好熱性細菌
Thermus thermophilus 無機ピロリン酸加水分解酵素の
error-prone PCR 変異導入による熱凝集抑制
杉山 典子、前田 歩、田中 浩史、花房 英一郎、金井 拓、佐藤 高則
*徳島大学大学院ソシオ・アーツ・アンド・サイエンス研究部生物化学研究室 770-8502 徳島市南常三島町 1-1
Thermus thermophilus Inorganic Pyrophosphatase Variants
by Error-prone PCR Suppressed Thermal Aggregation
Noriko SUGIYAMA
1, Ayumi MAEDA
1, Hiroshi TAKANA
1,
Eiichiro HANAFUSA
2, Taku KANAI
2and Takanori SATOH
3,*
1Faculty of Integrated Arts and Sciences, The University of Tokushima, 2 Graduate school of Human and Natural
Environment Sciences, The University of Tokushima, 3 Institute of Integrated Arts and Sciences, The University
of Tokushima, Tokushima city, Tokushima 770-8502, Japan
*To whom correspondence should be addressed. e-mail: [email protected]
ABSTRACT
Thermus thermophilus Inorganic pyrophosphatase (E.C. 3.6.1.1., Tth PPase) exhibits high thermostability, but thermal aggregation was observed on heating above 85 °C. In addition, we reported that sole cysteine in C-terminal region plays a key role in the thermostability and thermal aggregation of Tth PPase [Kohaku, Y. et al. (2008) Natl.Sci.Res., 22, 75-84]. In this study, we approached the suppression of its thermal aggregation by
error-prone PCR mutagenesis of whole molecule or C-terminal region. Firstly, we obtained thermostable four variants (Q119H/L162F, L162F, K173E and K159E/A170T) by error-prone PCR mutagenesis. Moreover, we examined thermostabilities of four variants in terms of the enzyme activity, tertiary and quaternary structure.
Although conformation and quaternary structure of four variants were almost the same as those of wild type enzyme in native state, K173E and K159E/A170T variants showed higher thermostabilities than wild type in tertiary and quaternary structure. In particular, thermal aggregation of these two variants would be suppressed after heating at 85°C. Therefore, it was suggested that Lys159 and Lys173 in the molecular surface of C-terminal region may contribute to the formation of thermal aggregation of Tth PPase.
Keywords: Thermophile, Pyrophosphatase, Error-prone PCR, Directed evolution, Thermus thermophilus
__________________________________________________________________________________________
1. 緒言
本研究対象である無機ピロリン酸加水分解酵 素(PPase)は動物から植物まで広く存在し、Mg2+ などの二価金属イオンの存在下で 1 分子の無機 ピロリン酸を 2 分子の無機リン酸に加水分解す る反応を触媒する酵素であり、DNA や RNA 合成 の駆動力として働く1)。これまでに原核生物由来のPPase として Eschericia.coli (E.coli PPase)2)、高
PPase)3)、中等度好熱性細菌Thermophilic bacterium PS‐34)、Bacillus stearothermophilus (Bst PPase) 5) 由来のPPase などが報告されており、さらに古細 菌 Sulfolobus acidocaldarius 6) や Pyrococcus horikoshii PPase 7)の三次元立体構造と機能の相関 についても報告されている。このうち本研究対象 であるTth PPase は、一次構造および X 線結晶解 析による三次元立体構造が既に解明されており、 単量体あたり174 アミノ酸残基からなる 6 量体を
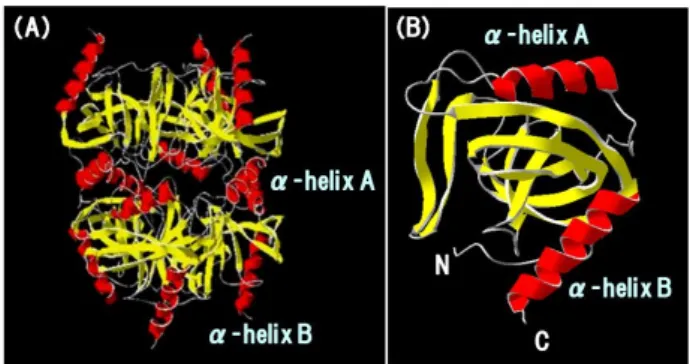
Fig. 1 Three-dimensional structure of Tth PPase The three-dimensional structure of hexameric (A) and monomeric (B) Tth PPase. The figure was drawn by a Molecular Graphics Program, Swiss-Pdb Viewer Ver.4.0 (Swiss Institute of Bioinformatics). The X-ray structural data for Tth PPase was obtained from the
Protein Data Bank (PDB code 2PRD) 8).
の加熱後も酵素活性および四次構造が安定であ り高い耐熱性を示すが、85℃以上の加熱で熱凝集 を起こすことが報告されており3), 10), 11)、この熱凝 集に関与する部位に関する知見はほとんど無い。 この観点から我々は、Tth PPase 分子表面に存在 する2本のα-helix(A および B)に着目し、このう ちα-helix A は 6 量体の 3 量体―3 量体界面を形 成し、Thr138 や Ala141 などのアミノ酸残基側鎖 同士の相互作用が高温における四次構造形成能 に関与することを明らかとした12)。さらに3 量体 ―3 量体界面の Thr138 と Ala141 を His138 と Asp141 に改変することで三量体界面のイオン性 が増し、三量体間界面が安定化することによって 六量体の安定性が向上することや 13)、このα -helix A に続く flexible loop の Ala144-Lys145 領域 を欠損させると熱凝集が抑制されることも明ら かにしている11)。 一方、C 末端側にあるα-helix B は変性の開始 点と考えられており、熱変性で露出した Cys168 が他分子と架橋し高分子化することで、熱凝集を 引き起こす可能性が示唆されている。この部位を Ala に置換した C168A 変異体は、熱凝集を抑制す ることが報告されているが14)、その他の熱凝集生 成または抑制因子については明らかではない。そ こで本研究では、α-helix B を含む C 末端領域 において熱安定性や熱凝集に関与するアミノ酸 残 基 を 新 規 に 探 索 す る た め 、 進 化 工 学 的 手 法 (error-prone PCR)15)を用いてこの領域にランダム 突然変異を導入後、スクリーニングにより耐熱性 を示す変異型酵素を選別し、得られた変異型酵素 の変異箇所と熱安定性および熱凝集との相関を 検討した。
2. 実験方法
試 薬 お よ び 材 料 モリブデン酸アンモニウム四水和物、 L‐ア スコルビン酸ナトリウム、ピロリン酸ナトリウ ム・10 水和物、トリス(ヒドロキシメチル)ア ミノメタン、 酢酸ナトリウム三水和物は、和光 純薬工業より購入した。T4 DNA ligation Kit ver.2 は宝酒造より、制限酵素は Promega, New England Bio Labs および宝酒造から購入した。塩化マグネ シウム六水和物は片山化学より、DEAE-cellulose は 和 光 純 薬 工 業 よ り 、Sephacryl S-200HR は Pharmacia よりそれぞれ購入した。 その他の試薬 はすべて特級品を使用した。 Error-prone PCR による Tth PPase 遺伝子への ラ ン ダ ム 突 然 変 異 導 入 ランダム変異導入にはerror-prone PCR 法15)を 用いた。通常の PCR 反応液の組成に、Mn2+、 dATP/dTTP、または dCTP/dGTP を各々の最終濃 度が 0.1∼0.5mM、1mM になるよう加え、Tth PPase が 挿 入 さ れ た プ ラ ス ミ ド ベ ク タ ー pUCTPPCR を鋳型として、Taq DNA Polymerase (Promega)により PCR (94℃, 1 分→61℃, 1 分→ 74℃, 1 分を 30 サイクル)を行った。PCR プライ マーは、upper プライマーが 5’-GACCTACAAGG CCCTCGAGGCCAAGAA-3’ (No.5 primer; 27mer, Tth PPase の Glu137─Lys146 に 相 当 ) ま た は 5’-GTGTGGAATTGTGAGCGGATAAC-3’ (No.8 primer; 23mer, pUCTPPCR の Tth PPase 遺伝子の上 流 側 の プ ラ ス ミ ド 内 塩 基 配 列 に 相 当) 、 lower primer が、5’-TGCTGCAAGGCGATTAAGTTGGG TAAC-3’ (No.9 primer; 27mer, pUCTPPCR の Tth PPase 遺伝子の下流側のプラスミド内塩基配列に 相当) を用いた。得られた遺伝子増幅断片を、Tth PPase の発現ベクターpETTP の対応する領域と入 れ換え、変異型発現ベクターpETTPEPCR を構築 した。 変 異 型 酵 素 の ス ク リ ー ニ ン グ 変異型発現ベクターを大腸菌 BL21(DE3)株で 形質転換し、LB/Amp 寒天培地上で生育させたコ ロニーを初期ライブラリーとし、各コロニーを 96 穴プレートの LB/Amp 液体培地に植菌し、37℃ で16h 培養した。培養液に Bugbuster (Novagen) を加え25℃で 20 分間穏やかに振盪して大腸菌を 溶菌し、粗抽出液とした。次にこの96 穴プレー トを各条件にて加熱した後、氷上で10 min 冷却 し、上清の活性測定を行った 16)。96 穴プレート に1mM MgCl2と 1mM PPiを含む 80mM Tris‐ HCl 緩衝液(pH8.0) 25μl に上清(酵素溶液)を3 μl 加え、37℃で 10 分間保温した後、3%(v/v)過 塩素酸溶液を25μl 加えて懸濁し、酵素反応を停止させた。次に 150μl の 5μM 硫酸銅溶液、15 μl の 1%(w/v)モリブデン酸アンモニウム溶液、 10μl の 1%(w/v) L‐アスコルビン酸ナトリウム 溶液を加え、10 分間放置後、発色の強度のより 強いクローンを選別した。選別したクローンより プラスミドDNA を調製し、DNA 塩基配列決定を 行い、変異部位の同定を行った。 野 生 型 酵 素 お よ び 変 異 型 酵 素 の 精 製 野生型酵素および変異型酵素の精製は以下の 手順で行なった。野生型酵素遺伝子および変異型 酵素遺伝子を含む発現ベクターpETTP および pETTPEPCR を大腸菌 BL21(DE3)株で形質転換し た。得られた形質転換株を大量培養後に菌体を回 収し、超音波破砕を行い粗抽出液を得た。これを DEAE‐Cellulose 陰イオン交換クロマトグラフィ ー、Sephacryl S‐200HR ゲルろ過クロマトグラフ ィーによるカラム操作を行い、電気泳動的に単一 な精製標品を得た。 酵 素 活 性 の 測 定 と タ ン パ ク 質 定 量
PPase の酵素活性は、Peel と Loughman の方法 に従い 37℃にて行なった 16)。また、タンパク質 濃度の決定は、bovine serum albumin を標準タン パク質として、Bradford 法により行なった17)。 遠 紫 外 部 CD スペクトルの測定 CD スペクトルは、千葉工業大学総合科学研究 所にて測定を行った。測定試料をタンパク質濃度 を0.1mg/ml に調製し、室温にて J-720 dichrograph (JASCO)により測定を行った。1mm 石英セルを用 いて、200nm から 250nm の楕円率を測定し、分 子楕円率〔θ〕(deg・cm2/dmol)に変換した。また、 α-helix 含量は、次式から算出した18)。 α-helix 含 量 (%) = ‐ ( [ θ ]222nm +2340) 100/ 30300 蛍 光 ス ペ ク ト ル の 測 定 野生型酵素および変異型酵素の蛍光スペクト ルの測定は、F-2500 spectrofluorometer (Hitachi)に より、タンパク質濃度を0.05 mg/ml とし、20mM Tris-HCl buffer (pH 7.8)を溶媒として室温で行っ た。Trp 蛍光スペクトルは Tth PPase 内の Trp 残基 (Trp149 および Trp155)を内在性プローブとし、励 起波長295nm、蛍光測定波長を 320∼ 400 nm に 設定し測定した。熱安定性における蛍光スペクト ル測定は、タンパク質濃度0.05mg/ml に調製した 酵素溶液を 40∼90℃の各温度で 1 時間加熱後氷 冷し、上記の条件で蛍光スペクトルを測定した。 四 次 構 造 の 分 析 野生型酵素および変異型酵素の分子量の測定 は、50mM Tris-HCl buffer (pH 7.8) を移動相に用 いて、HPLC に接続した TSKgel G3000SW カラム (Tosoh, 7.5mm I.D. × 30cm) により室温で行なっ た。流速 0.3ml/min で試料 5μg を注入し、波長 280nm で検出した。溶媒および試料は全て 0.45 μm フィルター(Millipore)でろ過したものを使用 した。熱安定性における四次構造の分析は、タン パク質濃度0.1mg/ml に調製した酵素溶液を 40∼ 90℃の各温度で 1 時間加熱後氷冷し、さらに 0.45 μm フィルターろ過後の試料を、上記の条件で測 定した。得られた溶出曲線の面積は、chromatopac CR‐6A (Shimadzu) を用いて算出した。
3. 実験結果
変 異 型 酵 素 の ス ク リ ー ニ ン グ お よ び 変 異 箇 所 の 同 定 まず、Tth PPase 遺伝子の Thr138 から Gly174 を含む C 末端領域に相当する部分に対し、No.5 upper primer および No.9 lower primer を用いて、 error‐prone PCR を行い、ランダム変異を導入し た。このとき、dNTP の濃度において dATP と dTTP、 またはdCTP と dGTP が通常の反応組成より 1mM 高いものとMn2+濃度を0.1、0.25、0.3、0.4、0.45mM と変えたものを、組み合わせて PCR を行った。 その結果、1mM dCTP、dGTP と 0.45mM Mn2+を 含む反応液でのみ PCR 増幅断片が確認できなか った。そこで PCR 増幅断片が確認できた1mM dCTP、dGTP および 0.45mM Mn2+を含む反応液以 外の試料各々を混合し、制限酵素による切断と T4 リガーゼによる ligation により、変異遺伝子を 発現ベクターに組み込んだ。次にこのベクターを 用いて大腸菌 BL21(DE3)株を形質転換しコロニ ーを得た後、96 穴プレートで培養を行い溶菌後、 条件を変えた 1 次∼3 次スクリーニング(一次: 90℃、2 時間加熱、二次:90℃、3 時間加熱、三 次:95℃、30 分加熱)により 1948 個中 5 個の耐熱 性を保持した陽性コロニーを得た。これら5 個の コロニーの発現確認を行い、DNA 塩基配列決定 により変異箇所を同定した。その結果5 個のうち 2 個は野生型 Tth PPase であったが、残る 3 個は それぞれ、L162F(TTG→TTT)、K173E (AAG→ GAG)、K159E (AAG→GAG)/A170T (GCC→ACC) のアミノ酸残基置換が確認された。次に、Tth PPase 遺伝子全領域に対しても同様 にerror-prone PCR を行い、ランダム変異を導入し た。No.8 upper primer および No.9 lower primer を 用いて、Tth PPase 遺伝子に先と同様の条件で PCR を行い、得られた変異遺伝子を発現ベクターに組 み込み、大腸菌BL21(DE3)株を形質転換した。96 穴プレートで培養を行い溶菌後、条件を変えた1 次∼3 次スクリーニング(一次:80℃、1 時間加熱、 二次:90℃、1 時間加熱、三次:100℃、1 時間加 熱)により 2400 個中 1 個の耐熱性を保持した陽性 コロニーを得た。この変異型酵素遺伝子を DNA
塩 基 配 列 決 定 し た と こ ろ 、Q119H(CAG → CAT)/L162F(TTG→TTT)のアミノ酸残基置換が 確認された。以上の結果より、Tth PPase のラン ダム変異体として、4 種の変異型酵素(L162F、 K173E、K159E/A170T、Q119H/L162F)が得られた。 野 生 型 酵 素 お よ び 変 異 型 酵 素 の 発 現 と 精 製 上記で得られた 4 種の変異型酵素の発現ベク ターを大腸菌BL21(DE3)株に形質転換し、培養後 粗抽出液を調製した。これらを、15%ゲルを用い た SDS-PAGE により分析したところ、変異型酵 素の発現が確認された(Data not shown)。さらに、 大量培養を行い、超音波破砕により溶菌させて粗 抽出液の得た後、DEAE‐cellulose 陰イオン交換 クロマトグラフィー、Sephacryl S‐200HR ゲルろ 過クロマトグラフィーの二本のカラム操作によ り、電気泳動的に単一な4 種の変異型酵素を得た。 野生型酵素についても、同様の手法により精製標 品を得た。 野 生 型 酵 素 お よ び 変 異 型 酵 素 の Initial characterization まず、変異導入による酵素への影響を検討する ため、試料のタンパク質濃度を0.1 ㎎/ml(蛍光ス ペクトルのみ0.05mg/ml)に調整し、未加熱状態 (Native 状態)における野生型および変異型酵素の 酵素活性、二次構造(CD スペクトル)、三次構造(蛍 光スペクトル)、四次構造(ゲルろ過クロマトグラ フィー)について検討した(Table I)。 酵素活性では、野生型酵素活性を100%として 相対活性を表すと、L162F は 106%、K173E は 95%、 K159E/A170T は 93%であり、変異導入による酵 素活性の顕著な差異は見られなかった。次に、野 生型および変異型酵素の二次構造を検討するた めに、遠紫外部CD スペクトルを測定した。その 結果、野生型酵素と比較して、各変異型酵素の二 次構造およびα-helix 含量に顕著な差異は観察さ れなかった。さらに、変異導入に伴う三次構造へ の影響を検討するため、分子内Trp 残基を蛍光プ ローブとして、励起波長 295nm で蛍光スペクト ルを測定した。その結果、各変異型酵素のTrp 残 基周辺の立体構造変化は生じていないことが示 唆された。最後に変異導入による四次構造への影 響を検討するため、未加熱状態において、HPLC を用いた TSK‐gel G3000SW ゲルろ過クロマト グラフィーによる分析を行った。その結果、四次 構造はいずれもHexamer であり、変異導入による 四次構造への影響は見られなかった。以上より、 変異導入に伴う酵素活性や立体構造への顕著な 影響は無いものと考えられる。 野 生 型 酵 素 お よ び 変 異 型 酵 素 の 酵 素 活 性 の 熱 安 定 性 次に、酵素活性を指標として、野生型酵素およ び変異型酵素の酵素活性の熱安定性を検討した (Fig.2)。各試料(0.1mg/ml)を 40∼90℃で 1 時間加 熱し、10 分間氷冷後、酵素活性の測定を行った。 その結果、Q119H/L162F の熱安定性は野生型酵素 と同等であったが、他の変異型酵素は野生型酵素 に比べ熱安定性の低下が見られた。特に 80℃加 熱後では、野生型酵素の残存活性は61.4%であっ たが、L162F は 38.8%、K159E/A170T では 34.0%、 K173E では 18.8%と顕著な差異が観察された。 野 生 型 酵 素 お よ び 変 異 型 酵 素 の 三 次 構 造 の 熱 安 定 性 野生型酵素および変異型酵素の三次構造の熱安 定性の差異を検討するため、各試料を 0.05mg/ml に調製後、40-90℃で 1 時間加熱し、Trp 蛍光スペ クトルによる分析を行った(Fig.3)。その結果、 Q119H/L162F, L162F 変異体は野生型酵素と同様 の 熱 安 定 性 を 示 し た が 、K173E お よ び
a The specific activity of wild type Tth PPase was taken as 100%. b λmax is the maximum wavelength of
tryptophan fluorescence spectra.c Fmax is the fluorescence intensity (arbitrary unit) at the emission maximum of
tryptophan fluorescence spectra. d α - helix content was calculated by the following equation 18); α - helix content
(%) = ‐([θ]222nm + 2,340) × 100 / 30,300. eQuaternary structure was estimated from the elution profile on
Fig.2 Thermostability of enzyme activity for wild type Tth PPase and variants
The enzyme (0.1mg/ml) was incubated in 20mM Tris-HCl buffer (pH 7.8) at the indicated temperatures for 1 h. Then the enzyme activity was measured at 37℃ after rapid cooling, and the activity at 40℃ was taken as 100%. Symbols: , wild type; , Q119H/L162F, , L162F; , K173E; Δ, K159E/A170T.
Fig.3 Thermostability of Tryptophan fluorescence spectra for wild type Tth PPase and variants The enzyme (0.05mg/ml) was incubated in 20mM Tris-HCl buffer (pH 7.8) at the indicated temperatures for 1 h. Then, the emission spectrum of each sample was measured with excitation at 295 nm after rapid cooling. On heating above 85 ºC, each sample was centrifuged after heating, and then its supernatant was measured. The fluorescence intensity at maximum wavelength was plotted as relative value to that at 40 °C. Symbols: , wild type; , Q119H/L162F, , L162F; , K173E; Δ, K159E/A170T. K159E/A170T 変異体では 85℃以上において三次 構造の熱安定性の上昇が見られた。このことは、 両変異型酵素では、85℃加熱後も約 70%の可溶性 分子が存在しており、両変異型酵素では熱凝集が 抑制される可能性が示唆された。 野 生 型 酵 素 お よ び 変 異 型 酵 素 の 四 次 構 造 の 熱 安 定 性 各試料を40℃から 90℃で 1h 加熱し、HPLC を用いた TSK‐gel G3000SW ゲルろ過クロマト グラフィーによる分析を行った(Fig.4)。その結果、 L162F は野生型と比較して 80℃での可溶性成分 の減少および熱凝集生成の増加が見られた。また K173E、K159E/A170T においてのみ、80℃、85℃ 加熱後で、高分子化成分(oligomer、>200kDa)の生 成が観察された(Fig.4(D),(E))。そこで 85℃におけ る野生型、K173E、K159E/A170T の四次構造の熱 安定性を経時的に分析した(Fig.5)。各試料を 85℃ で10∼30 分間加熱し、HPLC を用いた TSK‐gel G3000SW ゲルろ過クロマトグラフィーによる分 析を行った。その結果、野生型では熱凝集成分を 除去して分析しているため顕著なピークは観察 されなかったが、K173E および K159E/A170T で は可溶性成分(oligomer, hexamer, trimer)が検出さ れ、加熱時間の経過に伴い、hexamer の減少、 oligomer の増加、trimer の生成が見られた。また、 K159E/A170T に比べ K173E では 10min 加熱後で
のoligomer の生成量が多く見られた。このことか ら、K159E/A170T および K173E では、85℃で熱 凝集の抑制に伴う可溶性高分子化成分の増加が 生じており、Tth PPase における熱凝集生成は、 分子が高分子化する過程を経て起きる可能性が 示唆された。
4. 考察
本研究では、Tth PPase の熱安定性や熱凝集生 成に関与する新規のアミノ酸残基に関する知見 を得るため、比較的研究例の少ないTth PPase の α-helixB を含む C 末端領域に着目し、Tth PPase 遺伝子のα-helixB を含めた C 末端領域および全 領域に進化分子工学的手法(error-prone PCR)を用 いたランダム突然変異型酵素を作製した。その結 果、四種の変異型酵素(Q119H/L162F、L162F、 K173E、K159E/A170T)が得られた(Fig.6)。 各変異型酵素は、Native 状態では酵素活性、二 次構造、三次構造、四次構造において、野生型酵 素 と の 顕 著 な 差 異 は 見 ら れ な か っ た こ と か ら (Table I)、変異導入は、酵素活性と立体構造に大 きな影響を与えないと考えられる。このことは、Fig.4 Oligomeric thermostability for wild type Tth PPase and variants
Panels: (A) Wild type, (B) Q119H/L162F, (C) L162F, (D) K173E, (E) K159E/A170T. The enzyme (0.1mg/ml) was incubated in 20mM Tris-HCl buffer (pH 7.8) at the indicated temperatures for 1 h. After rapid cooling, the gel filtration chromatography was performed as described in MATERIALS AND METHODS. Composition was estimated from each peak area of hexamer, trimer, monomer, and oligomer (component having much higher molecular weight than hexamer) in the HPLC elution profiles. 100% was taken as the sum of all peak area after heating at 40ºC. Symbols: ●, hexamer; □, trimer; Δ, monomer; ■, oligomer.
上記の変異箇所(Fig.6)が酵素の活性部位やサブ ユニット界面に含まれておらず、直接酵素活性や 四次構造には影響を与えないためであると考え られる。 また、酵素活性を指標とした熱安定性に関して は、三種の変異型酵素(L162F、K159E/A170T、 K173E) で 80℃以上において熱安定性の減少が 観察され(Fig.2)、三次構造(Trp 蛍光スペクトル) ではK173E および K159E/A170T で 85℃以上で熱 安定性の上昇が見られた(Fig.3)。一方、四次構造 の熱安定性においては、K173E、K159E/A170T に おいてのみ、80℃、85℃加熱後で、oligomer と trimer の生成が観察された(Fig.4)。さらにこの二 種の変異型酵素については、85℃における四次構 造の経時変化より、hexamer および oligomer の存 在が確認されたことから(Fig.5)、Lys159、Ala170、 Lys173 におけるアミノ酸残基の置換が熱凝集 (aggregation) 抑制に関与していることが示唆さ れた。また、Tth PPase における熱凝集生成は、 hexamer→oligomer→aggregation のように進行し ていると推定され、このことはα-helix B を発端 に分子の立体構造が崩れていき14)、Cys168 によ る酵素分子同士の会合とLys159 や Lys173 などの イオン性残基の関与により、高温での溶解度の減 少を導き、活性型hexamer の減少と高分子化を経 て、不活性型熱凝集成分が増加すると思われる。 また、各変異型酵素におけるアミノ酸残基の役 割については、L162F および Q119H/L162F の場 合、野生型酵素と同様に熱凝集生成が見られ高分 子化成分が見られないことから共通する変異部 位(L162F)は熱凝集への関与は低いと考えられる。 また、酵素活性の熱安定性への影響については、 Q119H/L162F における Q119H 変異が、近接する 活性部位残基(Asp97、Asp102、Lys104 など)や単 量 体 界 面 残 基(Arg114) 、 周 囲 の イ オ ン 結 合 Asp120-Arg158)への微細環境に影響を及ぼすと 推定されるが(Fig.6)、この点に関してはさらに検 討する必要がある。 一方、Lys173 は C 末端に位置し、他残基と相 互作用を形成していないため側鎖のフレキシビ
Fig. 5 The elution profiles of TSK-gel G3000SW gel filtration chromatography (HPLC) for wild type Tth PPase and variants after heating at 85 °C
The flow rate was 0.3 ml/min. Each sample (0.1mg/ml) was heated at 85°C for the indicated times, and filtered by Millex filter (pore size 0.45μm, Millipore). 50μl of each protein after was injected to column, followed by eluted with 50mM Tris-HCl (pH 7.8), and then analyzed as described in MATERIALS AND METHODS.
リティが高いと推定される。この残基の電荷の逆 転(K173E)が、C 末端のカルボキシル基や、周囲 のArg166 や Arg171 に作用して、間接的に分子内 部にある Glu や Asp などとのイオン性相互作用 ( G l u 1 4 - A r g 1 6 6 - G l u 1 6 3 , G l u 6 4 - A r g 1 7 1 , Asp120-Arg158) に影響を与え(Fig.6) 8), 9)、活性 部位を含む分子の立体構造を変化させたため、酵 素活性の熱安定性(Fig.2)が低下したと考えられ る 。 また、この K173E における 85℃における熱凝 集の抑制(Fig.5)は、変異導入によって酵素分子の 表面電荷の負電荷が増加し、可溶性成分(oligomer, hexamer, trimer)の割合が上昇したためだと思わ れる。この高分子化や熱凝集の抑制は、部位は異 なるが同じく分子表面に位置する Ala144,Lys145 の欠損変異体においても観察されている11)。本研 究の結果でも、分子表面の Lys173 は熱凝集生成 に関与しており、Tth PPase の分子表面の電荷の 相違が、高温での分子の溶解度や分子間会合に影 響を与えることが改めて示唆された。 最後に、K159E/A170T については、得られた四 種の変異型酵素の中でも耐熱性を最も保持して いた。このことは、Lys159 は活性部位と離れて おり、さらに A170T のアミノ酸置換により OH 基が主鎖のNH 基と水素結合を形成し分子を安定 化している可能性もあり、酵素活性への影響は K173E より少なかったと思われる。また、85℃に おける熱凝集生成の抑制は、K173E と同様に、変 異導入による酵素分子の表面電荷の負電荷が増 加したためだと考えられ、酵素分子表面の負電荷 の増加は、酵素分子の高温での溶解度の増加を導 き、熱凝集生成を抑制すると推定される。 以上のことから、Tth PPase 分子のα-helix B に 含まれるイオン性残基(Lys159 および Lys173)は、 Tth PPase の分子全体の電荷に大きく影響するこ と 、 ま た こ の 電 荷 変 化 は 分 子 の 変 性 状 態 転 移 (hexamer→oligomer→aggregation)に影響を与え、 特に熱凝集の過程に関与する事が明らかとなっ た。 今後、本研究のC 末端領域に関する知見や 他の耐熱化因子に関する知見を基に、耐熱化因子 を組み合わせることによりTth PPase の更なる人 工耐熱化が可能であると考えられる。
謝辞
本研究は、実吉奨学会(Saneyoshi Foundation) お よび徳島大学教育改善推進費(奨励研究)の支援に より実施された。CD スペクトルの測定ならびに 研究の実施に際し、ご協力頂きました故・三浦謹 一郎東京大学名誉教授(元千葉工業大学教授)、 東京大学大学院の武井俊朗博士に深く感謝いた します。参考文献
1. Lahti, R. (1983) Microbial inorganic pyrophosphatases. Microbiol. Rev. 47, 169-179 2. Harutyunyan, E. H., Oganessyan, V. Y.,
Oganessyan, N. N., Avaeva, S. M., Nazarova, T. I., Vorobyeva, N. N., Kurilova, S. A., Huber, R., and Mather, T. (1997) Crystal structure of holo inorganic pyrophosphatase from Escherichia coli at 1.9Å resolution. Mechanism of hydrolysis. Biochemistry, 36, 7754-7760
3. Satoh, T., Samejima, T., Watanabe, M., Nogi, S., Takahashi, Y., Kaji, H., Teplyakov, A., Obmolova, G., Kuranova, I., and Ishii, K. (1998) Molecular cloning, expression and site-directed mutagenesis of inorganic pyrophosphatase from Thermus thermophilus HB8. J. Biochem., 124, 79-88
4. Maruyama, S., Maeshima, M., Nishimura, M., Aoki, M., Ichiba, T., Sekiguchi, J., and Hachimori, A. (1996) Cloning and expression of the inorganic pyrophosphatase gene from thermophilic bacterium PS-3. Biochemistry and Molecular Biology International., 40, 679-688
Fig. 6 Three-dimensional structure of Tth PPase and substituted amino acids by error-prone PCR .
The three-dimensional structure of monomeric Tth PPase. The figure was drawn by a Molecular Graphics Program, Swiss-Pdb Viewer Ver.4.0 (Swiss Institute of Bioinformatics). The X-ray structural data for Tth PPase was obtained from the Protein Data Bank (PDB code
2PRD) [12].Amino acids in the α-helix B forming ionic
interactions were drawn by red (acidic amino acid) and blue (basic amino acid). Substituted amino acids (Gln119, Lys159, Leu162, Ala170, Lys173) by error-prone PCR were shown in green.
5. Satoh, T., Shinoda, H., Ishii, K., Koyama, M., Sakurai, N., Kaji, H., Hachimori, A., Irie, M., and Samejima, T. (1999) Primary structure, expression, and site-directed mutagenesis of inorganic pyrophosphatase from Bacillus stearothermophilus. J. Biochem. 125, 48-57 6. Leppänen, V. M, Nummelin, H., Hansen, T.,
Lahti, R., Schäfer, G., Goldman, A. (1999) Sulfolobus acidocaldarius inorganic pyrophosphatase: structure, thermostability, and effect of metal ion in an archael pyrophosphatase. Protein Sci. 8, 1218-1231
7. Liu, B., Bartlam, M., Gao, R., Zhou, W., Pang, H., Liu, Y., Feng, Y., and Rao, Z. (2004) Crystal structure of the hyperthermophilic inorganic pyrophosphatase from the archaeon Pyrococcus horikoshii. Biophys. J. 86, 420-427
8. Teplyakov, A., Obmolova, G., Wilson, K.S., Ishii, K., Kaji, H., Samejima, T., and Kuranova, I. (1994) Crystal structure of inorganic pyrophosphatase from Thermus thermophilus. Protein Sci. 3, 1098-1107
9. Salminen, T., Teplyakov, A., Kankare, J., Cooperman, B.S., Lahti, R., and Goldman, A. (1996) An unusual route to thermostability disclosed by the comparison of Thermus thermophilus and Escherichia coli inorganic pyrophosphatases. Protein Sci, 5, 1014-1025 10. Satoh, T., Takahashi, Y., Oshida, N., Shimizu, A.,
Shinoda, H., Watanabe, M., and Samejima, T. (1999) A chimeric inorganic pyrophosphatase derived from Escherichia coli and Thermus thermophilus has an increased thermostability. Biochemistry, 38, 1531-1536
11. Satoh, T., Oshida, N., Ono, M., Hattori, M., Ohta, T., Watanabe, M., Shinoda, H., Takahashi, Y., Lee, J.S., and Samejima, T. (1999) Deletion of Ala144-Lys145 in Thermus thermophilus inorganic pyrophosphatase suppresses thermal aggregation. J.Biochem. 125, 858-863
12. Kouzai, M., Kohaku, Y., Sugiyama, N., Yamaguchi, Y., and Satoh, T. (2008) Threonine 138 is crucial for the Quaternary Structure and the Thermostability of Thermus thermophilus Inorganic Pyrophos- phatase. Natural Science Research, 22, 65-73 13. Tanaka, H., Maeda, A., Sugiyama, N.,
Kyogoku, H., Kouzai, M., Takei, T., Miura, K., and Satoh, T. (2008) Thermostabilization by the Improvement of Intertrimeric Residues in Thermus thermophilus Inorganic Pyrophos- phatase. Natural Science Research, 22, 85-96 14. Kohaku, Y., Maeda, A., Kouzai, M., Sugiyama,
N., Fukuhara, K., Kanai, T., Takei, T., Miura, K. and Satoh, T. (2008) Effect of Cys168 substitutions on the Thermostability and the Thermal Aggregation of Thermus thermophilus Inorganic Pyrophosphatase. Natural Science Research, 22, 75-84
15. Song, J. K and Rhee, J. S. (2000) Simultaneous enhancement of thermostability and catalytic activity of phospholipase A(1) by evolutionary molecular engineering. Appl Environ Microbiol.
66, 3, 890-894
16. Peel, J.L. and Loughman, B.C. (1957) Some observations on the role of copper ions in the reduction of phosphomolybdate by ascorbic acid and their application in the determination of inorganic orthophosphate. Biochem.J., 65, 709-716
17. Bradford, M. M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-254
18. Chen, Y. H., Yang, J. T., and Matinez, H. M. (1972) Determination of the secondary structures of proteins by circular dichroism and optical rotatory dispersion. Biochemistry, 11, 4120-4131
論文受付 2013 年 9 月 3 日 改訂受付 2013 年 9 月 30 日 論文受理 2013 年 9 月 30 日