 (Pparg2)
3 結果 3-1 身体および血液生化学パラメータ 第一章第一節ではSTZ 投与 WKY ラットも含めた 3 群間での比較を行なったが、本節で はSHR/NDmc-cp ラットと対照 WKY ラットとの 2 群間の比較を行なった。SHR/NDmc-cp ラットにおける体重、血漿のインスリン濃度および中性脂肪濃度、血糖値は、対照のWKY ラットよりも有意に高かった(Table 3)。 3-2 WKY ラットおよび SHR/NDmc-cp ラットの腓腹筋における遺伝子発現解析 WKY ラットおよび SHR/NDmc-cp ラットの腓腹筋より抽出した総 RNA を用いてマイ クロアレイ解析を行なった。WKY ラットと比較して SHR/NDmc-cp ラットにおいて発現 が高かった 42 遺伝子のうち、各カテゴリへ分類された遺伝子数は以下の通りであった:
energy/metabolism は 14 遺 伝 子 、 signal transduction は 6 遺 伝 子 、 transcription/chromatin は 2 遺伝子、structural proteins は 1 遺伝子、immune response
は 3 遺伝子、transport nutrients/ions は 3 遺伝子、stress は 2 遺伝子、other および
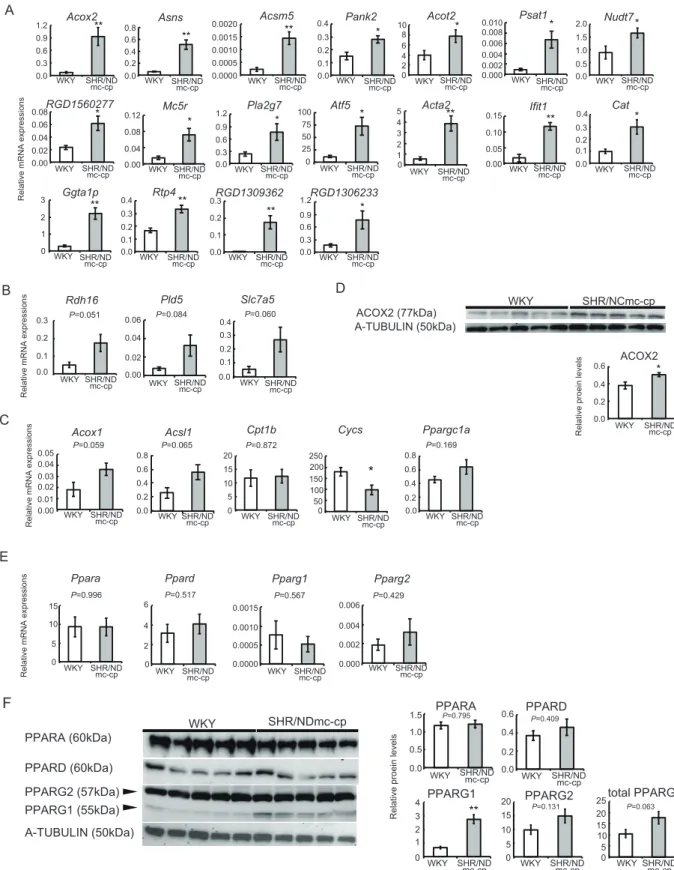
unknown は 11 遺伝子(Table 4)。 3-3 SHR/NDmc-cp ラットの腓腹筋において発現が高かった遺伝子のリアルタ イム RT-PCR およびタンパク質レベルの解析 上述の遺伝子についてリアルタイムRT-PCR を行い、遺伝子発現レベルを測定した。上 述の42 遺伝子のうち、対照の WKY ラットよりも SHR/NDmc-cp において発現量が有意 に高かったものは以下の 18 遺伝子であった:energy/metabolism カテゴリでは Acox2、 Asns、Acsm5、Pank2、Acot2、Psat1、Nudt7、signal transduction カテゴリでは RGD1560277、Mc5r、Pla2g7、transcription/chromatin カテゴリでは Atf5、structural
protein ではActa2、immune response カテゴリでは Ifit1、stress カテゴリでは Cat、other
カテゴリおよびunknown カテゴリではGgta1p、Rtp4、RGD1309362、RGD1306233(Fig.
2A)。有意な差は観察されなかったものの、SHR/NDmc-cp ラットにおいて WKY ラット よりも発現が高い傾向にあった遺伝子は以下のとおりであった:energy/metabolism カテ
ゴリのRdh16(P = 0.051)、signal transduction カテゴリの Pld5(P = 0.084)、transport
nutrients/ions カテゴリのSlc7a5(P = 0.060)(Fig. 2B)。脂肪酸酸化関連遺伝子である
アシル CoA オキシダーゼ 1(Acox1)、長鎖アシル CoA 合成酵素 1(Acsl1)、Cpt1b、シ
トクロムc(Cycs)、Ppargc1a の発現量を調べたところ、SHR/NDmc-cp ラットにおいて
WKY ラットよりもAcox1 および Acsl1 の遺伝子発現量が高い傾向にあったものの、有意
ではなかった(Fig. 2C)。ACOX2 タンパク質レベルは、SHR/NDmc-cp ラットにおいて WKY ラットよりも有意に高かった(Fig. 2D)。 PPAR 各サブタイプ(Ppara、Ppard、Pparg1、Pparg2)の遺伝子発現量は、SHR/NDmc-cp ラットおよび WKY ラットの間で差はみられなかった(Fig.1E)。PPAR 各サブタイプで は、PPARG1 タンパク質の SHR/NDmc-cp ラットにおける発現が、WKY よりも有意に高 かった。他のサブタイプのタンパク質レベルは、両群に差は見られなかった(Fig. 2F)。 3-4 発現が高かった遺伝子における PPRE 探索 マイクロアレイおよびRT-PCR の解析結果より、SHR/NDmc-cp ラットにおいて WKY ラットよりも発現が有意に高かった 18 遺伝子の上流域(-5000 bp から-1 bp)を、
PPRESearch プログラム[70,71]にて PPRE を探索した。PPRE の存在が推定される遺伝
energy/metabolism カテゴリのAcox2、Asns、Acsm5、Pank2、Nudt7、signal transduction
カテゴリのRGD1560277、Mc5r、transcription/chromarin カテゴリの Atf5、structural
protein の Acta、stress カテゴリの Cat、other および unknown の Ggta1p、Rtp4、
Body weight (g) 402 ± 7.88 603 ± 23.00 **
Plasma triglyceride (mg/dL) 64 ± 14.22 946 ± 230.36 **
Plasma glucose (mg/dL) 172 ± 17.56 326 ± 99.91 **
Plasma insulin (ng/mL) 2.10 ± 0.75 14.01 ± 0.29 **
Values are indicated as mean ± SD of 5–6 animals.
**P < 0.01 by student's t-test
WKY SHR/NDmc-cp