博士学位論文
国内産ドジョウに内包される隠蔽種に関する
遺伝学的・生態学的研究
平成 30 年 1 月 9 日
近畿大学大学院 農学研究科 環境管理学専攻 岡 田 龍 也1
目 次
第1 章 序論・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・4 第2 章 福井県中池見湿地におけるドジョウ 2 系統集団間の遺伝学的・生態学的研究 1 節 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・12 2 節 ドジョウ 2 系統集団間における生殖的隔離の実態・・・・・・・・・・・・・・・・・13 2-2-1 緒言 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・13 2-2-2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・14 2-2-3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・18 2-2-4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・20 3 節 ドジョウ 2 系統集団間における生態的分化の検証・・・・・・・・・・・・・・・・・22 2-3-1 緒言 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・22 2-3-2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・23 2-3-3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・24 2-3-4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・25 第3 章 隠蔽種の両性生殖集団と交雑由来の 3 倍体・クローン集団の系統関係 1 節 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・28 2 節 新規集団における系統判別・・・・・・・・・・・・・・・・・・・・・・・・・・・31 3-2-1 緒言 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・312 3-2-2 材料と方法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・32 3-2-3 結果および考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・33 3 節 新規集団における遺伝子型の判別 ・・・・・・・・・・・・・・・・・・・・・・・34 3-3-1 緒言 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・34 3-3-2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・34 3-3-3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・36 3-3-4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・37 4 節 新規集団における交雑の確認 ・・・・・・・・・・・・・・・・・・・・・・・・・38 3-4-1 緒言 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・38 3-4-2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・39 3-4-3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・40 3-4-4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・40 5 節 mtDNA 分析によるドジョウ科の系統解析 ・・・・・・・・・・・・・・・・・・・・42 3-5-1 緒言 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・42 3-5-2 材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・42 3-5-3 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・43 3-5-4 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・46 第4 章 総合考察 1 節 共存集団と融合集団・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・53 2 節 共存集団系統の起源・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・54
3 3 節 融合集団系統の起源・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・55 4 節 各グループの集団形成・・・・・・・・・・・・・・・・・・・・・・・・・・・・・56 5 節 Type I 系統の分布および集団形成・・・・・・・・・・・・・・・・・・・・・・・・62 6 節 ドジョウの分類学的提言・・・・・・・・・・・・・・・・・・・・・・・・・・・・69 引用文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・74 謝辞 図表
4 1章 序論 隠蔽種とは,形態的に区別がつかず同一種として扱われてきたが,実際には 生殖的に隔離された別種のことを指す (Mayr, 1970).従来,形態的差異に基づい た分類学によって多くの種が記載されてきた一方で,形態的に判別のつかない 隠蔽種が見落とされてきた.しかし,近年の分子系統学的研究の発展によって 隠蔽種の存在が明らかにされるようになってきた.例えば,Asai et al. (2011) に よる分類学的整理においては,日本人なら誰もが知っている小魚である「メダ カ」が,ミナミメダカOryzias latipes とキタノメダカ Oryzias sakaizumii の 2 種に
別れるという点で社会的にも大きな話題となった.我々日本人にとって身近な 淡水魚であるメダカでさえ隠蔽種を含んでいたという事実は,多くの生き物の 種分化やその実態がまだまだ明らかになっていないことを物語っている.実際, その他の淡水魚においても近年になって新たな隠蔽種が次々と報告されている ( 例. ヨ ド ゼ ゼ ラ Biwia yodoensis, Kawase and Hosoya, 2010; カ マ ツ カ
Pseudogobio esocinus の一種, Tominaga et al., 2016; タンゴスジシマドジョウ
Cobitis takenoi, Nakajima, 2016).「種とは互いに交配しうる自然集団で,それは他
のそのような集団から生殖の面で隔離される」という生物学的種概念[Mayr, 1942(1999); マイヤ,1994]に基づくと,隠蔽種の存在を検証するには生殖的隔 離の確認が非常に重要である.しかし,生殖的隔離に関する研究は多くの魚類
5
で不充分であり,前述のメダカ 2 種においても生殖的隔離について充分な検証
は成されていない.そして,メダカと並んで日本人にとって身近な淡水魚であ る「ドジョウ」においても隠蔽種の存在が示唆されてきたが,生殖的隔離の実 態について明らかになっていない.
ドジョウMisgurnus anguillicaudatus (Cantor, 1842) はコイ目ドジョウ科ドジョ
ウ属に分類される体軸方向に伸長した体形を持つ種であり (Fig. 1),日本全土, 台湾,中国大陸および朝鮮半島など極東域に分布しているが,北海道と琉球列 島では自然分布か明らかになっていない(斉藤,1989;藤田,2015).最近の分 類学の見解では,ヨーロッパから東アジアにかけて同属7 種(Misgurnus nikolskyi,
Misgurnus fossilis, Misgurnus dabryanus, Misgurnus mohoity, Misgurnus buphoensis, Misgurnus multimaculatus, Misgurnus tonkinensis)が分布すると報告されており
(Kottelat, 2012; 細谷,2013),これらのうち,日本国内には在来種のドジョウと 外来種であるM. dabryanus(カラドジョウ)の 2 種の生息が確認されている.ド ジョウは水田や用水路など身近な環境に生息し,「どぜう鍋」や「柳川鍋」など の食用として,あるいは釣りや動物園で飼育される鳥類の餌としての利用や, 島根県に代表される「どじょうすくい」に登場するなど古くから民謡・童謡に 歌われ,神事・祭事にもドジョウがまつわるなど,文化的にも人の暮らしに非 常に密接した淡水魚である(中島・内山,2017).ドジョウと聞けば誰しもが, 土色で,細長い口髭があり,にょろにょろとした姿を思い浮かべることができ
6 るほどに,記号化された存在となっている. ドジョウの体にある模様には様々なパターンがあり地理的変異や個体変異が 非常に大きく,形態的に分化した集団が日本国内で見つかっており(久保田・ 小野,1964; 菫ほか,1999),複数の種を含んでいることが示唆されてきた.さ らに,ドジョウに関する分子系統学的研究も盛んに行われており,遺伝的に分 化したグループも日本国内で報告されている(菫ほか,1999; Khan and Arai, 2000, Morishima et al., 2008; Šlechtová et al., 2008; 小出水ほか,2009; 清水ほか,2011; Kitagawa et al., 2011; 鹿野ほか,2012).これらの研究から,日本国内の在来集団 には大きく 2 つに分化したミトコンドリア(mt)DNA 系統である Type I 系統と Type II 系統を持つドジョウが確認されている (Kitagawa et al., 2011).これら 2 系
統は他の先行研究ではそれぞれ A クレード,B クレードとも呼ばれている
(Morishima et al., 2008; 小出水ほか,2009).これら 2 系統の分化程度の指標とな る mtDNA 遺伝子領域の塩基置換率はおよそ 13-18 %であり (Morishima et al., 2008; 小出水ほか,2009),ヨーロッパに生息するシマドジョウ属の亜属間を上 回る程度の塩基置換率である(11.8-13.6 %; Perdices and Doadrio, 2001).Type I 系統は日本国内の北日本を中心に不連続に検出され,大陸に生息する他のドジ ョウ属と近縁な関係性を示す.Type II 系統は日本国内全域で検出され,ドジョ ウ科シマドジョウ属 Cobitis と近縁な関係性を示す.これら 2 系統は核 DNA が
7
子浸透が関係していると先行研究で述べられている (Šlechtová et al., 2008; Kitagawa et al., 2011).すなわち,およそ 700-1000 万年前に生息していたドジョ ウの祖先種の一部の集団がシマドジョウ属の祖先種と交雑を起こし,ドジョウ の祖先種のmtDNA にシマドジョウ属の祖先種の mtDNA が浸透することで Type II 系統が誕生した.一方,Type I 系統はドジョウの祖先種の中でシマドジョウ属 の祖先種と交雑することなく進化してきた系統である.つまり,日本人が目に する機会の多いType II 系統のドジョウとは遺伝子浸透を受けたものであり,日 本国内で極稀に確認される Type I 系統こそが真のドジョウ系統と言える.中国 大陸ではType I 系統の存在が確認されておらず,後から誕生した Type II 系統に よって駆逐されてしまった可能性から,日本国内の縁辺部に不連続に分布する Type I 系統は本来のドジョウ系統の遺存(レリック)集団とされている(Kitagawa et al., 2011).以上のように,これら 2 つの mtDNA 系統は異なる進化的背景を持 つことや大きく遺伝的に分化していることから,同一の個体群として取り扱う ことは不適切と考えられているが(例えば,清水,2014),生物学的な種を定義 するうえで重要である生殖的隔離の成立については研究されていない.一方で, この mtDNA の遺伝的差異のみを根拠として現行のドジョウから,ドジョウ
Misgurnus anguillicaudatus,キタドジョウ Misgurnus sp. (Clade A),ヒョウモンド
ジョウMisgurnus sp. OK,シノビドジョウ Misgurnus sp. IR の 4 種に分けること
8
観に基づいており,分類学的な混乱が生じているため早急な研究が求められて いる.
ドジョウの中には北海道や石川県を中心とした地域で「クローン」個体が稀 に生息していることが報告されている (Morishima et al., 2002; Morishima et al., 2008; 荒井,2009).一般的な多くの生物種は両性生殖を行うが,クローンドジ ョウは雌性発生によって生じていることが確認されている (Morishima et al., 2002).また,同じように北海道や石川県を中心とした地域で倍数性変異を起こ した3 倍体個体が確認されており (Zhang and Arai, 1999; Arai, 2003; Morishima et al., 2008),ドジョウの雌性発生クローンが稀に産する 2n の非還元卵が偶発的に 精子を受精することで自然 3 倍体個体が生じることが報告されている(小野里 ほか,1993;森島ほか,1999;Morishima et al., 2002;糸納・荒井,2004).さら には,2n の卵が雌性発生によって無性生殖的に繁殖することで 2 倍体クローン が生じることも確認されている (Morishima et al., 2002).いずれの場合も,ドジ ョウの特殊な生殖様式を持つ集団は全てクローンや 3 倍体という現象が関係し ているとされているが,広域なドジョウの分布域の中ではこれらは限られた地 域でしか生じていない.ドジョウ集団内の多くは両性生殖を行う 2 倍体が確認 されており,少なくとも 3 倍体・クローン個体を含む集団とは異なる集団であ ると思われるが,その関係性など多くのことが明らかになっていない. 近年,河川や用水路の改修,護岸工事,圃場整備などによる生息地の減少あ
9 るいは消失,農薬や家庭排水,侵略的外来種の導入などの影響による生息条件 の悪化,これらの要因でドジョウの個体数は激減しており,環境省のレッドリ ストでは情報不足(環境省,2015),各都道府県のレッドデータブックでも 13 の府県で絶滅危惧種I 類から情報不足などに位置付けられている(野生生物調査 協会・Envision 環境保全事務所,2012).また,上述した外来種のカラドジョウ が現在多くの地域で確認されており,生息場所や餌料面での競合が懸念されて いる(加納ほか,2007;加納・今井,2008;松沢・瀬能,2008).さらに,ドジ ョウの場合は同種による外来種問題も生じている.中茎ほか(2007)は栃木県 内と東京中央市場におけるドジョウ流通実態を調査し,その取扱量のほとんど が中国産であること,産地表示が国内となっているものは納入業者の所在地で あり,すべて中国からの輸入であることを報告した.また,清水・高木(2010) は,愛媛県内で中国産として販売されているドジョウと同じミトコンドリア・ ハプロタイプを自然水域から確認しており,国内へ輸入されているドジョウが なんらかの過程を経て,すでに一部の在来個体群に侵入していることが明らか となっている.非在来性ドジョウの自然水域への侵入過程としては,食材や釣 り餌の放逐,養殖地からの逸散のみならず,保護対象となっている鳥類のトキ
Nipponia nippon やコウノトリ Ciconia boyciana,ナベヅル Grus monacha の餌と
しての他産地からの導入(総務省,2013; 吉郷,2007; 景平ほか,2009;Kano et al., 2011),仏教儀式の放生会における市場等で販売されているドジョウの放流
10 ( 例 え ば , 梅 原 總 山 墓 相 研 究 所 ホ ー ム ペ ー ジ ,http://www5b.biglobe. ne.jp/~bosou/intoku/intoku2.htm, 2013-8-19 参照)などがあり,非在来性の遺伝子 系統の拡散要素は多方面にわたっている.中国産のドジョウは日本産のドジョ ウと遺伝的には容易に識別可能であり,早期の発見によって更なる国外遺伝子 の侵入を抑えられる可能性があるが,養殖として利用するドジョウを国内の他 地域から持ち込み,逸出してしまった他地域産ドジョウが自然域に拡大した場 合も国内外来種として在来集団に影響を及ぼすと考えられるが,より遺伝的に 近い集団間では非在来性の判断が困難になる場合が多い.全国のドジョウの遺 伝解析を実施した研究では,国内のいくつかの地域集団で遺伝的に地域固有性 がみられないなどすでに影響が広がりつつあり(小出水ほか,2009),今後の在 来性を判断する際に困難をきたすと予想される. 昨今では,保全・外来種問題はどの生物にも当てはまる問題となってしまっ ているが,ドジョウの場合は何を守るべきか,その実態さえ曖昧となってしま っている.即ち,上述したようにドジョウの中には大きく分化した異なる遺伝 的系統,雌性発生クローンや倍数性変異を起こした 3 倍体および非在来性系統 が含まれており,一括りにできない複雑な遺伝的・分類学的背景があり,種の 実態が定まらないまま今日にいたるまで様々な用途で利用され続け,人為的撹 乱が進行しているのである.誰もが知り,利用できる身近な生物でありながら, その種の正体が明らかになっていないというのはドジョウならではと言える.
11 本研究では遺伝学的手法を用い,分類学上 1 種として一括りにされてきた日本 国名のドジョウ集団に内包される種の実態を明らかにするものである.さらに, 異なる遺伝的集団間の生態学的研究,分子系統学的研究も行い,それぞれの関 係性についても調査・研究を行ってきた. 本論文は全 4 章からなる.第 2 章では福井県中池見湿地に焦点を当て,ドジ ョウにみられる 2 つの mtDNA 系統の生殖的隔離と生態的分化について調べた. 続く第3 章では日本国内全域から収集したドジョウの DNA 情報を基に系統関係 を調べ,それぞれの集団の系統差および集団の成り立ちについて検証した.最 後に第 4 章の総合考察として,第 2 章,第 3 章で明らかになった結果を基に現 時点で考えらえるドジョウの集団形成を探るとともに分類学的な提言を行い, 今後の研究課題について述べた.
12 2章 福井県中池見湿地におけるドジョウ 2 型集団間の遺伝学的・生態学的研究 1 節 緒言 生殖的隔離とは,ある生物群の間で有性生殖による遺伝子の交流が存在しな いこと,あるいは交流後に個体群が維持されないことである.つまり,個体の 間で交配ができない状況がある場合,それらは速やかに生殖的に隔離されてい ることを意味する.生殖的隔離には交配前隔離と交配後隔離があり,交配前隔 離には生息場所や繁殖期の違いなど自然界での出会いや繁殖機会がないことが 挙げられ,交配後隔離には不妊や雑種の生存力の低下などがある.また,地理 的に隔離されている 2 つの集団間では物理的に交配は起きないため,異所的に 分布する生物群の生殖的隔離を証明することは困難である.一方,対象とする2 つの集団が同所的に生息している場合,両者の生殖的隔離の有無について直接 的に検証することができる.また,生物群の中で生態的に分化が生じている場 合,異なる環境への適応によって隔離障壁が進化し,生殖的隔離が発達する種 分化の様式と成り得る(松林・藤山,2016).非常に密接な近縁種が同所的に生 息する場合,近縁種同士が共存するメカニズムが強固になる傾向があり (Taper and Case, 1985),同所的に生息する近縁種同士の繁殖生態が異なることがいくつ かの種において報告されている(例.アカガエル属 Rana, Hillis, 1981; ショウジ ョウバエ属 Drosophila, Seiger and Khamis, 1987).
13
本研究では,同所的に生息する異なる2 つの mtDNA 系統(Type I 系統,Type II 系統)をそれぞれ有するドジョウに着目し,両者が生殖的に隔離された別種と して異なる繁殖集団を維持しているのか,共存するためのメカニズムの一部と
して生態的分化が生じているのか,この2 点についてそれぞれ検証した.
2 節 ドジョウの 2 つの mtDNA 系統集団間における生殖的隔離の実態 2-2-1 緒言
日本国内に生息するドジョウに含まれるmtDNA の Type I 系統と Type II 系統
は大きく遺伝的に分化しているが,異なる 2 系統は種内多型として存在してい
るのか,異なる繁殖集団の存在を示しているのかわかっていない.ドジョウType I 系統と Type II 系統が同所的に出現する地域がいくつか知られおり (Morishima et al., 2008; 小出水ほか,2009; 清水・高木,2010; Fujimoto et al., 2017) (Fig. 2), 日本国内全域を対象にしたドジョウの遺伝学的調査によって,福井県中池見湿 地においても2 つの mtDNA 系統が同所的に検出された.そこで,福井県中池見
湿地においてドジョウ 2 系統の生殖的隔離について調査を行った.中池見湿地
に同所的に生息するドジョウ2 系統には核 DNA においても分化がみられるのか, 遺伝的交流はあるのか,そして遺伝的分化は表現型として形態的差異に表れて
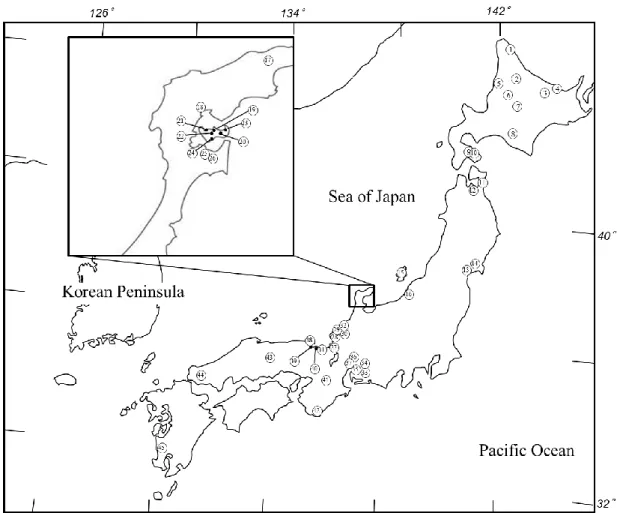
14 いるのか,遺伝学的および形態学的解析を行った. 2-2-2 材料と方法 採集とmtDNA 系統判別 中池見湿地とは福井県敦賀市に位置する総面積 250,000 m2の内陸性低湿地で ある (Fig. 3; 坂巻,2003).地下水を水源とし,湿地内の水は“後谷”と称される 1 本の水路に集められ笙の川水系に連なる木の芽川に流れ出る. 2013 年 5 月から 2015 年 5 月にかけて,タモ網(目合い約 3.5 mm,底辺長約 300 mm)を用いて中池見湿地内に生息するドジョウ 54 個体を採集した.採集さ れた個体は 70 %エタノールで固定し,一部の個体は近畿大学農学部の所蔵標本 として登録・保存した(KUN-P 045276-045285).固定された鰭組織からフェノ ール・クロロホルム法 (Asahida et al., 1996) によって全 DNA を抽出した.即ち, 1.5ml チューブに鰭組織と TNES-6Ubuffer 緩衝液を 250µl,Protenase K (TE 溶 解 20mg/ml)を 5µl 加え,37℃のインキュベーターに一晩静置して鰭組織を溶解 した.溶解したサンプルにフェノール・クロロホルム溶液を 250µl を加え 10 分 間撹拌し,遠心分離機(10,000rpm,10 分,15℃)によってスピンダウンした. その後,DNA が含まれている上澄み液を新しいチューブに移してジエチルエー
15 テルを 250µl 加え,3 分間撹拌した後に軽くスピンダウンし,不要な上層・中間 層を取り除いた.さらに,5M NaCl を 25µl と無水エタノールを 500µl 加えて 5 回反転させ,-80℃で 1 時間以上冷却保存した.そして,遠心分離機(15,000rpm, 15 分,4℃)でスピンダウンした後に上澄み液を捨て,70 %エタノール溶液 500µl を加え 5 回反転した.その後,再度遠心分離機(15,000rpm,15 分,4℃)によ ってスピンダウンした後に上澄み液を捨て,1 時間以上の静置乾燥によりペレッ ト状の DNA を得た.乾燥後,TE を 150µl 加え冷蔵保存した. 各個体の mtDNA 系統を判別するため,全 54 個体で mtDNA の制限酵素断片長 多型(RFLP)分析を行った.Šlechtová et al. (2008) に従い,プライマーセット Glu–L.Ca14337–14359(5′-GAA GAA CCA CCG TTG TTA TTC AA-3′)と Thr–
H.Ca15598–15548(5′-ACC TCC RAT CTY CGG ATT ACA-3′)を用いて mtDNA シトクロムb 遺伝子(cyt b)の PCR 増幅を行った.反応条件は,95℃・120 秒の熱 変性の後,95℃・30 秒,55℃・30 秒,72℃・60 秒を 30 サイクル行い,72℃・ 300 秒の最終伸長とした.1 %アガロースゲル(Agarose LO3,タカラバイオ株式 会社,滋賀県)を用いて電気泳動をした後,ミドリグリーン染色(日本ジェネ ティクス株式会社,東京都)および紫外線照射によって増幅の有無を確認した. 続いて,Kitagawa et al. (2011) に報告されている対象領域の配列情報を基に,タ カ ラ バ イ オ 株 式 会 社 の ホ ー ム ペ ー ジ に あ る 制 限 酵 素 認 識 配 列 検 索 ツ ー ル (http://www.takara-bio.co.jp/enzyme/enzyme_search.php)を用いて 2 系統の判別に
16
有効な制限酵素を探索し,2 種類の制限酵素 Dde I,Rsa I(タカラバイオ株式会 社,滋賀)を用いて制限酵素反応を行った.この時の反応は各試薬に添付され ているプロトコールに従った.反応後,3 %アガロースゲル(Agarose LO3,タ カラバイオ株式会社,滋賀県)上で電気泳動し,ミドリグリーン染色(日本ジ ェネティクス株式会社,東京都)および紫外線照射によってバンドパターンを 検出することで2 系統の判別を行った. 核DNA の単一遺伝子分析 単一の核遺伝子に対する PCR-RFLP 分析について全 54 個体で行った.DNA データベース[日本DNA データバンク(DDBJ),欧州分子生物学研究所(EMBL), および米国国立生物工学情報センター(GeneBank)]に登録されている M. anguillicaudatus の発現配列タグ(EST)を用いた予備実験の結果に基づき,リボ ヌクレアーゼサブユニット H2 遺伝子(rnaseh2)領域(アクセッション番号: BJ818489)を 2 系統の多型分析に選択した.PCR 増幅には新たに設計したプラ イマーセットBJ818489F(5′-GTT AGT TGC CGC TTG TCC TC-3′)と BJ818489R (5′–CCC CAC AGT CCA AAG CAT AC–3′)を用いた.反応条件は,95℃・120 秒の熱変性の後,95℃・30 秒,62℃・30 秒,72℃・60 秒を 30 サイクル行い, 72℃・600 秒の最終伸長とした.続いて,PCR 産物を特異的な制限酵素 Alu I(タ カラバイオ株式会社,滋賀)で消化した.この時の反応は各試薬に添付されて
17 いるプロトコールに従った.反応後,3 %アガロースゲル(Agarose LO3,タカ ラバイオ株式会社,滋賀県)上で電気泳動し,ミドリグリーン染色(日本ジェ ネティクス株式会社,東京都)および紫外線照射によってバンドパターンを検 出することでヘテロ接合体の有無および2 系統の多型について調べた. AFLP 法によるゲノムフィンガープリンティング
Type I 系統と Type II 系統間でゲノム DNA を共有しているかを確認するため, 中池見湿地内で同じ調査時期の同地点から採集されたType I 系統 8 個体と Type II 系統 7 個体において,増幅断片長多型(AFLP)法によるゲノムフィンガープ
リンティングを行った.AFLP 法は詳細な遺伝的変異を評価する分析手法であり,
ゲノム全体を網羅的に分析できること,高い再現性を有していることからクロ ーン構造を調べるのに適していることが示されている (Bensche and Åkesson, 2005; Barreto et al., 2011).AFLP 分析には AFLP plant mapping kit(Applied Biosystems 社,アメリカ)を用い,操作手順は添付のプロトコールに従った.予 備実験としてプライマー選定を行い,明瞭なDNA 断片の波形が確認できた Eco RI-AGC,AAG,ACA と Mse I -CTG,CTA,CTT の 3 通りの組み合わせを用い た.増幅産物はGeneScan 500 Rox size standard(Applied Biosystems 社,アメリカ) とともにApplied Biosystems 3130 Genetic Analyzer で分析した.検出された DNA 断片はPeak Scanner Software 2.0(Applied Biosystems 社,アメリカ)を用いて収
18
集した.50-540bp の範囲の DNA 断片を 1 または 0 としてコードし,それぞれの
個体の各DNA 断片の存在または不在を示した.コード化されたデータに対して
Arelequin 3.5 (Excoffier and Lischer, 2010) を用いて分子分散分析(AMOVA)を行 った.2 系統間の遺伝的分化は Wright(1965)によって定義されたペアワイズ固 定指数(Fst)によって評価した. 形態学的解析 全54 個体において胸鰭の形状観察(塚原,1948)および解剖による生殖腺の 観察を行い,各個体の性別を決定した.雄の胸鰭基部にある骨質盤の形状と総 脊椎骨数の2 つはドジョウ科において重要な分類形質である(相澤,1981; 澤 田・相澤,1983).2 系統それぞれの雄個体が持つ胸鰭基部の骨質盤について光 学顕微鏡を用いて,総脊椎遺骨数については 2 系統の各個体において軟 X 線撮 影によってそれぞれ観察した.また,得られた結果は R 統計ソフトウェアパッ
ケージ(R Development Core Team, Austria, Inc.)が実装する Wealch の t 検定によ
って統計的に評価した.
19
中池見湿地内で採集されたドジョウ54 個体(雄 31 個体,雌 23 個体)が確認 できた箇所は全9 地点であった (Fig. 3).mtDNA の PCR-RFLP 分析によって確
認されたDde I 処理による DNA 断片は,Type I 系統でおよそ 380,350,250 お
よび120bp,Type II 系統でおよそ 700,330 および 70bp,Rsa I では,Type I 系統 でおよそ1100bp,Type II 系統でおよそ 560,470 および 120bp であった.この結 果,Type I 系統が 16 個体,Type II 系統が 38 個体確認された (Table 1).Type I 系統では雄10 個体,雌 6 個体,Type II 系統では雄 21 個体,雌 17 個体されてお り,2 系統ともに雌雄両方が生息していることがわかった.また,2 系統が同所 的に確認されたのは4 地点であった.
核DNA の PCR-RFLP 分析の結果,確認されたバンドパターンは Type I 系統お よびType II 系統に特異的であり,rnaseh2 遺伝子においても遺伝的な分化がみら
れた.Alu I 処理による DNA 断片は,Type I 系統でおよそ 400,320,130,70 お
よび30bp,Type II 系統でおよそ 240,210,160,150,100,80,50,50,50 お よび40bp であった.2 系統でヘテロ接合体は検出されなかった.
AFLP 法によって得られた DNA 断片の総数は Type I 系統で 53-59,Type II 系 統は87-99 であった (Table 1).AGC + CTG(EcoRI + Mse-I),AAG + CTA(Eco-RI + Mse-I)および ACA + CTT(Eco-RI + Mse-I)の各プライマーセットで得られた DNA 断片は,Type I 系統ではそれぞれ 20-24,11-14 および 20-23,Type II 系統 ではそれぞれ28-36,18-23,37-51 であった.DNA 断片の共有率を示す BSI(Band
20
Sharing Index)の範囲は,Type I 系統内で 0.87-0.96,Type II 系統内で 0.85-0.94 であり,クローン個体は確認されなかった.また,2 系統間の平均 BSI はおよそ 0.30 であり,ゲノム共有率は低いことが示された.AMOVA で示された遺伝的変 異は2 系統間で 87.73 %,系統内で 12.27 %であり,2 系統間の方が系統内より遺 伝的に大きな変異が蓄積されていた.さらに,Type I 系統と Type II 系統間にお けるFstは0.877 であり,2 系統間で有意に分化していた(permutation test; P<0.01). 雄個体にみられる胸鰭基部の骨質盤の形状は,2 系統間で大きさに若干の差異 がみられたが定量化するには至らなかった.一方で,総脊椎骨数は Type I 系統 で48-51 本,Type II 系統で 44-48 本であり,2 系統間で有意な差がみられた(Wealch のt 検定; P<0.01)(Table 1). 2-2-4 考察 これまで,日本国内に分布するドジョウM. anguillicaudatus にはいくつかの遺 伝的に分化したグループが報告されてきたにもかかわらず,生殖的隔離につい ての研究例はなく,分類学的研究に基づく整理は不充分であった.本研究で示 された核の単一遺伝子やAFLP 法による分析では,同所的に生息するドジョウ 2 系統はゲノムDNA を共有していないことが明らかになった.また,2 系統とも
21 に雌雄が確認され,雌雄間のBSI の範囲において各系統で差異はなかった(Type I 系統,雄集団内 0.87-0.94,雌集団内 0.88-0.96,雌雄間 0.90-0.96;Type II 系統, 雄集団内 0.89-0.92,雌集団内 0.89-0.94,雌雄間 0.85-0.92).これらから,雌雄 両方が共通の遺伝子プールを持っていることを 2 系統ともに示していることが わかった.さらに,雌雄両方および 2 系統ともにほとんどの個体において成熟 した生殖腺が確認され,2 系統ともに両性生殖を行っていることが明らかになっ た.結論として,Type I 系統と Type II 系統は生殖的に隔離されていることがわ かった.これを裏付けるように両者の間には形態的な分化も認められた.した がって,2 系統は 2 つの異なる生物種として認識されるべきであると考えられた. 以降,本稿ではmtDNA 系統を示す場合には Type I 系統および Type II 系統,種 を示す場合にはType I 種および Type II 種とする. ドジョウに充てられた学名M. anguillicaudatus は中国の舟山島の標本に対応し ている (Cantor, 1842).日本国内以外で Type I 系統は確認されておらず,分布域 から中国大陸に生息するドジョウはType II 系統(遺伝子移入系統)であると考 えられており,学名であるM. anguillicaudatus はドジョウ Type II 種に適用される べきである.そのため,少なくとも中池見湿地には 2 系統の違いに由来する別 種が生息していることは明らかで,中池見湿地の Type I 系統(レリック)は新 たに確認された隠蔽種のドジョウType I 種(Misgurnus sp.)として記載されるべ きであり,今後の分類学的研究が求められる.一方で,mtDNA で Type I 系統を
22 指すもの全てが別種のType I 種というわけではなく,Type I 系統内には倍数性変 異を示す個体やクローン個体が存在しているため,中島・内山(2017)が示す ように単純な分類はできないと思われる.倍数性変異個体やクローン個体につ いては第3 章,分類学的提言に関しては第 4 章で詳しく述べる. 3 節 ドジョウ 2 系統集団間における生態的分化の検証 2-3-1 緒言 本章内の 2 節で示したように,中池見湿地内に生息するドジョウ 2 系統は生 殖的隔離が生じた互いに共存している別種(Type I 種,Type II 種)である.その ため,2 種間の生態学的特徴の差異が共存を維持している可能性が示唆される. 特に,繁殖は全ての生物種にとって重要なイベントであるため,繁殖期は両者 の生態的特徴の差異が明確な時期である可能性が高い.繁殖期における空間 的・時間的な生息環境の差異を例とした生態学的特徴は,2 種の隔離に決定的な 影響を及ぼしているかもしれない. 本研究では,ドジョウの繁殖期と非繁殖期において中池見湿地内でドジョウ の定量的採集を行い,2 種の空間的・時間的な生息環境の差異について調査した.
23 2-3-2 材料と方法 中池見湿地内におけるドジョウの採集調査は2016 年の 4 月から 8 月の間にお よそ月に 1 回のペースで行った.ドジョウの一般的な繁殖期は 6 月から 7 月で あるため(斉藤,1989),本調査期間は繁殖期および非繁殖期を含んでいる.ド ジョウに関する産卵行動として,普段は水路などの恒久的な水域に生息し,繁 殖期になると水田などの一時的水域に移動して産卵することが報告されている (斉藤ほか,1988;田中,1999).そのため予備的な採集調査を行い,比較的ド ジョウの採集が容易で異なる環境を採集地点として選んだ[水田 2 地点(St. 1 およびSt. 2),灌漑水路(St. 3)](Fig. 4).これらの採集地点は互いに直接的には 繋がってはいないが,増水などの季節的な変化やそれに伴う繁殖期における水 田への侵入など,ドジョウ 2 種の出現頻度の変化が反映されると予想された. 中池見湿地内は水位変動が激しいことから採集地点の範囲を一定にすることは 困難であったため,努力量を一定にする[3 人×20 分/地点,D フレームネット (目合い約0.5 mm,底辺長訳 300 mm)の使用]ことで定量採集調査とした.採 集された個体は,標準体長,雌雄を記録し,尾鰭を数 mm カットした後に,個 体を採集地点に放流した.採取した鰭組織はすぐに70 %エタノールで固定した. 田中(1999)に従い,体長 50 mm 以下は稚魚,それ以上は未成魚・成魚として
24
記録した.成魚における雌雄の判別は塚原(1948)に従い,雄の胸鰭の形状に 及ぼす二次的性的特徴の外部観察によって行った.未成魚の雌雄は基本的に不 明とした.各サンプルの種判別(Type I 種または Type II 種)は,mtDNA の cyt b におけるPCR-RFLP 分析によって行った.手法については 2-1 で述べた通りであ る.各地点における2 種の生息割合を比較し,R ver. 3.2.3 (R Development Core Team, 2013) を用いて Bonferroni 法による補正および x2検定の多重比較によって 統計解析を行った.
2-3-3 結果
全調査期間を通じて採集された成体の個体数は 82 個体であり,各地点で採集 された個体数は異なっていた(St. 1 で 51 個体,St. 2 で 18 個体,St. 3 で 13 個体) (Fig. 5) (Table 2).採集された成体の中で Type I 種が確認されたのは 11 個体 (13.4 %)のみであり,Type II 種に対して中池見湿地内における Type I 種の生 息が非常に少ないことが確認された (Table 2).灌漑水路(St. 3)から採集され た Type I 種の個体数は他の 2 地点(St. 1 および St. 2)より有意に多かった(St. 1 と St. 2 vs. St. 3, P < 0.001).また,St. 1 では Type I 種は全く確認されなかった. 一方で,St. 1 および St. 2 で採集された Type II 種の個体数は St. 3 より有意に多
25 かった(St. 1 と St. 2 vs. St. 3, P < 0.001).各地点で採集された 2 種の割合に調 査回毎で有意な差はみられなかった (Fig. 5). 調査期間中に採集された稚魚は全 48 個体であり大部分は水田からであった (St. 1 で 24 個体,St. 2 で 23 個体,St. 3 で 1 個体).採集された中で最も標準 体長の小さい個体は 24 mm であり,調査期間中に当歳魚と考えられた個体は確 認されなかった.St. 2(4 個体)および St. 3(1 個体)から採集された Type I 種 の稚魚は 5 個体のみであった. 2-3-4 考察 灌漑水路(St. 3)におけるドジョウ Type I 種の高い生息割合は 2 種間の生息環 境の相違を示唆した.さらに,この結果の妥当性を検証するため 2016 年 10 月 23 日および 2017 年 7 月 17 日に St. 3 で追加調査を実施したところ,どちらの結 果からも今回の結論を支持するものが得られた(Type I 種 : Type II 種 = 9: 1, 計 10 個体)(Fig. 5) (Table 2). 水田と灌漑水路では水温の上昇率やプランクトンの種類および多様性に違い があることが報告されており (William and Coad, 1979; 田中,1999),ドジョウ 2 種の好適生息環境の差異がそれぞれの生息場所を異所的にした可能性が考えら
26 れた.また,2 種は総脊椎骨数など形態的に異なっており,それが生息環境への 適応に影響しているかもしれない.一方で,単純に今回選択された調査地点の 位置関係による影響によって 2 種が採集された割合が地点毎で異なっていた可 能性も考えらえた.本研究は水田と灌漑水路という最も異なる環境下において2 種の生息割合の検証を行ったものであり,今後は地点数を増加してより詳細な 環境差異で検証を行う必要がある.しかし,中池見湿地の面積は大きく,全て の環境下で採集調査を実施するのは現実的ではない.さらに,採集地点は泥深 い環境であり,ドジョウを採集するには採集者の能力に影響され客観性を維持 するのが困難になってくる.そのため,これらの課題を解決する手法として環 境(e)DNA 分析の導入が考えられた.eDNA 分析は水中に含まれる DNA を直 接分析することで種の在・不在やバイオマス量を推定する手法であり(山中ほ か,2016),調査員の力量に影響されず客観的なデータが得られ,なおかつ短時 間で多くの地点を分析することが可能となる.既に本研究の継続研究によって eDNA 分析は実施されており,断片的ではあるが成果も挙げられている.今後は eDNA 分析によって多くの地点でドジョウ 2 種の割合を抽出し,各地点の物理環 境や餌資源を調査する必要がある. 近年のドジョウの個体数の減少が考慮され,環境省が発行する最新のレッド リストにドジョウは絶滅危惧種として掲載されている(環境省,2015).本章 2 節で述べたように,日本国内に生息するドジョウには遺伝的・形態的に異なる2
27 種が含まれていることが明になったため,これら 2 種は別々の保全単位として 扱われるべきである.これまでの先行研究では,Type II 系統の分布域でしか生 態学的研究は行われおらず(斉藤ほか,1988;田中,1999),レリックかつその 分布域の狭さから希少生物と考えられるドジョウ Type I 種に関して,生態学的 情報を収集することは急務と言える.
28 第3 章 隠蔽種の両性生殖集団と交雑由来の 3 倍体・クローン集団の系統関係 1 節 緒言 第2 章で述べてきたように,ドジョウ Type I 系統と Type II 系統は中池見湿地 内で同所的に確認されており,これらは互いに生殖的に隔離された別種(Type I 種,Type II 種)であることが明らかになった.自然界において, 種間では基本的 に生殖的隔離が起きているため,雑種が生じることは稀であるが (Dobzhansky, 1951),古くからいくつかの魚類において種間交雑に関する報告があることも事 実である (Hubbs, 1955).国内の淡水魚においても最近の遺伝学的研究によって ようやくその実態が明らかになってきたが(例.チチブTridentiger obscurus とヌ
マチチブT. brevispinis, Mukai et al., 2000; ニホンイトヨ Gasterosteus nipponicus と
太平洋系イトヨG. aculeatus, Yamada and Goto, 2003),その報告例は僅かである.
同所的に生息する近縁種間の自然交雑は一部分で生じるに留まり,基本的には それぞれが別種として独自性を維持している.一方で,交雑によって新たな集 団を形成している事例も報告されており,メダカ種群(ミナミメダカとキタノ メダカ)では過去に両種が交雑した結果生じた“Boundary population”という新た な地域集団群を生み出したとされている(酒泉, 1987; Takehana et al., 2016).ま た,北米産のコイ科魚類Phoxnus eos-neogaeus は P. eos と P. neogaeus との雑種起
29
ることで3 倍体を生み出すことがあり (Dawley and Goddard, 1988; Goddard et al., 1989),これらは一般的な有性生殖を行う集団や 2 倍体集団のような通常の種と は異なると言える.
ドジョウの場合,中池見湿地以外でも2 つの mtDNA 系統である Type I 系統と Type II 系統が同所的に出現する地域が確認されており,これらの多くの地域で はクローンドジョウや3 倍体ドジョウが確認されている (Morishima et al., 2008; 小出水ほか,2009; 清水・高木,2010, Yamada et al., 2015; Fujimoto et al., 2017) (Fig. 6).また,クローンドジョウや 3 倍体ドジョウは特に北海道の集団から多く確認 されているが (Morishima et al., 2008),北海道に生息しているドジョウは古くか ら在来性が疑われてきた(稗田,1984; 斉藤,1989; ).
最近の研究で,3 倍体・クローンドジョウは Type I 系統と Type II 系統の核 DNA をヘテロ接合の状態で有していることが確認され,集団の形成には Type I 系統 とType II 系統の交雑が起因となっていることが明らかになっている (Yamada et al., 2015; Fujimoto et al., 2017).荒井,(2009),Yamada et al. (2015) の報告をまと めると,異なるゲノムをそれぞれもつType I 系統と Type II 系統が交雑した場合,
異質なゲノムであることから減数分裂時に異常をきたし,通常の n 卵ではなく
非還元の 2n 卵を産することがある.この 2n 卵は受精時に精子核を排除して雌 性発生によってクローン 2 倍体ドジョウを生み出す.また,クローン 2 倍体ド ジョウが産する2n 卵が偶発的に精子核を取り込むことによって 3 倍体ドジョウ
30 が稀に生じる.さらに,クローン由来の自然 3 倍体ドジョウでは 1 セットの異 質ゲノムが排除され,残りの相同性の高い 2 ゲノムで減数分裂がおこり n 卵を 形成する「減数分裂雑種発生」機構が発生する.すなわち,「Type I 系統と Type II 系統の交雑」→「クローンドジョウの形成」→「3 倍体ドジョウの発生」とい う段階を踏んで集団中に新たな繁殖様式を行う個体が誕生し,これらが混在す る場合がある. 上記の異なる繁殖様式を持つ個体を含む集団がある一方,中池見湿地内で生 息するType I 種と Type II 種は交雑することなくそれぞれ別種として独自性を維 持している.これは,中池見湿地内で互いに両性生殖を行う集団として共存し ていることから,本研究ではType I 種と Type II 種の「共存集団」と呼称する. 一方で,3 倍体・クローンドジョウは Type I 系統と Type II 系統の交雑によって 誕生したものであり,Type I 系統,Type II 系統,クローン,3 倍体,それぞれが 互いに交雑を繰り返すことで集団を維持している.これらには上記で述べたあ らゆる段階のものを含むが,本研究ではType I 系統と Type II 系統の「融合集団」 と呼称する. これまで共存集団の存在は明らかになっていなかったため,Type I 系統と Type II 系統が同所的に出現する地域では両者間の交雑によって融合集団が形成され るというのが一般的な考えであった(例えば,荒井,2009).しかし,共存集団 の存在が明らかになった本研究では,同じType I 系統と Type II 系統が同所的に
31 出現する場合,共存集団と融合集団という異なる集団形成が生じたことに関し て疑問があり,それぞれの集団形成には異なる現象が生じている可能性が考え られた.即ち,共存集団と融合集団では異なる起源によって別々の集団形成が 起きてきたと推察され,融合集団においても複数の起源によって様々な集団形 成が生じてきた可能性が考えられた. ミトコンドリアは細胞質を通して母系遺伝するため,核DNA のような組み換 えが起こらないことから,集団の起源を探るにはmtDNA による分子系統解析が 有効な手段である(小池・松井,2003).また,細かな系統の差を探索すること で共存集団および融合集団の起源となった集団を検証できる可能性も考えられ た.本研究では,共存集団と融合集団を用いてmtDNA の分子系統解析を行うこ とで,それぞれの系統関係および起源について推定することを目的に調査を実 施した.始めに,本研究によって日本国内各地から採集・収集されたドジョウ のmtDNA 系統の判別をし(2 節),これらの研究で新たに得られた集団と既存研 究との遺伝子型の比較し(3 節),共存集団と融合集団を構成する遺伝子型の関 係性について検討した(4 節). 2 節 新規集団における系統判別 3-2-1 緒言
32
融合集団であるクローンや3 倍体個体は,mtDNA において Type I 系統を示す ことが先行研究で明らかになっている(Morishima et al., 2008; Yamada et al., 2015; Fujimoto et al., 2017).そのため,特に融合集団の起源を探るには mtDNA の Type I 系統を含む集団を特定することが重要となる.また,Type I 系統を含む集団の 出現は稀であり,その分布に地理的なまとまりを示さないことから,広域な網 羅的な採集が必要である.そこで,Type I 系統の検出が多く確認されている中 部・東日本を中心に新たにドジョウの採集・収集を行い,mtDNA の cyt b 遺伝子 領域におけるPCR-RFLP 分析による mtDNA 系統の判別を行った. 3-2-2 材料と方法 中部・東日本を中心に採集・収集した計45 地点 335 個体のドジョウを採集し た (Table 3) (Fig. 7).採集した個体は 70 %エタノールで固定し供試材料とした. 供試材料は,筋組織または鰭組織から2-2-2 と同様にフェノール・クロロホルム
法(Asahida et al., 1996)によって全 DNA を抽出し,DNA サンプルとしてその 後の実験に使用した.系統判別のためのPCR-RFLP 分析の手順は 2-2-2 と同様に 行った.
33 3-2-3 結果および観察 全45 地点 335 個体のうち,Type I 系統が 83 個体,Type II 系統が 252 個体確認 された.Typ I 系統は北海道のクッチャロ湖,名寄市,網走,女満別,古丹別, 北竜町,蓴菜沼,大沼,石川県の輪島市,向田町,新保町の計11 地点の 83 個体 から検出された (Table 4).本研究によって新たに Type I 系統が確認されたのは 北海道のクッチャロ湖,古丹別,北竜町,蓴菜沼,大沼,石川県の輪島市,向 田町,新保町の計8 地点であった.また,Type I 系統と Type II 系統の同所的な 出現が確認されたのは,北海道の北竜町,蓴菜沼,大沼,石川県の輪島市,向 田町,新保町の計6 地点であった. 本研究によって Type I 系統は北海道や石川県で多く確認され,既存研究と同 様の傾向であった.また,確認された地点数,個体数ともにType I 系統は Type II 系統より少なく,希少性が高いことが改めて確認された.Type I 系統と Type II 系統が同所的に生息していることが確認された地域では,交雑が生じている可 能性が考えられたため,これらの集団を対象とした解析を次に行った.
34 3 節 新規集団における遺伝子型の判別 3-3-1 緒言 融合集団に含まれるクローンや3 倍体個体は Type I 系統の mtDNA を保持して いること,特に,mtDNA 調節領域(D-loop)においては特定の遺伝子型(ハプ ロタイプIII)を有することが確認されており(Morishima et al., 2008),この遺伝 子型に代表される Type I 系統のドジョウ集団が融合集団の形成に母系起源とし て深く関わっていることを示している.しかし,ドジョウと同じクローンや倍 数性変異がみられるギンブナCarassius sp.では,クローンの起源には複数の系統 が関与していることが示唆されており(大原,2010),ドジョウの融合集団にお いてもハプロタイプIII 以外の系統が起源となっている可能性は十分考えられる. 本節では,本研究で新たに採集された地域の集団について,ドジョウにおいて すでに多くの系統の情報が蓄積されている D-loop の塩基配列決定を行い,既知 のハプロタイプと照合した. 3-3-2 材料と方法 材料には,新規集団からType I 系統を 27 個体,Type I 系統と同所的に生息し
35
ている集団を中心にType II 系統 8 個体,中池見湿地(共存)集団から 12 個体, 計47 個体の DNA サンプルを用いた (Table 3) (Table 5).清水・高木(2010)に 従い,プライマーセットL15923(5′-TTA AAG CAT CGG TCT TGT AA-3′)と H16498(5′-CCT GAA GTA GGA ACC AGA TG-3′)用い D-loop の PCR 増幅を行 った.反応条件は,95℃・120 秒の熱変性の後,95℃・30 秒,62℃・30 秒,72℃・ 60 秒を 30 サイクル行い,72℃・600 秒の最終伸長とした.全ての PCR 産物は 1 % アガロースゲル(Agarose LO3,タカラバイオ株式会社,滋賀県)を用いて電気 泳動を行い,ミドリグリーン染色(日本ジェネティクス株式会社,東京都)お
よび紫外線照射によって増幅の有無を確認した.増幅が確認されたPCR 産物は,
NucleoSpin® Gel and PCR Clean-up(MACHEREY-NAGEL 社,ドイツ)を用いて 精製し,シークエンス反応の鋳型とした.シークエンス反応はApplied Biosystems 3730xl DNA Analyzer によって解析受託している株式会社マクロジェン(京都府) に依頼した.この時の反応プライマーにはPCR 時と同じものを用いた.各 DNA サンプルから得られた塩基配列に加え,DDBJ に登録されている本領域を含んだ ドジョウの塩基配列のうち,Morishima et al. (2008)で検出された Clade A(本研 究における Type I 系統)に属するハプロタイプ(I,II,III,IV,VIII,X,XI, XII)と,Clade B-1,Clade B-2(本研究における Type II 系統)に属するハプロタ イプ(V,VII),Saitoh et al. (2003),Saitoh et al. (2006),清水・高木(2010)で検 出された近縁種 Misgurnus nikolskyi, Misgurnus dabryanus, Misgurnus mohoity,
36
Cobitis striata striata のデータを取得後,ClustalW(Thompson et al., 1994)を用い
て多重アライメントを行い,近隣結合法(NJ)による系統解析を行った.NJ に おける遺伝的距離の計算は Kimura(1980)の 2 パラメータモデルを選択した. 各枝の結びつきについては1,000 回のランダム・サンプリングによるブートスト ラップ確率により信頼性を得た.これらの計算と系統樹の作成には系統解析ソ フトMEGA7 ver. 7.0.14(Kumar et al., 2016)を用いた.
3-3-3 結果 本研究で用いた新規の集団のうち,北海道のクッチャロ湖集団,網走集団か ら検出された個体がハプロタイプ I のグループに,北海道のクッチャロ湖集団, 網走集団から検出された個体がハプロタイプ II のグループに,北海道のクッチ ャロ湖集団,北竜町集団,蓴菜沼集団,大沼集団から検出された個体がハプロ タイプIII にそれぞれ含まれた (Table 4, 5).石川県の輪島市集団,向田町集団か らそれぞれ確認された個体はハプロタイプ III に遺伝的に近い新たな遺伝子型 (ハプロタイプ A)が検出された.また,中池見湿地の Type I 種は報告されて いるどのハプロタイプとも遺伝的に離れた単一の遺伝子型(ハプロタイプD)を 有していた.以上これらは全てType I 系統で確認されたハプロタイプであった.
37 Type II 系統からは,既存研究のハプロタイプ V,VII,XI と新たな遺伝子型(ハ プロタイプB,C)が検出された.なお,本研究で用いた集団からはハプロタイ プIV,VIII,X,XII を持つ個体は検出されなかった. 3-3-4 考察 本研究でハプロタイプIII に含まれた北海道のクッチャロ湖集団,北竜町集団, 蓴菜沼集団,大沼集団の個体は融合集団と同一の遺伝的系統であることが示唆 れた.一方で,石川県の輪島市集団,向田町集団からそれぞれ検出された遺伝 子型はハプロタイプ III と遺伝的に近いが明確に異なっている新たな遺伝子型 (ハプロタイプA)が得られた.新規集団の中でハプロタイプ III およびハプロ タイプ A は,クローン判別および倍数性確認の予備実験によって 3 倍体および クローン個体が検出されており(高橋・小野里,未発表),これらは新規に確認 された融合集団であることが示唆された.ハプロタイプIII とハプロタイプ A は 遺伝的に近いことから,融合集団と密接に関係している可能性が考えられるが, 融合集団の成立には複数の系統の集団が関わっていることが示唆された.また, 共存集団である中池見湿地からは,既存研究にはない新たな単一の遺伝子型で 構成されていたことが示され,融合集団の起源となったものとは異なる系統に
38 属していることが示唆された.これらにより,Type I 系統内に代表される各集団 の遺伝的な違いが共存集団と融合集団の違いを引き起こしている可能性が示唆 された. 4 節 新規集団における交雑の確認 3-4-1 緒言 3 倍体やクローン個体のドジョウは,Type I 系統と Type II 系統の交雑を起因に して誕生し,さらに特定の遺伝子型(ハプロタイプ III)を持つことが先行研究
によって確認されており(Yamada et al., 2015; Fujimoto et al., 2017),本研究では 交雑を介して3 倍体,クローン,Type I 系統,Type II 系統が含まれているものを 融合集団と定義した.即ち,融合集団を形成する個体にはType I 系統と Type II 系統の両方の核DNA を有していることが考えられた. 3 節で融合集団が示唆されたハプロタイプ III と同一の集団(北海道のクッチ ャロ湖集団,北竜町集団,蓴菜沼集団,大沼集団)の個体,ハプロタイプIII と 遺伝的に近い新たな遺伝子型(ハプロタイプA)の集団(石川県の輪島市集団, 向田町集団)の個体において,交雑によって異なる遺伝子型を構成しているか を検証するため核DNA 分析を行った.
39
3-4-2 材料と方法
材料には,融合集団であることが示唆された集団からハプロタイプIII を有す る9 個体とハプロタイプ A を有する 4 個体,共存集団(中池見集団)から 86 個 体,その他のハプロタイプ(I, II, V, VII, XI, B, C, D)または D-loop を対象にした
遺伝解析が行われていないためハプロタイプが不明な計21 個体を用いた.分析
手法には核DNA の 2 遺伝子(rnaseh2,RAG-1)を対象に PCR-RFLP 分析を行った (Table 3, 7).rnaseh2 遺伝子領域に対する PCR-RFLP 分析のプロトコールは第 2 章で述べた通りである.RAG-1 に関しては,第 2 章における rnaseh2 の PCR-RFLP 分析でType I 種,Type II 種と判別された中池見産のサンプルを用い,マーカー としての有効性を予備実験で検証した.DNA サンプルから Šlechtová et al. (2008) に従い,プライマーセットRAG-1F(5′-AGC TGT AGT CAG TAY CAC AAR ATG-3′) とRAG-RV1(5′-GTG TAG AGC CAG TGR TGY TT-3′)を用いて PCR 増幅を行っ た.反応条件は,95℃・120 秒の熱変性の後,95℃・30 秒,54℃・30 秒,72℃・ 60 秒を 30 サイクル行い,72℃・300 秒の最終伸長とした.得られた塩基配列は タカラバイオ株式会社のホームページにある制限酵素認識配列検索ツール (http://www.takara-bio.co.jp/enzyme/enzyme_search.php)を用いて 2 系統の判別に
40 有効な制限酵素を探索し,選ばれた制限酵素 Pst I(タカラバイオ株式会社,滋 賀)を用いて PCR-RFLP 分析を行った.この時のプロトコールは制限酵素に付 随するマニュアルに従った. 3-4-3 結果 ヘテロ接合体はType I 系統内でみられるハプロタイプ III およびそれに近い新 たな遺伝子型ハプロタイプA を持つ個体でみられた (Table 4, 7) (Fig. 8).以上の ことは核DNA の 2 遺伝子とも同様であった.また,Type I / Type I のホモ接合体 は46 個体,Type II / Type II のホモ接合体は 62 個体,Type I / Type II のヘテロ接 合体は12 個体確認された.
3-4-4 考察
ハプロタイプIII の遺伝子型を持つ個体から TypeI / Type II のヘテロ接合体であ ることが確認されたため,新規集団のうち北海道の蓴菜沼集団,大沼集団,石 川県の向田町集団は融合集団であると結論付けられた.また,共存集団である
41 中池見湿地内の個体において,Type I 系統,Type II 系統ともにヘテロ接合体は確 認されず,2 章と同様に交雑によって異なる遺伝子型を形成しない共存集団であ ることが改めて示された. その他,新たな Type I 系統の遺伝子型(ハプロタイプ A)を持つ石川県の輪 島市集団,向田町集団の個体においてもTypeI / Type II のヘテロ接合体が確認さ れたため,融合集団であると考えられた.また,ハプロタイプIII 以外の遺伝子 型を持つ個体で交雑を示すヘテロ接合体が確認されたのは本研究が初めてであ り,ハプロタイプ A を持つ個体は重要なサンプルになり得ると思われた.さら に,ハプロタイプIII とハプロタイプ A を持つ交雑個体では,遺伝子型が若干異 なっていることから,融合集団の形成に系統的差異が生じていると考えられた. 即ち,ハプロタイプIII とハプロタイプ A を持つ交雑個体の系統を詳しく調べる ことで融合集団の起源を探ることに繋がると思われた. 2 節から 4 節の検証によって先行研究と本研究で用いられたサンプルの情報が 整理できた (Table 3, 7).5 節では融合集団と共存集団を用いてそれぞれの系統関 係および起源について推定することを目的にmtDNA の分子系統解析を行い,特 に,共存集団,融合集団が含まれているType I 系統を中心に論じていく.
42 5 節 mtDNA 分析によるドジョウ科の系統解析 3-5-1 緒言 1 節および 2 節において,融合集団を中心に既存研究との照合,特に,ハプロ タイプIII の遺伝子型と交雑による異なる遺伝子構成の有無について検証してき たが,本章の目的である共存集団と融合集団の起源を探るには,他集団との細 かな系統差を見出すことで集団の形成が推察できる可能性がある. ドジョウの起源はユーラシア大陸にあるため(澤田,1987; Šlechtová et al., 2008),集団の形成を推察するには大陸に分布する近縁集との関係性も重要とな る.それぞれの起源を考察することは目的とし,日本国内だけでなく大陸産の ドジョウ科に関するデータも多く蓄積されている領域であるmtDNA の cyt b を 対象としてドジョウ科の系統類縁関係を明らかにし,その中での融合集団と共 存集団の遺伝子型における位置づけについて検討した. 3-5-2 材料と方法 2 節から 4 節で情報が整理された共存集団および融合集団も含む計 45 地点 335
43
おいて,cyt b の PCR 増幅を行った.この時のプロトコールと増幅に用いたプラ
イマーは2-2-2 の通りである.増幅の確認後シークエンス反応を行い,その時の
手順は 3-2-2 と同様であった.各 DNA サンプルから得られた塩基配列は,
ClustalW (Thompson et al., 1994) を用いて多重アライメントを行った.得られた
塩基配列にDDBJ に登録されているドジョウとその近縁種の配列データを加え,
最尤法(ML)と近隣結合法(NJ)による系統解析を行った.この時引用した塩
基配列データの種名は DDBJ の登録名および最近の論文で使用されている種名
に従った.Find Best DNA/Protein Models (ML)を用いて ML の最適なモデルを計 算し,その結果HKY+G+I model (Hasegawa, Kishino and Yano, 1985) が選ばれた. NJ における遺伝的距離の計算は Kimura (1980) の 2 パラメータモデルを選択し た.各枝の結びつきについては1,000 回のランダム・サンプリングによるブート ストラップ確率により信頼性を得た.これらの計算と系統樹の作成には系統解 析ソフトMEGA7 ver. 7.0.14 (Kumar et al., 2016) を用いた.系統樹の外群には Kitagawa et al. (2011) に従って,Sabanejewia balcanica[AY887851; Šlechtová et al. (2008)]および Pangio pangia[EF508583; Šlechtová et al. (2008)]を選択した.
44
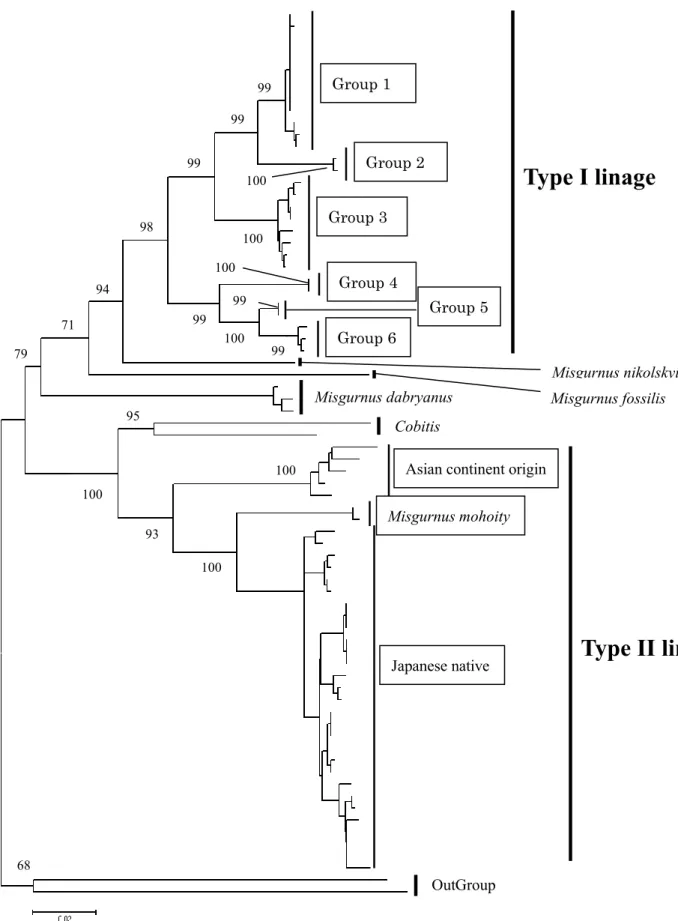
得られたcyt b の塩基配列をアライメントした結果,部分塩基配列 1014bp が決 定された.系統樹では,ドジョウは2 つの大きな系統である Type I 系統と Type II 系統に分けられ,それぞれ高いブートストラップ率で支持された(98.0 %および 93.0 %) (Fig. 9).また,ML・NJ ともに同様の系統関係を示す樹系図が得られ た.Type I 系統は大陸に分布するドジョウ属の各種(M. nikolskyi, M. fossilis, M.
dabryanus)に,Type II 系統はシマドジョウ属と近縁な関係を示し,先行研究と
同様の結果が得られた.Type I 系統内ではさらに単系統性が支持された 6 つのグ ループ(Group 1-6)に細分化され,グループ間の塩基置換率は 1.9-9.0 %であっ た.Type II 系統内には,日本産ドジョウとともに,大陸産ドジョウ,M. mohoity が含まれ,Type II 系統内の塩基置換率は 6.8-11.9 %であった.
cyt b により得られた樹系図の構成は D-loop のものと同様であり,Group 1a は ハプロタイプIII,Group 1b はハプロタイプ A,Group 2 はハプロタイプ D,Group 3 はハプロタイプ I,II,Group 6 はハプロタイプ XI に対応した (Table 4).なお, DDBJ から引用したデータである Group 4,5 は D-loop との対応の確認ができな かったが,ハプロタイプX,XII のいずれかに相当するものと考えられた.よっ て,融合集団から検出されたmtDNA 系統は全て Type I 系統の Group 1 に含まれ た.下記に,ドジョウにみられる共存集団と融合集団の差異において重要な Type I 系統内の各 Group の結果についてまとめる.
45
Group 1 (Fig. 10)
グループ内でさらに2 つのサブグループ(Group 1a, Group 1b)がみられた.Group 1a は北海道のクッチャロ湖,名寄市,女満別,北竜町,蓴菜沼,大沼,青森県 の五所川原市,岩手県の奥州市,石川県の半浦,向田町,さらに近縁種の M. nikolskyi サハリン集団から得られた個体で構成されていた.ほぼ全ての個体から は同じ塩基配列が得られ,1 個体で 1 塩基の置換が確認されただけで遺伝的均一 性の極めて高いグループであった.Group 1b は石川県の輪島市,向田町,福井 県の越前市の集団で構成され,Group 1a との塩基置換率は 0.43 %であった. Group 2 (Fig. 11) 共存集団である中池見集団から得られた全ての個体と,中池見湿地と水域が繋 がっている笙の川水系の上流部に位置する池河内湿原の集団の個体という,中 池見周辺の集団のみで構成されていた.Group 2 で確認されたハプロタイプは 2 種類であった. Group 3 (Fig. 12) クッチャロ湖,名寄市,女満別,網走,今金町,共和町などの北海道の広い地 域で検出された集団,さらに近縁種のM. nikolskyi サハリン集団から得られた個 体で構成されていた.各地域で形成されるサブグループはみられず,固有性も
46 確認されなかった. Group 4 (Fig. 13) 宮城県今泉集団とM. nikolskyi サハリン集団から得られた個体で構成され,塩基 配列は同一であった. Group 5 (Fig. 14) 福島県須賀川市集団とM. nikolskyi サハリン集団から得られた個体で構成され, 塩基配列は同一であった. Group 6 (Fig. 15) 栃木県さくら市集団と石川県新保町集団から得られた個体で構成され,それぞ れ3 つと 5 つのハプロタイプが確認された. 3-5-4 考察
Kitagawa et al. (2011) では,シマドジョウ類から mtDNA の遺伝子移入を受け たドジョウ属をType II 系統としており,本研究ではこれに大陸産ドジョウ,M.